


Visual Overview
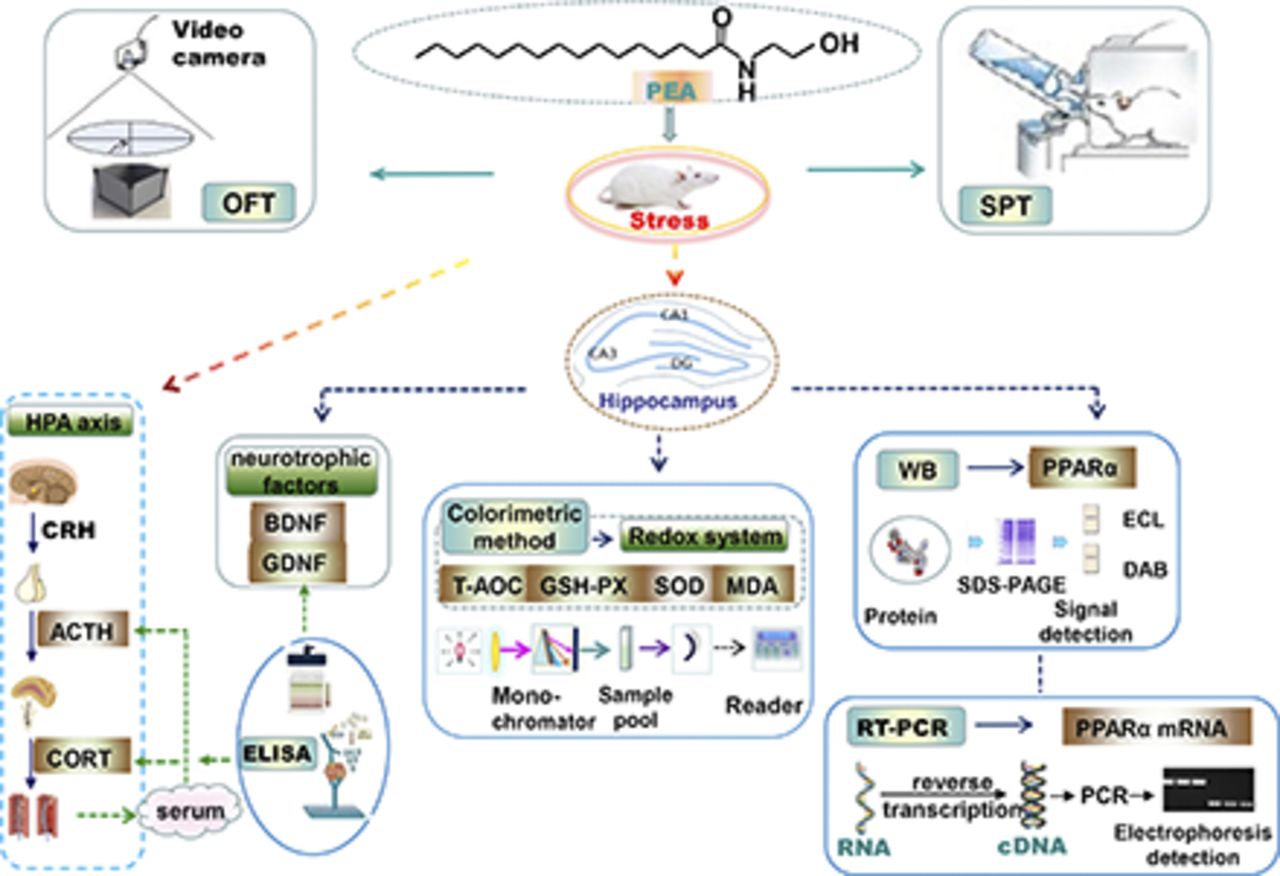
Abstract
N-Palmitoylethanolamide (PEA), an endocannabinoid-like molecule, participates in controlling behaviors associated with mental disorders as an endogenous neuroprotective factor. On the basis of accumulating evidence and our previous data, we tested the hypothesis that the antidepressant-like effects of PEA observed during chronic unpredictable mild stress (CUMS) are mediated by possible targets in the peroxisome proliferator–activated receptor alpha (PPARα) pathway. In this study, rats were subjected to 35 days of CUMS and treated with drugs such as PEA (2.5, 5.0, or 10 mg/kg, by mouth), fluoxetine (10 mg/kg, by mouth), or the combination of PEA and MK886 (1-[(4-chlorophenyl) methyl]-3-[(1,1-dimethylethyl) thio]-α,α-dimethyl-5-(1-methylethyl)-1H-indole-2-propanoic acid). After behavioral tests, the animals were sacrificed and their hippocampi were dissected for subsequent studies. PEA normalized weight gain, sucrose preferences, locomotor activity in an open-field test, and levels of the PPARα mRNA and protein in the hippocampus, and it reduced serum adrenocorticotropic hormone (ACTH) and corticosterone (CORT) levels in rats subjected to CUMS. PEA reversed the abnormal levels of several oxidative stress biomarkers and increased the concentrations of two neurotrophic factors in the hippocampus of CUMS-induced rats. In addition, PEA alleviated the decrease in hippocampal weight. However, the aforementioned effects of PEA were completely or partially abolished by MK886, a selective PPARα antagonist. On the basis of these findings, the PPARα pathway in the hippocampus is a possible target of the antidepressant effects of PEA, and the maintenance of a stable hypothalamic-pituitary-adrenal axis, the antioxidant defenses, and normalization of neurotrophic factor levels in the hippocampus are involved in this process.
Introduction
Depression is characterized by a significant and persistent negative mood and is associated with clinical symptoms such as decreased physical activity, sluggish thoughts, feelings of helplessness, and cognitive dysfunction. Most patients have a tendency to relapse and may even have suicidal attempts or behaviors, placing a substantial mental and economic burden on their families and society (Lueken and Hahn, 2016).
