


Abstract
Kinin B1 receptors are known to be highly induced after inflammatory stimuli in several biological systems. We report that incubation of pig iris sphincter with lipopolysaccharide from Escherichia coli caused a marked and time-related up-regulation of B1, accompanied by a reduction of B2 receptor-mediated contractile responses. The up-regulation of B1 receptors by lipopolysaccharide stimulation was decreased by the inhibitors of protein synthesis, cycloheximide and actinomycin D, and by dexamethasone and the nuclear factor-κB (NF-κB) inhibitor pyrrolidinedithiocarbamate (PDTC). In addition, lipopolysaccharide-induced up-regulation of B1 receptors in the pig iris sphincter was significantly reduced by the p38 inhibitor 4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl)1H-imidazole (SB203580) and to a lesser extent by the extracellular signal-regulated kinases 1 and 2 (ERK1/2) blocker 2′-amino-3′-methoxyflavone (PD98059). Molecular biology experiments demonstrated that in vitro incubation with lipopolysaccharide resulted in a time-dependent and remarkable activation of NF-κB and of p38 and ERK1/2 mitogen-activated protein (MAP) kinases, in pig iris sphincter preparations. While attempting to verify how MAP kinases are part of the B1 receptor-activated signaling transduction pathways, we observed that PD98059 was able to markedly reduce the contraction induced by B1 receptor activation in lipopolysaccharide-pretreated pig iris sphincter muscle but that this response was only partially decreased by SB203580. Our results extend the previous evidence on the mechanisms underlying the B1 receptor upregulation processes and demonstrate for the first time how this takes place in an ocular tissue, the pig iris sphincter. It is therefore possible to define B1 receptors as therapeutic targets for the treatment of infectious and inflammatory alterations of the eye.
Kinins represent a group of biologically active peptides generated at the sites of tissue damage, either in response to stimuli such as trauma or infection or during most inflammatory processes (Moreau et al., 2005). The role of kinins has been described in numerous systems, and their main pharmacological effects consist of smooth muscle contraction and relaxation, vasodilatation, increase of vascular permeability, and sensitization of sensorial fibers (Regoli and Barabé, 1980; Leeb-Lundberg et al., 2005). Kinins mediate most of their effects through the stimulation of two different G-protein-coupled receptors, classified as B1 and B2 (Regoli and Barabé, 1980; Calixto et al., 2000, 2001; Leeb-Lundberg et al., 2005). These receptors were initially defined based on pharmacological criteria and subsequently by molecular cloning studies (Regoli and Barabé, 1978; Hess et al., 2002). The B2 receptor is constitutively expressed in both the peripheral and central nervous systems, mediating most of the physiological actions evoked by kinins and exhibiting high affinity for bradykinin and kallidin. In contrast, the B1 receptor shows higher affinity for the kinin metabolites des-Arg9-bradykinin and des-Arg10-kallidin, and is generally absent under normal conditions, although it may be strongly upregulated after tissue trauma, during certain inflammatory states, or by the action of proinflammatory cytokines or bacterial products (e.g., Escherichia coli endotoxins) (Regoli and Barabé, 1980; Calixto et al., 2000, 2001; Leeb-Lundberg et al., 2005).
Lipopolysaccharide is the major component of the outer membrane of Gram-negative bacteria. It is now well known that administration of lipopolysaccharide elicits several relevant effects, such as endotoxic shock, inflammation, endotoxin tolerance, and Th1-type immune responses through massive production of proinflammatory cytokines and chemokines (Beutler and Rietschel, 2003). The majority of lipopolysaccharide's effects are mediated by its interaction with a member of the toll-like receptor family named TLR-4, and its responses are linked to the activation of several intracellular transduction pathways. Next, lipopolysaccharide can induce the association of the adapter MyD88 with TLR4, followed by the recruitment and activation of IL-1R-associated kinase 1 (Miyake, 2004). This leads to the subsequent activation of two different pathways involving the mitogen-activated protein (MAP) kinase family members ERK1/2, p38, and c-Jun NH2-terminal kinase and the Rel family transcription factor NF-κB (Hwang et al., 1997; Lawrence et al., 2001; Beutler, 2002; Miyake, 2004). Of high interest, these signaling pathways have been postulated as being involved in the up-regulation of B1 receptors (Larrivée et al., 1998; Ni et al., 1998; Schanstra et al., 1998; Campos et al., 1999; Sabourin et al., 2002; Medeiros et al., 2001, 2004). Furthermore, many pieces of evidence now indicate that lipopolysaccharide can lead to the overexpression of B1 receptors when assessed in both in vitro and in vivo models, as verified in various animal species such as rats, mice, rabbits, pigs, and nonhuman primates (Marceau et al., 1998; Calixto et al., 2004; Leeb-Lundberg et al., 2005). However, the precise sequence of signaling events bridging lipopolysaccharide to the B1R up-regulation, especially in smooth muscle, is not yet fully understood.
