


Abstract
Biguanides are a class of drugs widely used as oral antihyperglycemic agents for the treatment of type 2 diabetes mellitus, but they are associated with lactic acidosis, a lethal side effect. We reported previously that biguanides are good substrates of rat organic cation transporter 1 (Oct1; Slc22a1) and, using Oct1(−/−) mice, that mouse Oct1 is responsible for the hepatic uptake of a biguanide, metformin. In the present study, we investigated whether the liver is the key organ for the lactic acidosis. When mice were given metformin, the blood lactate concentration significantly increased in the wild-type mice, whereas only a slight increase was observed in Oct1(−/−) mice. The plasma concentration of metformin exhibited similar time profiles between the wild-type and Oct1(−/−) mice, suggesting that the liver is the key organ responsible for the lactic acidosis. Furthermore, the extent of the increase in blood lactate caused by three different biguanides (metformin, buformin, and phenformin) was compared with the abilities to reduce oxygen consumption in isolated rat hepatocytes. When rats were given each of these biguanides, the lactate concentration increased significantly. This effect was dose-dependent, and the EC50 values of metformin, buformin, and phenformin were 734, 119, and 4.97 μM, respectively. All of these biguanides reduced the oxygen consumption by isolated rat hepatocytes in a concentration-dependent manner. When the concentration required to reduce the oxygen consumption to 75% of the control value (from 0.40 to 0.29 μmol/min/mg protein) was compared with the EC50 value obtained in vivo, a clear correlation was observed among the three biguanides, suggesting that oxygen consumption in isolated rat hepatocytes can be used as an index of the incidence of lactic acidosis.
Metformin, a biguanide, is used for the treatment of hyperglycemia in patients with type 2 diabetes mellitus. It was developed during the late 1950s, first marketed in Europe in 1959, and has been available in the United States since 1995 (Davidson and Peters, 1997). Metformin seems to ameliorate hyperglycemia by improving peripheral sensitivity to insulin, reducing gastrointestinal glucose absorption and hepatic glucose production (Caspary and Creutzfeldt, 1971; Hundal et al., 2000;Borst and Snellen, 2001). Recently, metformin has also become available for the treatment of polycystic ovary syndrome (Velazquez et al., 1994;Nestler, 2001) and has been found to improve vascular function (Katakam et al., 2000), prevent pancreatic cancer (Schneider et al., 2001), and reverse fatty liver diseases (Lin et al., 2000) in experimental animals. Thus, a re-evaluation of its pharmacological activity is now underway. Lactic acidosis is a severe adverse effect of biguanides, and phenformin was withdrawn from the market in the 1970s for this reason (Assan et al., 1975; Kwong and Brubacher, 1998). Lactic acidosis is a life-threatening condition characterized by low arterial pH (<7.35) and elevated arterial lactate levels (5.0 mEq/l in humans), and more than 50% of the patients died when lactic acidosis took place under phenformin administration (Brown et al., 1998; Kwong and Brubacher, 1998). Metformin also associated with lactic acidosis in the lower incidence of approximately 3 cases per 100,000 patients per year, compared with a 10- to 20-fold higher incidence for phenformin (Pearlman et al., 1996; Lalau and Race, 2000; Kruse, 2001). Lactic acidosis is observed in patients with renal dysfunction, and because renal secretion is the major elimination route of biguanides (Davidson and Peters, 1997), renal dysfunction will cause a significant increase in plasma biguanide concentration, resulting in lactic acidosis. Metformin has been shown to reduce the oxygen consumption and glucose production in isolated hepatocytes in a concentration-dependent manner (El-Mir et al., 2000; Owen et al., 2000). This has been explained by the inhibition of mitochondrial respiratory complex I, although whether this occurs via direct or indirect inhibition remains unknown (El-Mir et al., 2000; Owen et al., 2000). Excessive inhibition of mitochondrial respiration by biguanide may cause lactic acidosis.
