


The human organic anion transporter 4, hOAT4 (SLC22A11, solute carrier family 22 member 11) is expressed at the apical membrane of cells of renal proximal tubule and due to its strategic localization, is thought to play a key role in the renal disposition of negatively charged compounds (Hagos et al., 2007; Liu et al., 2019; Louisse et al., 2023); however, whether hOAT4 works by reabsorbing and/or eliminating organic anions (OAs) remains uncertain at this point in time. hOAT4 is unambiguously involved in reabsorbing uric acid (Xu et al., 2017; Zhang et al., 2021) or environmental toxins like poly- and perfluoroalkyl compounds (Louisse et al., 2023) but is remarkably less effective in reabsorbing other OA like para-aminohippuric acid; as a matter of fact, para-aminohippuric acid is a much better substrate for efficient elimination from blood in the glomerular filtrate, showing that hOAT4 may equally well assist the apical exit of OAs (Hagos et al., 2007). It was in keeping with the latter possibility that dianionic angiotensin II receptor blockers (ARBs) such as azilsartan, candesartan, carboxylosartan, valsartan, and olmesartan were identified as potential substrates for OAT4-mediated renal excretion, provided that extracellular Cl– was available to function as an exchangeable anion; however, for other ARBs, such as the monoionic losartan, there was less dependence on extracellular Cl– (Noguchi et al., 2021). These findings may help to translate concepts and dissect the overall process of OAs excretion in two distinct steps: 1) basolateral entry, usually but not universally mediated by hOAT1-3 in exchange for an intracellular anion (Otani et al., 2017), and 2) apical exit, which may or may not involve hOAT4. The lack of a murine ortholog of hOAT4 clearly limits experimental approaches for understanding if and how hOAT4 may work to promote kidney excretion of OAs.
The current issue of the Journal of Pharmacology and Experimental Therapeutics offers an interesting contribution that scrutinizes similarities and differences between hOAT4 and murine Oat5 and may thus shed light on this complex scenario (Martinez-Guerrero et al., 2024). mOat5 (Slc22a19, solute carrier family 22 member 19) is genetically distinct from hOAT4 (less than 50% sequence analogy) but shares the same apical localization of hOAT4 in cells of kidney proximal tubule (Burckhardt and Burckhardt, 2011); moreover, mOat5 has long been known to work with some substrates that are also involved in hOAT4 driven processes (e.g., estrone sulfate or ochratoxin A) (Koepsell, 2013; Nigam et al., 2015). Building on these premises, the authors generated Chinese hamster ovary (CHO) cells that stably expressed hOAT4 or mOat5 and could be exploited to decipher how similar or different the two transporters were in promoting OAs excretion.
As an encouraging preliminary finding, the authors found that hOAT4 and mOat5 exhibited similar Ktapp (apparent Michaelis constant of transported substrate) and Jmax (maximal rate of mediated substrate transport) for radiolabeled ochratoxin (OTA), used as prototypic anionic substrate; moreover, but similar to the aforesaid differences in hOAT4-mediated transport of distinct ARBs, preloading CHO cells with glutarate or succinate as exchangeable anions resulted in transstimulation of the uptake of OTA but not of 6-carboxyfluorescein. Next, the authors evaluated how effectively OTA transport was inhibited by drugs like nonsteroidal anti-inflammatory agents (indomethacin, diclofenac) or diuretics (probenecid, hydrochlorothiazide). For each compound, the IC50 values were reasonably similar for hOAT4 and mOat5 but noticeable differences were observed across different competitors, with indomethacin and diclofenac showing significantly lower IC50 values compared with probenecid and hydrochlorothiazide. On balance, the IC50 values were in the range of those reported by others in separate and methodologically different experiments (Burckhardt, 2012). Having characterized functional similarities between hOAT4 and mOat5 in terms of uptake of a prototypical OA like OTA, the authors moved on to characterize if the two OATs were also similar in promoting substrate efflux, which was in fact the main goal of their explorations. Both hOAT4 and mOat5 were shown to promote OTA efflux but again some unexpected effects of exchangeable anions were observed, with glutarate and succinate causing transinhibition of OTA efflux by hOAT4 or mOat5, respectively.