In the present study, we investigated whether the treatment with lipopolysaccharide is able to promote the up-regulation of B1R in the pig iris sphincter by use of an in vitro system. In addition, we examined the requirement of NF-κB and MAP kinase pathways for B1 receptor induction. Finally, we also evaluated the temporal profile for the activation of these molecules before B1 receptor up-regulation.
Materials and Methods
Drugs and Regents. The following drugs were used: des-Arg9-bradykinin, des-Arg9[Leu8]-bradykinin, bradykinin, dexamethasone, pyrrolidinedithiocarbamate (PDTC), bacterial lipopolysaccharide (from E. coli, serotype 0111:B4), actinomycin D, cycloheximide, captopril, carbachol (all from Sigma Chemical, St. Louis, MO), PD98059, and SB203580 (both from Tocris Cookson, Ellisville, MO). d-Arg0-[Hyp3,Thi5,d-Tic7,Oic8]-bradykinin (Hoe 140) was from Aventis Pharma Deutschland GmbH (Frankfurt Main, Germany). R-715 and the benzodiazepine antagonist were kindly donated by Dr. Férnand Gobeil (Department of Pharmacology, University of Sherbrooke, Sherbrooke, QC, Canada) and by Merck Research Laboratories (West Point, PA), respectively. The stock solutions for all peptides used were prepared in phosphate-buffered saline (1-10 mM) and kept in siliconized plastic tubes at -18°C until use. All other drugs were dissolved in phosphate-buffered saline to the desired concentration just before use. In all experimental groups, at least one parallel control experiment was carried out in the presence of the vehicle used to dilute the drugs.
Preparation of Pig Iris Sphincter for Organ Bath Studies. The methodology used was the same as that described previously by El Sayah and Calixto (2003a,b). Pig eyes were obtained from a local slaughterhouse and were immediately transferred on ice to the laboratory. Tissues were used within 6 h after animal death. The pig iris sphincters were isolated and set up in organ chambers. Preparations were allowed to equilibrate for at least 90 min before drug addition. During this period, the Krebs-Henseleit solution was changed every 15 min. After a 90-min period of stabilization, preparations were exposed to a single concentration of KCl (80 mM) as a standard stimulus. After washout, replacement with normal medium, and return to the original baseline, complete concentration-response curves (CRCs) for des-Arg9-bradykinin or bradykinin (1-3000 nM) were obtained (at 60-min intervals) by means of the cumulative method. Contractile responses are expressed as the percentage of maximal KCl (80 mM)-induced contraction.
Effect ofE. coliEndotoxin. To analyze the effect of in vitro exposure to E. coli endotoxin (lipopolysaccharide) on kinin-induced contraction of the pig iris sphincter, CRCs were constructed for des-Arg9-bradykinin or bradykinin at 2, 4, 6, 12, and 24 h after continuous incubation of the preparations with lipopolysaccharide (1 μg/ml). Different concentrations of lipopolysaccharide were tested, and 1 μg/ml was selected as the best on account of its ability to up-regulate B1 receptors. Krebs-Henseleit solution containing lipopolysaccharide was renewed every 15 min. The specificity of lipopolysaccharide to kinin-mediated responses was determined by comparing its effects on the contractile responses elicited by carbachol in the pig iris sphincter. Only one agonist was tested in each preparation.
Effect of Kinin Receptor Antagonists. To confirm the involvement of kinin receptors in the contractile response of pig iris sphincter to des-Arg9-bradykinin after lipopolysaccharide incubation, CRCs were obtained in the absence or presence of selective kinin B1 and B2 receptor antagonists. Preparations were exposed continuously to lipopolysaccharide for 4 h, and then they were incubated with the selective B2 receptor antagonist Hoe 140 (30 nM) or with the selective peptide B1 receptor antagonists des-Arg9[Leu8]-bradykinin (3-30 μM) and R-715 (1-10 nM) or the novel nonpeptide B1 receptor antagonist with a benzodiazepine core (3-30 nM) (Wood et al., 2003). Only one CRC was plotted for each preparation. Therefore, separate control and test tissues were studied simultaneously. Antagonists were added to the preparation at least 10 min before challenge with des-Arg9-bradykinin. Distinct periods of incubation with the antagonists were tested, and a contact time of 10 min was selected.
