


Abstract
The relationship between receptor number and agonist-induced intracellular responses has been well studied in receptors coupled to adenylate cyclase; however, for receptors coupled to phospholipase C (PLC), very little is known about the effect of receptor number on receptor-mediated processes. To explore this issue, we investigated the effect of the number of receptors for gastrin-releasing peptide (GRP) on ligand affinity and on the ability to activate intracellular messengers [PLC, tyrosine phosphorylation of p125 focal adhesion kinase (p125FAK)] and cause receptor modulation (internalization, desensitization, down-regulation) and ligand degradation. Three BALB 3T3 cell lines were made that stably expressed the gastrin-releasing peptide receptor (GRP-R) with receptor numbers varying by 280-fold (GRP-R-Low, GRP-R-Med, and GRP-R-Hi). Each cell line had the same affinity for agonist. The efficacy for bombesin to increase [3H]inositol phosphates but not tyrosine phosphorylation of p125FAK correlated well with receptor number. In contrast, the EC50 value for [3H]inositol phosphate generation for bombesin was the same in each cell line. Receptor number did not alter internalization. In the absence of protease inhibitors, there was an inverse correlation between receptor number and receptor down-regulation and desensitization. However, with protease inhibitors present, GRP-R-Med and GRP-R-Hi down-regulated significantly less than the GRP-R-Low. Similarly, GRP-R-Low desensitized significantly more than GRP-R-Med or GRP-R-Hi. GRP-R-Hi caused significantly greater ligand degradation than GRP-R-Low, and protease inhibitors completely inhibited degradation by GRP-R-Low and inhibited degradation by 70% for GRP-R-Hi. In conclusion, we show that for the PLC-coupled GRP-R, receptor number had little or no effect on binding affinity, potency for activating PLC, tyrosine phosphorylation of p125FAK, or extent of receptor internalization. In contrast, receptor number had an effect on ligand degradation, down-regulation, desensitization, and efficacy of PLC activation without altering the efficacy of tyrosine phosphorylation of p125FAK. These results demonstrate that the effect of receptor number differs for the different functions mediated by the GRP receptor and differs from that reported for adenylate cyclase-coupled receptors such as receptors mediating the action of adrenergic agents, secretin, and opioids.
The mammalian bombesin-related peptides, GRP and neuromedin B, have effects in many diverse cell systems (1), including the central nervous system (e.g., regulation of circadian rhythm, thermoregulation, satiety), the gastrointestinal tract (1) [e.g., release of numerous gastrointestinal hormones, gallbladder and smooth muscle contraction, pancreatic secretion (2)], and the immune system (1) (e.g., stimulation of chemotaxis, natural killer cell activity, antibody-dependent cellular cytotoxicity), and developmental effects (1) (e.g., normal lung maturation, stimulation of fetal chondrocytes). GRP is one of the most potent neuropeptides modulating growth effects (1, 3). GRP stimulates the growth of both normal tissue (3) (e.g., bronchial endothelial cells, uterine stromal cells, Swiss 3T3 cells) and many different tumor cells (3, 4) (e.g., human small-cell lung cancer cells, breast and prostate cancers). In some cells, such as human small-cell lung cancer cells, GRP-related peptides have an autocrine growth function (4). Pharmacological and cloning studies have demonstrated that the actions of GRP are mediated by the GRP-R, a member of the seven-transmembrane G protein-linked receptor superfamily (5). These receptors are coupled to PLC in all tissues examined (1-3) through heterotrimeric G proteins, and activation results in an increase in IP, mobilization of intracellular Ca2+, and activation of protein kinase C. Agonist activation of the GRP-R results in tyrosine phosphorylation of a number of proteins such as p125FAK and paxillin (6) and causes the receptor to undergo numerous modulatory processes, such as internalization, down-regulation (7, 8), and desensitization (7, 8).
A number of studies have proposed that either the acquisition of GRP-Rs by tissues or the autocrine growth effect of GRP on various tissues may be important in the growth and perhaps pathogenesis of a number of neoplasms (4, 9, 10). In normal and tumor cells, the number of GRP-Rs expressed have been shown to vary greatly (2, 11-13). This finding becomes potentially very significant in light of recent studies that demonstrate that alterations in receptor down-regulation or desensitization can have important regulatory effects on cell growth and cell cycle progression (14, 15). The relationship between receptor number and agonist-induced second messenger responses has been well studied for a number of receptors coupled to adenylate cyclase, including receptors for β-adrenergic agents (16-22), secretin (23), δ-opioid (24), m2 muscarinic cholinergic (25), 5-HT1A(26), 5-HT1B (27), luteinizing hormone, and vasopressin (20). In contrast to receptors coupled to adenylate cyclase, there have been few studies of the effect of receptor number on activation of second messengers or binding parameters for receptors preferentially coupled to PLC such as GRP-R (28). Furthermore, no studies for any receptor have examined the effect of receptor number on ligand degradation or on such receptor modulatory processes as internalization, and only a few studies of adenylate cyclase-coupled receptors have investigated the relationship between receptor number and receptor down-regulation and desensitization (23, 24). The GRP-R transfected into BALB 3T3 cells is an excellent system in which to investigate the effect of receptor number on these cellular processes for a PLC-coupled receptor because a recent study shows that GRP-R transfected into BALB 3T3 cells behave in an identical fashion to native receptors in Swiss 3T3 cells in terms of binding, generation of [3H]IP, and receptor modulation (i.e., internalization, down-regulation, and desensitization) (7).
To address the question of the relationship of receptor number to receptor coupling to various intracellular pathways (PLC, tyrosine phosphorylation of p125FAK) and receptor modulation (internalization, down-regulation, desensitization), we compared GRP-R-transfected BALB 3T3 cell lines with different receptor numbers, ranging from approximately the same number as on native Swiss 3T3 cells to the same cells containing one half of this number or 150-fold more than this number.
Experimental Procedures
Materials
[Tyr4]Bombesin and GRP were obtained from Peninsula Laboratories (Belmont, CA). Anti-p125FAK antibody and anti-phosphotyrosine mAb clone PY20 were from Transduction Laboratories (Lexington, KY). Recombinant protein A-agarose was from Upstate Biotechnology (Lake Placid, NY). 12-O-Tetradecanoylphorbol-13-acetate and deoxycholic acid were from Calbiochem (La Jolla, CA). PBS, pH 7.4, was from Biofluids (Rockville, MD). DMEM, fetal bovine serum, and the aminoglycoside G-418 were from GIBCO (Waltham, MA). Tris and glycine were from Schwarz/Mann Biotech (Cleveland, OH). EDTA, bacitracin, SBTI, dimethylsulfoxide, and Triton X-100 from Sigma Chemical (St. Louis, MO). Phenylmethylsulfonyl fluoride was from Fluka Chemical (Ronkonkoma, NY). bovine serum albumin fraction V was from Miles (Kankakee, IL). Aprotinin, leupeptin, and HEPES were from Boehringer-Mannheim Biochemicals (Indianapolis, IN). Goat anti-mouse IgG and IODO-GEN were from Pierce Chemical (Rockford, IL). Dowex AG1-X8 anion exchange resin (100–200 mesh formate form), SDS, 2-mercaptoethanol, and protein assay dye reagent were from BioRad (Richmond, CA). Hyperfilm ECL, enhanced chemiluminescence detection reagents, and Na125I were from Amersham (Arlington Heights, IL). myo-[2-3H]inositol (10–20 Ci/mmol) was from DuPont-New England Nuclear (Boston, MA). Hydrofluor scintillation fluid was from National Diagnostics (Atlanta, GA). BALB 3T3 fibroblasts were obtained from the American Type Culture Collection (Rockville, MD). Nitrocellulose membrane was from Schleicher & Schuell (Keene, NH). Standard buffer consisted of 98 mm NaCl, 6 mmKCl, 25 mm HEPES, 5 mm pyruvate, 5 mm fumarate, 5 mm glutamate, and 0.1% SBTI.
Methods
Growth and selection of GRP-R-transfected BALB 3T3 cell lines.
BALB 3T3 cells devoid of GRP-R were selected by clonal expression after assaying for GRP-R by RNase protection and binding studies. A mouse GRP-R clone was generated from Swiss 3T3 cells as described previously (7). After an EcoRI digest, a full-length GRP-R cDNA was isolated and ligated into a modified version of the mammalian expression vector pCD2 (7). A BALB 3T3 cell line that contained approximately the same number of receptors as native Swiss 3T3 cells (GRP-R-Med) and a cell line that contained fewer receptors than this cell line (GRP-R-Low) were constructed and identified as outlined below. A BALB 3T3 cell line expressing GRP-R has been previously described (29) that possesses >106receptors/cell and was characterized fully; in the current study, it was used for the GRP-R-Hi. This cell line contained a mycepitope at the 5′ end of the GRP-R, which has been shown previously not to affect cell function (29).
