

Abstract
P2X7 receptor subunits form homomeric ATP-gated, calcium-permeable cation channels. In this study, we used Western blots and immunocytochemistry to demonstrate that P2X7 receptors are abundant on presynaptic terminals of mossy fiber synapses in the rat hippocampus. P2X7-immunoreactive protein was detected using a specific P2X7 antibody in Western blots of protein isolated from whole hippocampus and from a subcellular fraction containing mossy fiber synaptosomes. P2X7 immunoreactivity was colocalized with syntaxin 1A/B-immunoreactivity in mossy fiber terminals in the dentate hilus and stratum lucidum of CA3. Extracellular and whole-cell voltage-clamp recordings in CA3 revealed that bath application of the potent P2X7 agonist 2′,3′-O-(4-benzoylbenzoyl)-ATP (Bz-ATP) caused a long-lasting inhibition of neurotransmission at mossy fiber–CA3 synapses. Consistent with a presynaptic action at mossy fiber synapses, Bz-ATP had no significant effect on neurotransmission at associational–commissural synapses in CA3 but increased paired-pulse facilitation during depression of mossy fiber evoked currents. In addition, Bz-ATP had no postsynaptic effect on holding current or conductance of CA3 neurons. Bz-ATP-induced mossy fiber synaptic depression was blocked by the P2X7 antagonist oxidized ATP but not by the P2X1–3,5,6 antagonist pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid or the P2Y antagonist reactive blue 2. Finally, an antagonist of p38 MAP kinase activation [4-(4-fluorophenyl)2-(4-methylsulfinylphenyl)5-(4-pyridyl)imidazole] but not extracellular signal-regulated kinase 1/2 MAP kinase (2′-amino-3′-methoxyflavone) blocked the synaptic depression mediated by Bz-ATP, suggesting that this presynaptic inhibition was mediated by activation of p38 MAP kinase. The results of the present study demonstrate that activation of presynaptic P2X7receptors depresses mossy fiber–CA3 synaptic transmission through activation of p38 MAP kinase.
ATP is released from synapses throughout the peripheral nervous system and CNS (White, 1977, 1978;Jahr and Jessell, 1983; Edwards et al., 1992; Edwards and Gibb, 1993), where it can act on P2X receptors to modulate neurotransmission. P2X receptors are ligand-gated, calcium-permeable cation channels (Khakh, 2001) that are activated by extracellular ATP. There are seven known P2X receptor subunits, P2X1–7. Of the seven subunits, only P2X7 subunits are thought to function exclusively as homomeric receptors (North and Surprenant, 2000). Activation of P2X7 receptors can lead to the initiation of signaling cascades through second messengers, such as phospholipase D (Kusner and Adams, 2000), p38 MAP kinase (Hu et al., 1998; Hide et al., 2000; Panenka et al., 2001), or the transcription factor nuclear factor-κB (Ferrari et al., 1997). Recent data suggest that the initiation of these signaling cascades could be mediated through putative protein interactions with the long cytoplasmic C terminus of the P2X7 subunit (Denlinger et al., 2001; Kim et al., 2001). In some circumstances, the pore formed by the P2X7 receptor may allow permeation of large cations (North and Barnard, 1997; North and Surprenant, 2000) that may eventually lead to cytolysis (Di Virgilio, 1995; Baricordi et al., 1999; Mutini et al., 1999).
P2X7 receptors are only activated by high extracellular concentrations of ATP. Low concentrations (nanomolar) of ATP that are ineffective at activating P2X7receptors are known to increase neuronal excitation and synaptic activity in the nervous system. For example, ATP-induced activation of P2X receptors can evoke single-channel cation currents from chick ciliary ganglion nerve terminals (Sun and Stanley, 1996) and enhance the frequency of miniature endplate currents at the frog neuromuscular junction (Fu and Poo, 1991). Activation of P2X receptors has also been shown to increase the frequency of miniature postsynaptic currents in dorsal root ganglion dorsal horn neuronal cocultures (Gu and MacDermott, 1997; MacDermott et al., 1999) and increase excitation in the hippocampus (Wieraszko and Seyfried, 1989; Inoue et al., 1992,1995). However, high concentrations of ATP (micromolar) are known to induce a long-lasting form of synaptic depression that cannot be explained by the degradation of ATP into adenosine (Wieraszko and Seyfried, 1989). The synaptic depression mediated by high concentrations of ATP could be explained by the activation of presynaptic P2X7 receptors. Recently, Deuchars et al. (2001) reported the presynaptic localization of P2X7 receptors in the spinal cord and brainstem of the rat. We subsequently investigated the anatomical distribution of P2X7 receptors in the rat hippocampus. When we discovered that P2X7 receptors were abundant on hippocampal mossy fiber terminals, we used whole-cell and extracellular field recordings to determine the actions of P2X7receptor activation on neurotransmission at this synapse. In the following study, we provide physiological and pharmacological evidence that activation of these presynaptic P2X7receptors results in rapid and long-lasting synaptic depression that is mediated through a p38 MAP kinase signaling cascade.