Effect of Different Inhibitors on the Up-Regulation of des-Arg9-Bradykinin-Induced Contraction. To assess the possible involvement of protein synthesis in the lipopolysaccharide-induced enhancement of pig iris sphincter contractile response to des-Arg9-bradykinin, CRCs were plotted 4 h after lipopolysaccharide incubation (the previously selected best period of pretreatment) in the absence or presence of PDTC (300 μM, NF-κB activation inhibitor), actinomycin D (1 μM, mRNA transcription inhibitor), cycloheximide (70 μM, translation/N-glycosylation inhibitor), or dexamethasone (100 μM). The role of MAP kinases was also evaluated. To this end, the preparations were exposed to SB203580 (10 μM, p38 MAP kinase inhibitor) or to PD98059 (10 μM, ERK1/2 inhibitor). Tissues were continuously exposed to lipopolysaccharide (4 h) in the absence (control) or presence of one of the inhibitors indicated above throughout the whole period of incubation.
To determine any possible nonspecific effect of the inhibitors on the contractile response to des-Arg9-bradykinin, at the end of experiments, a complete CRC for carbachol (1-3000 nM) in the absence or presence of each inhibitor was obtained.
Effect of MAP Kinase Inhibitors on des-Arg9-BradykininInduced Signaling Transduction. In another set of experiments, to verify the participation of the MAP kinase activation in B1 receptor-activated signaling transduction pathways, the receptor was induced by the continuous incubation with lipopolysaccharide and the contractile response to des-Arg9-bradykinin was confirmed at 3.5 h after establishment of the preparations (control). Then, preparations were incubated with SB203580 (10 μM, a p38 MAP kinase inhibitor) or PD98059 (10 μM, an ERK1/2 inhibitor) for 30 min, and they were again exposed to the B1 receptor agonist des-Arg9-bradykinin. The concentrations of all inhibitors used in this work were selected on the basis of previous studies (Medeiros et al., 2001, 2004).
Determination of MAP Kinase Activation: Western Blot Analysis. To determine whether MAP kinases are activated after incubation with lipopolysaccharide, pig iris sphincters were carefully dissected from adhering tissues (two preparations per eye) and placed individually in 24-well plates filled with serum-free Dulbecco's modified Eagle's culture medium. The incubation was performed at 37°C in 5% CO2 humidified air, for different periods of time (0-180 min) in the absence (control) or presence of lipopolysaccharide (1 μg/ml). Some tissues were incubated with lipopolysaccharide plus one of the MAP kinase inhibitors. After the incubation period, the pig iris sphincters were frozen in liquid N2. Some tissues were frozen immediately after isolation (basal group).
Cytoplasm protein extraction and Western blot analysis were performed as described previously (Medeiros et al., 2004). Membranes were incubated with antibodies for total and phosphorylated forms of p38 MAP kinase or ERK (Santa Cruz Biotechnology, Santa Cruz, CA). The proteins were visualized by means of the ECL detection system (GE Healthcare, Sao Paolo, Brazil).
Determination of NF-κB Activation: Electrophoretic Mobility Shift Assay. The effect of lipopolysaccharide incubation on NF-κB activation was also evaluated by using electrophoretic mobility shift assay. Pig iris sphincters were incubated with lipopolysaccharide (1 μg/ml) for several different intervals of time (0-24 h) and collected as described above. Nuclear protein extraction and electrophoretic mobility shift assay were performed as described before (Fernandes et al., 2003; Medeiros et al., 2004). For this purpose, the Gel Shift Assay System Kit from Promega (Madison, WI) was used according to the manufacturer's instructions. In brief, NF-κB doublestranded consensus oligonucleotide probe (5′-AGTTGAGGGGACTTTCCCAGGC-3′) was end-labeled with [γ-32P]ATP (GE Healthcare) in the presence of T4 polynucleotide kinase for 10 min at 37°C. Unincorporated nucleotides were removed by passing the reaction mixture over a Sephadex G-25 spin column (GE Healthcare). In a total volume of 20 μl, nuclear extracts (10 μg) were incubated with gelshift binding buffer [(10 mM Tris-HCl, pH 7.5, 1 mM MgCl2, 50 mM NaCl, 0.5 mM dithiothreitol, 0.5 mM EDTA, 4% glycerol, and 1 μg of poly(dIdC)] for 20 min at room temperature. Furthermore, each sample was incubated for 30 min at room temperature with 25,000 cpm of 32P-labeled NF-κB consensus oligonucleotide. Protein-DNA complexes were resolved by nondenaturing 6% acrylamide/bisacrilamide (37.5:1) in 0.25× Tris-borate/EDTA buffer at 150 V for 2 h. The gel was vacuum-dried and analyzed using a FUJIX BAS 2000 (Düsseldorf, Germany) PhosphorImager system.
Statistical Analysis. Results are expressed as the mean ± S.E.M. of four to seven independent experiments.

