


Visual Overview

Abstract
Type 2 diabetes (T2D) is a rising pandemic worldwide. Diet and lifestyle changes are typically the first intervention for T2D. When this intervention fails, the biguanide metformin is the most common pharmaceutical therapy. Yet its full mechanisms of action remain unknown. In this work, we applied an ultrahigh resolution, mass spectrometry-based platform for untargeted plasma metabolomics to human plasma samples from a case-control observational study of nondiabetic and well-controlled T2D subjects, the latter treated conservatively with metformin or diet and lifestyle changes only. No statistically significant differences existed in baseline demographic parameters, glucose control, or clinical markers of cardiovascular disease risk between the two T2D groups, which we hypothesized would allow the identification of circulating metabolites independently associated with treatment modality. Over 3000 blank-reduced metabolic features were detected, with the majority of annotated features being lipids or lipid-like molecules. Altered abundance of multiple fatty acids and phospholipids were found in T2D subjects treated with diet and lifestyle changes as compared with nondiabetic subjects, changes that were often reversed by metformin. Our findings provide direct evidence that metformin monotherapy alters the human plasma lipidome independent of T2D disease control and support a potential cardioprotective effect of metformin worthy of future study.
SIGNIFICANCE STATEMENT This work provides important new information on the systemic effects of metformin in type 2 diabetic subjects. We observed significant changes in the plasma lipidome with metformin therapy, with metabolite classes previously associated with cardiovascular disease risk significantly reduced as compared to diet and lifestyle changes. While cardiovascular disease risk was not a primary outcome of our study, our results provide a jumping-off point for future work into the cardioprotective effects of metformin, even in well-controlled type 2 diabetes.
Introduction
Type 2 diabetes (T2D), often associated with obesity, is a rising pandemic worldwide, with 9.3% of adults estimated to have T2D globally: a number that is predicted to rise to 10.9% (700 million individuals) by 2045 (International Diabetes Federation, 2020). T2D comes at a high cost, both in terms of human life and economic damage due to loss of productivity and global health expenditures (Ng et al., 2014; International Diabetes Federation, 2017). In 2020, diabetes was the eighth leading cause of death in the United States (Ahmad and Anderson, 2021). Individuals with diabetes also suffer from a host of comorbidities whose severity increases the longer blood glucose remains inadequately controlled, including neuropathy, nephropathy, retinopathy, and microvascular and macrovascular complications (Nathan, 1993). Identifying ways to more effectively treat diabetes and its complications are of critical importance.
T2D is exemplified by chronically elevated blood glucose levels caused by both insufficient insulin release from the pancreatic β-cells and resistance of the body’s tissues to the effects of insulin. Hemoglobin A1c (HbA1c) is a clinical test that quantifies the glycemic state over the previous 2 to 3 months. For T2D individuals, current HbA1c targets for vary from 6.5% to 8%, depending on age and comorbidities (Qaseem et al., 2018; Garber et al., 2020; American Diabetes Association Professional Practice Committee, 2021). T2D disease management typically focuses first on diet and lifestyle changes, progressing to pharmaceutical interventions once those fail. Diet and lifestyle management focuses on five principles: improving diet, increasing exercise, reducing smoking, reducing alcohol consumption, and reducing body weight (Schlesinger et al., 2020). Yet, in a vast majority of individuals, diet and lifestyle modifications are insufficiently effective in achieving blood glucose targets, particularly long term (Khunti et al., 2013; American Diabetes Association Professional Practice Committee, 2021).
Once pharmaceutical intervention is called for, the biguanide metformin is nearly always the first-line therapeutic (American Diabetes Association Professional Practice Committee, 2021). Metformin is generally accepted as a medication with few severe complications and, other than initial gastrointestinal side effects, is usually well tolerated (American Diabetes Association Professional Practice Committee, 2021). Metformin has also been suggested to have cardioprotective benefits in individuals with T2D (Eurich et al., 2005; Evans et al., 2006; Holman et al., 2008). With increasing evidence that achieving HbA1c targets alone is insufficient to prevent adverse outcomes (Finfer et al., 2009), particularly with regards to cardiovascular events (Herman et al., 2005; Skyler et al., 2009; Giorgino et al., 2016), there has been a push for advancing pharmaceutical interventions in concert with diet and lifestyle changes (Altaf et al., 2015; Padhi et al., 2020). With nearly 70 years of clinical use and a well-established safety profile, metformin is a logical candidate (Holman et al., 2008).
Although there is still some debate in the field, metformin’s primary blood glucose-lowering mechanisms are thought to be reduced glucose production and release by the liver and increased lactate production in the small intestine (Pernicova and Korbonits, 2014; McCreight et al., 2016; Song, 2016). Both of these mechanisms have been linked with alterations in circulating metabolites (Wang-Sattler et al., 2012; Floegel et al., 2013; Walford et al., 2014; Guasch-Ferré et al., 2016; Lu et al., 2016; Pallares-Méndez et al., 2016; Yu et al., 2016; Lai et al., 2020; Zhao et al., 2020). Metabolomics, the endpoint of the ‘omics’ cascade, is well suited to study metabolic disorders like T2D (Dunn and Hankemeier, 2013; Guasch-Ferré et al., 2016; Pallares-Méndez et al., 2016). As high-throughput and high-sensitivity/specificity techniques are continually being developed, metabolomics has emerged as an approach well suited to provide coverage of the changes that occur in clinical biosamples because of disease status and intervention.
