


Abstract
Brain histamine H3 receptors are predominantly presynaptic and serve an important autoregulatory function for the release of histamine and other neurotransmitters. They have been implicated in a variety of brain functions, including arousal, locomotor activity, thermoregulation, food intake, and memory. The recent cloning of the H3 receptor in our laboratory has made it possible to create a transgenic line of mice devoid of H3 receptors. This paper provides the first description of the H3receptor-deficient mouse (H3 −/−), including molecular and pharmacologic verification of the receptor deletion as well as phenotypic screens. The H3 −/− mice showed a decrease in overall locomotion, wheel-running behavior, and body temperature during the dark phase but maintained normal circadian rhythmicity. H3 −/− mice were insensitive to the wake-promoting effects of the H3 receptor antagonist thioperamide. We also observed a slightly decreased stereotypic response to the dopamine releaser, methamphetamine, and an insensitivity to the amnesic effects of the cholinergic receptor antagonist, scopolamine. These data indicate that the H3receptor-deficient mouse represents a valuable model for studying histaminergic regulation of a variety of behaviors and neurotransmitter systems, including dopamine and acetylcholine.
The neurotransmitter histamine, which originates from tuberomamillary nuclei in the posterior hypothalamus, projects diffusely throughout the central nervous system (CNS) and has been implicated in the regulation of many functions, including sleep/wake, food and water intake, thermoregulation, memory, and other homeostatic processes (Wada et al., 1991; Brown et al., 2001). Four subtypes (H1, H2, H3, and H4) of histamine receptors are currently recognized (Hill et al., 1997; Hough, 2001). The H3 subtype is predominantly located presynaptically and serves as an autoreceptor to regulate the synthesis and release of histamine (Hill et al., 1997). The H3 subtype also has heteroreceptor functions and influences CNS dopamine, γ-aminobutyric acid, noradrenaline, acetylcholine, and serotonin levels (Arrang et al., 1983, 1987b;Schlicker et al., 1988; Clapham and Kilpatrick, 1992; Hill et al., 1997). Behavioral correlates of H3 receptor function have primarily been studied in the context of pharmacologically blocking the receptor using the specific H3 receptor antagonist, thioperamide. For instance, thioperamide has been used to increase the amount of wakefulness (Monti et al., 1991), to prevent scopolamine-induced amnesia (Giovannini et al., 1999), and to decrease food intake (Itoh et al., 1999; Attoub et al., 2001) in rats. The recent cloning of the H3 receptor in our laboratory (Lovenberg et al., 1999) has made it possible to create a transgenic line of mice devoid of H3 receptors and to explore at a molecular level the importance of this receptor in a variety of behaviors. This paper provides the first description of 1) generating the H3 receptor knockout mice, 2) verifying the deletion with radioligand binding and a pharmacologic challenge, and 3) testing the neurochemical and behavioral consequences of deletion of the H3 receptor.
Materials and Methods
Generation of Histamine H3−/− Receptor Animals
Mouse H3R gene clones were isolated from a 129/Ola mouse genomic library, and phage clones covering 13 kilobases of the mouse H3R gene were isolated. TheXhoI DNA fragments containing the second exon of the mouse H3R gene were used to prepare the knockout construct. A cassette containing a neomycin resistance gene was used to replace a 0.7-kilobase region covering part of the first intron and the 5′ end of the second exon of the gene. A HSV-thymidine kinase cassette was placed at the 3′ end of the construct. The mouse H3R gene, the neomycin resistance gene, and the HSV-thymidine kinase gene in the construct are in the same orientation of transcription. The DNA construct was introduced into embryonic day 14 embryonic stem cells by electroporation. Cells were cultured in the presence of 400 μg/ml geneticin (G418) and 0.2 μM ganciclovir. Embryonic stem cells with the disrupted gene were detected by polymerase chain reaction (PCR) and then confirmed by Southern hybridization using a DNA probe flanking the 3′ end of the construct. Chimeric mice were generated from embryos injected with embryonic stem cells. Germline mice were obtained from breeding of chimeric male mice with C57BL/6J females. Germline mice heterozygous for the disrupted H3R gene were identified by PCR. H3R-deficient mice carrying only the disrupted H3R gene were obtained from cross-breeding of heterozygous mice. Wild-type and homozygous animals were produced from the breeding of the germline heterozygotes. These wild-type and homozygous mice were further bred to produce the animals used for this study.
Northern Blot and RT-PCR
Northern Blot Hybridization.
Twenty micrograms of total RNA from H3 +/+ and H3 −/− mouse brains were run on a formaldehyde gel and transferred onto a Nytran nylon membrane (Schleicher & Schuell, Keene, NH). The membrane was prehybridized with hybridization buffer: 50% formamide, 5× sodium chloride/sodium phosphate/EDTA, 5× Denhardt's solution, 0.1% SDS, and 200 μg/ml of single-stranded DNA. The membrane was then hybridized overnight at 42oC with a 32P-labeled probe against mouse H3 exon 3 DNA. As control, a mouse beta actin DNA probe was used in a parallel experiment.
Reverse-Transcriptase PCR.