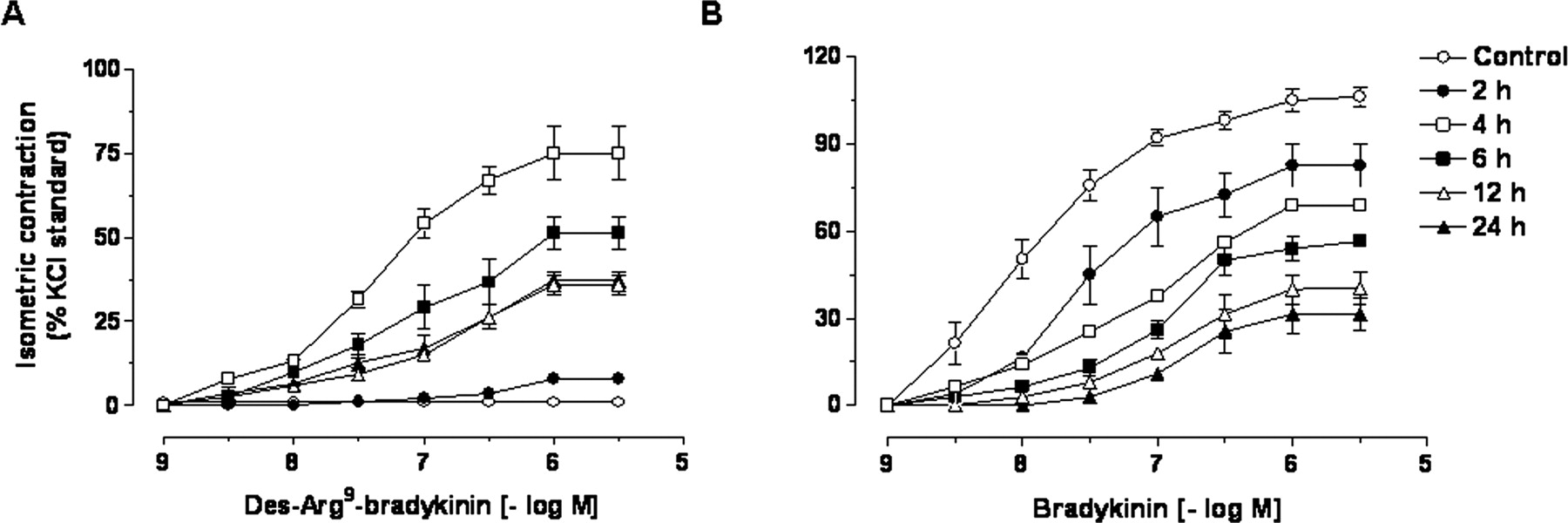
Mean cumulative concentrationresponse curves for des-Arg9-bradykinin (A) and bradykinin (B) in control and lipopolysaccharide (2-24 h)-incubated pig iris sphincter preparations. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.

Mean cumulative concentration-response curves for des-Arg9-bradykinin in lipopolysaccharide (4 h)-incubated pig iris sphincter preparations in the absence or presence of selective kinin B1 des-Arg9-[Leu8]-bradykinin (A, 3, 10, and 30 μM), R-715 (B, 1, 3, and 10 nM) or benzodiazepine (C, 3, 10, and 30 nM), or B2 Hoe 140 (D, 30 nM) receptor antagonists. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.
Results
As described beforehand (El Sayah and Calixto, 2003a), bradykinin (1-3000 nM) caused a marked and concentrationdependent contraction of the pig iris sphincter muscle with a maximal response corresponding to 100% of the carbachol effect, whereas the selective B1 receptor agonist des-Arg9-bradykinin completely failed to cause any detectable contraction of this preparation, even when tested at high concentrations (up to 1 μM). The results in Fig. 1A show that continuous incubation of preparations with lipopolysaccharide (1 μg/ml) resulted in a striking increase of des-Arg9-bradykinin-induced contractile responses. This event was time-dependent, reaching the maximum at 4 h (approximately 75% in relation to KCl) and remaining significant for up to 24 h (approximately 30% in relation to KCl). On the other hand, the contractile responses evoked by bradykinin were significantly low in pig iris sphincter preparations treated with lipopolysaccharide (Fig. 1B). The reduction of bradykinin-induced contraction was found to be a time-dependent process, being maximally lessened 24 h after lipopolysaccharide incubation (responses of approximately 30% in relation to KCl).