MATERIALS AND METHODS
SDS-PAGE and Western blotting. Hippocampal proteins were isolated from Sprague Dawley rats (3–4 weeks old) and homogenized in 0.32 m sucrose. Small (P2) and large (P3) mossy fiber synaptosomal fractions were then isolated according to previously published methods (Terrian et al., 1988, 1989). Proteins were subjected to SDS-PAGE on 10% gels and probed with the following antibodies: rabbit anti-P2X7 polyclonal (1:18,000; Alomone Laboratories, Jerusalem, Israel), rabbit anti-NMDA receptor subunit 1 (NMDAR1) (1:3000; Chemicon, Temecula, CA), mouse anti-β-tubulin (1:6000; Sigma, St. Louis, MO), and rabbit anti-synaptoporin (1:30,000; Synaptic Systems, Gottingen, Germany). Immunoreactive signals were visualized using peroxidase-labeled goat secondary antibodies (1:10,000; Jackson ImmunoResearch, West Grove, PA) and enhanced chemiluminescence (Lumi-Lightplus; Roche Diagnostics, Mannheim, Germany).
Immunocytochemistry. For immunocytochemistry, rats were anesthetized and transcardially perfused with 4% paraformaldehyde in 0.1 m phosphate buffer, pH 7.4. Brains were sectioned in the coronal plane (50 μm) on a vibrating microtome (VT100; Leica, Willowdale, Ontario, Canada) and processed for immunocytochemistry using standard procedures (Sloviter et al., 1996). The following primary antibodies were used: rabbit anti-P2X7 (1:3000; Alomone Laboratories), mouse anti-MAP-2 (1:20,000; Sigma), or anti-syntaxin 1A/B (1:5000; Stressgen, Victoria, British Columbia, Canada). The following secondary antibodies were used: biotinylated donkey anti-mouse or rabbit IgG, Cy2-conjugated donkey anti-mouse IgG and Cy3-conjugated donkey anti-rabbit IgG, or Cy5-conjugated donkey anti-rabbit IgG (1:1000;all from Jackson ImmunoResearch). Sections were imaged on an Axioskop LSM510 laser scanning microscope (Carl Zeiss Microscopy, Jena, Germany).
Electrophysiology. Hippocampal slices (300 μm thick) were obtained from 10- to 30-d-old rats, immersed in ice-cold artificial CSF (aCSF; see below), and incubated in a submersion chamber for ≥1 hr at room temperature. For recordings, individual slices were transferred to either an interface chamber (Fine Science Tools, Foster City, CA) for extracellular recordings or a submersion chamber for whole-cell voltage-clamp recordings. All recordings were done at room temperature. In either chamber, slices were superfused (2 ml/min) with aCSF consisting of (in mm): 119 NaCl, 2.5 KCl, 1.3 MgSO4, 26 NaHCO3, 1 NaH2PO4, 2.5 CaCl2, and 10 glucose, aerated with 95% O2/5% CO2. Extracellular recordings were obtained with glass micropipettes filled with HEPES-buffered aCSF (resistance, 1–3 MΩ). Extracellular recordings were filtered at 5 kHz, digitized at 10 kHz using a Digidata1200 interface (Axon Instruments, Foster City, CA), and stored on a Pentium III computer for later analysis using Clampfit (Axon Instruments). A bipolar tungsten-stimulating electrode was used to stimulate dentate granule cells, thereby activating mossy fibers. Mossy fiber–CA3 synaptic responses were measured in the stratum lucidum of the CA3 region and distinguished by their characteristic short latency, rapid rise time, large paired-pulse facilitation (PPF), and >70% inhibition by the metabotropic glutamate receptor (mGluR) agonist (2s,1′s,2′s)-2(carboxycyclopropyl)glycine (L-CCG-1).
Whole-cell recordings were obtained using patch pipettes filled with (in mm): 100 cesium methanesulfonate, 10 cesium-BAPTA, 40 HEPES, and 5 N-(2,6-dimethylphenyl carbamoylmethyl)triethylammonium bromide, adjusted to a pH of 7.3 with cesium hydroxide (resistance, 1–3 MΩ). CaCl2 and MgSO4 were increased to 4 mm in the aCSF for all whole-cell recordings. During paired-pulse facilitation experiments, picrotoxin (10 μm) was included in the patch pipette to block GABAA receptors (Nelson et al., 1994; Xiang and Brown, 1998). Series resistance in all recordings was <20 MΩ, and data were excluded if series resistance varied by >15%. All recordings were digitized at 5–10 kHz and filtered at 2 kHz.
Statistics. All statistics were performed using a paired (correlated groups) t test except for the comparison between the effect of 2′,3′-O-(4-benzoylbenzoyl)-ATP (Bz-ATP) and adenosine on slices incubated with 4-(4-fluorophenyl)2-(4-methylsulfinylphenyl)5-(4-pyridyl)imidazole (SB203580). In this case, a simple one-way ANOVA was used.
