


Visual Overview
Abstract
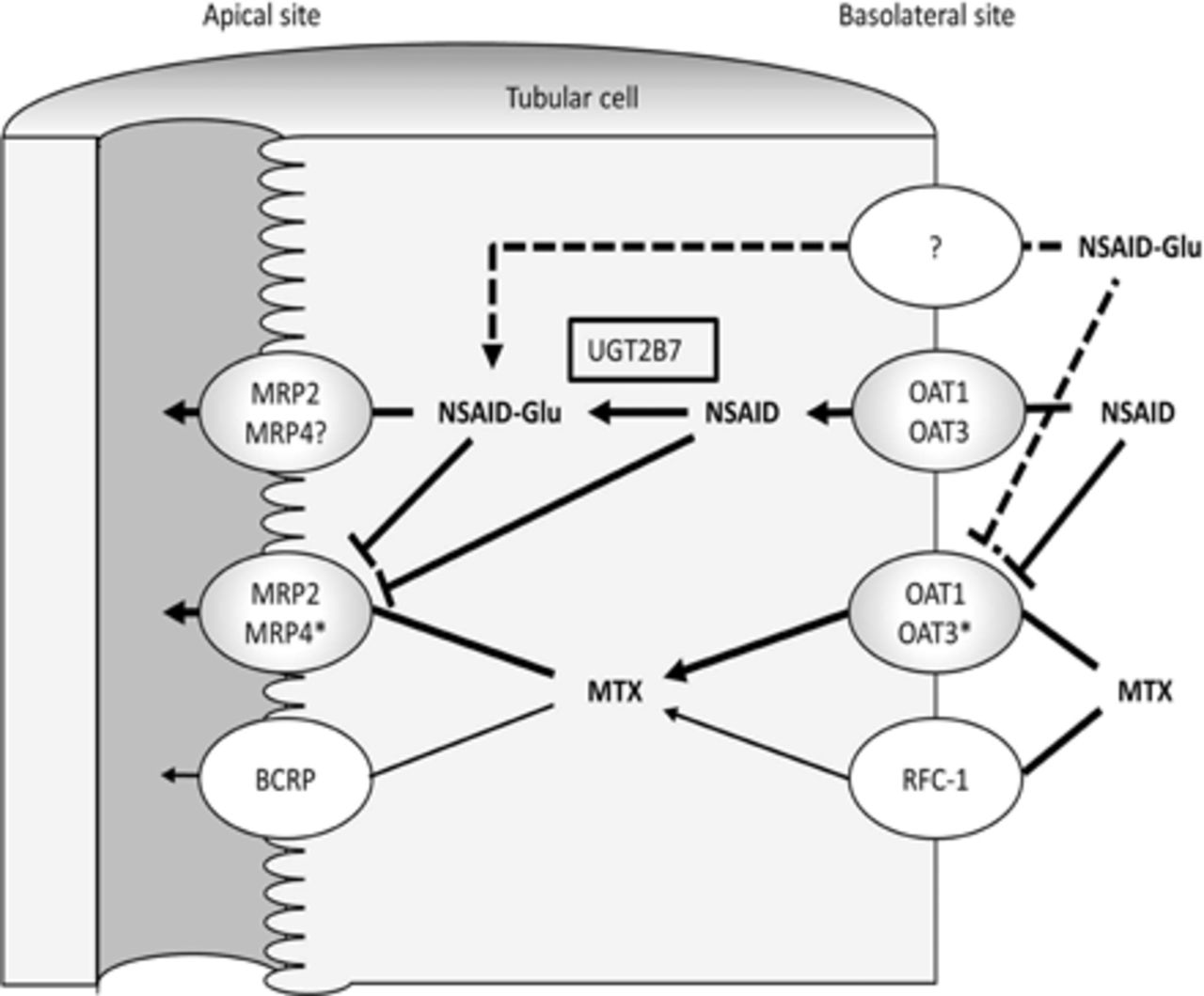
Combined administration of methotrexate (MTX) and nonsteroidal anti-inflammatory drugs (NSAIDs) can result in a decreased systemic clearance of MTX. To date, inhibition of renal uptake via organic anion transporters and efflux via multidrug resistance–associated protein (MRPs) by NSAIDs has been recognized as possible sites of drug interaction between MTX and NSAIDs. Although most NSAIDs are glucuronidated in kidney tissue and excreted mainly as glucuronide conjugates, it is not fully known whether the glucuronides of NSAIDs (NSAIDs-Glu) inhibit MTX excretion via MRP2 and MRP4. The purpose of this study was to investigate the inhibitory effects of the glucuronides of several NSAIDs (diclofenac, R- and S-ibuprofen, R- and S-flurbiprofen, and R- and S-naproxen), as well as the parent NSAIDs on MTX uptake using human MRP2- and MRP4-expressing inside-out vesicles. Results confirm that all NSAIDs and NSAIDs-Glu examined exhibited stereoselective and concentration-dependent inhibitory effects on MTX uptake via MRP2 and MRP4. Notably, NSAIDs-Glu potently inhibited MTX uptake via MRP2 and MRP4 compared with the corresponding parent NSAIDs except for naproxen in MRP2 and S-flurbiprofen in MRP4. The present results support that the glucuronides of NSAIDs, as well as the parent NSAIDs, are involved in the inhibition of urinary excretion of MTX via MRP2 and MRP4.
Footnotes
- Received September 3, 2015.
- Accepted December 7, 2015.
This work was supported by the “Antiaging” Project for Private Universities, with a matching fund subsidy from the Japanese Ministry of Education, Culture, Sports, Science and Technology (MEXT). This research was also supported in part by the MEXT-Supported Program for the Strategic Research Foundation at Private Universities, 2014–2018 (S1411037).
- Copyright © 2016 by The American Society for Pharmacology and Experimental Therapeutics
JPET articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|




{kind=link}