As described previously (7), stably transfected BALB 3T3 cells expressing GRP-Rs were generated by calcium phosphate precipitation. Selection with G-418 (800 μg/ml) was begun 48 hr after transfection and continued for 2–3 weeks. Clonal cell lines were screened for125I-[Tyr4]bombesin binding, and cells expressing saturable binding were further analyzed for receptor number using the least-squares curve-fitting program LIGAND. Cells with a moderate (1–3 × 105 receptors/cell) and a small number (<1 × 105 receptors/cell) of GRP-R were identified and characterized by binding studies and studies of [3H]IP generation. Each of the GRP-R-Med and GRP-R-Low showed similar results. One cell line containing a moderate receptor number (GRP-R-Med) (WT-5) and another cell line containing a low receptor number (GRP-R-Low) (WT-8) were selected for detailed characterization. Cells were maintained in DMEM containing 10% fetal bovine serum plus 280 μg/ml G-418 and passaged every 3–4 days at confluence using 0.1% trypsin in 1 mm EDTA. All cell lines were cultured at 37° in a 5% CO2 atmosphere.
Binding of 125I-[Tyr4]bombesin to GRP-R-transfected cells.
125I-[Tyr4]Bombesin (2200 Ci/mmol) was prepared as described previously (30). Binding studies using the GRP-R-transfected cells were performed as described previously (7). Briefly, disaggregated cells were suspended in binding buffer [standard buffer plus 1 mm MgCl2, 2.2 mm KH2PO4, 2 mmglutamine, 11 mm glucose, 0.1% (w/v) bacitracin, and 0.2% (w/v) bovine serum albumin, pH 7.4]. GRP-R-Hi cells were suspended at a concentration of 1 × 105 cells/ml, whereas other cells were suspended at a concentration of 3–10 × 106 cells/ml. Incubations contained 75 pm 125I-[Tyr4]bombesin and proceeded for 45 min at 22° unless otherwise stated. At the end of the incubation, 100 μl of cell suspension was centrifuged at 10,000 × g for 10 sec to separate bound from free ligand. Nonsaturable binding of125I-[Tyr4]bombesin was the amount of radioactivity associated with GRP-R-transfected cells when the incubation mixture contained 1 μm bombesin. Nonsaturable binding was <15% of total binding in all experiments, and all values are reported as saturable binding (i.e., total minus nonsaturable binding).
GRP-R down-regulation.
Down-regulation was performed as described previously (7). Briefly, cells were split 1:4 and 24 hr later were washed once in PBS. One fourth of the cells were resuspended in DMEM containing 6 nm bombesin for ≤ 24 hr, whereas the remaining cells were resuspended in DMEM alone. After incubation with or without 6 nm bombesin, cells were washed, and down-regulation was measured by assessing the binding of125I-[Tyr4]bombesin to determine cell number as described previously (7). Analysis of the binding data was performed using LIGAND, which permitted comparisons in mathematically derived receptor number (B max) and affinity (Ki ) between bombesin-pretreated cells and control cells as described previously (7, 30). Down-regulation was expressed as the percentage of the receptor number present on bombesin-pretreated cells compared with the untreated cells that were processed in parallel as described previously (7).
GRP-R internalization.
Internalization was performed using125I-[Tyr4]bombesin as described previously (7). Briefly, confluent cells were mechanically disaggregated, washed, and suspended at a concentration of 1 × 105 cells/ml in GRP-R-Hi cells and 3- 10 × 106 cells/ml in GRP-R-Med and GRP-R-Low cells in binding buffer with 0.1% bacitracin. Cells were incubated with 75 pm 125I-[Tyr4]bombesin for various times at 37°. After incubation, 100 μl samples were diluted 20-fold with 0.2m acetic acid, pH 2.5, in 0.5 m NaCl and incubated for 5 min at 4° to remove surface radioligand, as described previously (30). In all cases, parallel incubations were conducted in the presence of 1 μm unlabeled bombesin to determine changes in nonsaturable binding. Results were expressed as the percentage of saturable 125I-[Tyr4]bombesin added that is internalized (i.e., not removed by acid treatment).
Cell membrane preparation.
Cells were grown to confluence, mechanically disaggregated, washed twice with PBS at 4°, and resuspended in 10 ml of homogenization buffer (50 mmTris·HCl, pH 7.4, 0.2 mg/ml SBTI, 0.2 mg/ml benzamidine) as described previously (30). Cells were homogenized using a Polytron homogenizer (Brinkmann Instruments, Westbury, NY) for 30 sec with a power level 6 at 4°. The homogenate was centrifuged for 5 min at 500 ×g at 4°. The supernatant was centrifuged at 30,000 ×g for 30 min at 4°. The pellet was resuspended in membrane binding buffer [10 mm HEPES, pH 7.4, 118 mmNaCl, 4.7 mm KCl, 5 mm MgCl2, 1 mm EGTA, 0.2% (w/v) bovine serum albumin, 0.2 mg/ml benzamidine, 0.2 mg/ml SBTI, 1 mg/ml bacitracin] at a 1.0 mg of protein/ml concentration.
Binding of 125I-[Tyr4]bombesin to cell membranes.
Binding of 125I-[Tyr4]bombesin to cell membranes was performed as described previously (30). Briefly, membranes were diluted with membrane binding buffer to a protein concentration of 0.1–0.25 mg/ml for GRP-R-Low and GRP-R-Med cells and 0.0075 mg/ml for GRP-R-Hi cells. In a typical experiment, a 300-μl aliquot was incubated with 75 pm 125I-[Tyr4]bombesin in the presence or absence of various agents at 22° for 30 min. Duplicated 100-μl samples were layered over 300 μl of PBS (4°) in 500-μl polypropylene tubes and centrifuged at 10,000 × g for 3 min (Beckman Model B microfuge). After aspiration of the supernatant, the pellet were washed once with 300 μl of PBS (4°). The tops of the tubes were cut, and radioactivity associated with membrane was measured with a γ-counter. In all cases parallel, incubations in the presence of 1 μm unlabeled bombesin to determine nonsaturable binding were performed. Nonsaturable binding was always < 20% of the total binding.
Measurement of [3H]IP and desensitization studies.
Changes in [3H]IP and desensitization studies were performed as described previously (7). Briefly, cells were plated onto 24-well dishes at 2.0 × 105 cells/well in DMEM containing 10% fetal bovine serum and grown for 24 hr and then loaded with 3 μCi/ml myo-[2-3H]inositol in DMEM with 2% serum for 24 hr. Cells were washed and incubated for 15 min with IP buffer (standard buffer plus 10 mm LiCl, 2 mm CaCl2, 2% bovine serum albumin, and 1.2 mm MgSO4) and then incubated with varying concentrations of bombesin for 60 min at 37°. Reactions were halted with 1% HCl in methanol. Total [3H]IP were isolated by anion exchange chromatography as described previously (7). For the desensitization study, cells were plated onto 24-well dishes at 1.0 × 105 cells/well in DMEM containing 10% fetal bovine serum and grown for 24 hr and then loaded with 3 μCi/mlmyo-[2-3H]IP in DMEM without serum for 24 hr. Cells were washed with IP buffer and preincubated for 10 min with IP buffer and then incubated with varying concentrations of bombesin for 45 min at 37°. Desensitization was measured as described previously (7) by treating cells with either no additions or with 6 nmbombesin in parallel as described for down-regulation. The residual ability to increase cellular IP in cells preincubated with 6 nm bombesin was determined by incubating cells with 1 μm bombesin and expressing the maximal increase as the percentage of that caused by control cells processed in parallel (7).
Degradation of radioligands.
Each of the different GRP-R-transfected BALB 3T3 cell lines, at a concentration of 6.5 × 105 cells/ml, were incubated in the DMEM with 0.5 nm 125I-[Tyr4]bombesin and 5.5 nm unlabeled bombesin in the presence or absence of 0.1% bacitracin and 25 μg/ml leupeptin for 4 hr at 37°. Aliquots (500 μl) were centrifuged, and 400 μl of supernatant was injected onto an HPLC (Waters Model 204, Milford, MA) with a Vydac C18 column (0.46 × 25 cm). 125I-[Tyr4]Bombesin or its breakdown products were eluted using 22.5% acetonitrile in 0.25m triethylammonium phosphate, pH 3.5, at a flow rate of 1 ml/min as described previously for separation of125I-[Tyr4]bombesin.
Immunoprecipitation of tyrosine-phosphorylated proteins.