The data shown in Fig. 2 demonstrate the effect of the selective B1 receptor antagonists des-Arg9-[Leu8]-bradykinin (3-30 μM), R-715 (1-10 nM), and the recently developed nonpeptide benzodiazepinic antagonist (3-30 nM) on des-Arg9-bradykinin-induced contraction of pig iris sphincter preparations exposed to lipopolysaccharide (1 μg/ml) for 4 h. The preincubation of the preparations with the B1 receptor antagonists produced a concentration-dependent shift to the right, associated with the reduction of maximal responses elicited by des-Arg9-bradykinin. Conversely, the selective kinin B2 receptor antagonist Hoe 140 (30 nM) was not able to significantly alter des-Arg9-bradykinin-induced contraction of the pig iris sphincter preparations pretreated with lipopolysaccharide (Fig. 2).
We further assessed some of the possible mechanisms involved in the up-regulation of B1 receptor-mediated contractile responses induced by lipopolysaccharide in the pig iris sphincter. For this purpose, preparations were continuously exposed to lipopolysaccharide (4 h) in the absence or presence of various classes of inhibitors. The results obtained clearly indicate that incubation with the protein synthesis inhibitors actinomycin D (1 μM, mRNA transcription inhibitor), cycloheximide (70 μM, translation/N-glycosylation inhibitor), or dexamethasone (100 μM) caused a complete blockage of the contractile effects evoked by des-Arg9-bradykinin (1-3000 nM) in lipopolysaccharide-treated pig iris sphincter preparations (Fig. 3). Another set of data shows that PDTC (300 μM), an inhibitor of NF-κB pathway activation, was able to significantly reduce the contraction evoked by des-Arg9-bradykinin after the exposure to lipopolysaccharide (Fig. 4). In addition, the up-regulation of des-Arg9-bradykinin-induced contraction after the incubation with lipopolysaccharide was markedly prevented by the p38 MAP kinase inhibitor SB203580 (10 μM) and to a lesser extent by the ERK1/2 inhibitor PD98059 (10 μM) (Fig. 5). It is worth mentioning that none of the drugs tested in this section were capable of significantly affecting carbachol-induced contraction of the pig iris sphincter muscle (Table 1).
Effect of several classes of drugs on 3000 nM carbachol-induced contraction in pig iris sphincter
Results are expressed as the mean ± S.E.M. of four experiments.
To analyze whether the activation of MAP kinases is implicated in B1 receptor-activated signaling transductional pathways, SB203580 or PD98059 (both 10 μM, selective inhibitors of p38 and ERK1/2, respectively) were incubated 30 min before the construction of CRCs for des-Arg9-bradykinin in preparations continuously pretreated with lipopolysaccharide (4 h). Data revealed that PD98059 incubation fully reduced the contraction induced by des-Arg9-bradykinin in lipopolysaccharide-treated tissues, whereas this response was only partially reduced by SB203580 (Fig. 6).

Effect of continuous incubation with the protein synthesis modulators cycloheximide (A, 70 μM), actinomycin D (B, 1 μM), or dexamethasone (C, 100 μM) on the up-regulation of des-Arg9-bradykinin-evoked contractile responses in lipopolysaccharide (4 h)-stimulated pig iris sphincter preparations. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.
The relevance of p38 MAP kinase and ERK1/2 for the effects of lipopolysaccharide in the pig iris sphincter muscle was further determined by Western blot analysis. Lipopolysaccharide incubation (1 μg/ml) resulted in a time-dependent increase of both p38 MAP kinase and ERK1/2 activation, which was maximal between 5 and 30 min, returning to control levels after 120 min (Fig. 7). Finally, lipopolysaccharide was found to be able to induce a marked activation of the transcriptional factor NF-κB in the pig iris sphincter muscle, as indicated by the electrophoretic mobility shift assay experiments, which revealed an evident increase of NF-κB/DNA-binding activity that peaked between 2 and 4 h after the incubation of lipopolysaccharide (Fig. 8).

Effect of continuous incubation with the NF-κB inhibitor PDTC (300 μM) on the up-regulation of des-Arg9-bradykinin-evoked contractile responses in lipopolysaccharide (4 h)-stimulated pig iris sphincter preparations. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.
Discussion
The relevance of kinins and their receptors has been well demonstrated in several biological systems (Calixto al., 2001; Leeb-Lundberg et al., 2005; Moreau et al., 2005). Regarding the ocular system, only a few studies so far have assessed the role played by kinins. Bradykinin, acting via B2 receptors, evokes contractile, inflammatory, and nociceptive responses of the rabbit iris sphincter muscle (Griesbacher and Lembeck, 1987; Geppetti et al., 1990; Wang and Hakanson, 1992; Hall et al., 1995). More recently, El Sayah and Calixto (2003a) have shown that bradykinin produces potent and graded contraction of the pig iris sphincter by activating B2 receptors, whereas the selective B1 receptor agonist des-Arg9-bradykinin, even at high concentrations (up to 1 μM), has no detectable contractile effect on this preparation. These results indicate the presence of constitutive B2 but not B1 receptors in this pig tissue. The present study evaluates the ability of lipopolysaccharide to up-regulate the contractile responses evoked by des-Arg9-bradykinin in the pig iris sphincter muscle, attempting to determine the mechanisms that are possibly involved in this process.