Major depressive disorder (MDD) is now the second leading cause of disability worldwide and contributes to the high mortality associated with many comorbid medical conditions (Singh and Gotlib, 2014). Currently, drug therapy is the main treatment of moderate or severe depressive episodes, but these treatments still have many disadvantages, such as the delayed onset of action and numerous side effects of antidepressants. Therefore, researchers in the medical community have focused on identifying additional drug targets and developing safer and more effective antidepressants (Lueken and Hahn, 2016), and some endogenous substances may have certain therapeutic advantages.
The central endocannabinoid system (ECS) is a neuroactive lipid signaling system in the brain that plays important roles in neuroprotection and reward system activation, among others, by controlling neurotransmitter release (Boorman et al., 2016; Morena et al., 2016). Malfunctions in the central ECS may promote the development and maintenance of psychiatric disorders such as anxiety and depression (Hill and Patel, 2013; Wyrofsky et al., 2015). Several lines of evidence have suggested that facilitation of endocannabinoid signaling within the hippocampus and prefrontal cortex prevents stress-induced behavioral changes, and pharmacological augmentation of endocannabinoid signaling produces both anxiolytic and antidepressant behavioral responses (Lee et al., 2016; Schiavon et al., 2016).
N-Palmitoylethanolamide (PEA) is an endogenous lipid compound that is a member of the endogenous N-acylethanolamide (NAE) family. NAEs function as agonists of the nuclear peroxisome proliferator–activated receptor alpha (PPARα), a nuclear receptor that functions as a transcription factor (Ambrosino et al., 2013; Borrelli et al., 2015). Di Cesare Mannelli and colleagues (2013) found that PEA reduced inflammation in wild-type, but not in mutant, PPAR-α null mice (PPAR-α−/−). And a recent study also demonstrated the pivotal role of PPARα in the PEA pharmacodynamic mechanism to relieve pain (Boccella et al., 2019). These findings strongly suggest that PEA, via a PPARα-mediated mechanism, directly intervenes in the nervous tissue alterations responsible for pain and inflammation. On the basis of accumulating evidence, PEA may function as an endogenous neuroprotective factor and participate in controlling reward-related behaviors (Bilbao et al., 2013; Guida et al., 2015). A recent randomized, double-blind study in depressed patients also indicated a fast antidepressant action of PEA when associated with citalopram (Ghazizadeh-Hashemi et al., 2018). Our previous studies have also confirmed that orally administered PEA is an effective treatment of depression in mouse models subjected to forced stress (Yu et al., 2011). However, the mechanism underlying the antidepressant effects of PEA on stressed subjects is not clear and requires further investigation.
In the present study, the antidepressant-like effects of PEA were examined and compared with the antidepressant fluoxetine (a serotonin reuptake inhibitor) using a rat model of chronic unpredictable mild stress (CUMS). At the same time, a group of CUMS-induced rats was treated with the combination of PEA and MK886 (a selective PPARα antagonist, 1-[(4-chlorophenyl) methyl]-3-[(1,1-dimethylethyl) thio]-α,α-dimethyl-5-(1-methylethyl)-1H-indole-2-propanoic acid) to observe whether the effects of PEA were abolished or diminished by the MK886 pretreatment. Furthermore, the levels of the PPARα mRNA and protein were measured in the hippocampus to determine whether the PPARα pathway is a possible target of the antidepressant effects of PEA. In addition, we determined the serum levels of adrenocorticotropic hormone (ACTH) and corticosterone (CORT) as peripheral biomarkers representing the activity of the hypothalamic-pituitary-adrenal (HPA) axis; total antioxidant capacity (T-AOC), malondialdehyde (MDA) levels, glutathione peroxidase (GSH-PX) activity, and superoxide dismutase (SOD) activity in the hippocampus were evaluated as indicators of oxidative stress. The expression of two neurotrophic factors, brain-derived neurotrophic factor (BDNF) and glial cell line–derived neurotrophic factor (GDNF), in the hippocampus, as well as the decrease in the hippocampal weight, were measured to assess the occurrence of and improvements in hippocampal atrophy (Abdallah et al., 2015). Possible correlations between these factors analyzed in the present experiment were determined to further explore the possible mechanism underlying the antidepressant effects of PEA and its effective targets.
Materials and Methods
Compounds and Drugs.
PEA (chemical structure is shown in Fig. 1; synthesized at the College of Pharmacy, Yanbian University, China) or fluoxetine (Enhua, Jiangsu, China) was suspended in 0.3% methyl cellulose (Loba–Chemie, Shanghai, China) and administered daily via gavage. MK886 (Han Xiang, Shanghai, China) was dissolved in sterile saline and administered daily via an intraperitoneal injection. All doses are presented as milligrams per kilogram body weight.

Structure of N-palmitoylethanolamide (PEA).
Animals.
Experimental procedures were approved by the Institutional Animal Care and Use Committee of China and were conducted in accordance with the Yanbian University of Health Guide for Care and Use of Laboratory Animals. Male Sprague-Dawley rats (180–200 g; the Experimental Animal Center, Yanbian University, Jilin, China) were housed individually in cages for 3 days to allow adaptation to the environment under the following controlled conditions: 12-hour light/12-hour dark cycle, free access to tap water and food pellets, and ambient temperature and relative humidity at 22 ± 2°C and 55% ± 5%, respectively.
Experimental Procedures.