Recently, we demonstrated that the biguanides are good substrates of rat organic cation transporter 1 (Oct1; Slc22a1) (Wang et al., 2002). Oct1 is a polyspecific transporter for small and hydrophilic organic cations such as tetraethylammonium and 1-methyl-4-phenylpyridinium (Grundemann et al., 1994; Dresser et al., 2000; Inui et al., 2000). Oct1 is abundantly expressed in the liver and, to a lesser extent, in the kidney, where it is localized in the basolateral membrane. Previously, we demonstrated that the order of the transport activity (Vmax/Km) of biguanides by rOct1 is phenformin > buformin > metformin. Although no significant reduction was observed in the urinary excretion of metformin, the hepatic uptake of metformin was reduced markedly in Oct1(−/−) mice, and the distribution volume of metformin in the liver of Oct1(−/−) was very close to the hepatic extracellular space (Wang et al., 2002). Therefore, the Oct1(−/−) mouse is a good animal model in which to examine whether the liver is a key organ in the increase of blood lactate concentration.
In the present study, the increase in blood lactate concentration was compared in wild-type and Oct1(−/−) mice when metformin was given by constant intravenous infusion. The increase in lactate concentration was investigated in rats after the infusion of biguanides (phenformin, buformin, and metformin) at different rates, and their potency was compared with that in the reduction in oxygen consumption in isolated rat hepatocytes.
Materials and Methods
Animals and Materials.
Male Sprague-Dawley rats (8 weeks old, 250–280 g of body weight; Charles River Japan Inc., Kanagawa, Japan) and male Oct1(−/−) and wild-type FVB mice (12–16 weeks old) used in the present study (Jonker et al., 2001) were housed at a room temperature of 24 ± 1°C with food and water ad libitum. Metformin and phenformin were purchased from Sigma Chemical (St. Louis, MO). Buformin and perchloric acid (60%) were purchased from Wako Pure Chemicals (Osaka, Japan). Pentobarbital was from Dainippon Pharmaceutical (Osaka, Japan). The L(+)-lactate kit was purchased from Sigma Diagnostics (St. Louis, MO), and the model 5300 biological oxygen monitor was purchased from YSI Inc. (Yellow Springs, OH). All other chemicals were of analytical grade and were commercially available.
Lactic Acidosis Study of Metformin in Mice.
Mice were anesthetized with intraperitoneal sodium pentobarbital (50 mg/kg), and the femoral vein was catheterized with polyethylene tubing for infusion. Infusion was performed using a basic syringe pump (Harvard Apparatus Inc., Holliston, MA). Metformin dissolved in saline was administered at a rate of 8.0 ml/h/kg and a dose of 150 mg/h/kg for 3.5 h. Blood samples used for the determination of metformin were collected from an angular vein at 90, 150, and 210 min. After centrifugation, the plasma samples were deproteinized with four times their volume of acetonitrile and then subjected to HPLC. Blood samples used for the determination of lactate concentration were collected from the tail vein. Whole blood was mixed with double the volume of perchloric acid (8%) and vortexed. After centrifugation, the upper solution was used for lactate determination according to the manufacturer's protocol. After sampling at 210 min, the mice were killed, and the liver and femoral muscles from the opposite femur that had not been catheterized were removed immediately. The removed liver and muscles were homogenized with 4 volumes of phosphate-buffered saline, deproteinized with acetonitrile, and evaporated to dryness. Pellets were dissolved in 200 μl of water for HPLC analysis.
Lactic Acidosis Study of Biguanides in Rats.
Rats were anesthetized with diethyl ether, and the femoral vein was catheterized with polyethylene tubing for infusion. Infusion was performed using the basic syringe pump. Metformin, buformin, and phenformin dissolved with saline were administered for 4 h. The constant infusion doses were 250, 175, 100, 50, and 25 mg/h/kg for metformin, 50, 25, 12.5, 5, and 2.5 mg/h/kg for buformin, and 25, 12.5, 5, 2.5, and 1.0 mg/h/kg for phenformin, respectively. The administration rate of the saline was 8.0 ml/h/kg. Blood samples used for the determination of biguanides were drawn from the cervical vein at 150, 180, and 210 min. Sample treatment, determination of biguanides, and blood lactate concentrations were performed as described above. AUCs of the blood lactate concentration until 240 min were calculated by the linear trapezoidal rule, and EC50 values were estimated by the equation (Emax model) AUC = AUCmaxS/(EC50 +S), where S represents unbound biguanide plasma concentrations. The fitting was carried out by an iterative nonlinear least-squares method.