Total RNA was isolated from H3 −/− and H3 +/+ mouse brains using the TRIzol RNA purification kit (Invitrogen, San Diego, CA). cDNAs were synthesized from total RNA using Superscript II reverse transcriptase (Invitrogen) and random primers as described by the manufacturer. PCR was used to detect the H3 mRNA expression with brain cDNA from either H3 −/−or H3 +/+ mouse as templates, and with P1: 5′-CTCTGCAAGCTGTGGCTGGTGGTAGACTACCTACTGTGTG-3′ and P2: 5′-CTTCTTGTCCCGCGACAGCCGAAAGCGCTGGGTGATGCTT-3′ as primers. The PCRs were performed under conditions of 94°C, 40 s; 65°C, 40 s; 72°C, 2 min for 40 cycles. As control, mouse glycerol-3-phosphate dehydrogenase primers (BD Biosciences Clontech, Palo Alto, CA) were used to amplify glycerol-3-phosphate dehydrogenase cDNA in a parallel PCR reaction. The PCR products were run in a 1.5% agarose gel, stained with ethidium bromide, and visualized under UV light.
Measurement of Histamine H3 and H1 Receptor Binding
Preparation of Membrane Fraction.
Mice were euthanized by gas (CO2) and decapitated immediately. Brains were removed and stored at –80°C until used further. Forebrain tissue was homogenized with a Polytron homogenizer (Kinematica, Lucerne, Switzerland) in ice-cold 50 mM Na/K phosphate buffer, pH 7.5. The homogenates were centrifuged twice at 50,000g for 20 min at 4°C. The resulting pellets were resuspended in binding buffer and kept at 4°C until needed.
The Histamine H3 Binding Assay.
The histamine H3 receptor binding was assayed by a modification of the method of Arrang et al. (1987a). Briefly, to examine inhibition curves, 0.4-ml aliquots of the membrane suspensions (10 mg of tissue) were incubated for 60 min at 25°C with 1.5 nM (R)-[α-3H] methylhistamine. Specific binding was defined as that inhibited by 10 μM thioperamide. The reaction was terminated by addition of 5 ml of the ice-cold buffer and rapid filtration on a glass fiber filter (GF/B) precoated with 0.3% polyethyleneimine. The filters were washed three times with 5-ml volumes of the ice-cold buffer, and the radioactivity trapped on the filters was counted in 10 ml of ScintiVerse (Fisher Scientific, Atlanta, GA).
The histamine H1 binding was assayed by a modification of the method of Tran et al. (1978). In brief, 25 μg of brain membrane was used for the ligand-binding assay with the H1 receptor antagonist, [3H]pyrilamine and with 1 μM quinine to prevent binding to the cytochrome P450-like protein (Liu et al., 1994) (25°C). Concentrations of [3H]pyrilamine were used for Scatchard analysis. Nonspecific binding was determined in the presence of 50 μM triprolidine, an H1 receptor antagonist. The samples were counted as described above.
Measurement of Neurotransmitter Contents
After decapitation, brains were removed rapidly and divided into the cortex and cerebellum. The brain tissues were stored at –80°C until assayed. Brain tissue was homogenized in 3% perchloric acid containing 5 mM disodium EDTA and 5-hydroxy-N ω-methyltryptamine by a Polytron homogenizer (Kinematica) at a maximum setting for 10 s in an ice bath, and the homogenate was centrifuged at 10,000gfor 10 min at 4°C to obtain a clear supernatant. The histamine content was measured fluorometrically with O-pthalaldehyde (Shore et al., 1959) after separation on an HPLC system as described byYamatodani et al. (1985). Briefly, histamine was separated on a cation exchanger, TSK gel SP2SW9 (Tosoh, Tokyo, Japan; particle size 5 μm), eluted with 0.25 M KH2PO4at a flow rate of 0.6 ml/min using a constant flow pump (model CCPM; Tosoh). The histamine eluate was derivatized using an on-line automated Shore's O-pthalaldehyde method (Shore et al., 1959), and the fluorescence intensity was measured at 450 nm with excitation at 360 nm in a spectrofluorometer equipped with a flow cell (model C-R3A; Shimadzu, Kyoto, Japan) and a chromatographic data processor.
Monoamine content (dopamine, homovanillic acid, 3,4-dihydroxyphenylacetic acid, noradrenaline, serotonin, and 5- hydroxyindolacetic acid) in brain samples was measured with an HPLC system with an electrochemical detector (Yanai et al., 1998). They were separated using an HPLC system at 30°C on a reverse-phase analytical column (DS-80TM, 4.6 mm i.d. × 15 cm) and detected by an electrochemical detector (model ECD-100; Eikom Co, Kyoto, Japan). The column was eluted with 0.1 M sodium acetate-citric acid buffer, pH 3.5, containing 15% methanol, 200 mg/l sodium l-octanesulfate, and 5 mg/l disodium EDTA. All measurements were blinded with respect to brain structures and subject group.
Recording of Locomotor Activity, Body Temperature, and EEG/Waking Response to Thioperamide
Electrode and Transducer Implantation.
At 3 months of age, 10 H3 +/+ and 10 H3 −/− male (24–34 g) mice were implanted with chronic electrodes for polygraphic recording of frontoparietal electroencephalogram (EEG) and nuchal electromyogram (EMG) under deep anesthesia (i.p. injection of ketamine/xylazine). The implant consisted of two stainless steel screws (1-mm diameter) inserted through the skull, serving as EEG electrodes, and two insulated wires inserted into the nuchal muscles, serving as EMG electrodes. All electrodes were attached to a microconnector and fixed to the skull with dental acrylic cement. In addition, transducers (PDT-4000 E-Mitter, Mini-Mitter; Minimitter Co., Bend, OR) were inserted through a small incision off midline in the peritoneal cavity for biotelemetric recording of locomotor activity and body temperature. Aftersurgery, mice were individually housed and given 2 weeks to recover from the procedure.