RESULTS
P2X7 receptors were found on mossy fiber terminals
Western blotting and immunocytochemistry revealed that P2X7 receptors were abundant on presynaptic terminals of the rat hippocampus. We used a P2X7antibody that was raised against amino acid residues 576–595 of the rat P2X7 receptor subunit. This antibody recognized a single 70 kDa band in Western blots of proteins isolated from the hippocampus (Fig.1A,B), small (P2) hippocampal synaptosomes, and large, mossy fiber (P3) synaptosomes (Fig.1B). Inclusion of the P2X7antigenic peptide (1:1) with the antibody blocked detection of the 70 kDa P2X7-immunoreactive band. We did not detect any signal in the P3 fraction with an antibody against iba-1, a protein selectively expressed in microglia (Ito et al., 1998). This indicates that microglia did not contaminate our P3 synaptosome preparation (data not shown). Immunocytochemistry with this P2X7-selective antibody revealed dense immunoreactive terminals throughout mossy fiber termination zones in the dentate hilus and stratum lucidum of CA3 (Fig.1C, arrows). Fainter staining was also observed throughout the hippocampus and may represent immunoreactivity of other presynaptic terminals, or glial cells, such as microglia (Ferrari et al., 1996; Chessell et al., 1997; Di Virgilio et al., 1999) or astrocytes (Kukley et al., 2001; Panenka et al., 2001). Confocal microscopy confirmed that the P2X7-immunoreactive boutons were presynaptic mossy fiber terminals, because P2X7 immunoreactivity was colocalized with presynaptic syntaxin 1A/B immunoreactivity (Bennett et al., 1992;Ruiz-Montasell et al., 1996) (Fig. 1G–I) but not with dendritic MAP-2 immunoreactivity (Fig.1D–F).
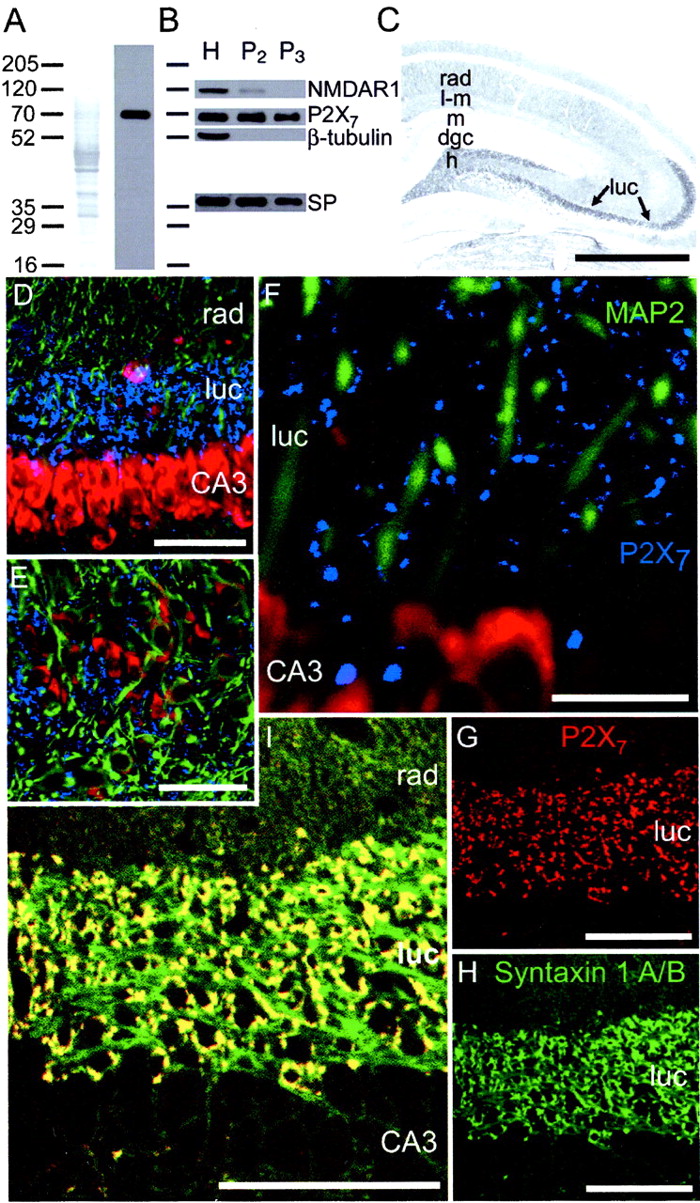
P2X7 receptors are located on presynaptic terminals of mossy fiber synapses. A, A Western blot of proteins isolated from the rat hippocampus showed that the P2X7 antibody recognized a single band of protein at an approximate molecular weight of 70 kDa. Lane 1, Amido Black-stained proteins that were first immunoblotted in lane 2. B, P2X7-immunoreactive bands of protein were present in proteins isolated from whole hippocampus (H), as well as small (P2) synaptosomal and large (P3 ) mossy fiber synaptosomal preparations. The presence of synaptoporin and relatively low abundance of NMDAR1 and β-tubulin indicates that P2X7 receptors were highly enriched in the fraction containing mossy fiber terminals (P3 ). C, Immunocytochemistry with this P2X7-selective antibody revealed dense immunoreactivity throughout mossy fiber termination zones in the dentate hilus (h) and stratum lucidum (luc) of CA3 (arrows). Fainter staining was also observed throughout the hippocampus and may represent immunoreactivity of other presynaptic terminals. rad, Stratum radiatum;l-m, stratum lacunosum-moleculare; m, molecular layer; dgc, dentate granule cell layer.D–L, Colocalization studies demonstrated that P2X7 immunoreactivity was located in the presynaptic terminals of mossy fiber synapses. D–F, P2X7 immunoreactivity (blue) was found throughout stratum lucidum; however, dendritic MAP2 immunoreactivity (green) did not colocalize with the punctate P2X7 immunoreactivity. Cell bodies were counterstained with ethidium bromide (red). G–I, In contrast, presynaptic syntaxin 1A/B immunoreactivity (green) was colocalized with the punctate P2X7 immunoreactivity (red), demonstrating that the mossy fiber terminals contained P2X7 receptors. Scale bars: C, 2 mm; D, 70 μm;E, 50 μm; F, 30 μm;G–I, 100 μm.