Respiratory Chain Inhibitory Effect of Biguanides in Isolated Rat Hepatocytes.
Hepatocytes were isolated from rats by the procedure described previously (Yamazaki et al., 1992). The experiment was prepared from the report of El-Mir et al. (2000). Briefly, hepatocytes (final protein concentration of 1.0 mg/ml) were incubated in closed vials at 37°C in a shaking water bath in 5 ml of Krebs-bicarbonate buffer (120 mM NaCl, 4.8 mM KCl, 1.2 mM MgSO4, 24 mM NaHCO3, 1.3 mM CaCl2, pH 7.4) saturated with a mixture of O2/CO2 (95%/5%) supplemented with 20 mM lactate, 2 mM pyruvate, 4 mM octanoate, and different concentrations of biguanides. Oxygen consumption for up to 30 min was monitored and used for data analysis. The concentration of biguanides caused a reduction of oxygen consumption to 0.29 μmol/min/mg protein of hepatocytes (75% of control value) was calculated by linear estimation.
HPLC Analysis.
The HPLC system involved a modell-7100 pump and a model l-7400 UV monitor (Hitachi, Tokyo, Japan) with a 300 × 3.9 mm i.d. C18 μBondapak (10 μm) column purchased from Waters (Milford, MA). The components of mobile phase consisted of 0.01 M phosphate buffer, pH 6.5, and acetonitrile at the ratio of 30:70. The flow rate was 1 ml/min. The wavelength for UV detection was 236 nm. The sensitivity was 1.5 ng for metformin and 3 ng for buformin and phenformin. The retention time was approximate 14, 11, and 9 min for metformin, buformin, and phenformin, respectively. The reproducibility was almost 90%, and no internal standard was used in the study. Biguanides were detected directly.
Statistical Analysis.
Statistical analysis was performed by one-way analysis of variance followed by Fisher's t test or Student's t test to identify significant differences between various treatment groups.
Results
Lactic Acidosis Induced by Metformin in Wild-Type and Oct1(−/−) Mice.
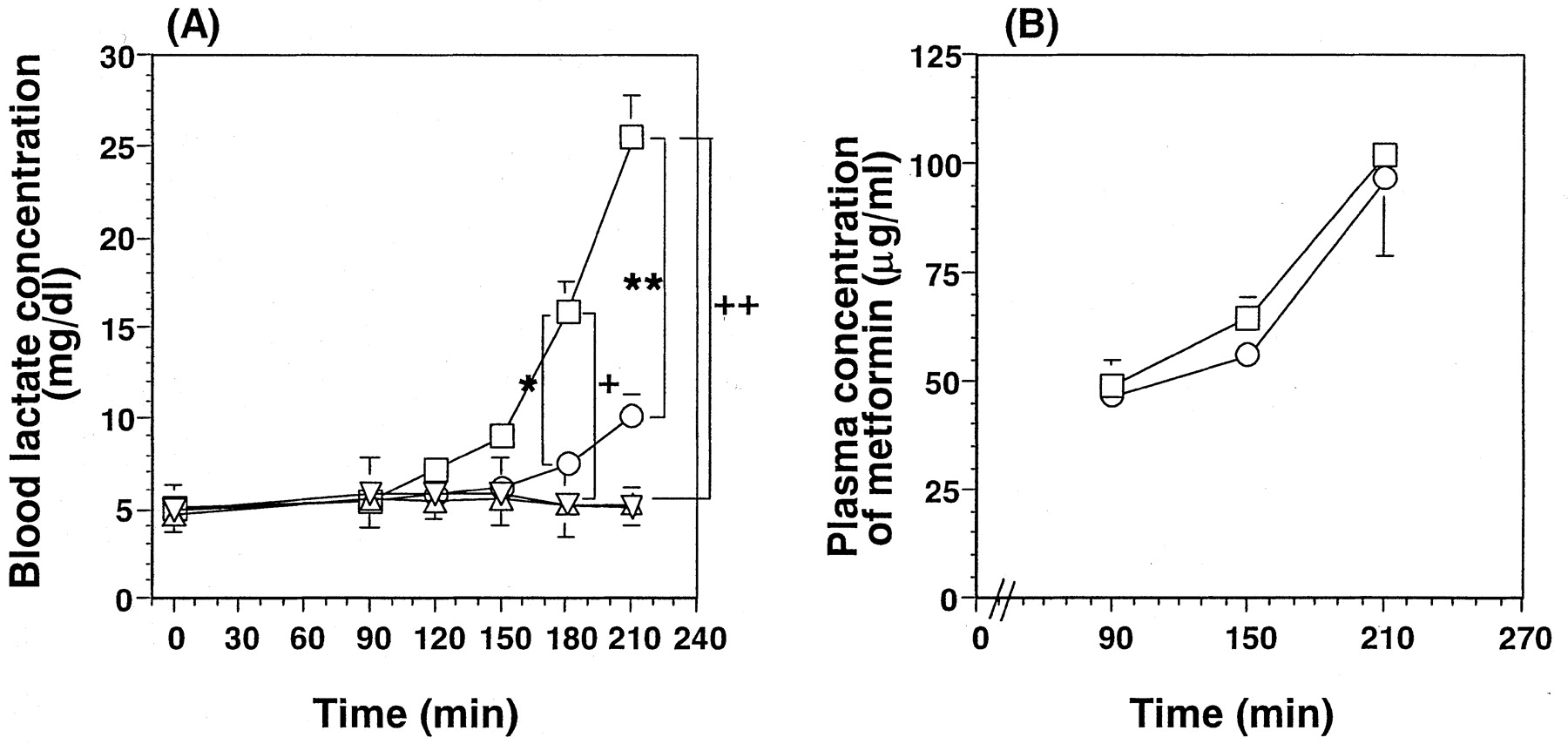