For behavioral tests of locomotor, body temperature, and EEG/EMG activity, it is necessary to individually house animals, which may be considered a stressor. To minimize stress as a confounding variable, animals were carefully adapted to the housing and recording environments before data collection, and each genotype was treated similarly under all experimental conditions.
Baseline Recordings of Locomotor Activity and Body Temperature.
After 2 weeks of recovery from surgery, the mice were transferred to a recording chamber for 2 to 3 days of adaptation followed by 48-h recording of locomotor activity and body temperature. The recording environment was temperature (23–24°C)- and light [12-h/12-h light/dark (LD) cycle]-controlled, with food and water available ad libitum. The biotelemetry transducers were precalibrated to produce radiofrequency signals indicating locomotor counts (number of movements per 10-s time period) and body temperature (accurate to 0.1°C). The transducers were powered by an induction coil, and output signals were detected by a radiofrequency receiver placed under each mouse cage. Data were collected using a software package developed in our laboratory (Multisleep 5.01; Actimetrics, Evanston, IL) for the analysis of EEG/EMG and circadian rhythm measurements.
Thioperamide Challenge.
After 2 weeks of recovery from surgery, H3 +/+(n = 6) and H3 −/− (n = 7) mice were connected to a cable/rotating swivel system for EEG/EMG recording and allowed 1 week of adaptation to a sleep recording chamber. Each mouse was then injected with saline subcutaneously (4 ml/kg body weight) at the onset of the light phase followed by 6 h of EEG/EMG recording. The next day, again at the onset of the light phase, each mouse was given 10 mg/kg thioperamide (dissolved in saline) subcutaneously, and EEG/EMG activity was collected for 6 h. EEG/EMG signals were fed into amplifiers (Grass model 12; Astro-Med Grass Instrument Division, West Warwick, RI). The signals were then digitized and stored on an on-line computer data acquisition program (Multisleep 5.01; Actimetrics Inc., Evanston, IL).
Data Analysis.
Polygraphic recordings were visually scored by 10-s epochs as either wake (W), non-rapid eye movement (NREM) sleep, or rapid eye movement (REM) sleep, according to standard criteria. Briefly, the different vigilance states were characterized as follows: W, low-voltage mixed frequency EEG activity and high EMG activity; NREM, continuous high amplitude low frequency activity in EEG and low EMG activity; and REM, low-voltage fast frequency cortical waves with a regular theta rhythm and absence of muscular tone. Sleep-wake parameters were analyzed over 2-h time intervals. The duration (minutes) of time spent in the different states of vigilance was expressed as a ratio of the total recording session duration (percentage of total recording time). Locomotor activity and body temperature were analyzed over 1-h time intervals, as well as over the 12-h light and dark phases and the total 24-h periods.
Data were analyzed using two-way repeated measures ANOVAs for between (genotype)- and within (time of day)-group factors. Significant interactions were followed up using Newman-Keuls post hoc analyses.
Acute Response of Locomotion and Stereotyped Behavior to Methamphetamine
Locomotion.
H3 +/+ and H3 −/− mice were injected with biological saline [0.9% (w/v) NaCl] or methamphetamine (1mg/kg i.p). Locomotor activity was measured with a photo-beam system (Hamilton-Kinder MotorMonitor System SmartFrame Cage Rack system with two-dimensional 4 × 8 beam; Hamilton-Kinder LLC, Julian, CA). Values for locomotor activity were calculated every 20 min, from 60 min before the injection of vehicle or methamphetamine until 180 min after the injection, and recorded with a personal computer. We used rat cages (9 × 16.93 × 7.87 inches) with a flat top grid that prevented the mice from climbing to the top lid. We used a floor grid with little bedding, which prevented the mice from making nests that could cut the photo-beam. The scores were analyzed by a two-way repeated ANOVA followed by a Student-Newman-Keuls test and a one-way ANOVA followed by Duncan's test.
Stereotyped Behavior.
H3 +/+ and H3 −/− mice were injected with biological saline [0.9% (w/v) NaCl] or methamphetamine (1 mg/kg i.p.). Stereotyped behavior was rated every 20 min, from 60 min before injection of the drug until 180 min after injection, according to the scale described by McLennan and Maier (1983). Scores were defined as follows: 0 = inactive; 1 = intermittent activity; 2 = continuous activity; 3 = intermittent stereotypy; 4 = continuous stereotypy over a wide area including stereotyped locomotor activity, sniffing, and rearing; 5 = continuous stereotypy over a restricted area (mainly sniffing and rearing); 6 = pronounced continuous stereotypy in a restricted area (mainly sniffing); 7 = intermittent licking or biting; and 8 = continuous licking or biting. The scores were analyzed by a two-way repeated ANOVA followed by a Student-Newman-Keuls test and a one-way ANOVA followed by Duncan's test.
Open Field Habituation
Open field habituation was assessed by a modification of the method of Molinengo et al. (1999). In brief, animals were placed individually in an open field apparatus (50 × 50 cm), and their locomotion distance was measured for 5 min using a videotracking system. Animals were tested for three consecutive days in the same activity chambers. To evaluate the effect of repetition of the test, the ambulation ratios (percentages) of the second and third days against the first day in the open field were calculated. Scopolamine (0.75 mg/kg) or vehicle (saline) was given intraperitoneally 30 min before the first and second test.