P2X7 receptor activation depressed mossy fiber–CA3 synaptic transmission
Next, we investigated the effect of P2X7receptor activation on synaptic transmission at mossy fiber synapses. First, we recorded evoked postsynaptic field potentials (fEPSPs) in the stratum lucidum of CA3 after stimulation of the dentate granule cells (Fig. 2). To ensure that we were recording from mossy fiber–CA3 synapses, we first applied L-CCG-I (20 μm), an mGluR agonist that selectively depresses mossy fiber inputs onto CA3 pyramidal cells (Manzoni et al., 1995; Schmitz et al., 2000). Bath application of L-CCG-I reversibly depressed the amplitude of the fEPSP (Fig. 2A). Subsequent bath application of the P2X7 receptor agonist Bz-ATP (30 μm) also depressed the fEPSP (Fig. 2A). However, application of L-CCG-I during the peak of the Bz-ATP response did not result in any additional depression of the synaptic response (fEPSP amplitude after Bz-ATP was 0.22 ± 0.12 of controls vs 0.17 ± 0.09 of controls in Bz-ATP plus L-CCG-I; mean ± SEM; n = 3), indicating that P2X7 receptor activation depressed the same population of synaptic inputs as L-CCG-I. Bz-ATP was also applied alone to monitor the time course of the P2X7-mediated synaptic depression without previous L-CCG-I application (Fig.2B). This prevented any potential interactions between progressive drug applications. As shown in Figure2B, Bz-ATP caused a rapid and long-lasting (>2 hr) statistically significant (t(5) = 10.37; p < 0.01) decrease in the fEPSP (fEPSP amplitude after Bz-ATP was 0.3 ± 0.05 of control amplitude; mean ± SEM; n = 6).

The P2X7 agonist Bz-ATP depressed mossy fiber fEPSPs but had no detectable effect on the presynaptic fiber volley. A, Averaged sample traces and plots of mossy fiber–CA3 synaptic responses recorded extracellularly from the stratum lucidum during one experiment at the indicated time points. L-CCG-I (20 μm) reversibly depressed the postsynaptic component of the field potential (indicated by ∗ in the first trace). Bz-ATP (30 μm) depressed the L-CCG-I-sensitive component of the fEPSP, and coapplication of L-CCG-I did not cause additional depression. B, Summary of separate experiments in which Bz-ATP was applied without preapplication of L-CCG-I. Averaged sample traces are shown before and after Bz-ATP application. Plot shows the mean values obtained from six slices. Bz-ATP depressed the mossy fiber fEPSP amplitude for >2 hr. C, Single plot and mean sample traces from a single experiment in which the field response was recorded in the presence of NBQX (20 μm) to monitor the presynaptic fiber volley. Bz-ATP had no effect on the presynaptic fiber volley. D, Summary of the effects of Bz-ATP on the fEPSP (n = 6) and the presynaptic fiber volley (n = 5). C, Control. ∗Statistical significance using a paired t test;p < 0.01. Calibration: A, 0.2 mV, 20 msec; B, 0.5 mV, 10 msec; C, 0.3 mV, 20 msec.