The time profiles of the blood lactate concentrations in wild-type and Oct1(−/−) mice at a constant infusion of a dose of 150 mg/h/kg of metformin are shown in Fig.1A, and the simultaneously determined plasma concentrations of metformin are shown in Fig. 1B. There was a marked difference in the response to metformin between wild-type and Oct1(−/−) mice after 180 and 210 min of intravenous infusion of metformin, although the plasma concentrations of metformin were comparable. At 210 min, the blood lactate concentration was 25.6 ± 2.2 mg/dl in metformin-treated wild-type mice, which was significantly increased compared with that in saline-treated mice (5.07 ± 1.05 mg/dl), whereas there was no statistical difference in the blood lactate concentration between metformin and saline-treated Oct1(−/−) mice (10.2 ± 1.1 and 5.25 ± 0.17 mg/dl, respectively). The blood lactate concentration in metformin-treated wild-type mice was 2.5-fold greater than that in metformin-treated Oct(−/−) mice. After the mice were sacrificed, the concentration of metformin in the liver and skeletal muscle was determined (Table1). In contrast to the significant reduction in metformin concentration in the liver of Oct1(−/−) mice, the concentration of metformin in muscle was similar in both groups.

Time profile of lactate concentration (A) and metformin plasma concentration (B) in wild-type and Oct1(−/−) mice during intravenous infusion of 150 mg/h/kg. Metformin was administered by constant intravenous infusion at a rate of 150 mg/h/kg. The whole blood lactate concentration in wild-type (■) and Oct1(−/−) mice (○) was compared with those in saline-treated wild-type and Oct1(−/−) mice (▵, A). B, there was no marked difference in the metformin plasma concentration. Statistical analysis was performed by one-way analysis of variance followed by Fisher's ttest. *, different tendency (p = 0.1; metformin-treated wild-type versus Oct1(−/−) mice at 180 min); significant difference at p < 0.05: + and ++, metformin-treated versus saline-treated wild-type mice at 180 and 210 min, respectively; **, metformin-treated wild-type versus Oct1(−/−) mice at 210 min. Results are shown as mean ± S.E. of three wild-type and four Oct1(−/−) mice.
Plasma concentration and tissue accumulation of metformin after i.v. infusion at the dose of 150 mg/h/kg for 210 min
Lactic Acidosis Induced by Biguanides in Rats.