Our results clearly indicate that lipopolysaccharide induced a time-dependent and noticeable increase of des-Arg9-bradykinin-induced contraction, which peaked after 4 h. A similar up-regulation of B1 receptor-mediated contractile responses has been demonstrated beforehand in other smooth muscle preparations (Marceau et al., 1998; Calixto et al., 2004; Leeb-Lundberg et al., 2005), but to our knowledge, this is the first experimental evidence on the functional induction of B1 receptors by lipopolysaccharide in the pig iris sphincter. Present data extend previous evidence showing that either pre-existing infection or treatment with lipopolysaccharide results in the up-regulation of B1 receptors in the vascular tissues of pigs (Schremmer-Danninger et al., 1998; Siebeck et al., 1998). El Sayah and Calixto (2003a) have shown that in vitro incubation of pig iris sphincter preparations for 6 h leads to a slight but detectable (responses approximately 15% of carbachol) up-regulation of des-Arg9-bradykinin-induced contraction. We might assume that B1 receptors represent pivotal molecules during infectious and inflammatory processes of the ocular system.
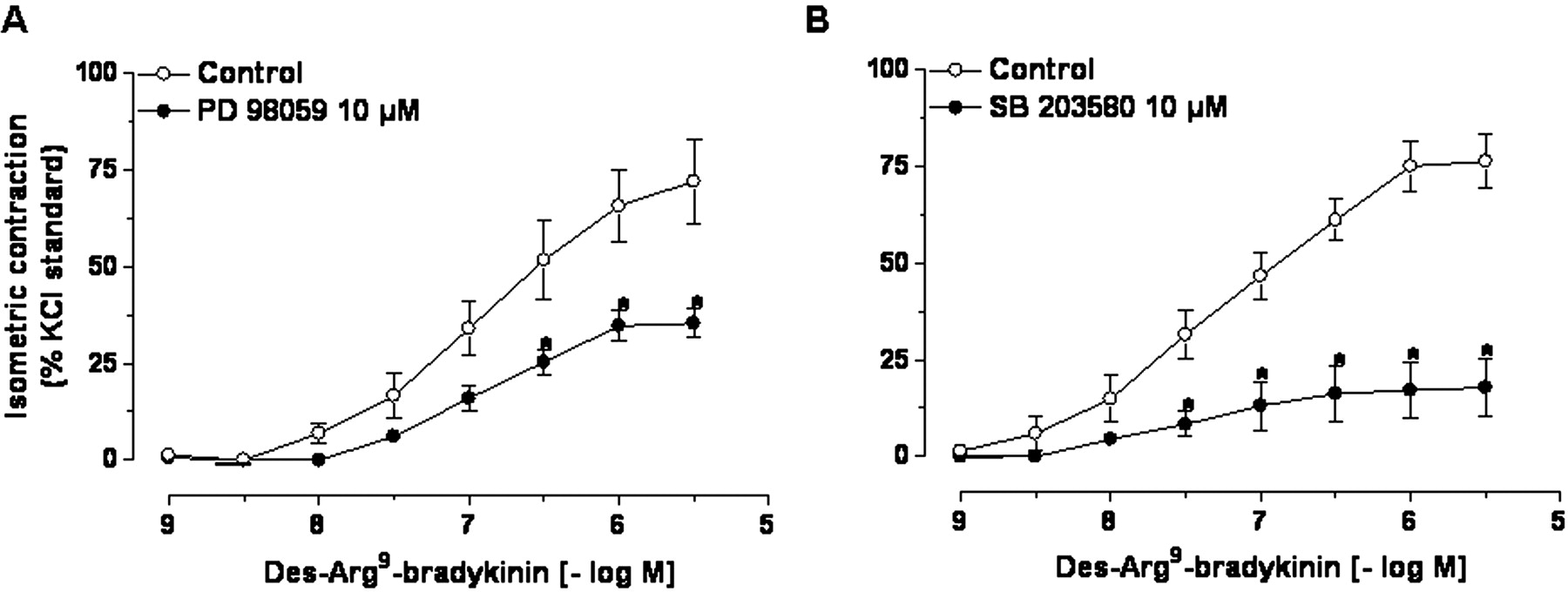
Effect of continuous incubation with ERK1/2 PD98059 (A, 10 μM) or p38 SB203580 (B, 10 μM) MAP kinase blockers on the up-regulation of des-Arg9-bradykinin-evoked contractile responses in lipopolysaccharide (4 h)-stimulated pig iris sphincter preparations. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.