In separate experiments, the postsynaptic response was blocked by 2,3-dihydroxy-6-nitro-7-sulfonyl-benzo[f]quinoxaline (NBQX), and the presynaptic fiber volley was monitored after Bz-ATP application (Fig. 2C). There were no significant (t(4) = −1.13; p > 0.05) alterations in the presynaptic fiber volley as a result of Bz-ATP application (n = 5; summarized in Fig.2D). Furthermore, we visualized mossy fiber terminals using a laser scanning microscope to see whether Bz-ATP induced the uptake of YO-PRO-1 via pore dilation and cell lysis (Virginio et al., 1999). We did not observe any uptake of YO-PRO-1 after Bz-ATP application (data not shown; n = 2). These data suggest that activation of P2X7 receptors with Bz-ATP does not induce cytolysis of mossy fiber terminals.
Next, we obtained whole-cell voltage-clamp recordings from CA3 pyramidal neurons to determine whether Bz-ATP selectively depressed mossy fiber–CA3 synaptic transmission or had a postsynaptic effect on AMPA receptors. As shown in Figure 3, Bz-ATP significantly (t(5) = 22.36;p < 0.01) depressed the amplitude of voltage-clamped mossy fiber EPSCs (mossy fiber EPSC amplitude after Bz-ATP was 0.33 ± 0.04 of controls; mean ± SEM; n = 6) but had no statistically significant (t(4) = 2.02; p > 0.05) effect on associational–commissural EPSCs (associational–commissural EPSC amplitude after Bz-ATP was 0.81 ± 0.1 of controls; mean ± SEM; n = 5). Associational–commissural responses were evoked by stimulation of the stratum radiatum in the presence of L-CCG-I to block mossy fiber synapses. Bz-ATP also had no significant effect on the CA3 whole-cell conductance (308 ± 34 pS before vs 288 ± 60 pS after Bz-ATP) or holding current (65.8 ± 8.2 pA before Bz-ATP application vs 71.6 ± 7.9 pA after Bz-ATP). Therefore, P2X7 receptor activation selectively depressed mossy fiber synapses and had no direct postsynaptic effect on CA3 neurons.

Activation of presynaptic P2X7receptors with Bz-ATP selectively depressed synaptically evoked mossy fiber currents in CA3. A, Plots of mean whole-cell voltage-clamp recordings in CA3 pyramidal neurons after stimulation of the mossy fiber (MF) pathway (n = 6 slices) or the associational–commissural (A/C) pathway (n = 5 slices). Bz-ATP significantly depressed the amplitude of voltage-clamped mossy fiber EPSCs but had no significant effect on associational–commissural EPSCs. B, Summary of the data presented inA. ∗Statistical significance using a pairedt test; p < 0.01. C, Average sample traces from evoked responses after stimulation of the mossy fiber pathway or the associational–commissural pathway.Con, Control. Calibration: C, 200 pA, 50 msec; 250 pA, 50 msec.
Bz-ATP-induced synaptic depression was blocked by oxidized periodate-ATP
To further delineate P2X7 receptor involvement in this Bz-ATP-induced effect, we assessed the ability of Bz-ATP to induce synaptic depression of mossy fiber–CA3 fEPSPs in the presence of the nonselective P2Y antagonist reactive blue 2 (RB2; 30 μm) (Fig.4A) or the P2X1–3,5,6 receptor antagonist pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS; 10 μm) (Fig. 4B). As shown in Figure 4, both RB2 (n = 5) and PPADS (n= 4) failed to block the effect of Bz-ATP on mossy fiber fEPSPs (t(4) = 13.81, p< 0.01; and t(3) = 26.5,p < 0.01, respectively). These data suggest that Bz-ATP-induced synaptic depression was not mediated by the nonselective activation of P2Y receptors or postsynaptically located P2X1–3,5,6 receptors.

The selective P2X7 antagonist o-ATP blocked Bz-ATP-induced depression of the mossy fiber–CA3 synaptic responses. A, Plots and average sample traces of mossy fiber–CA3 synaptic responses recorded extracellularly from the stratum lucidum of CA3 in the presence of the nonselective P2Y antagonist RB2 (30 μm; n = 5 slices). RB2 failed to block the effect of Bz-ATP on mossy fiber fEPSPs. B, Similarly, the P2X1–3,5,6 receptor antagonist PPADS failed to block the effect of Bz-ATP on mossy fiber fEPSPs (n = 4 slices). C, However, preincubation of the slices with the P2X7 receptor antagonist o-ATP (100 μm; n = 5 slices) significantly reduced the magnitude of the Bz-ATP-induced depression compared with controls (n = 5 slices).D, Summary of data presented in A–C.Con, Control. ∗Significance using a pairedt test; p < 0.01. Calibration:A, 0.5 mV, 10 msec.
To determine whether Bz-ATP-induced synaptic depression was mediated by activation of P2X7 receptors, we applied Bz-ATP in the presence of the P2X7 receptor antagonist oxidized periodate-ATP (o-ATP; 100 μm) (Murgia et al., 1993; Visentin et al., 1999). Consistent with Bz-ATP acting at presynaptic P2X7 receptors, Bz-ATP-induced synaptic depression was potently inhibited by 2 hr of preincubation with the P2X7 antagonist o-ATP (in matched slices, fEPSP amplitude after Bz-ATP was 0.3 ± 0.05 of controls vs 0.81 ± 0.12 of controls in slices preincubated with o-ATP; mean ± SEM; n = 5 for both) (Fig. 4C). This P2X7-like pharmacological profile combined with our inability to detect a postsynaptic current in CA3 pyramidal cells suggests that Bz-ATP acted presynaptically at P2X7 receptors to mediate mossy fiber synaptic depression.