The time profiles of the lactate concentration induced by constant intravenous infusion of phenformin, buformin, and metformin in rats are shown in Fig. 2. Compared with administration of saline, the blood lactate concentration was increased after each dose of metformin, buformin, and phenformin (Fig. 2, A-C). The correlation between increase in lactate AUC and the steady-state unbound plasma concentration of biguanides is shown in Fig. 2D. Taking the highest blood lactate AUC as 100%, the EC50 values of the biguanides were determined to be 734 ± 168, 119 ± 18, and 4.97 ± 0.87 μM for metformin, buformin, and phenformin, respectively (Table 2). There were 7- and 140-fold differences in the EC50 values for lactic acidosis between phenformin and buformin and between phenformin and metformin, respectively (Fig. 2D) (Table 2).

Time profiles of the lactic acid increase induced in rats by metformin (A), buformin (B), and phenformin (C) and the relationship between the induced lactate AUC and the plasma concentration of biguanides (D). The whole blood lactate increase induced by metformin (A), buformin (B), and phenformin (C) is shown. The constant infusion doses were 25 (◇), 50 (▵), 100 (○), 175 (⋄), and 250 mg/h/kg (■) for metformin; 2.5 (◇), 5 (▵), 12.5 (○), 25 (⋄), and 50 mg/h/kg (■) for buformin; and 1 (◇), 2.5 (▵), 5 (○), 12.5 (⋄), and 25 mg/h/kg (■) for phenformin. The lactate was significantly increased in biguanide-treated animals compared with control (⊞), and this increase was dose-dependent. D, the increase in lactate AUC with regard to steady-state unbound plasma concentration of biguanides; ▵, phenformin; ○, buformin; ■, metformin. Phenformin was the most potent inducer of blood lactate, and metformin was the least. Results are shown as the mean ± S.E. of three rats.
EC50 of lactic acidosis induced by biguanide compounds in rats
Oxygen Consumption Inhibition Study in Isolated Rat Hepatocytes.
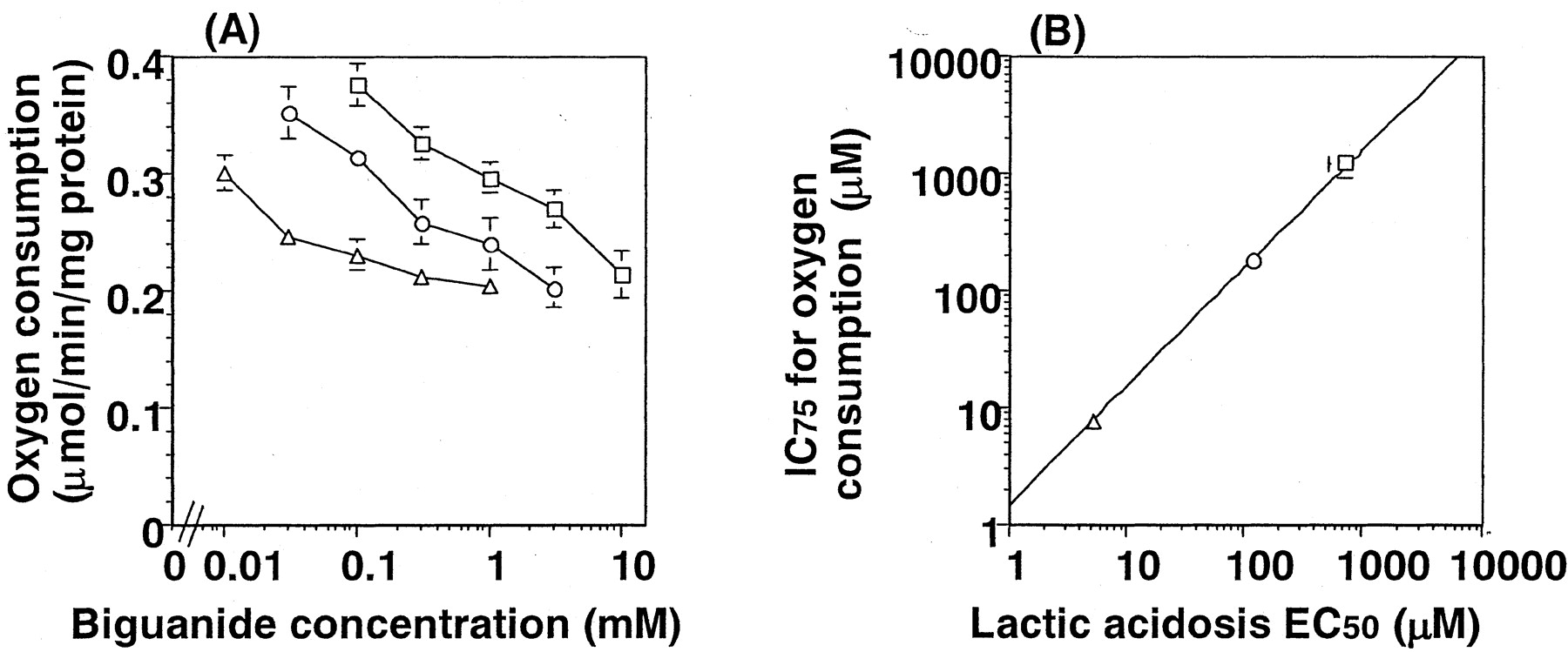