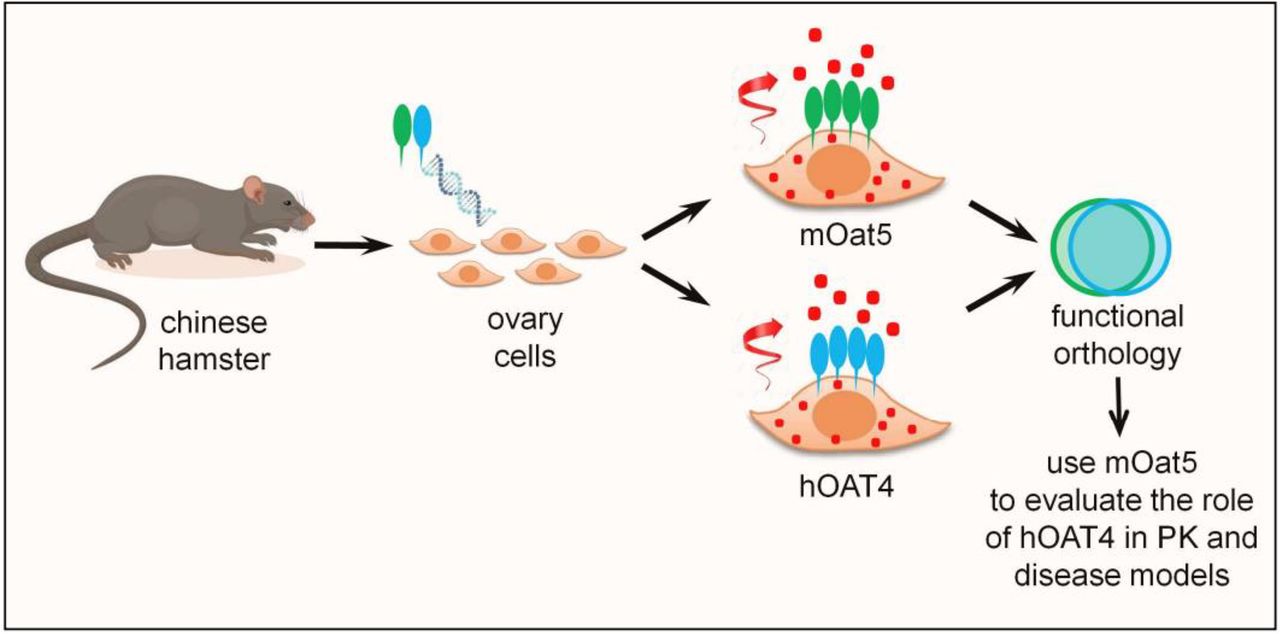
On balance, the work by Martinez-Guerrero et al. (2024) provides substantial advancements for readers conversant in this rather complex field of investigation. To summarize the strengths, the authors developed a thoughtful model for comparing hOAT4 and mOat5 in CHO cells and to demonstrate that the two OATs are functionally ortholog in many respects, including their ability to eject OAs. The transstimulatory effect of exchangeable anions still presents with some inconsistencies, with an apparent differential effect of glutarate or succinate on one OAT or the other; however, this is somewhat better compared with previous studies in which neither glutarate nor succinate caused effects on the transport functions of Oat5 expressed in Xenopus oocytes (Youngblood and Sweet, 2004). These facts pave the road to experiments in which Oat5, constitutively expressed in kidney proximal tubule of rodents, can be used with a reasonable level of confidence to understand how OAT4 works in humans and contribute to renal disposition of endogenous substrates or drugs or environmental agents. The strengths of the model developed by Martinez-Guerrero et al. (2024) are sketched in Fig. 1.

Experimental model developed by Martinez-Guerrero et al. (2024). hOAT4 and mOat5 were stably expressed in CHO cells and shown to be functionally orthologous in ejecting potential substrates. Functional orthology paves the road to using mOat5 to characterize the role of hOAT4 in experimental models of pharmacokinetics or disease.
Some weaknesses must nonetheless be taken into account. The authors suggest that functional orthology between hOAT4 and mOAT5 might foster research and deepen knowledge in the settings of metabolic dysfunction-associated steatohepatitis, a highly prevalent disease reportedly characterized by reduced liver expression of OAT1, OAT3, and OAT4 (Frost et al., 2022). This sounds like an interesting suggestion but many other opportunities were left without a due discussion. For example, it would be of interest to investigate OATs as determinants of efficacy and/or toxicity of cancer drugs like cisplatin, an alkylating agent used to treat urinary bladder cancer and many other tumors. On the one hand, OATs might modulate renal pharmacokinetics of cisplatin, and hence cisplatin activity in urothelial cancer cells (Selim et al., 2023); on the other hand, renal handling of cisplatin might also modulate nephrotoxicity that limits cisplatin treatment of nonrenal tumors (Hu et al., 2017). In rats, increased urinary excretion and reduced tubular expression of Oat5 were interpreted as potential markers of cisplatin-induced nephrotoxicity (Bulacio and Torres, 2015), which raises interest in extending similar concepts to hOAT4. Preclinical evidence of hOAT4 inhibition by other cancer drugs, like epirubicin and chlorambucil, is also available, which heralds drug–drug interactions when the cancer patient is given concomitant drugs liable to uptake or elimination by OAT4 (Toh et al., 2016; Liu et al., 2019). On a different note, the authors failed to emphasize that OAT4 expression and activity can be influenced by a plethora of factors. For example, gene polymorphisms can alter both substrate affinity and carrier efficiency, thus affecting drug disposition, efficacy, and safety (Lozano_et al., 2018; Yee and Giacomini, 2021). A good example of such possibility is given by single nucleotide polymorphisms that reduce OAT4 transport and renal clearance of the loop diuretic, torasemide (Vormfelde et al., 2006; Lozano et al., 2018). Functional orthology between genetic variants of hOAT4 and mOat5 should be investigated before the model developed by Martinez-Guerrero et al. (2024) could be universally exploited for pharmacokinetic investigations.
In sum, the work by Martinez-Guerrero et al. (2024) provides a solid rationale for studies that extend way beyond the otherwise interesting settings of metabolic dysfunction-associated steatohepatitis, and thus represents a must read contribution for anyone conversant in renal disposition of endogenous substances, drugs, and toxins. Functional orthology between nonortholog OATs is a paradox to exploit in many preclinical models of disease or drug disposition, making mOat5 a good molecular player to look at for surmising how hOAT4 would play in the patient or in subjects of the general population.
Footnotes
- Received May 16, 2024.
- Accepted June 10, 2024.
No author has an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- ARB
- angiotensin II receptor blocker
- CHO
- Chinese hamster ovary
- hOAT4
- human organic anion transporter 4
- mOat5
- murine organic anion transporter 5
- OA
- organic anion
- OTA
- ochratoxin
- Copyright © 2024 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}