Another part of our work revealed that continuous incubation of the preparations with lipopolysaccharide caused a time-related and pronounced reduction of the contractile responses evoked by bradykinin in the pig iris sphincter muscle. These results make it clear that up-regulation of B1 receptors by lipopolysaccharide in this tissue was associated with a marked down-regulation of B2 receptors, at least at the functional level. Previous evidence indicated that lipopolysaccharide treatment caused an increase of B1-receptormediated rat paw edema, accompanied by a marked decrease in B2 receptor-mediated edematogenic response (Campos et al., 1996). This has also been shown for the contraction evoked by B1 and B2 agonists in the stomach fundus of lipopolysaccharide-treated rats (Cabrini et al., 1996). A similar event has been described by Phagoo et al. (1999), suggesting a shift of kinin receptor subtypes from B2 to B1 in human lung fibroblasts in the presence of the IL-1β cytokine. Although the precise mechanisms implicated in this coordinated regulation of B1 and B2 receptors remain to be further clarified, it is possible that one or more lipopolysaccharideactivated signaling pathways (including the production of cytokines) could be associated with it.
It is noteworthy that our study indicates that contraction evoked by des-Arg9-bradykinin in lipopolysaccharide-treated pig iris sphincter preparations is dependent on the activation of kinin B1 (but not B2) receptors. Therefore, two peptidic (des-Arg9-[Leu8]-bradykinin and R-715) and one nonpeptidic (benzodiazepine) selective B1 receptor antagonists potently and concentration-dependently antagonized des-Arg9-bradykinin-induced contractile responses, whereas the selective B2 receptor antagonist Hoe 140 (even when tested in high concentrations) had no significant effect. It is worth mentioning that B1 receptor antagonists tested herein exhibited a clear mixed competitive and noncompetitive profile of action, producing a displacement to the right of the CRCs for des-Arg9-bradykinin, accompanied by a reduction of the maximal asymptote. This pharmacological outline has been demonstrated elsewhere for B1 receptor antagonists in other tissues and animal species (Larrivée et al., 2000).

Mean cumulative concentrationresponse curves for des-Arg9-bradykinin in lipopolysaccharide (4 h)-incubated pig iris sphincter preparations in the absence or presence of ERK1/2 PD98059 (A, 10 μM) or p38 SB203580 (B, 10 μM) MAP kinase inhibitors. Results are expressed as percentages of the contraction induced by 80 mmol of KCl. Each point represents the mean of five experiments, and the vertical lines show the S.E.M.

A, time-related effect of lipopolysaccharide (5-180 min) incubation on the activation of ERK1/2 and p38 MAP kinases in pig iris sphincter preparations. B, results were normalized by arbitrarily setting the densitometry of the basal group (0) and are expressed as mean ± S.E.M. of three independent experiments. PD98059 and SB203580 (both 10 μM) were used as positive control drugs to inhibit the activation of ERK1/2 and p38 MAP kinases, respectively.
B1 receptor up-regulation in the pig iris sphincter after incubation with lipopolysaccharide seems to represent a de novo synthesis process, as suggested by the subsequent set of experiments: 1) the functional up-regulation of B1 receptors in this tissue was a time-dependent process that required 4 h to reach the maximum; 2) the increase of B1 receptor-mediated contractile responses was fully inhibited by the protein synthesis inhibitors actinomycin D and cycloheximide; and 3) this event was abolished by dexamethasone, a drug that clearly inhibits the expression of several proinflammatory proteins. It has been widely demonstrated that pharmacological modulators of protein synthesis are able to prevent the up-regulation of B1 receptors in a number of in vivo and in vitro paradigms (Marceau et al., 1998; Sardi et al., 2000; Calixto et al., 2004). Of note in our experiments, actinomycin D, cycloheximide, and dexamethasone did not significantly affect the contractile responses evoked by carbachol in the pig iris sphincter, indicating that these drugs do not interfere with tissue viability and that this phenomenon is probably specific to B1 receptors.
A series of publications (using both in vivo and in vitro approaches) have provided convincing evidence of the relevance of NF-κB for the up-regulation of B1 receptors (Ni et al., 1998; Schanstra et al., 1998; Campos et al., 1999; Medeiros et al., 2001, 2004; Sabourin et al., 2002). Our data patently point to NF-κB as an important element for B1 receptor up-regulation induced by lipopolysaccharide in pig iris sphincter. First, lipopolysaccharide-evoked increase of des-Arg9-bradykinin-induced contraction was significantly prevented by the NF-κB inhibitor PDTC. Furthermore, lipopolysaccharide incubation with the pig iris sphincter produced a marked and time-related activation of NF-κB in periods of time that preceded the up-regulation of B1 receptors. Of interest, several recent publications have proposed a pivotal role for NF-κB in the model of uveitis induced by lipopolysaccharide in rats (Ohta et al., 2002; Chi et al., 2004; Fang et al., 2005; Yang et al., 2005). It is tempting to speculate that inflammatory responses after infectious stimuli in the ocular system probably involve the activation of transcriptional factor NF-κB.
