Article Text

Abstract
BACKGROUND The development of colitis in interleukin 10 (IL-10) deficient mice, together with the known anti-inflammatory and immunomodulatory properties of this cytokine have prompted consideration of IL-10 as a treatment for inflammatory bowel disease (IBD). However, studies using hrIL-10 in IBD models have yielded inconsistent results.
AIMS To examine the therapeutic potential of overexpressing the IL-10 gene before and after the induction of experimental colitis in rats.
METHODS Gene transfer was achieved by intraperitoneal injection of non-replicating human type 5 adenovirus bearing the IL-10 gene, either 24 hours before or one hour after intrarectal administration of dinitrobenzene sulphonic acid in rats. Colonic damage and inflammation was assessed macroscopically and by measuring myeloperoxidase activity and leukotriene B4 concentrations.
RESULTS Gene transfer increased IL-10 protein in serum for up to six days. IL-10 gene transfer prior to colitis improved colitis macroscopically and histologically, and significantly reduced colonic myeloperoxidase activity and leukotriene B4 concentrations. In contrast, IL-10 gene transfer after the onset of colitis had no beneficial effect.
CONCLUSIONS Gene therapy using an adenovirus-IL-10 construct was successful in preventing but not in reversing experimental colitis in the rat.
- gene therapy
- colitis
- interleukin 10
- adenovirus vector
- leukotrienes
- inflammatory bowel disease
- maintenance therapy
Statistics from Altmetric.com
- gene therapy
- colitis
- interleukin 10
- adenovirus vector
- leukotrienes
- inflammatory bowel disease
- maintenance therapy
There is increasing awareness of the role of cytokines in the pathophysiology of inflammatory bowel disease (IBD) and in their therapeutic potential.1 2 In the evaluation of the latter, an alternative to the administration of recombinant cytokine is the transfer of its gene for overexpression in the inflamed gut.3 Interleukin 10 (IL-10) exhibits both immunomodulatory as well as anti-inflammatory properties4; chronic enterocolitis develops spontaneously in mice lacking the IL-10 gene5 associated with increased concentrations of interferon α (IFN-α), tumour necrosis factor α (TNF-α), IL-1β, and IL-6 in the intestine.6 These findings indicate the therapeutic potential of IL-10; the preliminary finding that there is impaired IL-10 production by lamina propria T cells in human IBD7 suggests that it is an appropriate therapeutic candidate for IBD.
While the initial reports of amelioration of Crohn's disease following administration of human recombinant IL-10 (hrIL-10)8 9have been encouraging, some concerns remain. Firstly, the reported improvement of Crohn's disease following IL-10 treatment was based largely on clinical parameters with no indication of the effects of IL-10 on tissue injury and inflammation.8 9 Secondly, studies on the efficacy of IL-10 treatment in reducing inflammation in animal models of IBD have yielded conflicting results. While it has been shown that hrIL-10 improved colonic inflammation and tissue damage in the streptococcal peptoglycan-polysaccharide model of colitis in rats,10 little or no effect of IL-10 treatment has been reported in the TNBS (trinitrobenzene sulphonic acid) model of colitis.11 The reason for the apparent lack of effect of IL-10 in the TNBS colitis model is not fully understood. Firstly, it has been shown that the timing of IL-10 therapy relative to the onset of colitis is critical.10 Also, the poor efficacy of the administered cytokine may reflect the fact that hrIL-10 is rapidly cleared from the circulation after in vivo administration, limiting its availability to target tissues.12
To overcome the problem of limited bioavailability of hrIL-10 we used gene transfer technology to achieve overproduction of IL-10 in vivo in a similar model of colitis in rats. In addition, we sought to clarify the optimum therapeutic role of IL-10 by administering the gene prior to, or shortly after the induction of colitis. Because of the limited availability of TNBS, we have used dinitrobenzene sulphonic acid (DNB) which induces a colitis that is very similar to that induced by TNBS.13 14 Our results show that using this paradigm IL-10 successfully ameliorated colonic inflammation and tissue injury when the adeno-IL-10 construct was transferred before the induction of colitis. In contrast the adeno-IL-10 construct had little or no effect when administered after induction of the colitis. These results suggest that the therapeutic benefit of IL-10 may be more prominent in the prevention of relapses than in the suppression of established inflammation.