P2X7 receptor activation increased paired-pulse facilitation
If Bz-ATP depresses mossy fiber synaptic transmission by activating presynaptic P2X7 receptors, then Bz-ATP-induced depression should be associated with an increase in PPF (Regehr et al., 1994; Salin et al., 1996). We monitored PPF while recording whole-cell synaptic-evoked currents in CA3 neurons. Consistent with Bz-ATP activating presynaptic P2X7 receptors, we observed a significant (t(4) = −4.65; p < 0.01) increase in the ratio of the second EPSC amplitude to the first EPSC amplitude immediately after application of Bz-ATP (ratio before Bz-ATP was 1.74 ± 0.05; ratio after was 2.14 ± 0.09; mean ± SEM; n = 5 slices) (Fig.5). These data indicate that Bz-ATP decreased the probability of release at mossy fiber synapses.

Activation of presynaptic P2X7receptors increased mossy fiber PPF (50 msec). PPF was monitored by recording whole-cell synaptic currents evoked in CA3 neurons (n = 5 slices). A, Mean PPF ratio before and after application of Bz-ATP. ∗Statistical significance using a paired t test; p < 0.01. We observed a significant increase in the PPF ratio after Bz-ATP application, which is consistent with Bz-ATP acting on presynaptic P2X7 receptors. B, Average sample traces before and after Bz-ATP application. The lower trace on theright was rescaled so that the first current was the same size after Bz-ATP as it was in controls. Con, Control. Calibration: B, 200 pA, 50 msec; rescaled traces, 70 pA.
P2X7 receptor-mediated synaptic depression required activation of p38 MAP kinase
Recent evidence suggests that MAP kinase activity is potently activated by synaptic activity and is essential for some forms of synaptic plasticity (Impey et al., 1999). For example, extracellular signal-regulated kinase 1 (ERK1)/ERK2 MAP kinase activation is essential for the induction of long-term potentiation, and p38 MAP kinase activity is essential for the induction of long-term depression in CA1 of the hippocampus (Bolshakov et al., 2000). We have shown recently that activation of P2X7 receptors in cultured astrocytes leads to activation of p38 and ERK1/ERK2 MAP kinase (Panenka et al., 2001). To determine whether MAP kinase activity was necessary for the synaptic depression induced by Bz-ATP we preincubated the slices (2 hr) with the p38 MAP kinase inhibitor SB203580 (25 μm) or the ERK1/ERK2 MAP kinase inhibitor 2′-amino-3′-methoxyflavone (PD98059; 50 μm). Bz-ATP-induced synaptic depression of the L-CCG-I-sensitive mossy fiber–CA3 postsynaptic response was significantly (t(3) = 0.19; p > 0.05) blocked by inhibition of p38 MAP kinase activity with SB203580 (fEPSP amplitude after Bz-ATP was 0.93 ± 0.12 of control amplitude in slices preincubated with SB203580; n = 4) (Fig. 6A). In contrast, preincubation of the slices with the ERK1/ERK2 MAP kinase activity inhibitor PD98059 failed to block (t(3) = 8.27; p< 0.01) Bz-ATP-induced mossy fiber synaptic depression (fEPSP amplitude after Bz-ATP was 0.24 ± 0.11 of controls in slices preincubated with PD98059; n = 4) (Fig.6B). These data demonstrate that activation of p38 MAP kinase was necessary for P2X7receptor-mediated depression of mossy fiber–CA3 synaptic transmission.

P2X7 receptor-mediated mossy fiber synaptic depression required p38 MAP kinase activity. A, B, Plots of mean mossy fiber–CA3 synaptic responses recorded extracellularly from the stratum lucidum of CA3. A, Preincubation of the slices in the p38 MAP kinase inhibitor SB203580 (25 μm) completely blocked Bz-ATP-induced synaptic depression but had no effect on the L-CCG-I-induced depression of the mossy fiber fEPSP (n = 4 slices). B, Preincubation of the slices with the ERK1/ERK2 MAP kinase inhibitor PD98059 (50 μm) failed to have any effect on Bz-ATP-induced synaptic depression (n = 4 slices).C, Summary of data presented in A andB. Con, Control. ∗Significance using a paired t test; p < 0.01. Calibration: A, 0.5 mV, 10 msec.