In this study, we applied a high-throughput, ultrahigh resolution flow injection electrospray (FIE) Fourier transform ion cyclotron resonance (FTICR) mass spectrometry (MS) workflow to plasma samples from three groups—nondiabetic (ND) subjects, T2D subjects treated with diet and lifestyle modifications, and T2D subjects on metformin monotherapy—quantifying the relevant abundance of metabolic features among groups using an untargeted approach. Notably, all T2D subjects had good to excellent glucose control, and subjects were well matched for potential confounders such as age and body mass index (BMI). More than 3000 discrete metabolic features were significantly expressed per group, with more than 2000 annotated by chemical name or structure. Family and pathway analyses revealed key changes in the lipidome, particularly with regards to fatty acids and related molecules, were strongly associated with metformin monotherapy, independent of commonly used measurements of T2D disease control and cardiovascular disease (CVD) risk.
Materials and Methods
Human Subjects
All human subjects research was conducted in accordance with the standards set out by the World Medical Association Declaration of Helsinki Ethical Principles for Medical Research Involving Human Subjects as approved by the University of Wisconsin Health Sciences Institutional Review Board protocol number UW 2013-1082. Study design and participant recruitment has been previously described (Truchan et al., 2021). Briefly, potentially eligible subjects were University of Wisconsin Hospitals and Clinics endocrinology or internal medicine patients who met baseline inclusion and exclusion criteria by consent-waived electronic medical record search. Briefly, inclusion criteria included age 18 to 74 years old, not pregnant or lactating, no anemia or grossly abnormal kidney or liver function tests, no known autoimmune diseases or inflammatory disorders, and, for T2D subjects, no diagnosis of diabetes besides T2D. Potentially eligible subjects were contacted by phone. Those interested in participating provided written informed consent, and a fasting plasma sample was collected. The full clinical cohort comprised 35 ND and 132 T2D subjects, as described previously (Truchan et al., 2021; Fenske et al., 2022). For this work, plasma samples from 15 T2D subjects treated conservatively with diet/lifestyle modifications or metformin monotherapy and 14 ND controls matched for age and BMI were selected for downstream untargeted metabolomics.
Metabolite Extraction
The extraction was performed as previously described (Zhu et al., 2021). In brief, a 30 µL aliquot was taken from thawed plasma samples, mixed with 60 µL of chilled liquid chromatography-MS grade methanol. The samples were then vortexed, mixed with a nutating mixer, and centrifuged. Fifty µL of supernatant was mixed with 50 µL of water for FIE-FTICR MS analysis.
FTICR MS Metabolomics Analysis
FIE-FTICR MS experiments were performed using a Waters nanoACQUITY UPLC (Waters Corporation, Milford, MA, USA) coupled to a Bruker solariX 12 T FTICR mass spectrometer (Bruker Daltonics, Bremen, Germany) without a liquid chromatography column. Five µL of each metabolite extract was directly injected in triplicate from the Waters nanoACQUITY UPLC into the FTICR MS via 100 µm × 40 cm polyetheretherketone tubing. The mobile phase was 50:50 methanol:water with 0.1% formic acid or 10 mM ammonium acetate added for positive or negative modes, respectively, with a flow rate of 20 µl/min. Ions were accumulated for 0.1 second and an 8 M transient size applied, with 50 scans collected. The mass/charge ratio (m/z) range was set to 40 to 1500, with 50 m/z Q1 mass. Fifty scans were collected for each mass spectrum. Dry gas flow was set to 4 L/min at 150°C. Largest frequency values for octopole (5 MHz), quadrupole (2 MHz), and transfer hexapole (6 MHz) were used to improve ion transition. Time of flight was set to 0.8 millisecond. Sweep excitation power was set to 27%. The estimated resolving power at 400 m/z was 190,000. The FTICR MS was calibrated with 1 mM sodium trifluoroacetate in both positive and negative modes before experiments.