Materials and methods
ANIMALS
Specific pathogen free, male Sprague Dawley rats (180–200 g; Charles River Laboratories, Montreal, Canada) were kept in filtered cages in a level B clean room under controlled temperature and light-dark cycle (14:10 hours). All rats had ad libitum access to food and water before and during the study. All experimental procedures were approved by the Animal Care Committee at McMaster University and were in accordance with the guidelines of the Canadian Council on the Use of Laboratory Animals.
ADENOVIRUS VECTORS AND MATERIALS
Human type 5 adenovirus (Ad5) lacking the E1 region of the genome which regulates replication were transfected with one of the following cDNAs: murine IL-10 (Ad5IL-10), Escherichia coli LacZ (Ad5LacZ), or luciferase (Ad5Luc3) according to procedures published in detail elsewhere.15 The following antibodies used in the cytokine enzyme linked immunosorbent assays (ELISAs) were purchased from Pharmigen (San Diego, California, USA): rat antimouse IL-10, biotinylated rat antimouse IL-10, and recombinant murine IL-10. Nuncimmuno ELISA plates (Maxisorp TM) were obtained from Gibco BRL (Burlington, Canada). Rat anti-LTB4 antiserum was obtained from BIOTRAK (Amersham Canada, Oakville, Ontario, Canada). Unless otherwise stated, all other materials were purchased from Sigma Chemical Co. (St Louis, Missouri, USA).
INDUCTION OF COLITIS
Colitis was induced using a previously described technique,13 but substituting DNB for TNBS.14In brief, to induce colitis, each rat was lightly anaesthetised with Fluotane (halothane BP; Wyeth-Ayerst Canada, Inc., Montreal, Canada); 30 mg of DNB dissolved in 0.25 ml of 50% ethanol was instilled into the distal colon of each animal using a PE-50 cannula (Becton Dickinson Labware, Lincon Park, New Jersey, USA). Control rats were injected intracolonically using the same procedure with either normal saline (0.9% NaCl wt/vol) or 50% ethanol. DNB treated and control rats were kept in separate cages over the course of the study. Murine recombinant IL-10 was used for the following reasons: there is greater than 90% homology between the amino acid sequence of rat and mouse IL-1016; and there are demonstrable biological effects of mrIL-10 on rat cells in vitro.17
EXPERIMENTAL PROTOCOL
Ad5LacZ or Ad5IL-10 were suspended in 1 ml of Hanks's balanced salt solution (HBSS; pH 7.4) and injected into the peritoneal cavity either 24 hours before or one hour after the DNB enema. The route and time course of Ad5 administration was based on a previous study.15 We performed preliminary studies in order to identify the most effective dose of Ad5IL-10 by administering titres of Ad5IL-10 between 5 × 108 and 1 × 1010plaque forming units (pfu). Titres of 5 × 108 pfu and 109 pfu were both associated with improvement of colitis, while the titre of 1010 pfu exerted either no effect or worsening of colitis, sometimes with mortality. Thus 1 × 109 pfu of Ad5IL-10 were used in subsequent experiments.
On day 6 post-DNB, 1 ml of blood was obtained by cardiac puncture under anaesthesia and the rats were sacrificed by cervical dislocation. The colon was removed, rinsed in PBS, and opened lengthwise for a macroscopic evaluation of damage.
IN VIVO GENE TRANSFER
Assessment of transgene distribution
Ad5Luc3 was used to assess quantitative distribution of transgene expression after intraperitoneal Ad5 administration; 109pfu of Ad5Luc3 were suspended in 1 ml of HBSS and injected into the peritoneal cavity. The rats were sacrificed 24 hours later. The intra-abdominal organs were removed and placed in 5 or 10 ml of a lysis buffer (100 mM potassium phosphate containing 1 mM phenylmethylsulphonyl fluoride (PMSF) and 10 μg/ml aprotinin), homogenised, and sonicated on ice. A 1 ml sample of the homogenate was transferred to a 1.5 ml centrifuge tube and assayed for luciferase activity as described previously.18
Assessment of transgene expression
mrIL-10 concentrations were determined in 50 μl serum collected from rats 24 hours, and six days after intraperitoneal administration of Ad5IL-10 using a commercially available sandwich ELISA protocol (Pharmigen). An ELISA amplification system (Gibco BRL) was used to enhance the reaction. The monoclonal antibodies directed against murine IL-10 did not react with rat IL-10, thus we were able to determine specifically the amount of murine IL-10 produced after Ad5IL-10 injection into rats. A rmIL-10 was used to calculate IL-10 serum concentrations in all Ad5 infected rats.