Bombesin-stimulated tyrosine phosphorylation of p125FAK and paxillin was measured by a variation of the method used in Swiss 3T3 cells (6, 31). Briefly, quiescent and confluent cultures of cells in 100-mm dishes were preincubated twice with DMEM for 1 hr, treated with bombesin at the concentrations indicated, and lysed at 4° in 1 ml of a solution containing 50 mm Tris·HCl, pH 7.5, 150 mm NaCl, 1% Triton X-100, 1% deoxycholate, 0.1% (w/v) NaN3, 1 mm EGTA, 0.4 mm EDTA, 2.5 μg/ml aprotinin, 2.5 μg/ml leupeptin, 1 mmphenylmethylsulfonyl fluoride, and 0.2 mmNa3VO4. Lysates were centrifuged at 15,000 × g for 15 min. Protein concentrations of the supernatants were measured and adjusted to 0.5 mg/ml. The supernatants were incubated with 4 μg of anti-phosphotyrosine mAb PY20, 4 μg of goat anti-mouse IgG, and 30 μl of protein A-agarose overnight at 4°. The immunoprecipitates were washed three times with PBS and further analyzed by SDS-PAGE and Western blotting.
Western blotting and measurement of p125FAK tyrosine phosphorylation.
Immunoprecipitates were fractionated by SDS-PAGE, and the proteins were then transferred to nitrocellulose membranes. Membranes were blocked overnight at 4° using blotto [5% nonfat dried milk in a solution containing 50 mm Tris·HCl, pH 8.0, 2 mm CaCl2, 80 mm NaCl, 0.05% (v/v) Tween 20, 0.02% (w/v) NaN3] and incubated for 2–3 hr at 22° with 1 μg/ml anti-p125FAK mAb. The membranes were washed twice for 10 min in blotto and incubated for 40 min at 22° with anti-mouse IgG-horseradish peroxidase conjugate. The membrane were finally washed four times for 10 min with washing solution [50 mm Tris·HCl, pH 8.0, 2 mmCaCl2, 80 mm NaCl, 0.05% (v/v) Tween 20, 0.02% (w/v) NaN3], incubated with ECL detection reagents for 60 sec, and exposed to Hyperfilm ECL for ≤10 min. The density of bands on the film were measured using a scanning densitometer (Molecular Dynamics, Sunnyvale, CA).
Results
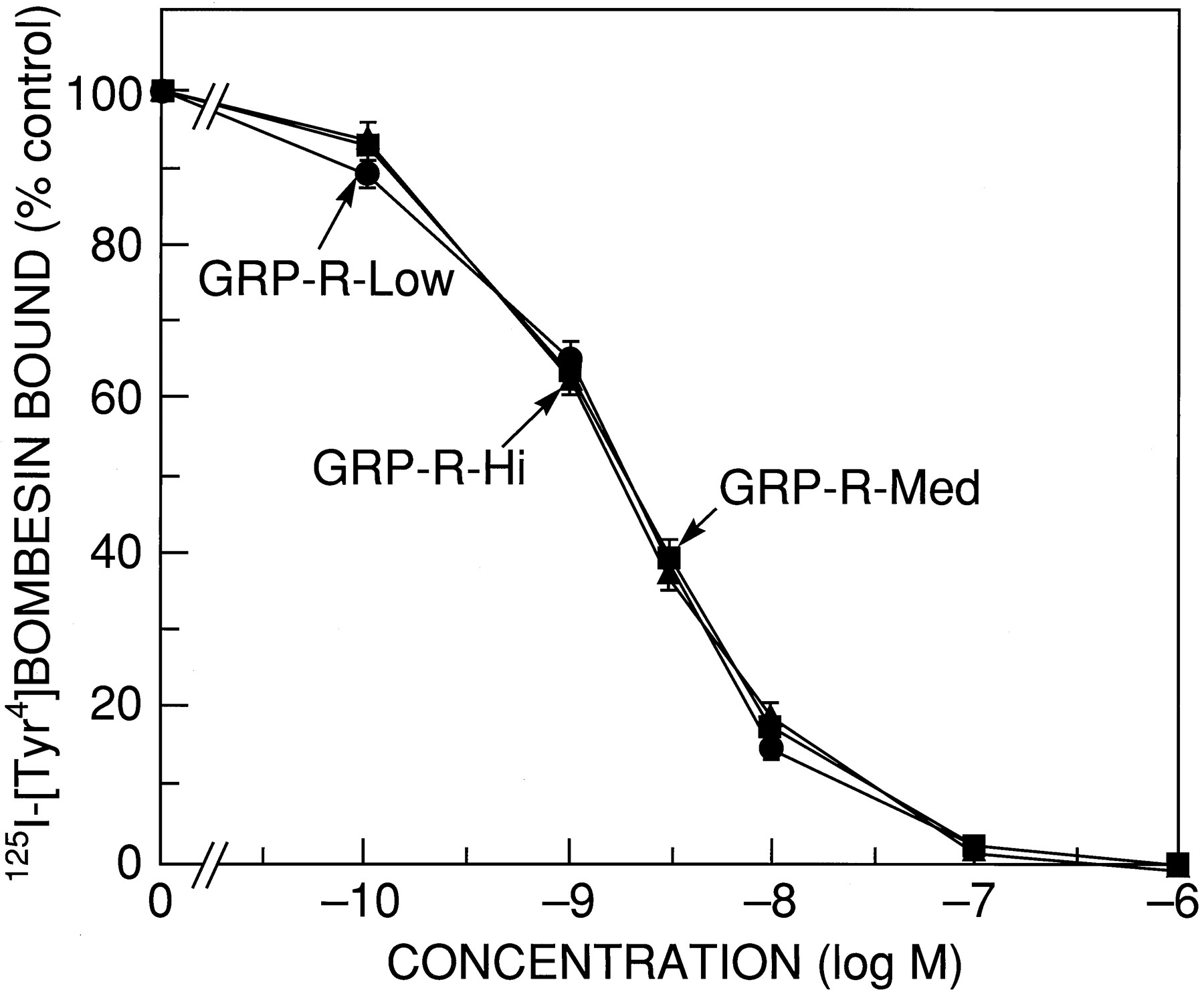
Each of the three GRP-R-transfected BALB 3T3 cells demonstrated similar dose-response curves for bombesin binding (Fig.1). Analysis of the bombesin dose-inhibition data demonstrated that receptor affinity was not altered by receptor number when analyzed in cells or in cell membranes (Table 1). Receptor numbers varied by 280-fold between GRP-R-Low and GRP-R-Hi when determined in intact cells, by 370-fold on membranes (i.e., per 106 cells), and by 160-fold when calculated using membrane protein. In membranes, GRP-R-Med had 3-fold more receptors than GRP-R-Low and had 112-fold fewer receptors than GRP-R-Hi when expressed on a per-cell basis (Table 1).

Ability of bombesin to inhibiting binding of125I-[Tyr4]bombesin to cell lines containing different numbers of GRP-Rs. GRP-R-transfected BALB 3T3 cells were grown to confluence. Cells were harvested and resuspended at a concentration of 3–10 × 106 cells/ml for GRP-R-Low and GRP-R-Med cells and 1 × 105 cells/ml for GRP-R-Hi cells in binding buffer with 75 pm 125I-[Tyr4]bombesin alone or with the indicated concentration of peptide for 45 min at 22°. Data are expressed as the percentage of saturably bound radioactivity in the absence of nonradioactive peptide (i.e., % control). For each experiment, each value was determined in duplicate, and the results are mean ± standard error of at least four experiments.