Data Analysis
Mass spectra were processed and analyzed using Data Analysis 4.3 (Bruker Daltonics, Bremen, Germany). Bucket (mass) lists in positive and negative ion modes were generated using the T-ReX 2D workflow in MetaboScape 4.0. The maximum Δm/z was set to 0.50 mDa. The maximum charge state was set to 3, and the intensity threshold was set to 0. The minimum number of features for the result was set to 5. Bucket lists in positive and negative ion modes were merged into one bucket list with 1.0 ppm m/z tolerance. Features with a ratio of sample average:blank average < 10 were deleted. The merged bucket list was annotated with the SmartFormula function in MetaboScape 4.0 (Bruker Daltonics, Bremen, Germany) with 2.0 ppm as the narrow Δm/z cutoff, 5.0 ppm as the wide Δm/z cutoff, 20 as the narrow mSigma cutoff, and 50 as the wide mSigma cutoff. Elements were set to CHNOPS, and element ratio filters were applied with common element ratio presets in MetaboScape 4.0, including 0.2 to 3.1 H/C ratio, 0 to 1.3 N/C ratio, 0 to 1.2 O/C ratio, 0 to 0.3 P/C ratio, 0 to 0.34 P/O ratio, and 0 to 0.8 S/C ratio. Electron configuration was set to even. Heuristic element count probability check was applied. The putative metabolites were annotated by METLIN with a 2 ppm mass error cutoff. In most cases, the isomer with the lowest METLIN number was reported. Lipids and lipid-like molecules were reported as a combined general chemical name with total fatty acid carbon number:number of double bonds. In a few cases, more than one isomer with similar structures and chemical properties were reported for one mass. SmartFormula and METLIN annotations provide information on the chemical formulas and chemical names, respectively (Smith et al., 2005; Guijas et al., 2018). InChI keys, HMDB IDs, and KEGG IDs were acquired using CTS servers from the METLIN name. InChI keys were used to classify the METLIN annotations using “ClassyFire,” and annotated metabolites were categorized according to their class information (Djoumbou Feunang et al., 2016). PubChem IDs were gathered using the PubChem server translator. Statistical analysis was performed using MetaboScape 4.0 and the online software MetaboAnalyst (Chong et al., 2018). MetaboScape was used for principal component analysis. MetaboAnalyst was used for pathway analysis, enrichment analysis, and statistical analyses (e.g., heat map, significant features). All P values are reported as false discovery rate-corrected. ChemRich plots were made using the database information from CTS, PubChem IDs, and P values from MetaboAnalyst to generate a plot of chemical similarity and significance.
Results
Validation of the Patient Cohort to Detect Treatment-Specific Changes in the Metabolome
Biobanked plasma samples from seven T2D individuals treated with diet and lifestyle modifications (T2D-DL), eight T2D individuals treated with metformin monotherapy (T2D-M), and 14 ND subjects, well matched for age and BMI, were selected from a larger clinical cohort described in Truchan and colleagues and Fenske and colleagues (Truchan et al., 2021; Fenske et al., 2022). The mean duration of disease was nearly identical between T2D-DL and T2D-M groups (∼3 years). As expected, HbA1c and fasting blood glucose were both elevated in T2D subjects as compared with ND, with no statistically significant differences between the T2D groups, and no T2D individuals had HbA1c levels above 8.1%, indicating good to excellent glucose control (Table 1). Fasting glucose and fasting insulin were used to calculate insulin resistance using the Homeostatic Model Assessment for Insulin Resistance (HOMA-IR) (Matthews et al., 1985). Neither fasting insulin nor HOMA-IR were significantly different among the groups (Table 1). Common CVD risk factors, including systolic blood pressure, diastolic blood pressure, total cholesterol, high-density lipoprotein, low-density lipoprotein, and triglycerides levels, were also similar (Table 1). As these values can be influenced by supplements or pharmaceuticals commonly prescribed to T2D patients, the rates of daily omega-3/fish oil supplement, statin, and prophylactic (i.e., low-dose) aspirin use were recorded, and none were significantly different among the T2D groups, although more T2D subjects were using prophylactic aspirin than ND (Table 1). Significantly more T2D subjects had been prescribed an angiotensin-converting enzyme inhibitor, angiotensin receptor blocker, or beta-blocker for high blood pressure than ND subjects, with no difference between the T2D groups (Table 1). Lipoprotein-associated phospholipase A2 (Lp-PLA2) is a marker of arterial inflammation associated with CVD risk (Dada et al., 2002), and there was a trend (P = 0.06) toward elevated Lp-PLA2 in the T2D groups as compared with ND, with no difference between the T2D groups (Table 1). No subjects had clinically elevated erythrocyte sedimentation rate or C-reactive protein, indicating the absence of acute inflammation or infection(Table 1). Taken together, both T2D groups had nearly identical diabetes control and CVD risk as measured by well-accepted clinical markers.
Demographic and clinical parameters of patient cohort
Unless otherwise indicated, data are presented as average ± standard deviation.