Inhibitory effects of adenosine were not mediated through p38 MAP kinase
ATP and some of its analogs can be rapidly degraded into adenosine by the actions of ectonucleotidase (Dunwiddie et al., 1997; Cunha et al., 1998). Thus, ATP application can inhibit synaptic transmission indirectly through adenosine formation and the activation of presynaptic A1 receptors (Dunwiddie et al., 1997; Cunha et al., 1998;Cunha and Ribeiro, 2000; Dunwiddie and Masino, 2001). We could not use an A1 antagonist, such as 1,3-dipropylcyclopentylxanthine, because blocking A1 receptors results in persistent seizure activity in the CA3 region, making it impossible to record stable mossy fiber responses as reported previously (Thummler and Dunwiddie, 2000). Therefore, to determine indirectly whether Bz-ATP-induced mossy fiber–CA3 synaptic depression was mediated through degradation of Bz-ATP into adenosine, we tested whether adenosine inhibits synaptic transmission through p38 MAP kinase activity. As shown in Figure7, preincubation of the slices with SB203580 (25 μm) blocked Bz-ATP-induced mossy fiber synaptic depression (n = 4) but failed to have any effect on adenosine-mediated (30 μm) mossy fiber synaptic depression (n = 4). In slices preincubated with SB203580, the fEPSP amplitude after Bz-ATP was 0.95 ± 0.15 of controls, versus 0.15 ± 0.09 of controls in adenosine (F(1,6) = 21.58,p < 0.01). Therefore, adenosine does not exert its inhibitory actions through p38 MAP kinase, and the presynaptic actions of Bz-ATP cannot be explained by the degradation of Bz-ATP into adenosine.

The p38 antagonist SB203580 blocked the actions of Bz-ATP but not the inhibition by adenosine (30 μm).A, Plot of an extracellular recording in which the p38 MAP kinase inhibitor SB203580 blocked the Bz-ATP-induced mossy fiber synaptic depression but failed to block the adenosine-induced inhibition of the mossy fiber fEPSP. B, SB203580 differentially affected the depression induced by Bz-ATP and adenosine. Therefore, adenosine does not exert its inhibitory actions through p38 MAP kinase, and the presynaptic actions of Bz-ATP cannot be explained by the degradation of Bz-ATP into adenosine by ectonucleotidase activity. ∗Statistical significance using a one-way ANOVA;p < 0.01.
DISCUSSION
The results of the present study demonstrate that activation of presynaptic P2X7 receptors results in the inhibition of neurotransmission at mossy fiber–CA3 synapses through a p38 MAP kinase-signaling pathway. First, we have used immunocytochemistry to demonstrate that P2X7receptors are abundant on presynaptic terminals of mossy fiber synapses in the rat hippocampus. Immunocytochemistry with a specific P2X7 antibody resulted in the labeling of small terminal-like puncta throughout the hippocampus. P2X7 immunoreactivity was particularly dense throughout the termination zones of hippocampal mossy fibers, where it was completely colocalized with the presynaptic marker syntaxin 1A/B but not the dendritic marker MAP-2 (Fig. 1). Syntaxin 1A is known to be present in the presynaptic mossy fiber terminals, and syntaxin 1B is present in the mossy fiber axons (Ruiz-Montasell et al., 1996). As demonstrated in Figure 1, all of the observed P2X7 immunoreactivity in the stratum lucidum was colocalized with the syntaxin 1A labeling of the presynaptic terminal. These data demonstrate that P2X7 receptors are located presynaptically in the stratum lucidum of the rat hippocampus. The specific presynaptic P2X7 receptor localization shown here is in contrast to the postsynaptic location of other known P2X receptors in the hippocampus of the rat (e.g., P2X2, P2X4, and P2X6) (Le et al., 1998; Rubio and Soto, 2001). These postsynaptically located receptors are likely to contribute to the increase in excitation that is observed in the hippocampus after application of low doses of ATP (Wieraszko and Seyfried, 1989).
Consistent with their presynaptic localization, activation of P2X7 receptors with Bz-ATP completely depressed the L-CCG-I-sensitive mossy fiber–CA3 synaptic response in extracellular field recordings. However, no direct effects of Bz-ATP on postsynaptic CA3 pyramidal neurons were observed when the conductance and holding current were monitored during whole-cell voltage-clamp recordings. Furthermore, we found no significant effect of Bz-ATP on AMPA receptor-mediated associationalcommissural synaptic transmission in CA3. This observation is consistent with our conclusion that Bz-ATP selectively activates presynaptic P2X7 receptors and suggests that at this concentration (30 μm), Bz-ATP did not activate other known postsynaptic P2X receptors (e.g., P2X2, P2X4, and P2X6). Although our conclusions support the involvement of P2X7receptors in presynaptic depression, the possible contribution of P2X4 cannot be totally eliminated. The enhancement of PPF during the Bz-ATP-induced synaptic depression is also consistent with a presynaptic site of action (Regehr et al., 1994;Salin et al., 1996) similar to what has been observed during mGluR-mediated depression in CA1 (Fitzjohn et al., 2001). Bz-ATP-induced synaptic depression was not blocked by the P2Y receptor antagonist RB2 (30 μm) or the P2X1–3,5,6 antagonist PPADS (10 μm). Other P2X receptors are antagonized by PPADS at this concentration, whereas P2X7 receptors are not (Surprenant et al., 1996). However, Bz-ATP-mediated synaptic depression required P2X7 receptor activation, because little or no synaptic depression was observed when the slices were preincubated with the irreversible P2X7 receptor antagonist o-ATP (Fig. 4) (Murgia et al., 1993; Visentin et al., 1999). Bz-ATP-induced synaptic depression also cannot be explained by the degradation of Bz-ATP into adenosine by local ectonucleotidases, because adenosine-mediated synaptic inhibition was not blocked by p38 MAP kinase inhibition, whereas the actions of Bz-ATP were blocked (see below).