Receptor affinities and receptor numbers for GRP-R-transfected BALB 3T3 cells studied
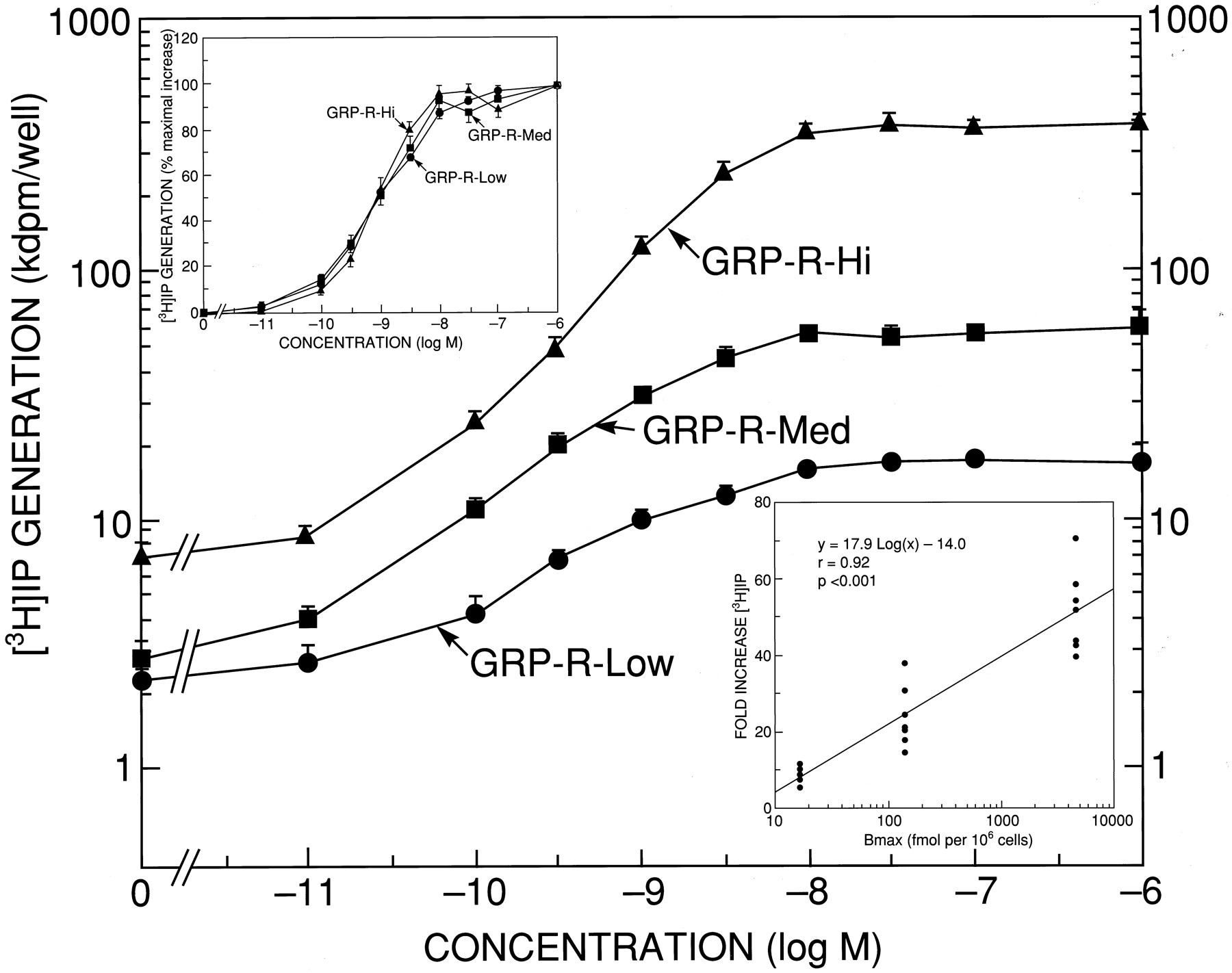
To determine the relationship between receptor number and the bombesin-induced increase in IP, we measured the ability of bombesin to increase [3H]IP in these cell lines (Table2). Each of these cell lines had the same potency (i.e., EC50 = 0.9–1.0 nm; Table 2). However, the cell lines differed slightly in basal values and remarkably in the maximum increase in [3H]IP (Fig. 2 and Table 2). GRP-R-Low increased [3H]IP 8-fold (from 2.3 ± 0.2 to 17.2 ± 0.7 kdpm) when maximally stimulated with 1 μm bombesin. As receptor number increased, the value of maximal increases of [3H]IP became higher to 24-fold in GRP-R-Med (from 2.8 ± 0.4 to 60.5 ± 4.2 kdpm) and 58-fold in GRP-R-Hi (from 7.2 ± 0.6 to 401.1 ± 21.3 kdpm). There was a close correlation (r = 0.92, p < 0.001) between the receptor number and the fold increase (Fig. 2,inset). In contrast, the EC50 values did not differ for the three cell lines containing different receptor numbers (Table 2).
Comparison of the potency and efficacy of bombesin to increase [3H]IP in GRP-R-transfected BALB 3T3 cells with different receptor numbers

Ability of bombesin to stimulate generation of [3H]IP in cell lines containing different numbers of GRP-Rs. GRP-R-transfected BALB 3T3 cells were plated onto 24-well plates at 2.0 × 105 cells/well in DMEM containing 10% fetal bovine serum. At 24 hr later, cells were incubated in DMEM supplemented with 2% fetal bovine serum and 3 μCi ofmyo-[2-3H]inositol for 24 hr. After a 15-min preincubation, cells were exposed to the indicated concentrations of bombesin for 60 min at 37°. [3H]IP were determined as described in Experimental Procedures. Data are expressed as dpm/well. Results are mean ± standard error of at least four experiments. Top inset, ability of bombesin to stimulate in generation of [3H]IP (% of maximal increase). [3H]IP generation was determined as described above, with the data expressed as the percentage of the maximal increase in each cell line caused by 1 μm bombesin for each cell line. Results are mean ± standard error of at least four experiments. Bottom inset, relationship between fold increase of [3H]IP and GRP-R number. [3H]IP generation was determined as described above, with the data expressed as fold increase over the basal value in each cell line. Points, results different experiments. The best fit and correlation coefficient, r, were calculated using a least-squares program.
The GRP-R has been reported to be rapidly internalized after exposure to agonist (29, 30, 32); therefore, we determined whether receptor number altered the internalization rate or extent of internalization. Each of three GRP-R-transfected cell lines rapidly internalized125I-[Tyr4]bombesin (Fig. 3). Receptor number did not alter the percentage of internalized GRP-R (Fig. 3 and Table 3). By 90 min, 77–82% of the radiolabeled 125I-[Tyr4]bombesin was internalized in the different cell lines (Table 3). The rate of internalization in GRP-R-Hi (t 1/2 = 7.3 ± 1.1 min) was not significantly different from that of the GRP-R-Low (t 1/2 = 5.7 ± 1.0 min) but was slightly slower (p < 0.05) than that of the GRP-R-Med (t 1/2 = 4.6 ± 1.7 min) (Fig. 3 and Table3).

Ability of cells with different GRP-R numbers to internalize 125I-[Tyr4]bombesin. GRP-R-transfected BALB 3T3 cells were grown to confluence. Cells were harvested and resuspended at a concentration of 3–10 × 106 cells/ml in GRP-R-Med and GRP-R-Low cells and 1 × 105 cells/ml in GRP-R-Hi cells in binding buffer. The cells were incubated with 75 pm 125I-[Tyr4]bombesin for the indicated times at 37°. After incubation, aliquots were exposed to 0.2 macetic acid in 0.5 m NaCl (pH 2.5) to remove surface-bound ligand as described in Experimental Procedures. Internalized ligand was the proportion of saturably bound counts not removed by exposure to acid wash. Results were expressed as the portion of saturably bound ligand at any time point that was not removed by acid exposure. For each experiment, each value was determined in triplicate.Points, mean ± standard error of at least four separate experiments.
Internalization, down-regulation, and desensitization of GRP-R-transfected BALB 3T3 cells with different receptor numbers
In native and GRP-R-transfected cells, the GRP-R is reported to undergo down-regulation and desensitization (7, 8). Previous studies of the wild-type GRP-R revealed that GRP-R down-regulation was rapid, with exposure to 6 nm bombesin causing >70% decrease by 6 hr and being maximal by 24 hr (7). In preliminary experiments, a similar time course was found for each of the cell lines expressing different receptor numbers. After 24-hr exposure to 6 nm bombesin without protease inhibitors present in the incubation medium, 84 ± 2% and 67 ± 3% down-regulation of the GRP-R occurred in GRP-R-Low and GRP-R-Med, respectively. In contrast, only 46 ± 7% of the GRP-R were down-regulated in GRP-R-Hi by bombesin over the same time period; this was significantly less than that seen in GRP-R-Med and GRP-R-Low (Fig.4). Given the prolonged incubation with agonist, we considered whether an increased rate of degradation of bombesin could contribute to the differences in GRP-R down-regulation. We therefore repeated these studies with two protease inhibitors (i.e., leupeptin and bacitracin) that are reported to inhibit bombesin degradation by GRP-R-containing cells. With protease inhibitors present (0.1% bacitracin and 25 μg/ml leupeptin), each of the GRP-R-expressing cell lines were more down-regulated than without protease inhibitor (Fig.4). With the protease inhibitors present, all cell lines demonstrated >70% down-regulation; however, the degree of down-regulation observed was greater for GRP-R-Low than for either GRP-R-Med or GRP-R-Hi (p < 0.01) (Table 3).