ASSESSMENT OF COLONIC DAMAGE
Colonic damage was assessed macroscopically and histologically using a modification of previously published criteria.19Briefly, the macroscopic criteria were based on the following: the presence of adhesions between the colon and other intra-abdominal organs, the consistency of colonic faecal material (as an indirect marker of diarrhoea), thickening of the colonic wall, presence and extension of hyperaemia, and macroscopic mucosal damage (assessed with a ruler). Microscopic criteria for damage and inflammation were assessed by light microscopy on haematoxylin and eosin stained histological sections obtained from whole gut specimens taken from a region of the inflamed colon immediately adjacent to the gross macroscopic damage. Histological criteria included: degree of mucosal architectural changes, cellular infiltration, external muscle thickening, presence of crypt abscess, and goblet cell depletion. All the parameters of macroscopic and histological damage were recorded and scored for each rat by two different observers who were blinded to the treatment conditions.
ASSESSMENT OF COLONIC MYELOPEROXIDASE ACTIVITY
Tissues (200–400 mg) for assessment of myeloperoxidase (MPO) were removed from the area of gross injury, snap frozen in liquid nitrogen, and assayed within seven days using a previously described method.20 MPO was expressed as U/mg of tissue, where 1 U corresponds to the activity of enzyme required to degrade 1 μmol of hydrogen peroxide in one minute at 24°C. MPO is a granule associated enzyme primarily contained in neutrophils and other cells of myeloid origin and is widely used as a marker of intestinal inflammation.21
ASSESSMENT OF LTB4 COLONIC CONCENTRATIONS
Leukotriene B4 (LTB4) concentrations were assessed on colonic tissue using an EIA protocol from BIOTRAK (Amersham Canada, Oakville, Ontario, Canada). Samples of colon were snap frozen and stored at −70°C until assayed. On thawing, samples were weighed, homogenised, and centrifuged at 4°C. A 50 μl aliquot of supernatant was then added to wells in triplicate. A 50 μl aliquot of rat anti-LTB4 was added to each sample and left to incubate at room temperature for two hours before adding LTB4-horseradish peroxidase conjugate. After one hour the plates were washed and TMB enzyme substrate (BIOTRAK; Amersham Canada, Oakville, Ontario, Canada) was added; the plates were incubated for 30 minutes before the absorbency was read at 620 nm.
DATA EXPRESSION AND STATISTICAL ANALYSIS
All the studies involved at least six rats and results were expressed as mean (SEM); n refers to the number of animals used for each experiment. Statistical significance was determined at p<0.05, using Student's t test for comparison of two means, or one way analysis of variance (ANOVA) for the comparison of more than two means, followed by Bonferroni's multiple range analysis.
Results
DISTRIBUTION OF LUCIFERASE ACTIVITY IN Ad5Luc3 TREATED RATS
Luciferase activity was evident in all intra-abdominal organs examined, with maximum expression detected in the liver (45%) and spleen (30%). The colon expressed 15% of the total luciferase activity (fig 1). This profile was not altered in the presence of DNB colitis (data not shown).

Luciferase activity following administration of Ad5Luc3 or Ad5IL-10. Results represent mean (SEM) from each group (n=8 rats for each group).
SERUM IL-10 CONCENTRATIONS IN Ad5IL-10 TREATED RATS
Circulating IL-10 concentrations rose 13-fold 24 hours after Ad5IL-10 treatment (fig 2). A small but significant increase in IL-10 serum concentrations was evident up to six days post-Ad5IL-10. Induction of colitis either 24 hours prior to, or one hour after the intraperitoneal administration of Ad5IL-10 did not significantly alter serum IL-10 concentrations (not shown). Administration of a smaller titre of Ad5IL-10 (5 × 108 pfu) was associated with undetectable IL-10 serum concentrations.

Serum IL-10 protein concentrations after administration of HBSS, Ad5LacZ, or Ad5IL-10. Results represent mean (SEM) from each group (HBSS, n=4; Ad5LacZ, n=4; Ad5IL-10, n=8). **Significantly different from controls (p<0.01).