Mossy fiber synapses contain vesicular ATP, and synaptosomes prepared from mossy fiber synapses release ATP in a Ca2+-dependent manner in response to K+-induced depolarization (Terrian et al., 1989). However, it is not known whether ATP is normally released from mossy fiber synapses or whether P2X7 receptors are normally activated during the evoked release of neurotransmitters from mossy fiber terminals. It is possible that presynaptic mossy fiber P2X7 receptors might only be activated during intense periods of mossy fiber activity, such as that observed during tetanus or seizure, when ATP release might reach levels high enough to activate P2X7 receptors. Therefore, P2X7 receptors might play an important role in limiting synaptic transmission when mossy fiber synaptic transmission is unusually high.
The results of the present study demonstrate that activation of p38 MAP kinase is necessary for P2X7 receptor-mediated depression of mossy fiber–CA3 synaptic transmission. Maruyama et al. (2000) have demonstrated recently that p38 MAP kinase is abundant in the terminals of mossy fiber synapses. As shown in Figure 6, Bz-ATP-induced synaptic depression was completely blocked by preincubation of the slices with the p38 MAP kinase activity inhibitor SB203580 but not the ERK1/ERK2 MAP kinase activity inhibitor PD98059. The presynaptic mechanism by which P2X7receptor-dependent p38 MAP kinase activity depresses mossy fiber synaptic transmission remains to be determined. However, recent evidence suggests that p38 MAP kinase activity is necessary for the inhibition of N-type calcium currents in neuroblastoma cells after bradykinin application (Wilk-Blaszczak et al., 1998). Therefore, it is possible that P2X7 receptor-dependent p38 MAP kinase activity depresses mossy fiber synaptic transmission through inhibition of calcium channels. However, mossy fiber terminals exhibit predominantly P-type calcium channel-dependent evoked neurotransmitter release and contain few N-type channels (Castillo et al., 1994).
Recent evidence suggests that MAP kinase activity is potently activated by synaptic activity and is essential for some forms of neuronal plasticity (Impey et al., 1999; Bolshakov et al., 2000). For example, translocation of ERK1/ERK2 MAP kinase to the nucleus of the presynaptic neuron is essential for long-term facilitation in Aplysianeurons (Martin et al., 1997), and p38 MAP kinase is essential for the induction of mGluR receptor-dependent, long-term depression in CA1 of the hippocampus (Bolshakov et al., 2000). Interleukin-1β has also been shown to increase p38 activation and modify long-term potentiation in perforant path synapses (Vereker et al., 2000). In the present study, we have shown that the rapid and reversible depression of mossy fiber synaptic transmission by the mGluR agonist L-CCG-I is unaffected by preincubation of the slices with the p38 MAP kinase inhibitor SB203580. Therefore, the mGluR-induced inhibition of mossy fiber synapses is apparently not mediated by p38 as it is in CA1.
In conclusion, we have provided evidence that P2X7 receptor subunits are abundant on presynaptic terminals of mossy fiber synapses in the rat hippocampus. Activation of these presynaptic P2X7 receptors with the P2X7 agonist Bz-ATP produced a rapid and long-lasting synaptic inhibition at mossy fiber–CA3 synapses. This presynaptic inhibition was mediated by the activation of p38 MAP kinase, because it was not observed when the slices were preincubated with a p38 MAP kinase inhibitor. Therefore, the results of the present study demonstrate that unlike any other member of the P2X receptor family, P2X7 receptors can decrease neurotransmitter release at mossy fiber–CA3 synapses by activating p38 MAP kinase in the presynaptic terminal.
Footnotes
This work was supported by the Canadian Institutes for Health Research (CIHR). B.A.M. is an Alberta Heritage Foundation for Health Research and CIHR Senior Scientist. T.B.B. was supported by studentships from Libin Neuroscience Canada Foundation Alberta, the Natural Sciences and Engineering Research Council of Canada, and the Alberta Heritage Foundation for Medical Research. We thank Denise Feighan and Naili Liu for their technical assistance.
Correspondence should be addressed to Brian A. MacVicar, Department of Physiology and Biophysics, Faculty of Medicine, University of Calgary, 3330 Hospital Drive Northwest, Calgary, Alberta, T2N 4N1 Canada. E-mail: macvicar{at}ucalgary.ca.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}