Effect of protease inhibitors and the ability of cells with different GRP-R numbers to undergo down-regulation. GRP-R-transfected BALB 3T3 cells were split 1:4; 24 hr later, cells were incubated for 24 hr with 6 nm bombesin in the presence or absence of 0.1% bacitracin and 25 μg/ml leupeptin in serum-free DMEM. Cells were washed, and binding studies were performed using the same conditions as described in the legend to Fig. 1. Computer analysis of the dose-inhibition curves using a curve-fitting program (LIGAND) demonstrated no change in receptor affinity with a decrease in receptor number (B max). Data are expressed as the percentage of receptors (B max) present in bombesin-treated cells compared with control cells processed similarly.Points, mean ± standard error of at least four separate experiments.
We next determined whether GRP-R numbers affected GRP-R desensitization. Desensitization was assessed after a preincubation with 6 nm bombesin in the presence and absence of protease inhibitors in the preincubation medium. A desensitization of 82 ± 2% occurred in the GRP-R-Low, whereas in GRP-R-Med and GRP-R-Hi bombesin caused only 44 ± 3% and 21 ± 3% desensitization, respectively, values that were significantly lower than that for GRP-R-Low (Fig. 5 and Table 3). To determine whether degradation of bombesin might also attenuate the ability of the receptor to undergo desensitization (similar to the down-regulation studies), we added 0.1% bacitracin and 25 μg/ml leupeptin to the preincubation medium. With protease inhibitors present with each of the GRP-R cell lines, more desensitization occurred than did in those without protease inhibitors present (Fig. 4 and Table 3). Similar to down-regulation, with the protease inhibitors present, all cell lines underwent >70% desensitization; however, the degree of desensitization was greater with GRP-R-Low (p< 0.01) than with GRP-R-Med or GRP-R-Hi, which did not differ significantly from one another.

Effect of protease inhibitors and the ability of cells with different GRP-R numbers to undergo desensitization. GRP-R-transfected BALB 3T3 cells were plated onto 24-well dishes at 1.0 × 105/well in DMEM containing 10% fetal bovine serum; 24 hr later, GRP-R-transfected BALB 3T3 cells were incubated with 6 nm bombesin and 3 μCi ofmyo-[2-3H]inositol in the presence or absence of 0.1% bacitracin and 25 μg/ml leupeptin in serum free DMEM for 24 hr. After washing, cells are preincubated for 10 min and then exposed to 1 μm bombesin for 45 min at 37°. The results are expressed with bombesin-treated cells as the percentage of maximal increase in [3H]IP caused by 1 μm bombesin compared with control cells processed in parallel.Points, mean ± standard error of at least four separate experiments.
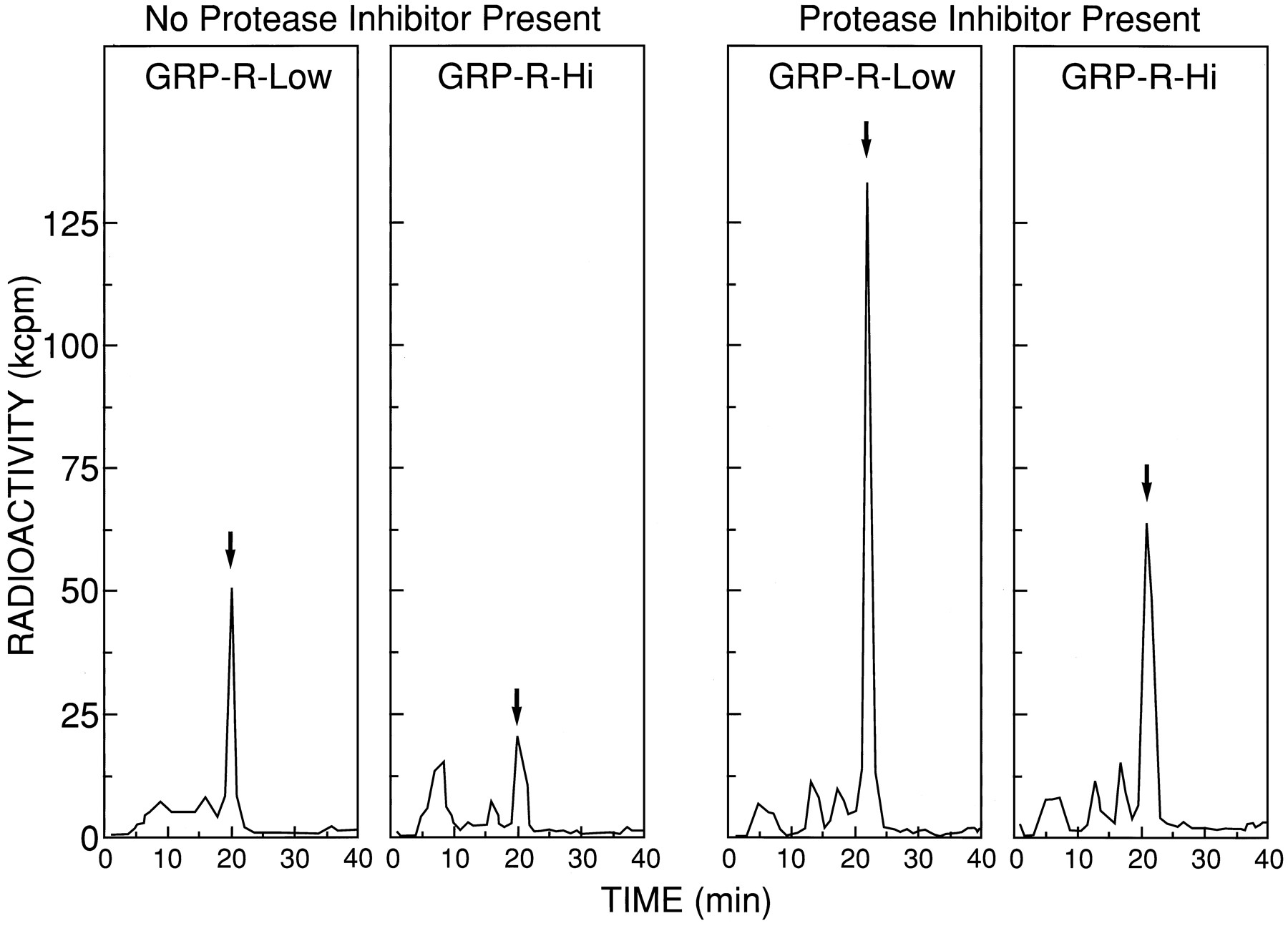
To directly assess the ability of the different cell lines containing different GRP-R receptor number to cause degradation of bombesin, 0.5 nm 125I-[Tyr4]bombesin and 5.5 nm unlabeled bombesin were incubated with each cell line at the same cell concentration (6.5 × 105cells/ml) for 4 hr, and the radioactivity in the supernatants was analyzed. Without protease inhibitors present, HPLC analysis showed that 35% and 59% of the 125I-[Tyr4]bombesin was degraded by GRP-R-Low and GRP-R-Hi, respectively (Fig.6). The addition of protease inhibitors markedly decreased ligand degradation in both cell types. HPLC analysis showed when protease inhibitors were present: 0% of the125I-[Tyr4]bombesin was degraded by GRP-R-Low or GRP-R-Med, and 20% was degraded by GRP-R-Hi (Fig. 6).

Effect of protease inhibitors on the ability of cell lines containing different GRP-R numbers to cause degradation of125I-[Tyr4]bombesin. HPLC elution profiles of supernatants after incubation of 0.5 nm 125I-[Tyr4]bombesin and 5.5 nmunlabeled bombesin with GRP-R-transfected BALB 3T3 cells with or without bacitracin and leupeptin in DMEM. Cells (6.5 × 105 cells/ml) were incubated with 0.5 nm 125I-[Tyr4]bombesin and 5.5 nmunlabeled bombesin for 4 hr at 37° in the absence or presence of 0.1% bacitracin and 25 μg/ml leupeptin. Arrows, elution time of 125I-[Tyr4]bombesin. After incubation, the cells were centrifuged, and the supernatant was applied to HPLC as described in Experimental Procedures. The supernatant from the cells were eluted isocratically with 22.5% acetonitrile in 0.25m triethylammonium phosphate as described in Experimental Procedures. Fractions (1 ml) were collected each minute, and radioactivity was determined in each fraction. These HPLC profiles are from one experiment and are representative of two others.Arrow, point of elution of intact125I-[Tyr4]bombesin.
To determine whether ligand degradation could be responsible for the lack of effect of receptor number on the potency of bombesin in causing PLC activation (Fig. 2), we examined the effect of protease inhibitors on the dose-response curve of bombesin for PLC activation in each cell line assessed by measuring [3H]IP (Fig.7). The presence of protease inhibitors in the same concentrations that markedly inhibited degradation (Fig. 6) had no effect on the dose-response curves of bombesin for stimulating increases in [3H]IP in GRP-R-Low, GRP-R-Med, or GRP-R-Hi (Fig. 7). Specifically, in the presence or absence of protease inhibitors, there was no change in the potency of bombesin for stimulation of [3H]IP in GRP-R-Low (0.7 ± 0.2 versus 2.4 ± 1.2 nm), GRP-R-Med (0.85 ± 0.25 versus 0.80 ± 0.11 nm), or GRP-R-Hi (1.1 ± 0.3 versus 1.4 ± 0.4 nm) or in the efficacy of maximal concentrations of bombesin in any of the cell lines (see Fig. 7legend).