All animals were randomly divided into one of the following two groups: a CUMS-treated group and a normal control (NC) group. The CUMS-treated rats were subjected to a series of variable stressors described by Jin et al. (2015). The animals were housed individually (330 × 215 × 200 mm) and subjected to the following stressors: 1) horizontal shock for 5 minutes (one shock/s); 2) swimming in 4°C water for 5 minutes; 3) swimming in 45°C water for 5 minutes; 4) food deprivation for 24 hours; 5) water deprivation for 24 hours; 6) exposure to wet bedding for 24 hours; 7) housing with other rats (five rats per cage) for 17 hours; 8) placement in an empty cage for 17 hours; 9) cage tilted at 45° for 24 hours; 10) foot shock for 1 minute (voltage 50 mV, frequency 0.05 Hz); 11) tail suspension for 1 minute; 12) foreign object placed in the cage for 6 hours; 13) light flashes for 10 minutes; and 14) reversed light/dark cycles (overnight illumination). The 14 stressors were scheduled semirandomly, with animals subjected to two different stressors daily. The rats in the CUMS group were exposed to the stressors for 35 days, and the same stressor was not administered on consecutive days to ensure that the rats were not able to anticipate the occurrence of specific stressors.
Untreated rats served as the NC group and were housed in a separate room in groups of five rats per cage (330 × 215 × 200 mm). Other than withdrawing food and water for 24 hours before the sugar preference experiment and withdrawing food for 6 hours before weighing, the rats were not subjected to additional stimuli. All behavioral tests were performed between 9:00 and 11:00 AM in a quiet, warm environment and were conducted in parallel for each experimental group. Biochemical parameters were measured on the last day of the experiment. After CUMS was induced for 7 days, the CUMS-treated rats were randomly divided into the following six groups (n = 10): 1) the CUMS control (vehicle) group; 2) 10-mg/kg fluoxetine group; 3) 2.5-mg/kg PEA group; 4) 5-mg/kg PEA group; 5) 10-mg/kg PEA group; and 6) 10-mg/kg PEA + 3-mg/kg MK886 group. Rats in these groups were treated with fluoxetine or PEA in 0.3% sodium carboxymethyl cellulose via gavage and MK886 via intraperitoneal injections daily for 28 days. The entire experimental period was 35 days (Fig. 2).

Experimental procedures. Animal grouping and treatments (A). Experimental procedures (B). Animals were exposed to CUMS for 35 days; drug treatments were administered for 28 days. Vehicle, PEA (2.5, 5, 10 mg/kg), fluoxetine (Fxt; 10 mg/kg), or MK886 (MK; 3 mg/kg) were administered for 28 days starting on day 8 after initiating the CUMS procedures.
Weight Monitoring.
The body weight gains reflect an animal’s appetite and digestive function. The body weights of the rats were recorded regularly once a week throughout the experimental period.
Sucrose Preference Test.
The sucrose preference test is used to assess the anhedonia of animals and indicates an animal’s reward sensitivity and depressive state (Remus et al., 2015). Initially, the rats were acclimated to a sucrose solution for 3 days before the test. On the 1st day, food and water were withheld for 24 hours; on the 2nd day, rats were exposed to a 1% sucrose solution (w/v) for 24 hours. On the 3rd day, a bottle of the 1% sucrose solution and a bottle of tap water were presented to each rat at random positions for 24 hours. During the test, all rats were exposed to the 1% sucrose solution (w/v) and tap water presented at random positions for 4 hours, followed by food and water deprivation for 24 hours. The bottles containing tap water or the sucrose solution were prepared 24 hours before the sucrose preference test and were stored in the same room to avoid leakage. The sucrose preference was calculated using the following formula: sucrose preference percentage (%) = sucrose consumption (g)/water consumption (g) + sucrose consumption (g) × 100.
Open-Field Test.
The open-field test (OFT) is commonly used to investigate the exploratory behavior and spontaneous activity of experimental animals (Jin et al., 2015). The OFT was performed using the TM-Vision behavioral experiment system (Tai-Meng, China). The OFT-100 experimental apparatus consisted of a nontransparent plastic container (625 × 740 × 510 mm) with an experimental area of 525 × 525 × 415 mm. The entire experimental process was recorded using a computer-controlled camera. The experimental area was divided into four active parts, namely, the sides, corners, perimeter, and center. Each rat was individually placed in the center of the open-field apparatus and allowed to explore its surroundings while the following parameters were recorded for 3 minutes: immobility time, locomotor time, and distance traveled. The walls and floor surfaces were thoroughly cleaned with 10% ethanol between the tests to remove possible biases caused by odor cues from previous rats.
Western Blot Analysis.
Nuclear proteins were extracted from rat brain tissues using a nuclear protein extraction kit (Beyotime, China), and the protein concentrations were detected using a commercially available BCA protein assay kit (Beyotime). Initially, 20 μg of the nuclear protein sample in lane marker loading buffer (Cwbiotech Company) were denatured at 100°C for 8 minutes. Brain lysates were separated on 10% Tris-glycine SDS-PAGE gel and transferred to a polyvinylidene fluoride membrane (0.45 μm pores; Millipore, Germany) using a wet electroblotting approach. The membrane was blocked with 5% (w/v) fat-free milk in PBS for 2 hours at room temperature and then incubated overnight at 4°C with the following primary antibodies: rabbit anti-PPAR-α (1:250; Sigma-Aldrich) and monoclonal mouse anti-β-actin (1:500; Sigma-Aldrich) as an internal loading control. All antibodies were diluted in Tris-buffered saline solution containing 0.1% Tween-20 (TBST). After thorough washes with TBST, the blots were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies (goat anti-rabbit IgG, 1:750, and goat anti-mouse IgG, 1:1000; Sigma-Aldrich) for 2 hours at room temperature, and then, the blots were developed using an enhanced chemiluminescence detection kit (ECL Plus; Beyotime) for 20 seconds and visualized using the lane one-dimensional gel analysis system (Beijing Sage Creation Science, Beijing, China). For densitometry analyses, the bands were scanned and quantified using lane gel analysis software (Beijing Sage Creation Science), and the results were expressed as the ratio of target protein to β-actin immunoreactivity, namely, the relative density value of PPARα/β-actin (the ratio of integrated density value, IDV).