EFFECT OF ADENOVIRUS INFECTION OF THE COLON
Infection with adenovirus, either as Ad5LacZ or Ad5IL-10, had no effect on the macroscopic or histological appearance of the colon of control rats compared with rats injected intraperitoneally with the HBSS vehicle (not shown).
DNB COLITIS IN THE ABSENCE OF Ad5IL-10 TREATMENT
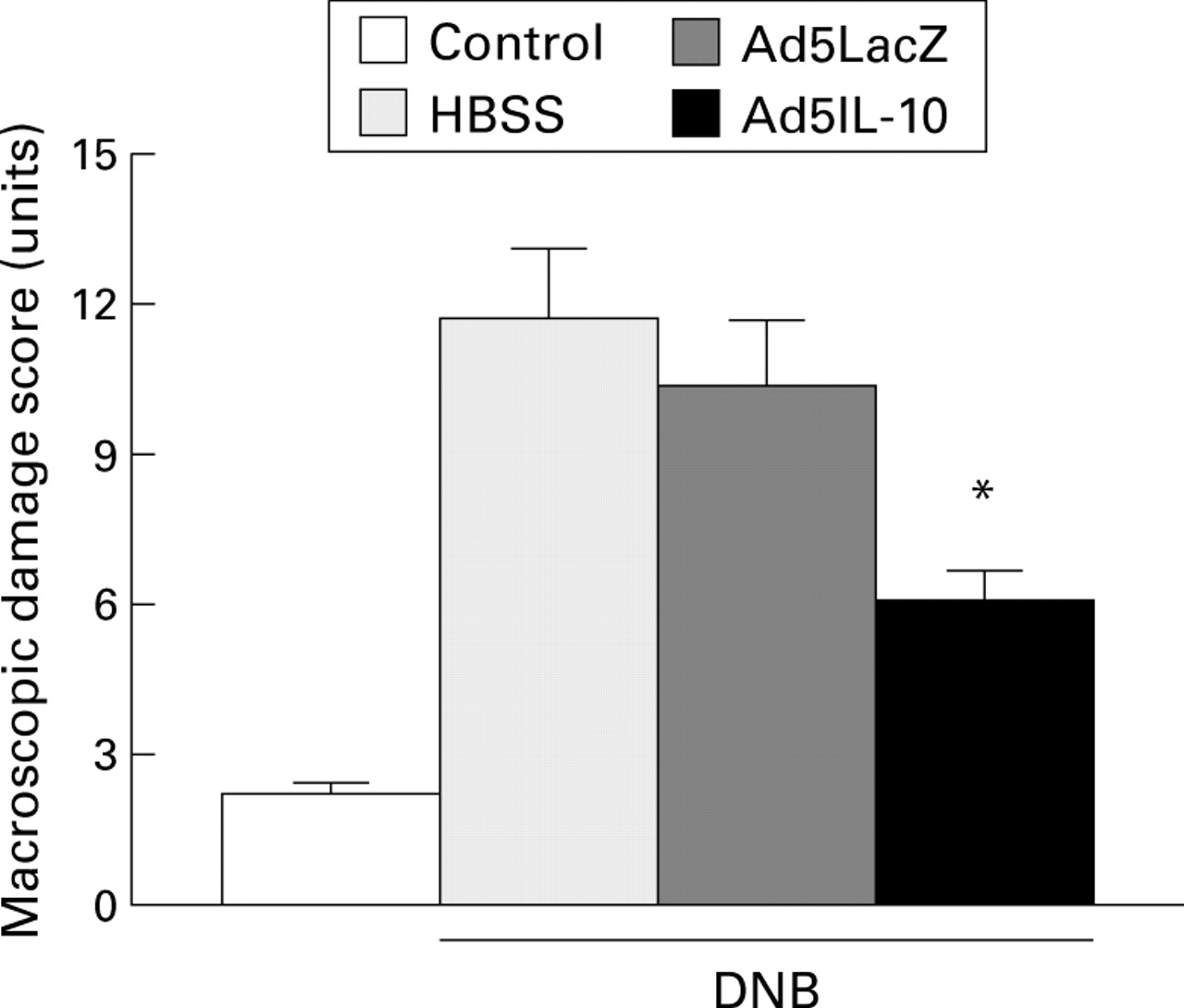
At day 6 post-DNB, the distal colon was thickened and ulcerated with evident areas of transmural inflammation. Adhesions were often present and the bowel was occasionally dilated. The colitis was characterised by an intense granulocyte infiltrate extending throughout the mucosa and submucosa, and often involving the muscularis propria which invariably appeared thickened. There was a greater than sixfold increase in both macroscopic and microscopic damage scores over the ethanol treated control animals (11.7 (1.4) versus 2.2 (0.2), p<0.01 for macroscopic damage; and 8.9 (1.1) versus 1.0 (0.4) for microscopic damage, p<0.01). Colitis was accompanied by a 15-fold increase in MPO activity compared with the ethanol controls (8.2 (1.7) versus 0.5 (0.2), p<0.01) and a 17-fold increased in LTB4 concentrations in the colon on day 6 post-DNB (table 2). The administration of Ad5LacZ had no effect on the morphology of the inflamed colon (table 1), or on MPO activity or LTB4 concentrations.