Several lines of evidence have emphasized the importance of different members of the MAP kinase family for the upregulation of B1 receptors (Calixto et al., 2004). It has been demonstrated that spontaneous or IL-1β-caused overexpression of B1 receptors in the rabbit aorta involves the stimulation of both ERK and p38 kinases (Larrivée et al., 1998). A similar participation of ERK and p38 has been shown for heat stress-induced B1 receptor up-regulation in vascular smooth muscle cells (Lagneux et al., 2001). On the other hand, the up-regulation of B1 receptors after tissue injury in the rat portal vein seems to depend on the activation of p38 and c-Jun NH2-terminal kinase (but not ERK) kinases (Medeiros et al., 2004). Our results revealed that p38 blocker SB203580 and to a lesser extent ERK1/2 inhibitor PD98059 significantly prevented the increase of B1 receptor-mediated contractile responses induced by lipopolysaccharide in the pig iris sphincter. In addition, Western blot analysis clearly showed that, in this preparation, lipopolysaccharide had a similar profile of activation for both p38 and ERK1/2 (which was maximal between 5 and 30 min). These results make it possible to infer a major role of p38 MAP kinase for the modulation of B1 receptors by lipopolysaccharide in the pig iris sphincter, whereas the activation of ERK1/2 might be associated with the regulation of other inflammatory proteins, rather than kinin B1 receptors.

A, time-dependent effect of lipopolysaccharide (30 min to 24 h) incubation on NF-κB-specific DNA-protein complex formation in pig iris sphincter preparations. B, results were normalized by arbitrarily setting the densitometry of the basal group (0) and are expressed as the mean ± S.E.M. of three independent experiments.
An aspect that must be considered is that MAP kinases can also represent important signaling transductional pathways associated with the stimulation of B1 receptors (Liebmann, 2001; Calixto et al., 2004). This hypothesis has been tested through an independent series of experiments in which p38 SB203580 or ERK1/2 PD98059 MAP kinase inhibitors were incubated after the period required for the up-regulation of B1 receptors by lipopolysaccharide. Of note, in contrast to that demonstrated for the up-regulation process, des-Arg9-bradykinin-induced contraction in lipopolysaccharide-pretreated preparations was markedly reduced by ERK1/2 inhibitor PD98059, whereas p38 SB203580 blocker had only a slight effect. We might suggest that the MAP kinases p38 and ERK1/2 are activated in a different manner to mediate up-regulation and signaling transductional pathways of the B1 receptors. Additional studies using different experimental models should be performed to confirm this idea.
In summary, the results of the present work indicate that lipopolysaccharide incubation causes a marked and timerelated up-regulation of B1 (associated with a striking reduction of B2) receptor-mediated contractile responses in pig iris sphincter. B1 receptor modulation seems to involve de novo protein synthesis, the activation of MAP kinases (mainly p38) and the stimulation of NF-κB pathways. We have also evidenced the participation of MAP kinase ERK1/2 as an intracellular signaling pathway activated after B1 receptor stimulation. This article provides new insights into the role of kinin receptors, especially the B1 subtype, during infectious and inflammatory responses of the ocular system and sheds light on B1 receptors as potential targets for the development of new therapeutic options in the treatment of eyerelated diseases.
Footnotes
-
This work was supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil), the Programa de Apoio aos Núcleos de Excelência (Brazil), and by the Fundação de Apoio à Pesquisa de Santa Catarina. R.M. and E.S.F. are PhD students in pharmacology receiving grants from CNPq and CAPES, respectively. M.M.C. holds a postdoctoral fellowship from CAPES.
-
ABBREVIATIONS: MAP, mitogen-activated protein; NF-κB, nuclear factor-κB; CRC, complete concentration-response curve; ERK1/2, extracellular signal-regulated kinases 1 and 2; IL, interleukin; PDTC, pyrrolidinedithiocarbamate; SB203580, 4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl)1H-imidazole; PD98059, 2′-amino-3′-methoxyflavone; R-715, Ac-Lys-[d-β-Nal7,Ile8]des-Arg9-bradykinin; Hoe 140, d-Arg0-[Hyp3,Thi5,d-Tic7,Oic8]-bradykinin.
- Received November 23, 2005.
- Accepted February 7, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}