


Abstract
Pro-inflammatory cytokines play a critical role in the development of autoimmune and inflammatory diseases. Targeting the cytokine environment has proven efficient for averting inflammation. In this study, we reported that 6-[4-(aminomethyl)-2-chlorophenoxyl]benzo[c][1,2]oxaborol-1(3H)-ol (AN3485), a benzoxaborole analog, inhibited TLR2-, TLR3-, TLR4-, and TLR5-mediated TNF-α, IL-1β, and IL-6 release from human PBMCs and isolated monocytes with IC50 values ranging from 18 to 580 nM, and the inhibition was mediated at the transcriptional level. Topical administration of AN3485 significantly reduced PMA-induced contact dermatitis and oxazolone-induced delayed-type hypersensitivity in mice, indicating its capability of penetrating skin and potential topical application in skin inflammation. Oral administration of AN3485 showed dose-dependent suppression of LPS-induced TNF-α and IL-6 production in mice with an ED90 of 30 mg/kg. Oral AN3485, 35 mg/kg, twice a day, suppressed collagen-induced arthritis in mice over a 20-day period. The potent anti-inflammatory activity in in vitro and in vivo disease models makes AN3485 an attractive therapeutic lead for a variety of cutaneous and systemic inflammatory diseases.
Introduction
Pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 have proven to be good therapeutic targets for rheumatoid arthritis (RA), psoriatic arthritis, and psoriasis (Kopf et al., 2010). Identification of the mechanism whereby these cytokines are induced will not only help to understand the pathogenesis of these diseases but also provide a potential target for drug development. The association of aberrant Toll-like receptor (TLR) responses with these diseases (Lai and Gallo, 2008; Miller, 2008; Huang and Pope, 2009; Carrasco et al., 2011) and the fact that activation of certain TLRs induces TNF-α, IL-1β, and IL-6 production allow speculation that TLR signaling may play a role in disease initiation and/or progression.
TLRs were initially identified as receptors for exogenous pathogen-associated molecular patterns (PAMPs) of microbial origin (Poltorak et al., 1998) and act as the first line of defense against bacterial or viral infection (Dinarello, 2009). In addition to PAMPs, TLRs also recognize endogenous damage-associated molecular pattern (DAMP) molecules, such as heat shock proteins, nucleic acids from necrotic cells, high mobility group protein 1 (HMGB1), and extracellular matrix molecules, and activation of TLRs by DAMPs results in production of inflammatory mediators and tissue repair gene synthesis by which to participate in tissue damage response and repair (Marshak-Rothstein, 2006; Piccinini and Midwood, 2010). Activation of TLRs is tightly controlled to maintain homeostasis; however, in some pathogenic contexts, TLRs are overactivated to produce excessive amounts of inflammatory mediators such as TNF-α, IL-1β, and IL-6, which subsequently contribute to the pathogenesis of inflammatory disorders. The association of TLR signaling with these diseases promotes sustained efforts to develop novel drugs to block these pathways for a variety of disease indications. Attempts have been directed toward interfering with the association of TLRs with ligands or intracellular adaptor molecules (Hennessy et al., 2010; Loiarro et al., 2010), inhibiting the activation of critical kinases in TLR cascades (O’Neill, 2003; Lindstrom and Robinson, 2010) and blocking the functions of inflammatory cytokines (Kopf et al., 2010). Antagonists of TLRs and specific kinase inhibitors are either in preclinical development or at various stages of clinical trials (Hennessy et al., 2010; Lindstrom and Robinson, 2010). Targeting cytokines, such as TNF-α, IL-1β, IL-6, and IL-23 with biologics, has proven beneficial and applied in clinical practice for different diseases (Kopf et al., 2010). However, a significant portion of patients have a suboptimal response to the biologic therapies (Cohen et al., 2002; Leonardi et al., 2003; Bousquet et al., 2008), and together with issues of cost, convenience, and safety (Shear, 2006; Tak and Kalden, 2011), there is a continuing unmet medical need for an orally available small molecule able to inhibit the production of inflammatory cytokines.