Wheel Running Activity
H3 +/+ (n= 8) and H3 −/−(n = 11) mice were individually housed in a 33- × 15- × 13-cm polycarbonate cage (Nalge Nunc International, Naperville, IL) equipped with a 12.5-cm-diameter stainless steel exercise wheel. Each wheel revolution triggered a microswitch (Cherry Electric, Pleasant Prairie, WI) mounted on the outside of the cage near the axle of the wheel. The trigger closed an electrical circuit, and these resistance changes indicated the number of wheel revolutions per minute. Data were collected by a DOS PC computer system (Chronobiology Kit; Stanford Software Systems, Stanford, CA). This software generates data output in numeric and graphic form.
Each mouse was recorded under a 12:12-h LD schedule followed by a session in constant darkness (DD). The data represent the number of wheel revolutions during the last 5 days of exposure to LD, before the animals were transferred to DD, and wheel revolutions over five consecutive circadian cycles after they had been exposed to DD for 10 days. Because both genotypes have similar circadian periods, the absolute interval of time for determining the number of wheel revolutions was the same in DD.
LD data were analyzed using two-way repeated measures ANOVA and Newman-Keuls post hoc tests to resolve interaction effects. Data from DD trials were analyzed using an independent-samples Student'st test for genotype comparisons.
Passive Avoidance Test
The apparatus consisted of two compartments, one (9.5 × 18.5 × 16 cm) being surrounded by a white wall and illuminated by a 60-W lamp, and the other (9.5 × 18.5 × 16 cm) being dark and surrounded by a black wall. The compartments were separated by a guillotine door (4.5 × 4.5 cm). All the mice were habituated to the dark chamber for 60 min before the test. On the first day of the passive avoidance test, H3 +/+and H3 −/− mice were divided into two groups. One group of H3 +/+ (n = 14) and H3 −/− (n = 18) mice was injected with scopolamine (0.75 mg/kg i.p., 30 min before the session), whereas the other group of H3 +/+ (n = 13) and H3 −/− (n = 14) mice was injected with biological saline, 0.9% (w/v) NaCl, (1 mg/kg i.p.). The mice were placed into the illuminated safe compartment for 30 s before being given free access to the dark box. The mice tended to escape into the dark compartment. When all four paws were on the grid floor of the dark compartment, a scrambled constant-current foot shock (1 mA, constant voltage 120 V, 50 Hz) was delivered to the grid for 1 s. Then the mice were returned to their home cages. Twenty-four hours later, the procedure, without the electric shock, was repeated. The time that elapsed before each mouse entered the dark compartment was measured. The latency value of 300 s was assigned when animals did not enter the dark compartment within 300 s. The results were analyzed by Student's t test.
Results
Confirmation of Successful Knockout of the Histamine H3Receptor Gene.
Mice lacking the H3 receptor gene (H3 −/−) were created via homologous recombination in embryonic stem cells (129SVJ), and germline chimeras were crossed onto a C57BL/6J background to generate heterozygotes (H3 +/−). H3 +/+ and H3 −/− mice were created by breeding of H3 +/− mice, and germline transmission was determined by polymerase chain reaction. F2 H3 −/− mice were born with an expected mendelian frequency, appeared phenotypically normal, were fertile, and appeared viable through adulthood. Growth curves for H3 +/+ and H3 −/− were parallel with the H3 −/− animals displaying a slightly lower, but not statistically significantly different, average body weight (not shown). The total absence of H3receptors in the transgenic mice was verified by Northern blot, RT-PCR, and radioligand binding studies (Fig 1). Whereas H3 receptors can be readily detected via radioligand binding in normal (H3 +/+) mouse brain homogenates, H3 −/− mice demonstrated a complete loss of H3 receptor binding sites as determined by (R)-[α-3H]methylhistamine binding (Fig. 1B). Heterozygous mice (H3 +/−) had the same (R)-[α-3H] methylhistamine binding affinity (0.43 nM) as the H3 +/+mice (0.47 nM), but only about half the number of binding sites in whole brain homogenates (47 fmol/mg of protein versus 91 fmol/mg of protein, respectively).