Total RNA Extraction and Reverse Transcription–Polymerase Chain Reaction Analysis.
Total RNA was extracted from the rat hippocampi, purified using an RNAiso Plus reagent kit (Takara, Japan) according to the manufacturer’s instructions, and quantified using a Micro-UV spectrophotometer (Q5000; Huayunante Technology). First-strand complementary DNAs (cDNAs) were synthesized from 2 μg of total RNA using a Primescript RT-PCR kit (Takara) according to the manufacturer’s instructions, and these cDNAs were used as the template for the reverse transcription–polymerase chain reaction analysis. The cDNA templates were then amplified by polymerase chain reaction in a 20 μl total reaction volume using a StepOne real-time PCR system (Applied Biosystems, CA). The primer sequences (Invitrogen) were as follows: PPARα, 5′-AGTGCCTGTCCGTCGGGAT-3′ (forward) and 5′-GCTAGTCTTTCCTGCGAGTATG-3′ (reverse) (222 bp). As an internal control, the GAPDH cDNA was coamplified using the primer sequences 5′-GTGCCAGCCTCGTCTCATAG-3′ (forward) and 5′-CTTGCCGTGGGTAGAGTCAT-3′ (reverse) (184 bp). The amplification was performed with the following parameters: preheating at 95°C for 5 minutes, followed by 30 cycles of denaturation at 95°C for 30 seconds, annealing at 52°C for 30 seconds, and extension at 72°C for 1 minute. The reaction was terminated by a final extension step at 72°C for 10 minutes. The PCR products were separated on 2.5% agarose gels, and the density of each band was analyzed using a one-dimensional gel analysis system (Sage Creation Science). The expression of the gene of interest was normalized to GAPDH. The relative integrated density value was computed as PPARα/GAPDH. The threshold cycle (CT) of PPARα production was determined and normalized to the GAPDH internal standard. The samples obtained from four independent tissues were used to analyze relative gene expression with the 2−ΔΔCT method.
Determination of ACTH and CORT Levels.
ELISAs were used to detect serum ACTH and CORT levels. The steps described below were performed according to the instructions provided with the rat ACTH and CORT ELISA kits (Shanghai Enzyme linked Biotechnology, China). We added 50 μl of the standard to the standard well and added 10 μl of the test sample and 40 μl of the sample diluent to the test well. We then added 100 μl of the HRP-conjugated reagent to each well, covered it with an adhesive strip, and incubated the plate at 37°C for 60 minutes. After the last washing step, we removed the remaining wash solution and then added 50 μl each of the chromogens A and B to each well. We gently mixed it and incubated it for 15 minutes at 37°C, followed by adding 50 μl of stop solution. The color in the wells changed from blue to yellow. We read the optical density (OD) at 450 nm using a microplate reader (LIUYI, Beijing, China) within 15 minutes. The serum ACTH and CORT concentrations are reported in picograms per milliliter and nanograms per milliliter, respectively.
Determination of T-AOC, GSH-PX, T-SOD and MDA Levels.
The T-AOC and MDA levels and the GSH-PX and total superoxide dismutase (T-SOD) activities in the supernatants of the hippocampal homogenates were determined using commercial analysis kits (Nanjing Jiancheng, China) according to the manufacturer’s instructions. Many endogenous antioxidants reduce Fe3+ to Fe2+, the latter of which forms a stable compound with phenanthroline; therefore, T-AOC levels were measured using a colorimetric method with a spectrophotometer at the visible wavelength of 520 nm (V-5800; METASH, China). A T-AOC unit in tissue is defined as follows: at 37°C, each milligram of tissue protein per minute increases the OD value of the reaction system by 0.01. T-AOC levels are reported as units per milligram of tissue protein. GSH-PX promotes the reaction of hydrogen peroxide (H2O2) with reduced glutathione to produce H2O and oxidized glutathione (GSSG); therefore, the activity of GSH-PX is determined by the rate of its enzymatic reaction. GSH-PX activity is reported as units per milligram of protein. The xanthine oxidase method was used for the T-SOD assays. Superoxide radicals (O2−) are generated by xanthine and the xanthine oxidase reaction system, which oxidizes hydroxylamine to form nitrite. This reaction produces a purplish red color via the chromogenic agent, and the absorbance is measured with a spectrophotometer at visible wavelengths. T-SOD activity is presented as units per milligram of protein. MDA levels were determined using the thiobarbituric acid (TBA) method. MDA levels in lipid peroxidation degradation products are condensed with TBA to form red products, resulting in a maximal absorption peak at 532 nm, and its OD value was measured using a colorimetric method at a wavelength of 532 nm. The MDA level is reported as nanomoles per milligram tissue protein.
Determination of BDNF and GDNF Levels.
The hippocampal BDNF and GDNF concentrations were determined using ELISAs according to the manufacturer’s instructions (Sbjbio, China). Briefly, the following steps were performed. Fifty microliters of each of the standards were added to respective wells at concentrations of 90, 60, 30, 15, and 7.5 ng/l. We then added 10 μl of the test sample and 40 μl of the diluent to the test well. After an incubation and washes, we added 50 μl of the HRP-conjugated reagent. We then added 50 μl each of chromogen A and chromogen B and incubated the plate in the dark for 15 minutes. We added 50 μl of stop solution to stop the reaction. We read the OD at 450 nm using a microplate reader (LIUYI) within 15 minutes. The BDNF and GDNF concentrations are presented as picograms per milligram of brain tissue.