Effect of protease inhibitors on the ability of bombesin to stimulate the generation of [3H]IP in cells containing different numbers of GRP-Rs. Experiments were performed as described in the legend to Fig. 2 except that during the 60-min incubation at 37° with different concentrations of bombesin, 0.1% bacitracin and 25 μg/ml leupeptin were either present or absent. Results are expressed as the percentage of the maximal increase in each cell line caused by 1 μm bombesin. In each cell line, the presence or absence of the protease inhibitors did not alter the magnitude of stimulation or the potency (see text). Specifically, in the GRP-R-Low cell line in the presence of protease inhibitors, the basal and maximal values were 2,587 ± 1,042 and 20,955 ± 7,138 dpm, and in their absence, the basal and maximal values were 2,342 ± 957 and 18,740 ± 6,612 dpm, respectively. In the GRP-R-Med in the presence of protease inhibitors, the basal and maximal values were 2,080 ± 294 and 52,500 ± 6,263 dpm, and in their absence, the basal and maximal values were 3,279 ± 457 and 67,200 ± 11,616 dpm, respectively. In the GRP-R-Hi in the presence of protease inhibitors, the basal and maximal values were 4,412 ± 995 and 277,400 ± 19,700 dpm, and in the absence of protease inhibitors, the basal and maximal values were 4866 ± 1,111 and 272,500 ± 19,600 dpm, respectively. All values are mean ± standard error of three separate experiments.
Recently, the novel cytosolic tyrosine kinase p125FAK was identified as a prominent tyrosine-phosphorylated protein in Swiss 3T3 cells stimulated by bombesin (6). To examine the relationship between receptor number and tyrosine phosphorylation of p125FAKstimulated by bombesin, we compared the ability of bombesin to increase tyrosine phosphorylation of p125FAK in GRP-R-transfected BALB 3T3 cell lines with different receptor numbers. Bombesin caused a similar 4-fold increase in tyrosine phosphorylation of p125FAK in both GRP-R-Low and GRP-R-Hi (Fig. 7). Neither the basal value nor the maximum level of tyrosine-phosphorylated p125FAK differed between the two cell lines (Fig.8). Specifically, the basal values for GRP-R-Low and GRP-R-Hi were 22.5 ± 5.6% and 15.2 ± 6.0%, respectively, of maximal tyrosine phosphorylation elicited by 0.1 μmbombesin stimulation in GRP-R-Hi.

Ability of bombesin to increase tyrosine phosphorylation of p125FAK in cells containing different receptor numbers of GRP-Rs. GRP-R-Low and GRP-R-Hi cells grown to confluence were exposed to 0.1 μm bombesin for 10 min and then treated with lysing buffer. p125FAK tyrosine phosphorylation was analyzed by immunoprecipitation using anti-phosphotyrosine mAb. Immunoprecipitates were applied to SDS-PAGE; then, the proteins transferred to nitrocellulose membranes, and phosphorylated p125FAK was detected using anti-p125FAK mAb and an ECL detection kit as described in Experimental Procedures. Top, result from a experiment representative of three others. Bottom, quantification of p125FAK tyrosine phosphorylation determined by scanning densitometry. Values are mean ± standard error from four experiments and are expressed as percentage of the maximal phosphorylation caused by 0.1 μm bombesin in GRP-R-Hi cells.
We next determined whether GRP-R number affected either the potency or efficacy of agonists to stimulate tyrosine phosphorylation of p125FAK (Fig. 9). The dose-response curves for GRP-R-Low and GRP-R-Med were superimposable with an EC50 value of 0.14 ± 0.09 nm and 0.22 ± 0.11 nm, respectively. The dose-response curve for the GRP-R-Hi demonstrate a small 2-fold shift to the right, with bombesin causing a half-maximal effect at 0.42 ± 0.29 nm (Fig. 9).
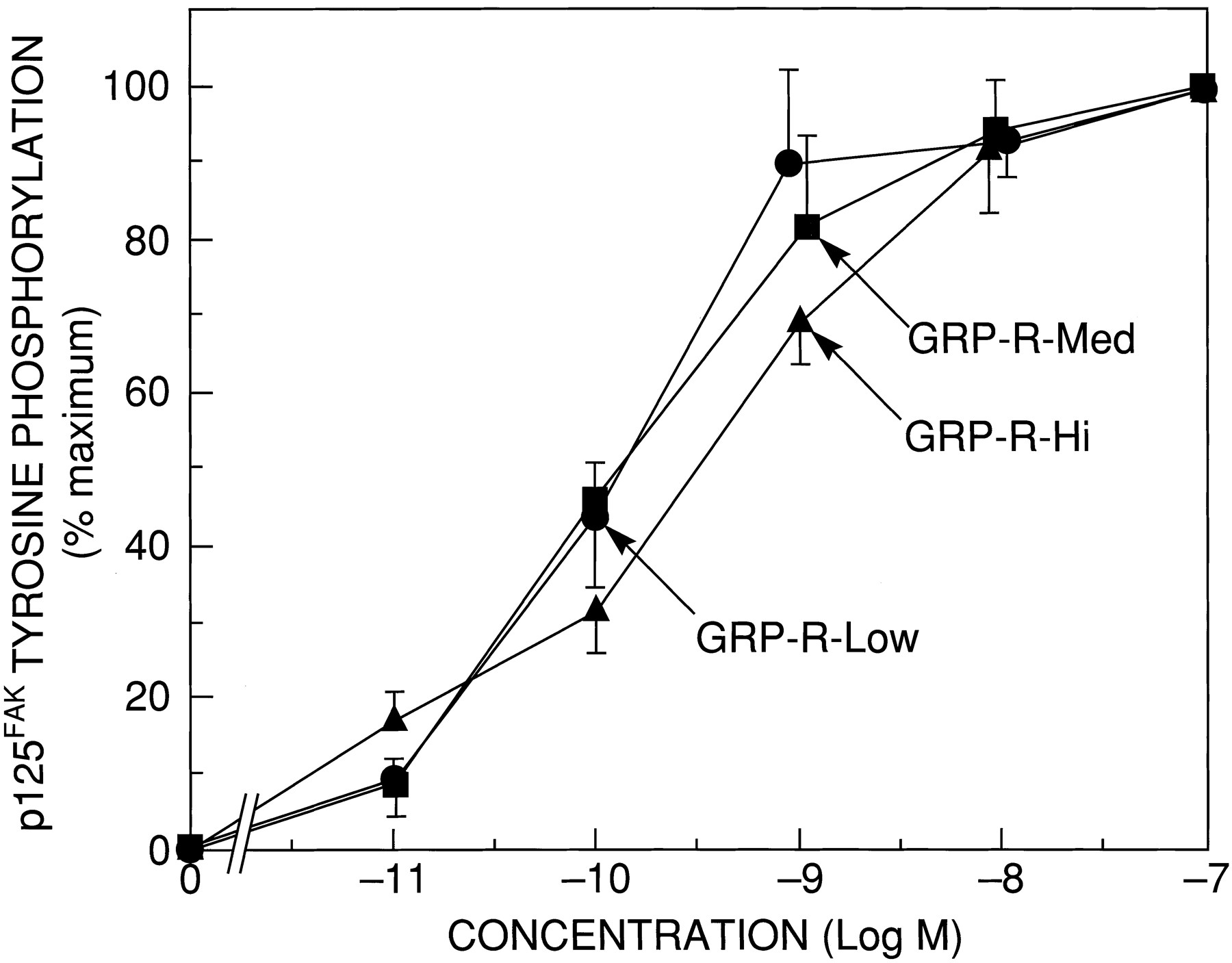
Dose-response curve of the ability of bombesin to stimulate tyrosine phosphorylation of p125FAK in cells containing different receptor numbers of GRP-Rs. Quiescent GRP-R-transfected BALB 3T3 cells were grown to confluence in 100-mm dishes and treated with varying concentrations of bombesin for 10 min; p125FAK tyrosine phosphorylation was determined by immmunoprecipitation and Western blotting as described in the legend to Fig. 7. Results are expressed as percentage of the maximal increase in p125FAK phosphorylation caused by 0.1 μmbombesin over the control level for each cell line. Values are mean ± standard error of at least three experiments.