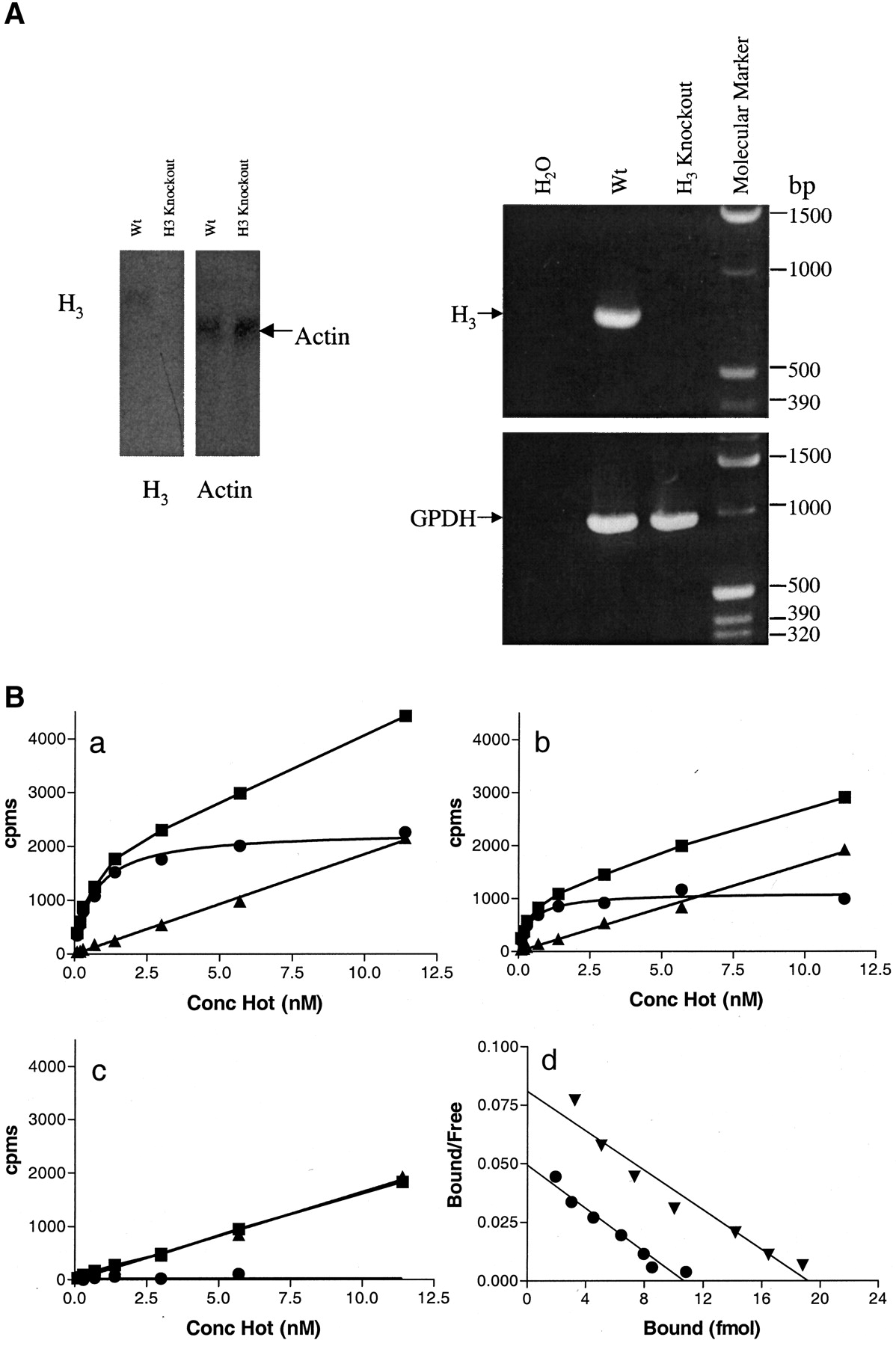
Absence of histamine H3 mRNA and (R)-[α-3H] methylhistamine binding in brains from H3 −/− mice. A, left, northern blot. Total RNA (20 μg) from H3 +/+ and H3 −/− mouse brains were transferred onto a Nytran nylon membrane and probed with a 32P-labeled probe against mouse H3 exon 3 DNA. As control, a mouse actin DNA probe was used in a parallel experiment. Right, reverse-transcriptase PCR. cDNA was synthesized from total RNA prepared from brains of H3 +/+ and H3 −/− mice and used as template in a PCR reaction. The PCR products were run on a 1.5 % agarose gel and detected under UV light. As controls, mouse GDPH primers were used in a parallel PCR reaction. B, receptor binding of (R)-[α-3H] methylhistamine to membranes prepared from H3 −/− (a), H3 +/− (b), or H3 −/−(c) mice. ▪, total binding; ●, specific binding; ▴, nonspecific binding. Counts bound (cpms) are plotted on the y-axis, and the concentration of radiolabeled (R)-[α-3H] methylhistamine is given on the x-axis. Whole brains (minus the cerebellum) were homogenized, and equal amounts of the resulting membrane preparation were included in each assay. d: Scatchard plots for the H3 +/+ mice (▴) and H3 −/− mice (●).
H3−/− Mice Have Normal Brain Levels of Dopamine, Norepinephrine and Serotonin, but Decreased Levels of Histamine.
We compared the brain levels of several neurotransmitters in H3 +/+ and H3 −/− mice. Neurotransmitter content of the cerebral cortex was measured and no significant differences were found for dopamine (H3 +/+ = 1.79 ± 0.28 nmol/g versus H3 −/− = 1.59 ± 0.16 nmol/g), norepinephrine (H3 +/+ = 0.37 ± 0.04 nmol/g versus H3 −/− = 0.37 ± 0.04 nmol/g), or serotonin (H3 +/+ = 0.57 ± 0.13 nmol/g versus H3 −/− = 0.47 ± 0.11 nmol/g), or any of their metabolites (not shown). However, significant differences were observed for cortical histamine content (H3 +/+ = 266.8 ± 45.1 pmol/g versus H3−/− = 156.1 ± 35.5 pmol/g, p <0.05).
H3−/− Mice Have Decreased Spontaneous Locomotor Activity and Wheel Running Behavior.