Hippocampal Weight and the Hippocampus Index.
On the last day of the experiment, the rats were anesthetized with 10% chloral hydrate (4 ml/kg; Chaoyang, Beijing, China) and sacrificed by decapitation. After rapidly removing and weighing the whole brain of each rat, the bilateral hippocampi were immediately dissected on an ice-cold table. The hippocampus index (the ratio of hippocampus weight to whole brain weight) was calculated using the following formula: the hippocampus index (%) = hippocampus weight (mg)/whole brain weight (mg) × 100%.
Results
Body Weight and Behavioral Changes in Rats after Exposure to CUMS and Drug Treatment.
After 35 days of CUMS and 28 days of PEA (2.5, 5, 10 mg/kg) or fluoxetine (10 mg/kg) treatment, the body weights and depression-like behaviors of the rats were examined (Fig. 3). The behavioral changes of the CUMS-treated rats were evaluated using the sucrose preference test and the open-field test. As shown in Fig. 3, the normal control (NC) group and the CUMS treatment group exhibited similar physiologic and behavioral indicators, as evidenced by their body weights (Fig. 3A), sucrose preference (Fig. 3B), and locomotor activities (data not shown) before CUMS induction. Thus, before the CUMS treatment, the basal body weights and behavioral parameters related to depression in each group of rats were similar. After 35 days of CUMS, the body weights (Fig. 3A), sucrose preferences (Fig. 3B), distance traveled and locomotor time (Fig. 3, C and D) were all significantly decreased, and the total immobility time was significantly increased in the rats in the vehicle group compared with the NC group. These results indicated the successful establishment of a chronic stress-induced depression model in rats. After 28 days of PEA or fluoxetine treatment, the body weight (Fig. 3A), sucrose preference (Fig. 3B), total distance traveled (Fig. 3C), and the total locomotor time (Fig. 3D) were all significantly increased, and the total immobility time (Fig. 3D) was significantly reduced in the 2.5-, 5-, and 10-mg/kg PEA groups and in the fluoxetine (10 mg/kg) group compared with the vehicle group. Nevertheless, significant differences in the aforementioned behavioral effects were not observed between the group that was coadministered MK886 and PEA and the vehicle group (Fig. 3). The group treated with the combination of MK886 and PEA showed significant differences compared with the group treated with PEA (10 mg/kg) alone (Fig. 3, A, B, and D), as well as with the NC group (Fig. 3, A, B, and D). On the basis of these findings, MK886, a selective PPARα antagonist, completely or partially abolished the effects of PEA.

Changes in body weight and behavioral indicators during the CUMS procedure and after PEA treatment. The body weights of the rats (A) and the results of the sucrose preference test (B) before CUMS and after 35 days of CUMS and 28 days of drug treatment are shown. The effect of PEA on the OFT performance (C and D) of CUMS rats after 35 days of CUMS and 28 days of drug treatment; the distance traveled (C) in the OFT, the locomotor time and the immobility time (D) in the OFT are shown. Veh (vehicle control group), Fxt (10 mg/kg fluoxetine group), PEA 2.5, 5, and 10 (2.5-, 5-, and 10-mg/kg PEA groups), PEA + MK886 (10 mg/kg PEA po + 3 mg/kg MK886 i.p.). Data are presented as the means ± S.E.M., n = 10. #P < 0.05; ##P < 0.01; ###P < 0.001 vs. NC; *P < 0.05; **P < 0.01; ***P < 0.001 vs. Veh; △P < 0.05 vs. 10 mg/kg PEA group.
PEA Restored the Nuclear Expression Patterns of the PPARα Protein and mRNA in the Hippocampus of CUMS-Induced Rats.
As shown in Fig. 4, after 35 days of CUMS, the nuclear expression of the PPARα protein and mRNA were both significantly increased in the hippocampus of CUMS-induced rats compared with those in the NC group. When animals were treated with drugs during the last 28 days of the experimental period, significant decreases in the nuclear levels of the PPARα protein (Fig. 4, A and C) were observed in the hippocampus of the PEA 2.5-, 5-, and 10-mg/kg treatment groups, and significant decreases in mRNA levels (Fig. 4, B and D) were observed in the PEA 5- and 10-mg/kg treatment groups compared with the vehicle group, but significant differences were not observed compared with the NC group. Thus, PEA normalized the nuclear levels of the PPARα protein and mRNA in the hippocampus of CUMS-induced rats, but a similar effect was not observed in the positive control treated with fluoxetine. Nevertheless, the effects of PEA were abolished by MK886 compared with those for PEA treatment alone. On the basis of these data, the PPARα pathway may be involved in the antidepressant effects of PEA.

Effects of PEA on nuclear levels of the PPARα protein and mRNA in the hippocampus of CUMS-induced rats after 35 days of CUMS and 28 days of drug treatment. Levels of the PPARα protein (A and C) were measured using Western blotting and normalized to β-actin levels. The histograms show the relative levels of the PPARα protein. Levels of the PPARα mRNA (B and D) were measured using reverse transcription–polymerase chain reaction. The level of the PPARα mRNA was normalized to GAPDH expression, and the histograms show the relative expression of the PPARα mRNA. The grouping and abbreviation of Veh, Fxt, and 2.5, 5, and 10 PEA groups and the PEA + MK886 group are the same as described in Fig. 3. Data are presented as the means ± S.E.M., n = 4. ##P < 0.01 vs. NC; *P < 0.05; **P < 0.01 vs. Veh; △△P < 0.01 vs. 10 mg/kg PEA group.