Discussion
In various normal tissues and tumor cells, the density of GRP-Rs can vary by > 6000-fold (2, 11-13, 33); however, almost nothing is known about the effects of variation of receptor number on cellular function. A number of results suggest that such alterations in GRP-R number could have important effects on cellular responses, particularly related to growth. Numerous studies demonstrate that a number of tumors either acquire the GRP-R or develop an increased number of GRP-Rs, and it has been proposed that bombesin may be important in the growth of these tumor cells, sometimes functioning as an autocrine growth factor (4, 9, 10). Similarly, recent studies with the PLC-coupled m3 muscarinic cholinergic receptor (14) and the neurokinin K2 receptor (15) provide evidence that alterations in coupling or receptor down-regulation or desensitization may be an important factor in affecting cell growth and cell cycle regulation. At present, almost no information is available on the effects of receptor numbers for PLC-linked receptors such as the GRP-R on cellular processes such as coupling, receptor down-regulation, or desensitization, which is in contrast to the β2-adrenergic receptor and other receptors coupled to adenylate cyclase, which have been extensively studied (16, 22). The GRP-R is a good model system in which to investigate these interactions for a number of reasons. Recent studies demonstrated that after binding to the GRP-R, GRP activates PLC and causes rapid internalization of the GRP-R, time-dependent homologous desensitization, and down-regulation (7). Also, activation of the GRP-R in some cell types (7) causes tyrosine phosphorylation of p125FAK and a number of other proteins (6). A recent study (7) demonstrated that when the murine GRP-R is transfected into BALB 3T3 cells, which do not possess this receptor but are closely related to the Swiss 3T3 cell from which the GRP-R was originally cloned, the GRP-R stably transfected cells behave in a similar fashion to cells possessing native GRP-R. In the current study, to investigate the relationship between receptor number and various cellular responses with the GRP-R, we studied three clonal cell lines of GRP-R-transfected BALB 3T3 cells with a 280-fold difference in receptor density. A number of tumor cells and native tissues have been reported to have GRP-R receptor densities in the range of 0.1–0.4 pmol/mg of protein (∼1,100–4,700 receptors/cell) (2, 12, 34), which is the range of the GRP-R cell line with low receptor numbers (GRP-R-Low) used in the current study. Similarly, a number of tumorous and native tissues possess GRP-R in the range of 1–4 pmol/mg of protein (12,000–50,000 receptors/cell), which is in the range of the GRP-R-Med cell line used in the current study, including some Swiss 3T3 cell lines that have been extensively used to investigate the cellular basis of action of the GRP-R (3). Last, some normal tissues, such as some Swiss 3T3 cell lines (13), gastric G cells (11), and some tumor tissues, have high GRP-R receptor densities of 20–60 pmol/mg of protein (∼234,000–750,000 receptors/cell), which is in the range of the GRP-R-Hi cell line used in the current study.
In our study, GRP-R number had a different effect on receptor affinity and receptor coupling to PLC. Increasing the receptor number had no effect on receptor affinity for various agonists determined directly by binding studies. These results are similar to those reported for a number of receptors coupled to adenylate cyclase such as the β2-adrenergic receptor (16, 17, 19, 21, 22), secretin receptor (23), δ-opioid receptor (24), m2 muscarinic receptor (25), and 5-HT1A (26) and 5-HT1B receptors (27). Our results differ from those of a recent study of the human GRP-R (28); in this study, the human GRP-R was transfected at two receptor densities, with one receptor density similar to our GRP-R-Med (42,000 receptors/cell) and a second cell line with 2-fold higher numbers than our GRP-R-Hi (1,340,000 receptors/cell), which had a 3-fold lowerKd value than the cell line with lower numbers (28). In contrast to the lack of effect on binding affinity in our study, increasing the receptor number resulted in only a small (3-fold) increase in basal PLC activity as assessed by measuring [3H]IP generation; however, it had a marked effect on agonist efficacy because there was a very close correlation (r = 0.92) between receptor number and fold increase in [3H]IP with agonist stimulation. These results are similar to those reported in studies of the effect of receptor number on adenylate cyclase activation for adenylate cyclase-coupled receptors (16, 18-23) and for the ability of the luteinizing hormone receptor to increase PLC activation assessed by measuring changes in intracellular Ca2+ concentration (20). In contrast to the efficacy, the potency of GRP in activating PLC was identical in each cell line and therefore was not influenced by receptor number. Although this result is similar to that found with the PLC-linked α1B-adrenoreceptor (35), these results, in general, differ from those of many (16-18, 20, 21, 23-25, 27) but not all (19, 26, 36) studies with adenylate cyclase-coupled receptors. With most adenylate cyclase-coupled receptors, increasing the receptor expression resulted in an increased potency for agonist stimulation of adenylate cyclase (16-18, 20, 21, 23-25, 27). Our results with PLC activation have both similarities and differences with a recent study that assessed the effect of GRP and five synthetic bombesin analogues on intracellular Ca2+ concentration increases in two cell lines transfected with human GRP-R that differ in receptor number by 30-fold (28). With the synthetic bombesin peptides, which functioned as partial agonists, but not with GRP itself, greater efficacy was seen in the cells with the highest human GRP-R numbers. In contrast to our results, GRP was more potent in the cells with the highest human GRP-R number (28). At present, the explanation for these differences remains unclear but could be due to a difference between [3H]IP and intracellular Ca2+ concentration stoichiometry with receptor number, a difference in coupling of the GRP-R in the different species, or to differences in the range of the receptor number used because in this latter study, the cell line with the highest number had >1 million receptors/cell, which was 2-fold higher than that of GRP-R-Hi in our study (28). Our results suggest the effect of receptor number on receptor coupling differs for receptors coupled to adenylate cyclase and those coupled to PLC. This conclusion is supported by recent results in receptors coupled to both the PLC and adenylate cyclase cascades (20, 25). Studies with m2 muscarinic receptor (25) and luteinizing hormone receptor (20) demonstrate that increasing receptor number causes an increased agonist potency for stimulating adenylate cyclase but does not change the agonist potency for PLC activation (20,25).
There have been few studies investigating the effect of receptor number on receptor modulation (internalization, down-regulation, desensitization) by agonists (23, 24), and none have been performed on PLC-linked receptors. In native GRP-R containing cells such as Swiss 3T3, AR 42J, and pancreatic acinar cells, 20–85% of radiolabeled agonists but not antagonists are rapidly internalized (13, 30, 34), and the receptor undergoes both down-regulation and desensitization. In our study, 80% of the radiolabeled agonist,125I-[Tyr4]bombesin, was internalized within 90 min in each of the cell lines with different receptor numbers, demonstrating that GRP-R number had no effect on the amount of internalization. Our results suggest that receptor number has a minimal effect on internalization rate because the cell line with highest density of receptors had a similar rate of internalization to the cell line with the lowest numbers but a slightly slower rate of internalization (p < 0.05) than those with an intermediate receptor number. The mechanism of this was not investigated, but it is unlikely to be due to ligand degradation because protease inhibitors were present in the incubation medium that were shown to inhibit ligand degradation. Another possibility that could be raised to explain the difference from the cell line with the highest numbers (GRP-R-Hi) and that with the intermediate numbers (GRP-R-Med) is that the amino-terminal myc epitope tag on the GRP-R-Hi altered cell function. In a previous study (29) in this cell line, cellular response was reported to be similar to cells without an epitope tag. Similarly, in this study, cells with the epitope-tagged receptor (GRP-R-Hi) did not differ from cells without this epitope in internalization extent, receptor binding affinity, EC50 value for PLC activation (GRP-R-Low, GRP-R-Med), internalization rate (GRP-R-Low), or the extent of down-regulation or desensitization (GRP-R-Med), further supporting the conclusion that its presence was not altering cell function. These data suggest that with the GRP-R, neither receptor number nor the amount number of ligand bound is a critical determinant of either receptor internalization rate or extent.