The effect of metformin, buformin, and phenformin on the oxygen consumption was examined using isolated rat hepatocytes. The control value of oxygen consumption by isolated rat hepatocytes was 0.389 ± 0.098 μmol/min/mg protein of hepatocytes. The concentration dependence of the reduction in oxygen consumption is shown in Fig. 3A. Oxygen consumption was decreased in the presence of biguanides in a concentration-dependent manner. Concerning the reduction in oxygen consumption, phenformin was the most potent drug. The concentrations of biguanides causing a reduction in oxygen consumption to 75% of the control value (0.29 μmol/min/mg protein) of rat hepatocytes were 1020 ± 136, 173 ± 15, and 7.25 ± 1.58 μM, and they correlated with the EC50 values determined in vivo for the increase of blood lactate (Fig. 3B).
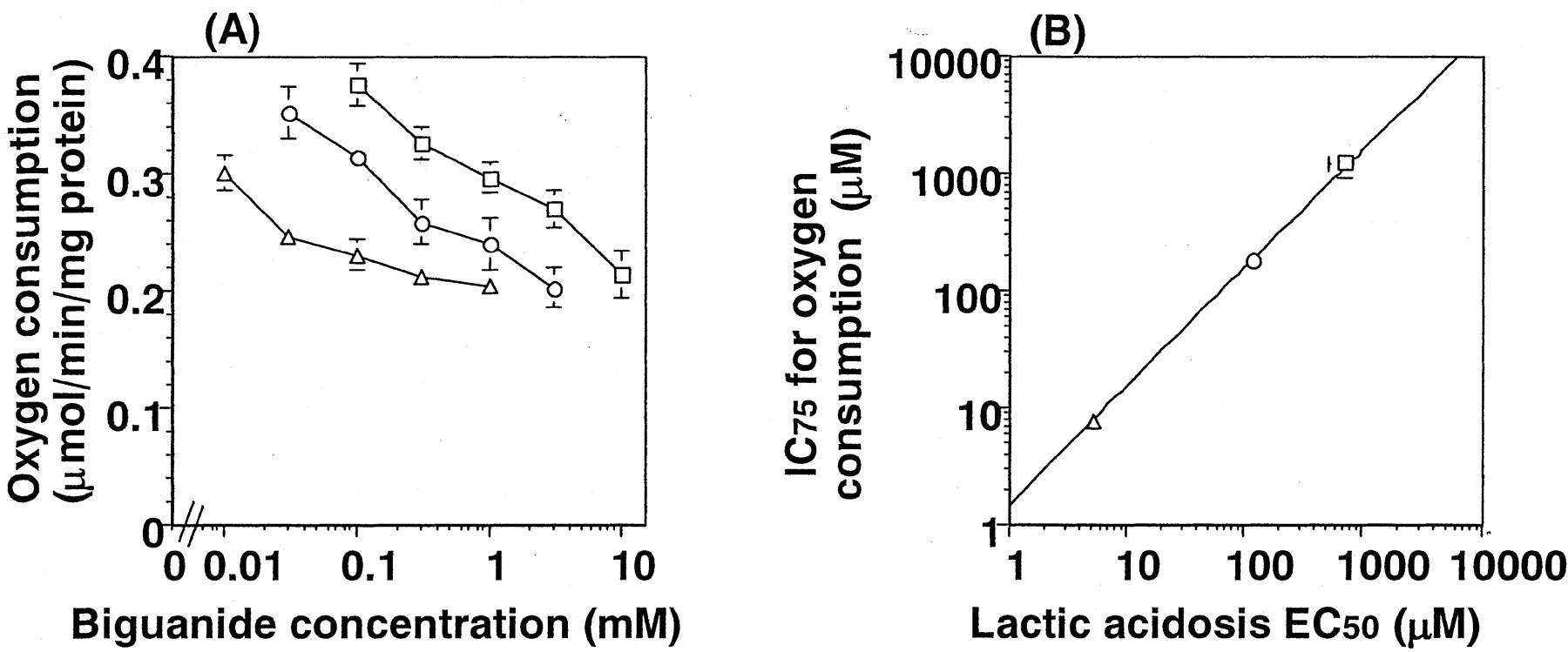
Respiratory chain inhibitory effect in isolated rat hepatocytes (A) and the correlation between lactic acidosis (in vivo) and respiratory chain inhibition (in vitro) (B). The inhibitory effect of biguanides on the mitochondrial respiration was determined in isolated rat hepatocytes. The reduction in oxygen consumption was taken as the index of respiratory chain inhibition. The oxygen consumption was reduced by biguanides compared with the control value of 0.389 ± 0.098 μmol/min/mg protein of hepatocytes. Metformin (■) showed the lowest inhibitory effect and phenformin (▵) the highest. Buformin (○) showed an inhibitory effect between those of metformin and phenformin. A correlation between the concentration of biguanides caused a reduction of oxygen consumption to 0.29 μmol/min/mg protein of hepatocytes (75% of control value), and their EC50values for lactic acidosis is shown in B. Results are the mean ± S.E. of three determinations. Error bars for some points lie within the symbols.
Discussion
The present study investigated whether the liver is the key organ for lactic acidosis, and it examined whether oxygen consumption by isolated hepatocytes can be used as an index of the incidence of lactic acidosis. Lactate, produced in the gut, liver, and peripheral tissues such as erythrocyte and skin, is used to form glucose in the liver (Cori cycle) (Radziuk