PEA Treatment Reduced the Serum ACTH and CORT Levels in CUMS-Induced Rats.
As shown in Fig. 5, after 35 days of CUMS treatment, the serum ACTH and CORT concentrations in rats were significantly increased compared with the NC group. After 28 days of drug treatment, ACTH and CORT concentrations in the fluoxetine (10 mg/kg) and PEA (2.5, 5 or 10 mg/kg) groups were significantly decreased compared with the vehicle group. Therefore, CUMS induced the HPA axis hyperactivity that was normalized by PEA. Furthermore, the CORT level was significantly increased in the group administered the combination of MK886 and PEA compared with that in the NC group or the group treated with PEA alone; however, no significant differences in ACTH levels were observed between the group that was coadministered MK886 and PEA, the NC group, and the group treated with PEA alone. These results suggested that the antagonistic effect of PEA on the HPA axis hyperactivity may be related to the PPARα signaling pathway.

Effects of PEA on serum ACTH and CORT levels in CUMS-induced rats. The serum ACTH and CORT levels were measured using ELISAs. The serum ACTH (A) and CORT (B) concentrations are reported in picograms per milliliter and nanograms per milliliter, respectively. The grouping and abbreviation of Veh, Fxt, and 2.5, 5, and 10 PEA groups and the PEA + MK886 group are the same as described in Fig. 3. Data are presented as the means ± S.E.M., n = 8. #P < 0.05; ##P < 0.01; ###P < 0.001 vs. NC; *P < 0.05; **P < 0.01; ***P < 0.001 vs. Veh; △P < 0.05 vs. 10 mg/kg PEA group.
PEA Treatment Increased the T-AOC Level, GSH-PX Activity, and T-SOD Activity and Reduced the MDA Content in the Hippocampus of CUMS-Induced Rats.
After 35 days of CUMS, the T-AOC level, GSH-PX activity and T-SOD activity were significantly decreased (Fig. 6, A–C) and the MDA concentrations were significantly increased (Fig. 6D) in the hippocampus of CUMS-induced rats compared with those in the control group. An explanation for this finding is that chronic stress disrupted the balance between the hippocampal oxidative and antioxidant systems and decreased the antioxidant capacity. After 28 days of drug treatment, the fluoxetine- (10 mg/kg) and PEA- (2.5-, 5-, and 10-mg/kg) treatment groups exhibited significant increases in GSH-PX and SOD activities and decreased MDA concentrations in the hippocampus. The PEA- (5- and 10-mg/kg) treatment groups exhibited significantly increased T-AOC levels, whereas the fluoxetine- (10 mg/kg) and PEA- (2.5 mg/kg) treatment groups did not show differences in the T-AOC levels, indicating that PEA exerted a dose-dependent effect on improving T-AOC levels compared with the vehicle treatment. Nevertheless, significant differences in the biochemical indices listed above were not observed between rats that were coadministered MK886 and PEA compared with the vehicle group (Fig. 6). However, the MK886 plus PEA–coadministration group showed significant differences in the T-AOC level, GSH-PX activity, and MDA concentrations compared with rats treated with PEA alone (Fig. 6, A, B, and D), suggesting that PEA partially restored the antioxidant capacity of the hippocampus and that MK886 may partially abolish these effects of PEA.

Effects of PEA on representative markers of oxidative stress in the rat hippocampus. The total antioxidant capacity (A), glutathione peroxidase activity (B), superoxide dismutase activity (C), malondialdehyde levels (D) were analyzed. T-AOC levels, GSH-PX activity, and T-SOD activity are reported as units per milligram of protein in tissues, and the MDA level is presented as nanomoles per milligram of protein in tissues. The grouping and abbreviation of Veh, Fxt, and 2.5, 5, and 10 PEA groups and the PEA + MK886 group are the same as described in Fig. 3. Data are presented as the means ± S.E.M., n = 8. ##P < 0.01; ###P < 0.001 vs. NC; *P < 0.05; **P < 0.01 vs. Veh; △P < 0.05 vs. 10 mg/kg PEA group.
PEA Treatment Increased the BDNF and GDNF Levels in the Hippocampus of CUMS-Induced Rats.
As shown in Fig. 7, after 35 days of CUMS and 28 days of drug treatment, BDNF and GDNF concentrations were significantly reduced in the hippocampus of rats in the vehicle group compared with those in the NC group, suggesting that CUMS decreased the levels of some neurotrophic factors in the rat hippocampus. Fluoxetine (10 mg/kg) and PEA (5 or 10 mg/kg) significantly increased the levels of BDNF and GDNF, and the BDNF concentration was also increased in the PEA 2.5-mg/kg treatment group compared with the vehicle group. However, significant differences in the BDNF and GDNF levels were not observed between rats coadministered MK886 and PEA and the group treated with PEA alone or the vehicle group, but a significantly lower BDNF content was observed than in the NC group. Thus, the PPARα pathway may be partially involved in the regulatory effect of PEA on neurotrophic factors in the rat hippocampus.

Effects of PEA on BDNF and GDNF levels in the hippocampus of CUMS-induced rats. The BDNF and GDNF levels were determined using ELISAs. The BDNF (A) and GDNF (B) concentrations are reported in picograms per milligram of brain tissue. The grouping and abbreviation of Veh, Fxt, and 2.5, 5, and 10 PEA groups and the PEA + MK886 group are the same as described in Fig. 3. Data are presented as the means ± S.E.M., n = 8. #P < 0.05; ##P < 0.01; ###P < 0.001 vs. NC; *P < 0.05; **P < 0.01 vs. Veh group.
PEA Treatment Alleviated the Decrease in the Hippocampal Weight in CUMS-Induced Rats.