Effect of Ad5IL-10 treatment on colonic leukotrience (LT) B4 concentrations 24 hours prior to colitis
Effect of Ad5IL-10 treatment on colonic damage and inflammation one hour after the induction of colitis
EFFECT OF PRIOR Ad5IL-10 TREATMENT ON COLITIS
Treatment with Ad5IL-10 prior to the induction of colitis caused a significant 40% improvement in macroscopic damage compared with rats pretreated with the control adenovirus Ad5LacZ (fig 3). This was accompanied by less microscopic damage and a reduced inflammatory cell infiltrate (fig 4). Similar results were obtained when a smaller titre of Ad5IL-10 (5 × 108 pfu) was administered; macroscopic damage scores fell from 10.3 (0.9) to 7.1 (0.8) (p<0.05).
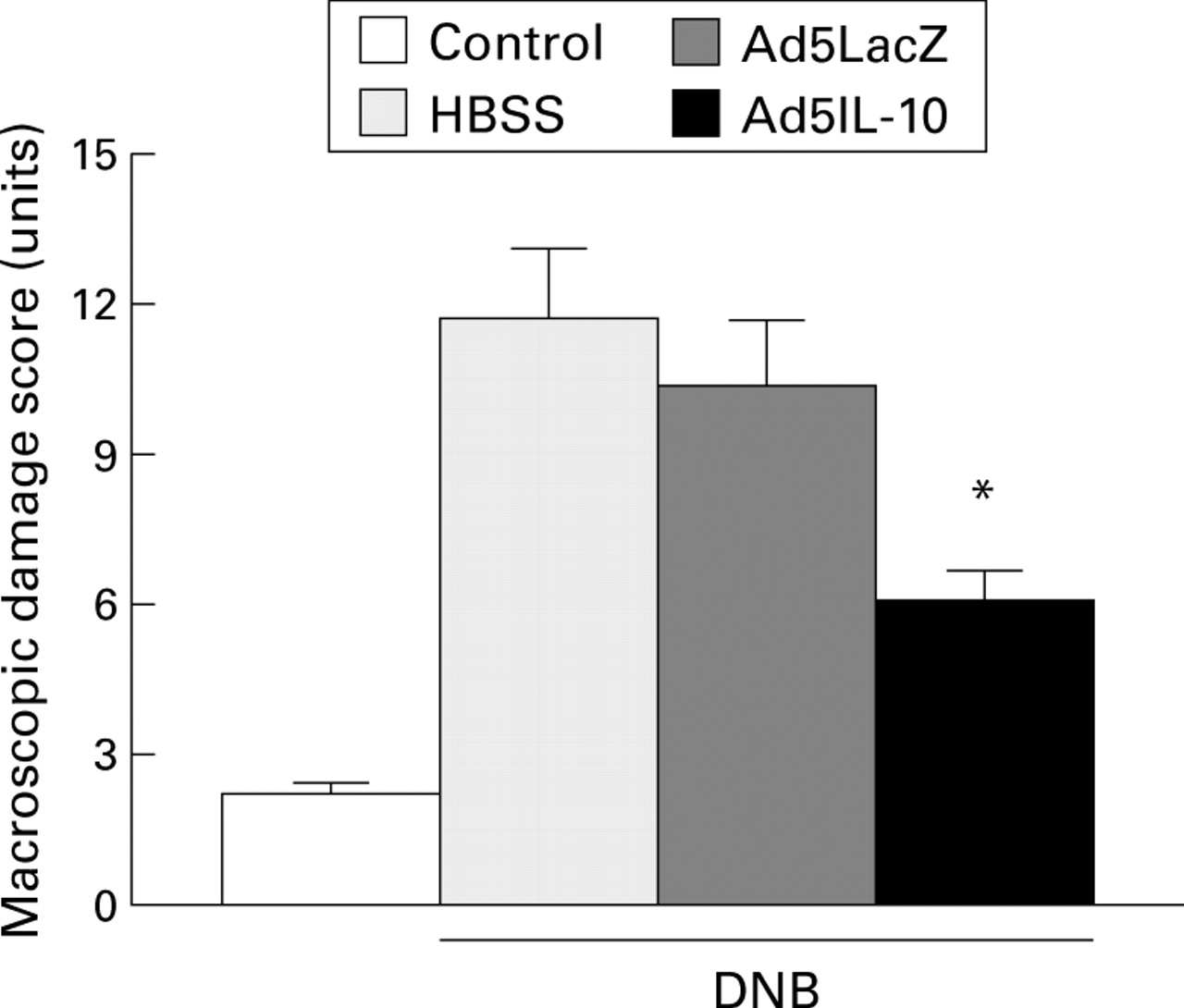
Macroscopic damage scores six days after dinitrobenzene sulphonic acid (DNB) induced colitis. Results represent mean (SEM) from each group (ethanol group, n=6; HBSS, Ad5LacZ, and Ad5IL-10, n=8). *Significantly different from Ad5LacZ (p<0.05).

Histological damage scores six days after dinitrobenzene sulphonic acid (DNB) induced colitis. Results represent mean (SEM) from each group (ethanol group, n=6; HBSS, Ad5LacZ, and Ad5IL-10, n=8). *Significantly different from Ad5LacZ (p<0.05).