To assess the effects of the H3 receptor on circadian rhythmicity and activity levels, both total locomotor activity and wheel running were evaluated. Significant interactions between genotype and time of day were detected for locomotor activity (day 1,F = 2.33(1,23), p < 0.001; day 2,F = 2.14(1,23), p < 0.01) and body temperature (day 1, F = 1.95(1,23), p< 0.01; day 2, F = 2.11(1,23), p < 0.01). The results of follow-up tests showed that the H3 −/− mice had markedly decreased locomotor activity during the dark phase of the circadian cycle (Fig 2a), and this was reflected by an impaired temperature elevation during the dark phase (Fig. 2B). Although there was no significant difference in the number of wheel running revolutions during the light phase, the significant dark phase effect (F = 4.33(1,17), p <0.05) resulted in an overall 22% decrease in wheel running behavior over the 24-hr light/dark cycle in the H3 −/−mice (Fig. 2C). This overall decrease in running wheel behavior persisted in continuous darkness where on average there was about a 24% decrease in the number of wheel revolutions over the circadian cycle in the H3 −/− mice (t = 2.32, df = 14, p < 0.05) (Fig. 2C). It is interesting to note that whereas the amplitude of the changes in locomotor activity and body temperature was blunted, circadian rhythmicity was maintained. We found no difference in either the phase angle of entrainment of the activity rhythm to the light/dark cycle, nor were there any differences in the free running period of the activity rhythm (period = 23.6 ± 0.1 h in H3 +/+ animals and 23.5 ± 0.1 h in H3 −/− mice).

Histamine H3 receptor knockout mice have decreased locomotor activity (A), body temperature (B), and wheel running behavior (C). A and B, mean (±S.E.M.) total activity counts (A) and body temperature (B) over 48 h in H3 +/+ (n = 10) and H3 −/− (n = 10) mice. Significant differences occurred between genotypes in the 12-h period of darkness (designated on the x-axis by black bars). Asterisks represent significant post hoc comparisons of genotypes at individual 1-h intervals (★, p < 0.05). C, mean (±S.E.M.) wheel revolutions in H3 +/+(n = 8) and H3 −/−(n = 11) mice during a 24-h (12:12-h LD) period (left three columns) and for a circadian cycle in constant darkness (DD) (right column). H3 −/− mice had significantly reduced wheel running activity during the LD (dark phase) and DD schedules compared with H3 +/+ mice. ∗,p <0.05, ∗, p <0.01
The H3−/− Mice Are Insensitive to the Wake-Promoting Effect of Thioperamide.
To test the role of the H3 receptor in mediating arousal states, we compared the wake-promoting effects of the H3receptor antagonist, thioperamide in H3 +/+ and H3 −/− mice. A significant interaction between genotype and drug occurred for percentages of wakefulness (F = 29.59(1,11), p < 0.001) and NREM sleep (F = 32.63(1,11),p < 0.001). Follow-up testing (Fig3) showed that thioperamide increased waking by 55% during the first 2 h after administration (at lights-on) in H3 +/+ mice. This increase in waking was associated with a 61% decrease in NREM sleep during the 2-h postinjection period, whereas REM sleep was not affected (Fig. 3). Importantly, there were no effects of thioperamide on the sleep-wake status of the H3 −/−mice, providing behavioral confirmation that the receptor had been deleted, and verifying that this behavioral effect of thioperamide is indeed mediated through the H3 receptor.

H3 receptor knockout animals are insensitive to the histamine H3 receptor antagonist thioperamide. Mean (±S.E.M.) percentage of recording time spent in each of the three vigilance states (wake, NREM, and REM sleep) during the first 2 h of the light phase in both H3 +/+ (n = 6) and H3 −/− (n = 7) mice. The open bars represent vehicle injections on baseline days, whereas the black bars represent thioperamide (10 mg/kg s.c.) injections given to the same animals the following day. Injections were performed at the onset of the light phase. ∗∗∗, p < 0.001
The H3−/− Mice Show a Decreased Sensitivity to Methamphetamine.
We examined the response of the H3 −/− mice to methamphetamine (1 mg/kg), which is known to increase locomotor activity and to induce stereotypic behavior, in part, by increasing dopamine release (Grilly and Loveland, 2001). After injections of methamphetamine, the decline of the stereotypy scores was faster in H3 −/− compared with H3 +/+ mice [F = 31.348(3,24), p < 0.001]. One-way ANOVA followed by Duncan's test at every time point showed that methamphetamine-injected H3 −/− mice had significantly lower stereotypy scores than H3 +/+ mice between 60 and 160 min (Fig. 4A). Likewise, there was an overall difference in the methamphetamine-induced locomotion between the H3 +/+ and H3 −/− mice [F= 4.051(3,24), p < 0.001]. A one-way ANOVA followed by Duncan's test indicated that the ambulation of methamphetamine-injected H3 −/− mice was significantly lower than that of H3 +/+ mice between 80 and 120 min after the injection. These data indicate that H3 −/− mice recover faster from methamphetamine-induced locomotion and stereotypy than H3 +/+ mice.

H3 receptor knockout mice exhibit decreased sensitivity to methamphetamine (A) and scopolamine (B). Means (±S.E.M.) of 7 saline-treated and 16 methamphetamine (MAP)-treated H3 +/+ mice, and 8 saline-treated and 26 MAP-treated H3 −/− mice are shown. Data were collected from 60 min before and 180 min after injections. A, left, rating of stereotyped behavior in response to MAP (1 mg/kg) treatment. Right, ambulation of mice treated with saline or MAP (1 mg/kg). ★, p < 0.05; ★★, p < 0.01; ★★★, p < 0.001. B, passive avoidance response. Left: mean (±S.E.M.) step-through latencies in untreated H3 +/+ (n = 13) and H3 −/− (n = 14) mice. Right, effect of scopolamine (0.75 mg/kg) on step-through latencies in H3 +/+ mice (n = 14) and H3 −/− mice (n= 18). The mice received scopolamine intraperitoneally 20 min before the first test in the passive avoidance chamber. No injection was given beforetesting on the second day. ∗∗∗, p < 0.001
The H3−/− Mice Are Resistant to the Amnesic Effect of Scopolamine in the Passive Avoidance Test.