After 35 days of CUMS, the hippocampal weight (Fig. 8A) and the hippocampus index (the ratio of hippocampus weight to whole brain weight, Fig. 8B) were significantly decreased in CUMS-induced rats. After 28 days of drug treatment, the fluoxetine- (10 mg/kg) and PEA- (2.5, 5, 10 mg/kg) treatment groups exhibited significant increases in the hippocampal weight (Fig. 8A) and the hippocampus index (Fig. 8B) compared with those of the vehicle group. Nevertheless, significant differences in the hippocampal weight and hippocampus index were not observed between rats treated with MK886 and the vehicle group. However, the hippocampus index of the group pretreated with MK886 was significantly different from the rats treated with PEA alone (Fig. 8B). On the basis of these data, PEA improved the phenotype of hippocampal atrophy to some extent, and more neuronal atrophy may occur in the hippocampus than in other brain tissues following CUMS. The effect of PEA on preventing hippocampal atrophy was partially reversed by MK886.

Effects of PEA on the hippocampus weight (A) and hippocampus index (B) in CUMS-induced rats. The whole brain and the bilateral hippocampi of the rats were rapidly removed and weighed. The weights of the brain and hippocampus are reported separately in milligrams. The grouping and abbreviation of Veh, Fxt, and 2.5, 5, and 10 PEA groups and the PEA + MK886 group are the same as described in Fig. 3. Data are presented as the means ± S.E.M., n = 8. ##P < 0.01 vs. NC; *P < 0.05; **P < 0.01 vs. Veh; △P < 0.05 vs. 10 mg/kg PEA group.
Discussion
Many social, environmental, and psychologic factors result in depression in humans. The CUMS animal model of depression was employed to mimic negative life events to which humans are exposed, and it is accompanied by two core symptoms, namely, the depressive state and anhedonia (Liu et al., 2016). In the present study, after 35 days of chronic stress exposure, the decreased sucrose preferences, exploratory behaviors, and body weight gains indicated that behavioral impairments and physiologic dysfunction in rats were similar to the symptoms observed in patients with depression, such as a loss of euphoria, loss of acute activity and curiosity, and weakening of appetite and digestive system function. After 28 days of PEA treatment, the rats’ sucrose preferences, exploratory behaviors, and body weight gains were increased, indicating improvements in reward sensitivities, acute activity, and curiosity, as well as in the appetite and emotions of the rats. Moreover, after coadministration of MK886, the aforementioned effects of PEA were abolished to different extents. Thus, PEA improved the depressive-like behaviors and the body weight loss of CUMS-induced rats, and its activity may be related to the PPARα signaling pathway.
The levels of the PPARα mRNA and protein were measured in the rat hippocampus to further confirm that the PPARα signaling pathway is a putative target of the antidepressant effects of PEA. PPARs are ligand-activated transcription factors that regulate lipid metabolism and energy homeostasis. PPARα is one of three subtypes of the nuclear PPAR family and is expressed by neurons in many regions of the brain (Roy and Pahan, 2015). PPARα plays an important role in regulating lipid synthesis and degradation through its ability to control the expression of key transport proteins and enzymes, and it is activated by endogenous ligands, including PEA (Hesselink, 2013). As shown in the present study, CUMS exposure upregulated PPARα expression at both the mRNA and protein levels, and the PEA treatment normalized the PPARα levels in the rat hippocampus. These effects of PEA were abolished by the MK886 pretreatment. Thus, exogenous PEA directly inhibited the expression of the PPARα receptor in the hippocampus of rats exposed to CUMS. Furthermore, after analyzing these data, we hypothesized that CUMS may suppress the synthesis of endogenous PEA or increase the consumption of PEA, leading to a decrease in the PEA content within the hippocampus. A long-term low level of PEA, which is an endogenous PPARα ligand, may induce a compensatory upregulation of PPARα expression. This upregulation may explain why elevated expression of the PPARα mRNA and protein was observed in CUMS-treated rats in the present study.
Notably, by supplementing rats with exogenous PEA, the low level of PEA was restored to normal levels, enabling levels of PPARα expression to return gradually to normal, as we observed in this study. However, MK886 prevented the ligand PEA from signaling through its receptor PPARα, and thus, the hippocampal levels of the PPARα mRNA and protein in the group treated with the combination of MK886 and PEA were similar to those in rats exposed to CUMS alone. We conducted another study to further confirm our hypothesis that the PPARα pathway is involved in the antidepressant-like effects of PEA in rats and to determine possible mechanisms underlying the antidepressant effects of PEA.
The HPA axis represents the main neuroendocrine system responsible for regulating the stress response and is a major integrated system that maintains body homeostasis (Gray et al., 2016). Chronic stress is known to alter the physiologic function of the HPA axis and plays a key role in the development of neuropsychiatric disorders (Yang et al., 2017). Alterations in the HPA axis (mainly elevated levels of stress-related hormones) have been observed in animal models of depression and patients with depression (Zhao et al., 2018). Serum ACTH and CORT levels were significantly increased in rats exposed to CUMS, suggesting that chronic stress increased the circulating levels of biomarkers of the HPA axis and induced persistent hyperactivity of the HPA axis. Termination of the response of the HPA axis to stress is mediated by multiple negative feedback loops, and recent studies have suggested that the ECS in the hippocampus participates in the regulation of negative feedback of the HPA axis through cannabinoid receptor type 1 (Hill and Tasker, 2012; Citraro et al., 2013). Our data also confirmed that PEA functions as an endocannabinoid analog to significantly decrease ACTH and CORT levels in CUMS-induced rats and that pretreatment with MK886 attenuated the effect of PEA. Thus, PEA ameliorated the hyperactivity of the HPA axis and participated in maintaining its homeostasis to ameliorate the consequences of stress, and the PPARα signaling pathway might also participate in this mechanism.