“Lipids and Lipid-like Molecules” Are the Largest Superclass of Plasma Metabolites Detected by an Untargeted FIE-FTICR MS Approach
We employed a FIE-FTICR MS workflow previously developed and validated for high-throughput, ultrahigh resolution untargeted metabolomics analysis of preclinical plasma samples (Schaid et al., 2021; Zhu et al., 2021). Using MetaboScape, more than 3840 discrete metabolic features, as defined by unique m/z, were detected above background, with most being shared among groups (Fig. 1A, hatched bars). Of these features, 2925 could be annotated by chemical formula, again, with most (1956) being shared among groups (Fig. 1A, solid bars, and Fig. 1B). Two hundred fifty-one were limited to the ND group, 11 limited to the T2D-DL group, and 10 limited to the T2D-M group (Fig. 1B). Performing a principal component analysis, samples from the T2D-DL group clustered the most strongly together, with significant overlap between the ND and T2D-M groups, with only the ND group having samples outside of the 95% confidence limit (Fig. 1C). A schematic of the analysis workflow can be found in Supplemental Fig. 1, and a full list of significantly expressed metabolic features with can be found in Supplemental File 1.

Overview of metabolic features, distribution, and classes among groups. (A) Bar chart displaying the total number of accurate mass features (hatched bars) and features annotated by chemical formula in MetaboScape (solid bars). (B) Venn diagram showing the number of shared and unique MetaboScape-annotated features among the three groups. (C) Principal component analysis plot of MetaboScape-annotated features among the three groups. (D) Chemical classification of METLIN-annotated metabolic features separated into nonpolar (yellow) and polar (blue) metabolites. (E) Heat map of MetaboScape-annotated features generated by unsupervised hierarchical clustering.
Based on the chemical ontology of annotated metabolites, the “lipids and lipid-like molecules” superclass comprised over 67% of annotated metabolites, with the combined “glycerophospholipids” and “fatty acyls” classes accounting for two-thirds (Fig. 1D). Of the polar metabolites, “organic acids and derivatives” was the second most abundant superclass, with 48 annotations, and was primarily composed of the subclass “amino acids, peptides, and analogues” (Fig. 1D). Unsupervised hierarchical clustering of the MetaboScape-annotated features revealed ND samples strongly segregated from the T2D groups, with some overlap between the two T2D groups (Fig. 1E).
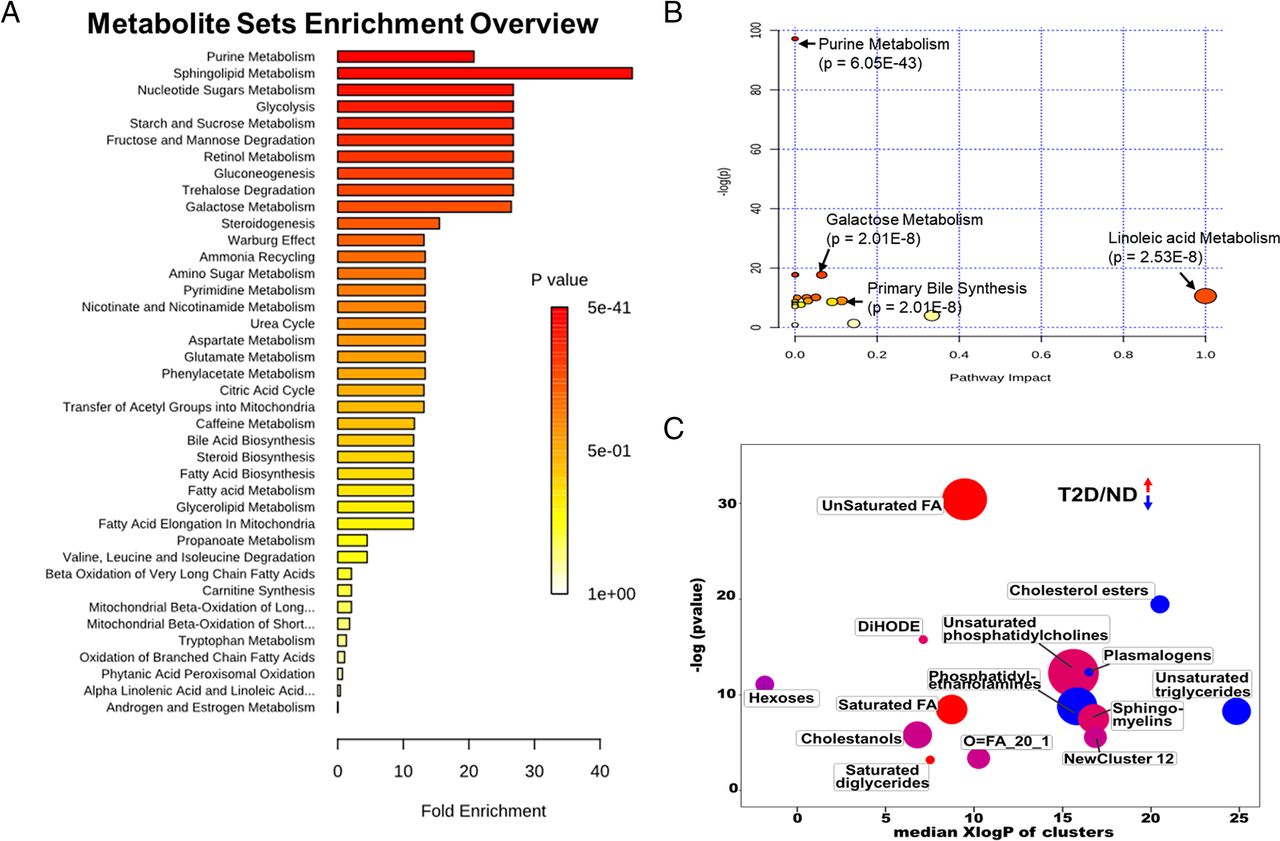
A Broad Shift to Steroidogenesis T2D groups
A wide range of metabolite sets were enriched in the combined T2D groups versus ND group. The metabolite set with the highest fold enrichment was “sphingolipid metabolism,” while “purine metabolism” had the highest statistical significance (Fig. 2A): both components in steroidogenesis, which was also significantly enriched. In agreement with the metabolite set analysis, the purine metabolism pathway was the most significantly altered, while changes to the linoleic acid metabolism pathway were the most highly affected by T2D status (Fig. 2B). A ChemRich analysis, which clusters based on chemical similarity, highlights the broad differences between the combined T2D group and ND (Fig. 2C). Among these differences were increases in fatty acid abundance, including both saturated and unsaturated fatty acids. Important to note is the observed increase in hexoses, as would be expected in plasma samples from T2D subjects. There were also changes to multiple lipid classes, including phosphatidylethanolamines (PEs), phosphatidylcholines (PCs), and sphingomyelins (SMs), with most of them being increased (Fig. 2C).