The effect of deletion of the H3 receptor on memory function was investigated using the step-through passive avoidance test. This test uses a light/dark preference and an acute aversive conditioning stimulus (mild foot shock), and has been used to demonstrate cognitive/memory enhancement of cholinesterase inhibitors. In a basic light/dark distribution test, there was no difference between H3 +/+ and H3 −/− mice either in time spent in the light or dark compartments or in the number of transitions from light to dark compartment (data not shown). There was no difference between H3 +/+ and H3 −/− mice in the basic passive avoidance test as both sets of mice were equally able to retain the recollection of the aversive stimulus (Fig. 4B). When the H3 +/+ mice were pretreated with the amnesic agent scopolamine (a muscarinic receptor antagonist) before their first exposure to the chamber, they failed to recall the aversive stimulus upon reintroduction to the chamber on the next day (Fig. 4B, right). However, the H3 −/−mice were completely unresponsive to the amnesic effects of scopolamine and responded similarly to the untreated group (Fig. 4B, left). We also tested the animals in the open field habituation test and found that the H3 −/−out animals displayed normal learning. However, in contrast to the results in the passive avoidance test, the analgesia-inducing effect of scopolamine was not decreased in H3 −/− animals in the open field habituation test (not shown).
Discussion
In the central nervous system, histaminergic neurons are found only in the tuberomammillary nucleus of the posterior hypothalamus. These cells project to numerous brain regions (Brown et al., 2001) and are involved in many brain functions. The histamine H3 receptor, a predominantly presynaptic inhibitory receptor, is involved in a number of physiological processes, including waking behavior and memory (Brown et al., 2001). The elucidation of its function has relied heavily on the use of H3 receptor-specific ligands. The cloning of the H3 receptor (Lovenberg et al., 1999) allowed a more molecular approach, including the production of knockout animals. In this report we describe some of the biochemical, behavioral, and pharmacological characteristics of the H3receptor knockout mouse.
We confirmed the absence of H3 receptor in the brain of the knockout mice by RT-PCR, Northern blot, and binding studies. Because it has been suggested that the H3 receptor may exist in several subtypes (West et al., 1990), binding studies using (R)-[α-3H]methylhistamine, [N-α-3H]methylhistamine, or [3H]histamine were performed in mouse brain homogenates. No binding could be detected even at high (>100 nM) concentrations, which indicates that all H3binding is due to a single gene product and argues against the existence of H3 receptor subtypes (data not shown). This also demonstrates that in whole brain homogenates, the recently identified H4 receptor (Nakamura et al., 2000; Liu et al., 2001) does not contribute to the observed binding of [3H]histamine. This is consistent with the lack of apparent distribution of H4 receptor messenger RNA in brain tissue (Liu et al., 2001).
Histamine receptors play a complex role in the regulation of brain levels of specific neurotransmitters. For instance, in H1 receptor knockout mice, the serotonin turnover rate is increased in specific brain regions (Yanai et al., 1998). It is known that stimulation of the H3 receptor increases the synthesis of histamine (Arrang et al., 1987b;Gomez-Ramirez et al., 2002). Also, Yates et al. (1999) reported that thioperamide, a histamine H3 receptor antagonist, enhanced the histamine turnover rate in rats. Therefore, we decided to measure brain levels of a number of neurotransmitters and their metabolites. The only change we found was a clear decrease in the levels of histamine in the cortex of H3 −/− animals, which we attribute to the removal of the stimulatory effect of the H3 receptor on the synthesis of histamine.
The presynaptic autoregulatory H3 receptor inhibits the neuronal release of histamine, which in turns leads to a decreased stimulation of postsynaptic H1receptors (Brown et al., 2001). The role of the central H1 receptor in arousal is extensively documented and underscored by the well known sedative effects of many H1 receptor antagonists (Kay, 2000). Thus, the absence of H3 receptors might be expected to promote histaminergic neurotransmission and thereby increase arousal in general. This was clearly not the case: the animals showed a decreased level of motor activity throughout the night (their active period) and decreased wheel running behavior that persisted during continuous darkness. There are several possible explications for this. First, the histamine H1 receptor might be down-regulated in the H3 −/− mice to compensate for the increased release of histamine at the synapse. We rejected this hypothesis after H1 receptor binding experiments showed that H3 −/− mice had comparable densities of H1 receptors compared with H3 +/+ mice (data not shown). Second, the removal of the H3 receptor may have led to perturbations in the homeostasis of neurotransmitter levels, an explanation that seems to be borne out by our results showing lower brain levels of histamine in H3 −/− animals. This leads to the interesting hypothesis that although the braking effect on histamine release exerted by the H3 receptor is removed in the H3 −/− mice, a compensatory decrease in the availability of histamine in the nerve terminals may lead to an overall reduction of histaminergic neurotransmission with reduced stimulation of H1receptors and decreased locomotion as a consequence. This hypothesis could be approached by experiments measuring the release of histamine from brain slices derived from H3 −/− mice and by in vivo microdialysis. Alternatively, histaminergic neurons may have diminished histamine content as a result of the lack of inhibition of the H3 receptor in its absence throughout the development of the animal. A careful study of developmental histamine turnover rates in H3 receptor-deficient animals may help address this question.