In response to stress-induced activation of the HPA axis, glucocorticoids are released and bind to the glucocorticoid receptor to increase glucose availability and the metabolic rate, which simultaneously increase the spontaneous production of free radicals (Lipina and Hundal 2016). When the neural antioxidant defenses of the organism are inadequate to counter the reactive oxygen species, oxidative homeostasis is disrupted (Bakunina et al., 2015), leading to oxidative stress. The accumulated free radicals induce a chain of neurobiological events and damage macromolecules, including DNA, proteins, and other molecules, either in the cytosol, nucleus, and/or mitochondria, along with a dysregulation of normal metabolism, ultimately resulting in neuronal death by apoptosis and/or necrosis (Brown et al., 2014). Furthermore, the hippocampus contains a high density of glucocorticoid receptors; this tissue is considered a target of psychologic stress and displays structural plasticity after both acute and chronic stress (Gobinath et al., 2016). In this study, we detected the levels of several representative and reliable markers of oxidative stress in the hippocampus. We observed increased levels of MDA, a typical product of lipid peroxidation, and decreased activity of SOD and GSH-PX (two key antioxidant enzymes, Palta et al., 2014; Rodrigues et al., 2014) and levels of T-AOC (a marker of the nonenzymatic and enzymatic antioxidant systems, reflecting the antioxidant capacity of the body, Talarowska et al., 2014) in the hippocampus of CUMS-induced rats. Chronic oral administration of PEA reversed the changes in the levels of these biomarkers, as mentioned above, but the MK886 pretreatment reversed the effects of PEA. On the basis of these data, PEA enhanced the antioxidant capacity of the hippocampus in CUMS-induced rats, and the PPARα pathway may have influenced this effect of PEA.
BDNF and GDNF, two important members of the neurotrophin family (Levy et al., 2018), are involved in the survival, differentiation, and growth of neurons (Halappa et al., 2018) and are considered two pivotal biomarkers of mood disorders. BDNF and GDNF participate in the neurobiology of depression and are involved in the therapeutic effects of antidepressants (Park and Lee, 2018). We observed decreases in the BDNF and GDNF levels in the hippocampus of CUMS-induced rats and increased concentrations after the chronic PEA treatment. MK886 reversed these effects of PEA, confirming the participation of the PPARα signaling pathway.
In addition, other researchers have confirmed neuronal atrophy in the granular cell and pyramidal cell layers of the hippocampus, along with a reduction in hippocampal volume in both patients with depression and animal models (Abdallah et al., 2015; Hacimusalar and Eşel, 2018). Likewise, in this experiment, the weight of rat hippocampus decreased in response to CUMS, indirectly indicating hippocampal atrophy. PEA treatment alleviated the decrease in the hippocampal weight, and the MK886 pretreatment partially reversed this effect of PEA, suggesting that PEA may ameliorate hippocampal atrophy and that the PPARα-signaling pathway is most probably involved in this process.
On the basis of the experimental results described above, chronic mild stress activated the HPA axis, produced an imbalance in hippocampal oxidative stress markers, and reduced the expression of two important neurotrophic factors. These changes all contributed to atrophy and cell loss in the hippocampus, which was manifested as a decrease in the hippocampus weight. The aforementioned changes were simultaneously associated with increased levels of PPARα mRNA and protein in the hippocampus, along with a depression-like behavioral phenotype in rats. Chronic treatment with PEA improved the depressive behaviors in animals, normalized the levels of PPARα mRNA and protein in the hippocampus, and was positively correlated with the normalization of the aforementioned biochemical indicators or biomarkers. At the same time, the effects of PEA were completely or partially abolished by MK886. These findings all further confirmed our hypothesis that the PPARα pathway is involved in the antidepressant-like effects of PEA on rats. If we could directly detect the correlation between PEA levels and PPARα expression in the hippocampus, our hypothesis would be more completely confirmed. This possibility should be studied further in future experiments, and its upstream and downstream targets require further exploration.
In conclusion, the PPARα pathway in the hippocampus is a possible target of the antidepressant effects of PEA; furthermore, the stabilization of the HPA axis and the antioxidant defenses, as well as the normalization of levels of neurotrophic factors in the rat hippocampus, are involved in this process.
Acknowledgments
The compound was synthesized and supplied free of charge by Zhe-shan Quan at the Yanbian University School of Pharmacy, Jilin Province, China.
Authorship Contributions
Participated in research design: Yu, Jiang.
Conducted experiments: Li, Wang, Bi, Piao.
Performed data analysis: Yu, Wang, Jiang.
Wrote or contributed to the writing of the manuscript: Yu, Wang.
Footnotes
- Received October 23, 2018.
- Accepted January 4, 2019.
This work was supported by the National Natural Science Foundation of China (Grant 81460217 and 81760650). The authors declare that there are no conflicts of interest relating to these studies.
Abbreviations
- ACTH
- adrenocorticotropic hormone
- BDNF
- brain-derived neurotrophic factor
- CORT
- corticosterone
- CUMS
- chronic unpredictable mild stress
- ECS
- endocannabinoid system
- GDNF
- glial cell line–derived neurotrophic factor
- GSH-PX
- glutathione peroxidase
- HPA
- hypothalamic-pituitary-adrenal axis
- HRP
- horseradish peroxidase
- MDA
- malondialdehyde
- NC
- normal control
- OD
- optical density
- OFT
- open-field test
- PEA
- N-palmitoylethanolamide
- PPARα
- peroxisome proliferator–activated receptor alpha
- SOD
- superoxide dismutase
- T-AOC
- total antioxidant capacity
- T-SOD
- total superoxide dismutase
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}