A new class of boron-containing small molecules has been developed over the past several years as potential drugs. Different from carbon, boron contains an electrophilic empty p-orbital that can form transient bonds with nucleophiles in an enzyme active site, which mimics a tetrahedral transition state of peptide bond cleavage in an enzymatic reaction (Baker et al., 2011). The benzoxaboroles, in which the boron atom is incorporated into a heteroaromatic ring system, are able to inhibit a number of important enzymes, including bacterial and fungi leucyl-tRNA synthetase (Rock et al., 2007), human phosphodiesterase-4 (PDE4) (Akama et al., 2009), and hepatitis C virus NS3/4A protease (Li et al., 2010). Three benzoxaboroles, AN2690 (Tavaborole; 5-fluorobenzo[c][1,2]oxaborol-1(3H)-ol), AN2728 [4-(1-hydroxy-1,3-dihydrobenzo[c][1,2]oxaborol-5-yloxy)benzonitrile], and AN3365 [(s)-3-(aminomethyl)-7-(3-hydroxypropoxy)benzo[c][1,2]oxaborol-1(3H)-ol] are in clinical trials for treatment of onychomycosis, psoriasis/atopic dermatitis, and Gram-negative bacterial infection and have been proven safe in humans when applied topically or systemically.
In this paper, we report that a novel benzoxaborole analog, AN3485, effectively inhibits LPS (TLR4)-induced cytokine synthesis in vitro and in vivo. It also suppresses zymosan (TLR2)-, polyinosinic-polycytidylic acid (Poly I:C, TLR3)- and flagellin (TLR5)-induced cytokine release from human PBMCs. In vivo anti-inflammatory activity of AN3485 in topical and systemic disease models is also demonstrated. These results indicate that this benzoxaborole analog may provide a new approach for treatment of autoimmune and inflammatory disorders.
Materials and Methods
Animals.
CD-1 and BALB/c female mice were purchased from Charles River Laboratories (Hollister, CA), and DBA/1J male mice were purchased from Jackson Laboratories (Bar Harbor, ME). All animal studies were performed with the approval of International Animal Care and Use Committees in accordance with the Guide for the Care and Use of Laboratory Animals.
Materials.
AN3485, 6-[4-(aminomethyl)-2-chlorophenoxyl]benzo[c][1,2]oxaborol-1(3H)-ol, was synthesized at Anacor Pharmaceuticals as described in patent application WO 2010028005 A1. All lots used had purity >95% as assessed by HPLC and 1H-NMR. Escherichia coli B2 lipopolysaccharide, polyinosinic-polycytidylic acid (Poly I:C), and Histopaque-1077 were purchased from Sigma (St. Louis, MO). Zymosan and recombinant flagellin were obtained from InvivoGen (San Diego, CA). Human TNF-α, IL-1β, and IL-6 cytokine determination kits were from Cisbio (Bradford, MA). EasySep human monocyte enrichment kit and RosetteSep human T cell enrichment cocktail were bought from StemCell Technologies (Vancouver, Canada). RNeasy mini kit was from Qiagen (Valencia, CA). Dynabeads human T-activator CD3/CD28, human TNF-α, IL-1β, IL-6, and GAPDH TaqMan qPCR probes and primers were from Invitrogen (Carlsbad, CA). CellTiter 96 aqueous one solution cell proliferation assay (MTS) was obtained from Promega (Madison, WI). Rapid Equilibrium Dialysis (RED) device was bought from Thermo Scientific (Rockford, IL).
Cell Isolation, Culture, and Stimulation.
Human buffy coats used in this study were from healthy donors randomly recruited by Stanford Blood Center (Palo Alto, CA) aged 17 years or older. PBMCs were isolated from pools of buffy coats, collected from 8 donors, by using Histopaque-1077, aliquoted and frozen in liquid nitrogen. To purify human primary monocytes, PBMCs isolated from an individual buffy coat were further enriched using EasySep human monocyte enrichment kit according to the manufacturer’s instructions. Human T cells were isolated from single donor buffy coat using RosetteSep human T cell enrichment cocktail based on the instructions from the manufacturer.
Human PBMCs, monocytes, and T cells were cultured in RPMI 1640 supplemented with 10% FBS, 2 mM glutamine, and 10 mM HEPES (complete media), in a 95% air /5% CO2 incubator. AN3485 was dissolved in 100% DMSO (dimethylsulfoxide) at 40 mM before further 10-fold serial dilution in cell culture media with final concentrations ranging from 0.1 nM to 100 μM. Cells cultured with 0.25% of DMSO were used as the untreated control in the in vitro assays. PBMCs (5 × 105) were seeded in each well of a 96-well plate for 3 hours and preincubated with AN3485 for 15 minutes before stimulation with 1 μg/ml of LPS in a final volume of 200 μl. After 24 hours, the cell culture supernatants were collected for cytokine determinations using Cisbio HTRF cytokine determination kits. The same procedures were used for the experiments stimulated with zymosan (30 μg/ml), Poly I:C (10 μg/ml), and flagellin (300 ng/ml). For the experiments using human monocytes, 1 × 105 freshly isolated monocytes were seeded in each well and cultured for 1.5 hours before stimulation. To stimulate human T cells, 2 × 105 freshly purified T cells were seeded in each well for 1.5 hours and then stimulated with Dynabeads human T-activator CD3/CD28 for 48 hours.