Comparison of enriched metabolic pathways for the ND group versus the combined T2D groups. (A) Pathway enrichment analysis showing pathways that are significantly altered at P = 0.05 level. (B) Pathway analysis with significantly altered metabolite groups highlighted. (C) ChemRich analysis using chemical similarity to show the trend in features that are modified. In C, blue is downregulated compared with control, red is upregulated, and purple is mixed.
Separate pairwise analyses were also performed among the ND, T2D-DL, and T2D-M groups. Similarly to the combined T2D analysis, “sphingolipid metabolism” was the most highly enriched and most significantly impacted metabolite set in the T2D-DL versus ND group (Supplemental Fig. 2A), purine metabolism was the most significantly altered pathway (Supplemental Fig. 2B). The T2D-DL group showed a broad shift in metabolism, with numerous fatty acid classes increased, including saturated, unsaturated, and hydroxy fatty acids (Supplemental Fig. 2C). PCs, PSs, and amino acids were increased, while cholesterol esters and unsaturated triglycerides were decreased (Supplemental Fig. 2C). Similar results for metabolite set and pathway impact analyses when comparing the T2D-M versus ND groups (Supplemental Figs. 3A, S3B). Interestingly, the ChemRich analysis showed far fewer differences in chemical similarity between the T2D-M and ND groups as compared with either the combined T2D or T2D-DL analyses (Supplemental Fig. 3C). As compared with the ND group, the T2D-M group had SM and PC lipid species that were increased, while others were decreased, and unsaturated triglycerides and cholesterol ethers were also changed as compared with the ND group (Supplemental Fig. 3C). Complete lists of significantly altered metabolites from each analysis can be found in Supplemental Tables 1 and 2.
Changes in Fatty Acid Metabolism by T2D Treatment
Based on the significant differences observed in the abundance of fatty acids, their conjugates, and related metabolite groups such as phospholipids (Table 2 and Supplemental Tables 1 and 2), we honed in on a subset of these metabolites for a more detailed analysis. Even though there were no significant differences between the two T2D groups with regards to T2D control and other important biometric and clinical parameters, there was a clear alteration in fatty acid metabolism in the T2D-DL group as compared with the ND group, with many of these changes being ameliorated with metformin treatment. The longer chain acylcarnitines octanoylcarnitine and palmitoylcarnitine were increased in the T2D-DL group, with no difference between the ND and the T2D-M groups (Fig. 3A). The building block carnitine was lowest in the T2D-M group, albeit not with statistical significance. Higher levels of palmitoyl carnitine have been linked to disorders of fatty acid oxidation (Bjørndal et al., 2018), and an analysis of fatty acid abundance among groups also showed increased levels in the T2D-DL group, with a return to ND levels in the T2D-M group (Fig. 3B). This trend held true for both saturated fatty acids, including palmitic and stearic acid, as well as unsaturated fatty acids, including oleic, linoleic, and arachidonic acid. Saturated hydroxy fatty acids were not altered among the groups, including the precursor to fatty acid esters of hydroxy fatty acids (FAHFAs), 2-hydroxy stearic acid (Fig. 3C). Unsaturated hydroxy fatty acids did not follow this trend, with ricinoleic acid being increased in both T2D groups as compared with ND (albeit with statistical significance only for T2D-DL), while the abundance of dimorphecolic acid and α-kamlolenic acid decreased (Fig. 3C). Finally, FAHFA abundance was increased across the board in the T2D-DL group as compared with ND, with a normalization in plasma from individuals treated with metformin (Fig. 3D).