Previous studies show that both the native GRP-R in various cells and the GRP-R transfected into BALB 3T3 cells (7) undergo homologous down-regulation after pretreatment with bombesin. In the current study, a similar degree of down-regulation (≥ 65%) was seen with the GRP-R-transfected cell line with a low (GRP-R-Low) or intermediate (GRP-R-Med) number of GRP-Rs when protease inhibitors were not present during the preincubation, whereas in the cell line with a high (GRP-R-Hi) number of GRP-Rs, significantly less down-regulation occurred. This result is similar to that reported with secretin receptors transfected into CHO cells at two receptor densities (23). In that study (23), preincubation of both cell lines with 0.1–10 nm secretin for 24 hr caused marked secretin receptor down-regulation. However, the degree of down-regulation with the cell line with low secretin receptor numbers was greater than that of the cell line with high secretin receptor numbers (23). In contrast, when the δ-opioid receptor was transfected into CHO cells (24) at high, medium, and low receptor numbers, down-regulation was the least with the cell line with the lowest receptor numbers. The results in our study and with the secretin receptors in CHO cells could be due to a direct effect of receptor number on down-regulation or to an indirect effect of increased degradation of the ligand during the preincubation. To distinguish between these two possibilities, we investigated the effect of GRP-R number on ligand degradation. We found that in the absence of protease inhibitors, degradation of125I-[Tyr4]bombesin occurred and the amount of degradation was much higher in cells with the highest GRP-R numbers. This result is consistent with previous studies that showed that a number of cells with native GRP-Rs degrade the radiolabeled agonist ligand 125I-[Tyr4]bombesin rapidly in the absence of protease inhibitors (13, 34). Various protease inhibitors, such as bacitracin, leupeptin, and phosphoramidon, have been shown to very efficiently inhibit this degradation. We found that with protease inhibitors present, each of the three cell lines was more significantly down-regulated than without protease inhibitors. Both the cell line with high GRP-R numbers (GRP-R-Hi) and the cell line with 61-fold lower GRP-R numbers (GRP-R-Med) caused 75% down-regulation, which was similar to that observed for GRP-R natively expressed by Swiss 3T3 cells (30). The degree of receptor down-regulation of the GRP-R cell line with low receptor numbers (GRP-R-Low) was significantly greater (p < 0.01) than that of the other two cell lines, even in the presence of the protease inhibitors. These data suggest that receptor number itself has an effect on the extent of down-regulation measured in cells with low receptor numbers, but for cells expressing high receptor numbers, further increasing the number of receptors has no effect. The results are consistent with the conclusion in the absence of protease inhibitors that ligand degradation is one of the principal factors in causing decreased down-regulation when the receptor numbers were increased to a very high density. In addition, these results suggest that one of the effects of increasing GRP-R numbers on various tumor cells could be to decrease the extent of agonist-induced down-regulation. Decreased down-regulation may confer an important growth advantage to some tumor cells; a recent study suggests that decreased receptor down-regulation might result in growth advantages by decreasing growth-inhibiting elements (14).
A number of previous studies demonstrated that natively expressed GRP-Rs can undergo homologous desensitization. We found that similar to down-regulation in the absence of protease inhibitors, the GRP-R cell with low receptor numbers (GRP-R-Low) was well desensitized, whereas the two with higher GRP-R numbers (GRP-R-Med and GRP-R-Hi) were poorly desensitized. This result is similar to that reported with secretin receptors transfected into CHO cells at two receptor densities (23) and with the δ-opioid receptor transfected into CHO cells with different receptor numbers (24). In our study and in the studies with the secretin and opioid receptors (23, 24), this decreased effect with higher receptor number could be due to ligand degradation. In the current study, when protease inhibitors that inhibited agonist degradation were present, each cell line demonstrated greater desensitization in a manner similar to that seen with down-regulation. Similar to down-regulation, these results demonstrate that in the absence of protease inhibitors, one of the mechanisms of decreased desensitization with very high receptor number is increased ligand degradation. With protease inhibitors present, each GRP-R cell line demonstrated >70% desensitization, and there was no difference between the cells with the highest receptor number (GRP-R-Hi) and those with a 61-fold lower receptor number (GRP-R-Med). However, the cell line with the lowest receptor number (GRP-R-Low) was significantly (p < 0.01) better desensitized than the other two cell lines with higher receptor numbers. Similar to that observed for down-regulation, the receptor number affected the extent of desensitization seen in cells with low receptor numbers, but with high receptor numbers, an additional increase in the number of receptors had no further effect. The decreased desensitization seen with increasing receptor numbers suggests the possibility that the increased GRP-R number that occurs in tumor cells might confer a selective growth advantage by decreasing desensitization. This hypothesis is supported by recent studies (14, 15) showing that receptor desensitization is an important regulator of various elements controlling cell growth; thus, if less desensitization occurs, rapid growth would be favored.
A number of previous studies suggest that chronic desensitization and down-regulation of the GRP-R are likely coupled processes, mediated by similar mechanisms, and these mechanisms differ from those mediating internalization (37-39) Our results support this proposal because there was a close relationship between the effect of receptor number on down-regulation and desensitization but not on internalization. Specifically, with both down-regulation and desensitization when receptor number was low, an increase in receptor number resulted in less down-regulation and desensitization; however, increases to a large number of receptors did not cause a further decrease in down-regulation or desensitization. In contrast, our study showed GRP-R number had no effect on the extent, and only a minimal effect on the rate of internalization, suggesting a different relationship exists between receptor number and this process compared with GRP-R down-regulation and desensitization.
Recent studies show that activation of receptors for a number of neuropeptides such as the GRP-R can cause tyrosine phosphorylation of a number of proteins, one of which is the novel cytosolic tyrosine kinase p125FAK (6), which may be important in mediating the growth effects (6, 31, 40). No studies have investigated the relationship between receptor number and activation of this cascade by neuropeptide receptors. We found that there was no significant difference in either the basal or maximal value of tyrosine phosphorylation of p125FAK between cells transfected with a low or high number of GRP-Rs even though they differed by >200-fold in receptor number. However, GRP-R number had an effect on the potency of bombesin for causing tyrosine phosphorylation of p125FAK in the different cell lines. The potency of bombesin for causing tyrosine phosphorylation in the cell line with high receptor number (GRP-R-Hi) was less than that in the cell lines with lower receptor numbers. Therefore, GRP-R number had a different effect on coupling of the receptor to PLC and to the tyrosine phosphorylation cascade. Although receptor number had no effect on potency of bombesin for PLC activation (only on its efficacy for activating PLC), for tyrosine phosphorylation, GRP-R number had the reverse effect, altering the potency but not the efficacy of the agonist. This difference is not due to a difference in receptor affinities because with the different cell lines with different receptor numbers, the binding affinity was the same. At present, the basis for this different effect of receptor number on coupling to the PLC and tyrosine phosphorylation cascade is unknown.
In conclusion, we described the effect of GRP-R number on ligand affinity, ability of agonist to activate intracellular messengers (PLC, tyrosine phosphorylation of p125FAK), and receptor modulation (internalization, down-regulation, desensitization). Increasing the receptor number affected different transduction pathways differently in that with increasing receptor number, the efficacy for bombesin-induced [3H]IP generation increased; however, it had no effect in the efficacy of tyrosine phosphorylation. It also had no effect on the receptor affinity in binding studies or in potency for activating PLC. Receptor number had a different effect on different receptor modulatory processes altering down-regulation and desensitization but not internalization. Specifically, high receptor number significantly (p < 0.01) attenuated the extent of agonist-induced down-regulation and desensitization but not the extent of internalization. An important unforeseen effect of increasing receptor number was to increase the degradation of agonists. This had a profound effect on processes involving prolonged incubations and therefore needs to be inhibited to truly assess the effects of receptor number only on the different processes. In previous studies of the effects of receptor number on various cellular processes (20), the possibility of effects on degradation were not considered, and any changes seen were attributed to the effect of the receptor numbersper se (23) on the processes. In many cases there was a decreased effectiveness of agonists and this could be due to increased ligand degradation rather than an effect of receptor number per se. These results demonstrate that the relationships between receptor numbers and receptor-mediated functions differ for the PLC-coupled GRP-R compared with those for various adenylate cyclase-coupled receptors. Furthermore, these results suggest that by altering receptor numbers, PLC-linked receptors in different cells can have marked effects on receptor-mediated processes that may have important effects on important functions such as cell growth.
Footnotes
- Received September 30, 1996.
- Accepted January 8, 1997.
-
Send reprint requests to: Dr. Robert T. Jensen, NIH/NDDKD/DDB, Bldg. 10, Room 9C-103, 10 Center Drive MSC 1804, Bethesda, MD 20892-1804. E-mail:robertj{at}bdg10.niddk.nih.gov
Abbreviations
- GRP
- gastrin-releasing peptide
- 5-HT
- 5-hydroxytryptamine
- mAb
- monoclonal antibody
- PLC
- phospholipase C
- CHO
- Chinese hamster ovary
- GRP-R
- gastrin-releasing peptide receptor
- IP
- inositol phosphates
- p125FAK
- p125 focal adhesion kinase
- PBS
- phosphate-buffered saline
- DMEM
- Dulbecco’s modified Eagle’s medium
- SBTI
- soybean trypsin inhibitor
- EGTA
- ethylene glycol bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid
- HEPES
- 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
- SDS
- sodium dodecyl sulfate
- PAGE
- polyacrylamide gel electrophoresis
- HPLC
- high performance liquid chromatography
- ECL
- enhanced chemiluminescence
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}