Third, it must be pointed out that the influence of the histaminergic system on locomotor activity is complex. For instance, the H1 knockout mice, which would be expected to show decreased locomotion throughout the light/dark cycle, actually displayed an increase in locomotor activity during the light phase (Inoue et al., 1996). In the present study, we saw no increase in activity during the light phase in H3 −/− mice, indicating that the components of the arousal system that are affected by knocking out the H1 and H3 receptors are different from one another. Finally, the mouse model described here is not a conditional knockout: the continuous absence of the H3 receptor throughout the development of the animals may be compensated for by changes in other genes.
One of the most significant functions of the H3receptor seems to be its role in the regulation of waking behavior. Administration of H3 receptor antagonists, such as thioperamide, increase wakefulness at the expense of REM and NREM sleep in rats (4 mg/kg i.p.) (Monti et al., 1991) and cats (2–10 mg/kg p.o.) (Lin et al., 1990). Thioperamide is a selective H3 antagonist with aK i of 4.2 nM at the rat H3 receptor (Lovenberg et al., 2001). It is active in vivo at doses between 2 and 20 mg/kg i.p.(Stark et al., 1996). We therefore tested the effect of this compound (10 mg/kg s.c.) in H3 −/− mice. The H3 +/+ animals responded to H3 receptor blockade with a decrease in NREM sleep and increased waking, whereas the H3 −/− animals were completely insensitive to the wake-promoting effects of thioperamide. These experiments confirm the important role of the H3receptor as a mediator of wakefulness.
As mentioned earlier, the presynaptic H3 receptor regulates the release of histamine and other neurotransmitters such as dopamine and acetylcholine (Hill et al., 1997). We therefore decided to investigate how these neurotransmitter systems were affected by ablation of the H3 receptor. We probed dopaminergic neurotransmission by using methamphetamine, which increases locomotor activity and induces stereotypic behavior through an increase in dopamine release (Grilly and Loveland, 2001). We found that the H3 −/− mice scored lower on a stereotypy scale after methamphetamine administration than did the H3 +/+ animals. The effect of methamphetamine on locomotor activity was less pronounced in the H3 −/− mice compared with H3 +/+ animals as well. These results may indicate that the H3 −/− mice have slightly decreased dopaminergic activity, which may contribute to the overall spontaneous decrease in total activity and wheel running behavior observed in these animals. Our observations are in agreement with those of Clapham and Kilpatrick (1994), who found that the H3 receptor antagonist thioperamide decreased amphetamine-induced locomotor activity in the mouse. However, because methamphetamine has several mechanisms of action, including serotonergic pathways, additional experiments with compounds selective for the various dopamine receptor subtypes will be needed to evaluate dopaminergic neurotransmission in more detail.
One of the most active areas of research in the H3 receptor field is the study of memory. For instance, Molinengo et al. (1999) showed that the effects of thioperamide on memory consolidation seemed dependent on the situation, with or without painful stress. Also, Passani et al. (2000) indicated that thioperamide did not improve memory in normal mice, but only in a learning deficit situation. Because blockade of the presynaptic H3 receptor leads to an increased release of acetylcholine (Hill et al., 1997), models exploring the role of the cholinergic system in memory formation are particularly interesting. It is known, for instance, that the acetylcholine receptor antagonist, scopolamine, induces amnesia in the passive avoidance test and that H3 receptor antagonists are able to prevent this effect (Giovannini et al., 1999). We found, using the passive-avoidance model, that H3 −/− mice showed normal learning behavior but were insensitive to the amnesia-inducing effects of scopolamine. This is consistent with previous reports showing that thioperamide could at least partially prevent the effects of scopolamine (Blandina et al., 1996; Onodera et al., 1998; Molinengo et al., 1999).
Interestingly, the H3 −/− mice were not insensitive to scopolamine in a second model, the open field habituation test. This test measures the habituation of exploratory activity and is a valid model of memory (Platel and Porsolt, 1982; Izquierdo et al., 1990). The fact that the H3 −/− mice were insensitive to the effect of scopolamine only when it was used in an aversion model may indicate a specific role of the H3 receptor in the memory processes associated with painful stimuli.
Histamine, as a neurotransmitter, has been implicated in the regulation of many peripheral and CNS functions. However, the precise role that histamine and its receptors play in CNS physiology is not entirely clear. The present study in mice devoid of the H3receptor indicate an important role for the H3receptor in the regulation of locomotor activity and body temperature. In addition, H3 −/− mice are resistant to the amnesic effect of the cholinergic antagonist, scopolamine, and show a decreased response to the dopaminergic-stimulating drug, methamphetamine. Because the H3 receptor has also been implicated in the regulation of a wide range of other physiological and behavioral processes, including food intake, digestion, cardiac and immune functions, cognition, and sleep, this new transgenic animal model should prove to be extremely important for elucidating the role of H3 receptors in a variety of peripheral and CNS functions as well as pathophysiological states that are associated with altered histaminergic activity.
Footnotes
- Received February 28, 2002.
- Accepted May 17, 2002.
-
This research was supported in part by National Institutes of Health Grants AG18200 and AL59598.
-
H.T. and C.D. contributed equally to this study.
Abbreviations
- CNS
- central nervous system
- PCR
- polymerase chain reaction
- RT-PCR
- reverse-transcriptase polymerase chain reaction
- HPLC
- high performance liquid chromatography
- LD
- light/dark
- EEG
- electroencephalogram
- EMG
- electromyogram
- NREM
- non-rapid eye movement
- REM
- rapid eye movement
- ANOVA
- analysis of variance
- DD
- constant darkness
- MAP
- methamphetamine
- U.S. Government





{kind=link}
{kind=link}
{kind=link}
{kind=link}