Significantly altered metabolites between T2D-DL versus T2D-M

Fatty acid metabolism shifts in response to T2D and metformin treatment. (A) Acylcarnitines are upregulated in T2D-DL group and return to baseline in the T2D-M group. (B) Fatty acids, both saturated (left) and unsaturated (right), are increased in T2D-DL and similar between ND and T2D-M. (C) Hydroxy fatty acids are similar between the groups with saturated (left) showing no difference and unsaturated (right) showing changes in the T2D groups compared with ND. (D) FAHFAs are all upregulated in T2D-DL and significantly decrease in the T2D-M group. *P value < 0.05, **P value < 0.01, ***P value < 0.001.
Changes in Phospholipid Abundance with Metformin Therapy
A volcano plot analysis revealed several metabolic features had a statistically significant fold change of > 2 between the two groups (Fig. 4A). In exploring the metabolites that were altered, we observed decreases in unsaturated fatty acids and phospholipids composed of these fatty acids, including PCs, PEs, and SMs, in the T2D-M group compared with T2D-DL group (Fig. 4B). In the proinflammatory state, Lp-PLA2 cleaves oxidized fatty acids from the sn2 position of phospholipids (Rosenson and Stafforini, 2012), and increased levels of Lp-PLA2 are associated with increased risk of CVD (Garza et al., 2007). While no differences in Lp-PLA2 levels were observed between the two T2D groups, the levels of oxidized PCs were significantly increased levels in the T2D-DL group as compared with ND and were normalized by metformin treatment (Fig. 4C). The trend of increased lipid abundance in the T2D-DL group was particularly strong for lipids with higher peak intensity, including SM (d34:1) and PC (38:6) (Fig. 4D). For lower intensity lipids, such as SM(d36:2) and PC(36:5), we also observed the same trend, indicating a global shift independent of any effect of lipid species abundance (Fig. 4E).

Metformin impacts lipid metabolism in T2D. (A) Volcano plot of T2D-DT2D-M/T2D-DL with 11 features increased in the T2D-M group and 39 decreased at cutoff of twofold change and P value<0.05. (B) ChemRich plot highlighting lipophilic compounds (Xlog P) values are greater than 7. (C) Selected oxidized lipids. (D) Selected highly abundant lipids. (E) Lower abundant lipids. *P value < 0.05, **P value < 0.01, ***P value < 0.001.
Discussion
Summary of Results
In this work, we compared the abundance of plasma metabolites in patients with well-controlled T2D treated conservatively with diet and lifestyle interventions or metformin monotherapy, thereby excluding any global effects of uncontrolled T2D on our results. Within the T2D groups, subjects had similar BMI, HOMA-IR, and fasting insulin levels, thereby excluding global effects of obesity and insulin resistance. Significant changes in a number of lipids and lipid-like molecules, including carnitines, free fatty acids (FFAs), hydroxy fatty acids, FAHFAs, and phospholipids, were found, with patients in the T2D-M group often having levels similar to ND controls.
Metabolites Associated with CVD Risk Are Reduced with Metformin Monotherapy
With recent studies debating the therapeutic benefits of intensive glycemic control in T2D (Rodriguez-Gutierrez et al., 2019), it is important to understand what mechanisms outside of glucose control may contribute to the benefits of T2D therapeutics. Previous studies have shown changes the levels of branched chain amino acids, phospholipids, fatty acids, triglycerides, acylcarnitines, and small molecular weight compounds in individuals with T2D (Wang-Sattler et al., 2012; Floegel et al., 2013; Walford et al., 2014; Guasch-Ferré et al., 2016; Lu et al., 2016; Pallares-Méndez et al., 2016; Yu et al., 2016; Lai et al., 2020; Zhao et al., 2020; Truchan et al., 2021). These shifts can occur prior to the development of hyperglycemia, highlighting the global changes of the disease (Tabák et al., 2009). Metformin has multiple known potential modes of action, including the inhibition of the mitochondrial enzymes, glycerol-3-phosphate dehydrogenase, and complex 1 of the electron transport chain (Minamii et al., 2018). Inhibiting these enzymes results in a reduction of substrate utilization and manifests systemically as reducing hepatic gluconeogenesis (Minamii et al., 2018).
The first lipid class we investigated was acylcarnitines, which function in transporting fatty acids into the mitochondria. The T2D-DL group had the highest levels of acylcarnitines, with subjects in the T2D-M group having similar levels to those in the ND group. Acylcarnitines have previously been reported as a proxy for fatty acid metabolism, with increased levels in circulation indicating β-oxidation dysfunction (Strand et al., 2017; Flam et al., 2022). This result is consistent with previous studies indicating that individuals with T2D have increased levels of short-, medium-, and long-chain acylcarnitines (Sun et al., 2016; Strand et al., 2017), which are reflective of cardiac metabolism (Makrecka-Kuka et al., 2017). However, our results differ from these previous studies, as the T2D-M group had similar levels of acylcarnitines to the ND group. Some hypothesize that medium-chain acylcarnitines are increased early in T2D pathogenesis, with an increase in long-chain at later time points as fatty acid oxidation is further disrupted (Schooneman et al., 2013). As all of our subjects had well-controlled and conservatively treated T2D, though, changes in acylcarnitines were specific to metformin treatment. Circulating acylcarnitines have been identified as correlating with CVD morbidity and mortality (Hosseinkhani et al., 2022; Paulin Beske et al., 2022; Storesund et al., 2022). Our results suggest metformin has beneficial effects on these outcomes independent of T2D disease status.