and Pye, 2001). Two possible mechanisms for lactic acidosis caused by biguanides have been proposed: 1) increased lactate production in the peripheral tissues, because metformin increases the glycolytic lactate production in peripheral tissue (Borst and Snellen, 2001); and 2) inhibition of lactate metabolism/transport in the liver and other tissues such as heart and muscle. Oct1(−/−) mice provide a good animal model to examine these possibilities because the hepatic concentration of metformin is drastically reduced in these mice at similar plasma concentrations. The importance of the intrahepatic concentration of metformin was shown by comparing the effect of metformin on the lactate concentration between wild-type and Oct1(−/−) mice (Fig. 1). At 210 min, the blood lactate concentration was significantly increased in metformin-treated wild-type mice compared with that in saline-treated mice, whereas there was no statistical difference in the blood lactate concentration between metformin and saline-treated Oct1(−/−) mice (Fig. 1A). The blood lactate concentration in metformin-treated wild-type mice was 2.5-fold greater than that in metformin-treated Oct(−/−) mice (Fig. 1A). The plasma concentration-time profiles of metformin were similar in wild-type and Oct1(−/−) mice (Fig. 1B). As summarized in Table 1, the hepatic concentration of metformin in Oct1(−/−) mice was much reduced, whereas its concentration in the skeletal muscle was comparable between Oct1(−/−) and wild-type mice. These results indicate the importance of the intrahepatic metformin, which may inhibit the lactate metabolism in the liver in causing lactic acidosis. Buformin and phenformin have been shown to be substrates of Oct1, with greater transport activity than metformin (Wang et al., 2002). Because Oct1 is responsible for the hepatic uptake of organic cations (Jonker et al., 2001; Wang et al., 2002), it is very likely that the hepatic uptake of buformin and phenformin is also accounted for by Oct1, and the lack of Oct1-mediated hepatic uptake will reduce their inducibility of blood lactate.