As fig 5 shows, in rats treated with Ad5IL-10 24 hours prior to the induction of colitis, there was a significant decrease in MPO activity compared with colitis rats pretested with the control adenovirus Ad5LacZ (from 7.2 (1.4) to 2.5 (0.5) U/g, p<0.01). Ad5IL-10 treatment 24 hours prior to colitis significantly decreased LTB4 colonic concentrations in comparison to rats treated with Ad5LacZ (table2).

Myeloperoxidase activity six days after dinitrobenzene sulphonic acid (DNB) induced colitis. Results represent mean (SEM) from each group (ethanol group, n=6; HBSS, Ad5LacZ, and Ad5IL-10, n=8). *Significantly different from Ad5LacZ (p<0.05).
EFFECT OF Ad5IL-10 ADMINISTERED AFTER THE INDUCTION OF COLITIS
As shown in table 1, treatment with Ad5IL-10 one hour after the induction of colitis caused only a 31% decrease in MPO activity compared with rats treated with Ad5LacZ. However, this was not accompanied by any significant improvement in either the macroscopic or microscopic damage scores, or in colonic LTB4 concentrations (table 1, fig 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of pretreatment of rats with Ad5IL-10 gene transfer on leukotriene (LT) B4 concentrations in the distal colon of rats six days after dinitrobenzene sulphonic acid (DNB) induced colitis. Results represent mean (SEM) from each group (ethanol group, n=6; HBSS, Ad5LacZ, and Ad5IL-10, n=8). *Significantly different from Ad5LacZ (p<0.05).