In the T2D state, a change in the abundance of fuel substrates exists, with a shift toward greater utilization of FFAs (Herman et al., 2005). FFAs and their metabolites are known to play a role in inhibiting insulin secretion and signaling, making them a crucial link between metabolic disorder and disease manifestation (Boden and Shulman, 2002; Bosma et al., 2022). Recently, higher circulating FFAs have also been linked with elevated risk of cardiovascular events, independent of T2D status and glycemic control (Yu et al., 2021; Hu et al., 2022; Lluesa et al., 2022; Thirumathyam et al., 2022). In the context of these previous studies, a reduction in circulating FFAs with metformin further supports beneficial effects of metformin even in well-controlled T2D patients.
FAHFAs were first reported to correlate with insulin sensitivity, with evidence they are reduced in insulin-resistant individuals (Yore et al., 2014). Yet circulating FAHFA levels were also found to correlate with several markers of cardiovascular function in healthy human subjects (Dongoran et al., 2020). In our study, individuals in the T2D-DL group had higher mean FAHFA abundance than those treated with metformin. As HOMA-IR was not statistically different between the T2D-DL and T2D-M groups, our data support an association of FAHFA levels with CVD risk independent of T2D status. Yet few of our subjects had clinically significant HOMA-IR values, and, in Yore and colleagues, the change in circulating palmitic acid esthers of hydroxystearic acid (PAHSA) levels occurred at a later time point in T2D disease progression than our cohort (Yore et al., 2014). Additionally, only some PAHSA isomers have been shown to elicit an anti-inflammatory response in mice. In this work, we measured total PAHSAs and not isomer-specific, which requires a targeted method (Moraes-Vieira et al., 2016; Kolar et al., 2018). An additional factor to consider is PAHSA has also been shown to increase with exercise (Brezinova et al., 2020), and we do not have exercise data for our clinical cohort. Taken together, these differences could explain some of the discrepancies in our data compared with previous reports and caveats of our study that will require additional investigation to deconvolute.
In our study, phospholipid abundance decreased with metformin treatment, indicating a change in nutritional overload. The acyl chains of phospholipids with higher degrees of unsaturation have previously reported to increase in diabetic patients by 45% to 64% compared with ND patients (Chuang et al., 2012). This is consistent with our findings of increased phospholipid levels in the T2D-DL group. However, we also observed these phospholipids decreased in the T2D-M group, indicating the potential that phospholipids are sensitive to overall energy state. Further, in T2D-M subjects, phospholipid levels were more similar to the ND group than the T2D-DL groups, demonstrating the effects of metformin on energy balance.
Oxidized phospholipids have previously been shown to increase in insulin-resistant individuals and be associated with the pathogenesis of oxidative stress-related diseases, including CVD (Fruhwirth et al., 2007; Sun et al., 2016; Que et al., 2018). Yet, in previous work, increased circulating oxidized phospholipids correlated with Lp-PLA2 levels (Pantazi et al., 2022). In our study, we found oxidized phospholipids were specifically elevated in the T2D-DL group, even though there was no statistically significant difference between the number of subjects in the T2D-DL and T2D-M groups who had clinically elevated Lp-PLA2 levels. Taken together with the increased levels of hydroxy fatty acids, this finding indicates metformin improves the oxidized lipid profile independent of this common clinical marker of CVD risk.
Hydroxy fatty acids have been linked to a variety of diseases, including cancer, inflammatory bowel disease, and neurodegenerative diseases (Li et al., 2020). Additionally, a link has been demonstrated between insulin-resistant states and increased vascular risk via hydroxy fatty acids and increased secretion of plasminogen activator inhibitor type 1 (Marx et al., 1999; Vangaveti et al., 2010). Our data shows the two T2D groups displayed similar trends in hydroxy fatty acids as compared with the ND group. As hydroxy fatty acids can be a proxy for reactive oxygen species (Wang et al., 2009), this indicates a potential underlying mechanism of T2D that is not controlled by metformin treatment. Yet, in our study, we found reduced levels of oxidized fatty acids, suggesting further study with a more comprehensive, targeted panel of hydroxy fatty acids is warranted.