The study using Oct1(−/−) mice thus indicates that lactic acidosis should be ascribed to the effect of biguanides on the liver. El-Mir et al. (2000) demonstrated that metformin reduced the oxygen consumption in a concentration-dependent manner in isolated rat hepatocytes. The relationship between the inhibition of oxygen consumption by the biguanides and the inducibility of blood lactate was investigated to determine the correlation between them. An increase in blood lactate was investigated in rats that were given biguanides at different infusion rates. Increasing the infusion rate of metformin, buformin, and phenformin caused a significant increase in blood lactate concentration (Fig. 2). Phenformin was the most potent drug to cause an increase in blood lactate concentration. The EC50values of phenformin, buformin, and metformin were determined using their unbound plasma concentration at steady state and the AUC of blood lactate. As summarized in Table 1, there was a 7-fold and 140-fold difference in the EC50 values between phenformin and buformin and between phenformin and metformin, respectively. As shown in Fig. 3, biguanides reduced oxygen consumption in isolated rat hepatocytes in a concentration-dependent manner. The EC50 values determined in vivo using unbound plasma concentration of biguanides were compared with their concentration needed to cause 75% of control value of oxygen consumption in isolated rat hepatocytes (Fig. 3). There was a clear linear correlation among the three biguanides examined in this study (Fig. 3), indicating that the reduction of oxygen consumption can be used as an index of the incidence of lactic acidosis. It is possible that excessive inhibition of mitochondrial respiration in the liver causes lethal reduction in the hepatic clearance of lactate. This should be examined in future studies, including an examination of the relationship between oxygen consumption and glucose production rate from lactate. Recently, human hepatocytes have become available for preclinical research, and these should allow the incidence of lactic acidosis to be predicted in humans.
The present study highlights two important issues, namely the transport activity of Oct1 and the saturation of hepatic uptake. Hepatic uptake is one of the main factors for determining the intrahepatic concentration of drugs. Basically, a biguanide that is transported more efficiently by Oct1 will achieve a higher intrahepatic concentration. Therefore, even though the IC50 value for the intrahepatic component, which plays a major role in lactic acidosis, is the same, a biguanide that is transported more efficiently by Oct1 will increase the blood lactate at lower plasma concentrations. Saturation of the hepatic uptake process may prevent increasing the intrahepatic concentration of biguanides to lethal levels. The IC50 values of metformin and phenformin for mitochondrial respiration that were determined with the use of isolated rat mitochondria were 15 mM and 50 μM, respectively (Owen et al., 2000). According to our previous report, theKm values of metformin and phenformin for rOct1 were 377 and 16 μM, respectively (Wang et al., 2002). Taking the key role of Oct1 in the hepatic uptake of metformin into consideration, the Km value determined in rOct1 expressing Chinese hamster ovary cells is very close to theKm values for the uptake of biguanides by hepatocytes. The IC50 value of metformin for mitochondrial respiration is much greater than theKm value for the hepatic uptake process, whereas the IC50 andKm values of phenformin are quite similar. Increasing the plasma concentration of metformin saturates the hepatic uptake process initially, and, therefore, this limits the increase in intrahepatic concentration of metformin. However, in the case of phenformin, the Km value and IC50 values are very similar, and even the concentration that causes saturation of the hepatic uptake process may lead to a significant reduction in oxygen consumption. The EC50 values of phenformin, buformin, and metformin obtained in this study are comparable with those of theirKm values for rOct1 (16, 50, and 377 μM, respectively). Taken from the results of the present study, two conditions need to be satisfied for the development of safer biguanides: 1) low transport activity by Oct1/OCT1, and 2)Km value for hepatic uptake process that is smaller than the IC50 value for mitochondrial respiration.
In conclusion, the Oct1-mediated hepatic uptake of biguanides plays an important role in lactic acidosis. Oxygen consumption and OCT1 cDNA-transfected cells can perhaps be used to evaluate the incidence of lactic acidosis in vivo.
Footnotes
-
This work was supported by Core Research for Evolutional Science and Technology (CREST) of Japan Science and Technology Corporation, Tokyo, Japan.
- Abbreviations:
- Oct1
- organic cation transporter 1
- rOct1
- rat organic cation transporter 1
- AUC
- area under the concentration-time curve
- HPLC
- high-performance liquid chromatography
- Received August 26, 2002.
- Accepted January 7, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}