Discussion
The intraperitoneal administration of the replication deficient adenovirus-IL-10 construct resulted in an increase in serum IL-10 concentrations that could be detected for at least six days, thus overcoming the constraint imposed by the short half life of exogenous rmIL-1012 and confirming results obtained by Xinget al.15
Tissue distribution of the virus in the abdomen was extensive. In addition to the anticipated expression in the liver,22 we also showed significant transgene expression in the distal colon—the site of DNB induced injury. These findings suggest that IL-10 may exert its biological effects at both a local and systemic level. Although we have not identified the infected cells, our recent work has suggested a propensity of the virus to infect the subserosal layers.23
Targeting of the gene to the mucosa of the inflamed colon would be ideal. In the past, attempts to deliver adenoviruses to the gastrointestinal mucosa in vivo were successful only in immunocompromised animals,24 in the neonatal intestine,25 or following selective intra-arterial injection of the adenovirus vector.26 The latter approach, however, involves laparotomy and bowel manipulation and is technically challenging; in our hands it exposed the rats to an unacceptably high mortality rate in conjunction with the administration of DNB. Furthermore, intra-arterial injection may produce undesired effects on endothelial cells.26 This may be critical in the context of the present study as IL-10 has been shown to induce expression of the proinflammatory cytokine IL-6.27
The beneficial effect of Ad5IL-10 was shown on all the parameters studied, including macroscopic and microscopic indices as well as on MPO activity and LTB4 concentrations in the inflamed colon. Our results indicate that the therapeutic effects of Ad5IL-10 are attributable to the cytokine itself rather than a consequence of the virus infection per se, as the control vector (Ad5LacZ) had no significant effect on the parameters studied. Indeed in another model, a replication deficient adenovirus vector (Ad5CMV-LacZ) potentiated inflammation in the airways.28
As our studies were performed during the acute phase of colitis (one to six days post-colitis) at a time when immune mediated mechanisms presumably play a minor role, we focused our attention on MPO activity. This closely reflects the tissue accumulation of granulocytes21 and previous studies have shown that the infiltrate in acute TNB colitis is comprised almost exclusively of neutrophils.29 Our results show that Ad5IL-10 treatment prior to the induction of colitis significantly suppressed MPO activity in the colon and that this was associated with reduced colonic damage at a macroscopic and microscopic level. These results are in keeping with studies in other models, such as that of antigen induced acute inflammation in the airways, which show that IL-10 is a potent suppressor of tissue granulocyte recruitment.30 Krakauer showed that IL-10 downregulates ICAM-1 and VCAM-1 expression on human endothelial cells.31 IL-10 also reduces both proinflammatory cytokine32 and chemokine33production by activated neutrophils and enhances neutrophil apoptosis, both in humans34 and in animal models of inflammation.35
Ad5IL-10 administration prior to the development of colitis significantly decreased colonic concentrations of LTB4. There is clear evidence from animal models of colitis indicating that leukotrienes in general, and LTB4 in particular, play a critical role in amplification of the inflammatory response.19 36 Wallaceet al have shown that successful inhibition of leukotriene synthesis results in substantial improvement of colonic damage induced by TNBS19 and that intrarectal administration of LTB4 enhanced colonic damage and inflammation produced by ethanol.36 Whether reduced LTB4 colonic concentrations reflected a direct effect of IL-10 on LTB4 synthesis or simply the reduction of colonic granulocytes cannot be answered by the results of our study. Indeed, previous studies have shown that in experimental colitis neutrophils are the major source of LTB4.36 However, regardless of the source, the reduction in colonic LTB4 almost certainly contributed to attenuation of colitis in view of the potent chemotactic activity of this metabolite.
This study emphasises the importance of the timing of IL-10 administration. Our results show that treatment of rats with Ad5IL-10 after the induction of colitis did not produce the macroscopic and microscopic improvement as was seen following pretreatment with the construct. We consider it unlikely that a higher titre of Ad5IL-10 would have changed this impression as our results suggest that we were using a maximally effective titre of the virus; similar results were obtained using a small titre of Ad5IL-10 (5 × 108 pfu). We were reluctant to increase the titre of Ad5IL-10, as in preliminary studies we found that high Ad5IL-10 titres (1010 pfu) showed either no effect or worsening of colitis which was sometimes fatal. This is in agreement with previous published findings showing that high systemic concentrations of IL-10 can exert proinflammatory activity in the non-inflamed murine intestine.15
The use of adenovirus vectors to treat inflammatory conditions of the gut is in its infancy, but the potential benefit of transient overexpression of a modulatory or anti-inflammatory cytokine is recognised.37 Two recent reports, suggesting that adenovirus vectors may be delivered directly to the gastrointestinal tract, provide a basis for some optimism.38 39 The results of the present study, showing that transient overexpression of IL-10 by prior administration of the vector attenuates colitis, should be viewed in light of these recent development in gene delivery to the gut, rendering gene therapy a viable option in the future treatment of inflammatory bowel disease.
Acknowledgments
Grant support was from the Medical Research Council of Canada.
References
Footnotes
- Abbreviations used in this paper:
- DNB
- dinitrobenzene sulphonic acid
- IBD
- inflammatory bowel disease
- IFN
- interferon
- IL
- interleukin
- LK
- leukotriene
- MPO
- myeloperoxidase
- TNF
- tumour necrosis factor