Improved Pancreatic β-Cell Function May Occur with Metformin Therapy
Prostaglandin E2 (PGE2), an arachidonic acid metabolite, is elevated in pancreatic islets from T2D mice and human organ donors, actively contributing to the β-cell dysfunction of the disease (Kimple et al., 2013; Neuman et al., 2017; Schaid et al., 2021; Zhu et al., 2021; Bosma et al., 2022). Circulating PGE2 is rapidly degraded, and a targeted lipidomics approach is required to detect arachidonic acid metabolites. Yet, in previous work, the abundance of arachidonic acid in membrane phospholipids was significantly elevated in pancreatic islets from T2D mice as compared with ND, correlating directly with the concentration of PGE2 produced (Kimple et al., 2013; Neuman et al., 2017), and incubating islets from T2D mice with eicosapentaenoic acid, which competes with arachidonic acid for the same site in membrane phospholipids, significantly improved T2D β-cell dysfunction (Neuman et al., 2017). In this work, we found elevated levels of arachidonic acid and its precursor, linoleic acid, in T2D subjects treated with diet and lifestyle modifications as compared with ND controls, with metformin therapy reversing these changes. While we did not directly measure β-cell function in our study, others have found metformin augments insulin secretion, even in the context of reduced insulin demand (Vazquez Arreola et al., 2022). Therefore, it is possible decreased β-cell PGE2 is at least partially responsible for metformin’s effects.
Limitations and Conclusions
Our results demonstrate in a small population of well-controlled T2D patients, metformin treatment significantly improved the circulating profiles of a number of lipids and lipid-like molecules, some of which have been independently correlated with CVD risk. Yet, as these metabolites are not clinically validated, we are unable to unequivocally conclude metformin reduces CVD risk independent of T2D control. Another limitation of our study is its design did not include longitudinal sample collection or long-term follow-up, a design required to confirm a reduced CVD risk with metformin therapy. Even so, our findings suggest an untargeted plasma metabolomics approach may provide a much richer set of biomarkers to quantify CVD risk. Future studies with larger populations and targeted approaches will be necessary to validate and advance our results into improved clinical care.
Acknowledgments
The authors would like to thank Stephanie Blaha for her assistance with patient recruitment.
Authorship Contributions
Participated in research design: Brasier, Cox, Davis, Ge, Kimple.
Conducted experiments: Wancewicz, Zhu, Fenske, Weeks, Wenger, Pabich, Daniels, Punt, Peter, Nall.
Contributed new reagents or analytic tools: Wancewicz, Zhu, Ge.
Performed data analysis: Wancewicz, Zhu, Fenske, Weeks, Kimple.
Wrote or contributed to the writing of the manuscript: Wancewicz, Zhu, Brasier, Cox, Davis, Ge, Kimple.
Footnotes
- Received October 24, 2022.
- Accepted January 17, 2023.
This work was funded in part by the U.S. Department of Veterans Affairs Biomedical Laboratory Research and Development Service [I01 BX003700]; National Institutes of Health National Center for Advancing Translational Sciences [Grant UL1-TR002373], National Institute of Diabetes and Digestive and Kidney Diseases [Grants R01-DK102598, F31-DK109698], National Institute of General Medical Sciences [Grants R01-GM125085, T32-GM081061], National Heart, Lung, and Blood Institute [Grants R01-HL109810, F31-HL152647], and NIH Office of the Director [Grant S10-OD018475]; a UW2020 WARF Discovery Initiative Grant from the University of Wisconsin-Madison Office of the Vice Chancellor for Research and Graduate Education and the Wisconsin Alumni Research Foundation; and Research Starter Grant in Translational Medicine and Therapeutics from the PhRMA Foundation. A.W. was supported by a VA Advanced Fellowship in Women’s Health. S.P. was supported by a Pearl Stetler Research Fund for Women Physicians Fellowship.
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health, the U.S. Department of Veterans Affairs, or the U.S. government. The study sponsors had no role in the study design; collection, analysis, or interpretation of data; the writing of the report; or the decision to submit the paper for publication. No author has an actual or perceived conflict of interest with the contents of this article.
↵
 This article has supplemental material available at dmd.aspetjournals.org.
This article has supplemental material available at dmd.aspetjournals.org.

Abbreviations
- BMI
- body mass index
- CVD
- cardiovascular disease
- FAHFAs
- fatty acid esters of hydroxy fatty acids
- FFA
- free fatty acid
- FIE
- flow injection electrospray
- FTICR
- Fourier transform ion cyclotron resonance
- HbA1c
- hemoglobin A1c
- HOMA-IR
- Homeostatic Model Assessment for Insulin Resistance
- Lp-PLA2
- lipoprotein-associated phospholipase A2
- MS
- mass spectrometry
- m/z
- mass/charge ratio
- ND
- nondiabetic
- PAHSA
- palmitic acid esthers of hydroxystearic acid
- PC
- phosphatidyl choline
- PGE2
- prostaglandin E2
- PE
- phosphatidyl ethanolamine
- SM
- sphingomyelin
- T2D
- type 2 diabetes
- T2D-DL
- type 2 diabetic treated with diet and lifestyle modifications
- T2D-M
- type 2 diabetic treated with metformin monotherapy
- U.S. Government work not protected by U.S. copyright.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}