RNA Isolation and Real-Time PCR.
Freshly isolated monocytes from individual donor’s buffy coat were cultured at 5 × 105 cells/ml for 1.5 hours and then treated with AN3485 for 15 minutes before stimulation with either LPS, zymosan, Poly I:C, or flagellin for indicated time periods. Cell culture supernatants were collected for cytokine determinations. Total RNAs were isolated from cell pellets by using an RNeasy mini kit based on manufacturer’s instructions. The mRNAs of TNF-α, IL-1β, and IL-6 were quantitated by real-time PCR using TaqMan probes and primers and normalized to the level of the endogenous control, GAPDH.
In Vitro Kinase Assay.
Kinase assays were performed in Reaction Biology (Malvern, PA) (Ma et al., 2008) with 30 μM of AN3485. For the assays with IRAK1/4, TAK1, TBK1, MAP3K8, MEK1/2, MKK4/6, p38 α/β/δ/γ, MAPKAPK2/3/5, ERK1/2, IKKα/β/ε, AKT1/2/3, JNK1/2/3, PKCα/β1, and PKA, 1 μM of [γP33]-ATP and appropriate substrates were used. For the assays with PI3Kα/β/γ/δ, 10 μM of [γP33]-ATP was added in each reaction.
Cytotoxicity (MTS Assay).
Human PBMCs and monocytes pretreated with AN3485 for 15 minutes were stimulated with LPS for 1 day. The cells were then suspended in 100 μl of fresh complete medium and incubated with 20 μl per well of MTS solution for 4 hours to allow color development. The absorbance at 490 and 690 nm was recorded. Inhibition of cell viability was calculated based on the formula: % inhibition of cell viability = 100 − (ΔODAN3485 treated / ΔODuntreated) × 100 · ΔOD = OD490-OD690.
Pharmacokinetics (PK) Study of AN3485.
Thirty-six CD-1 female mice were divided into 3 groups with 12 mice/group and received AN3485 either by intravenous injection with 5 mg/kg in a solution composed of 50% saline, 40% PEG400, and 10% DMSO (50/40/10 Saline/PEG400/DMSO, pH 5.7) or by gavage (PO) or subcutaneous injection (SC) with 10 mg/kg in 70/25/5 PEG400/PG/DMSO solution. Blood samples were collected in K2EDTA-coated tubes at eight time points during a 24-hour period with three mice at each time point. Plasma concentration of AN3485 was determined by LC/MS/MS, with Lower Limit of Quantitation (LLOQ) ≥8 ng/ml. The mean plasma concentration-time profiles were analyzed using WinNonlin Pro version 5.2 with a two-compartmental model for the intravenous data and a noncompartmental model for both PO and SC data.
LPS-Induced Cytokine Production in Mice.
The general procedures were as described by Gozzi et al. (1999) with some modifications. Female CD-1 mice (20–30 g), 10 mice per group, were orally dosed with AN3485, dissolved in vehicle 70/25/5 PEG400/PG/DMSO solution, 1 hour before intraperitoneal injection (IP) with 10 μg/kg of LPS in 0.9% saline. One hour post-LPS administration, the mice were euthanized, blood was collected in a K2EDTA-coated tube by cardiac puncture, and plasma was obtained for cytokine determination and PK analysis.
Topical PMA-Induced Irritant Contact Dermatitis.
CD-1 female mice (20-30 g), five mice/group, were used in this study based on a protocol described by Kuchera et al. (1993) with a few modifications. Five micrograms of PMA (phorbol 12-myristate 13-acetate) in 20 μl of ethanol was applied topically to the anterior and posterior surface of the right ear of each mouse. Twenty microliters of vehicle (ethanol:acetone, 1:1, vol/vol); 0.3 mg of dexamethasone (in 20 μl); 0.1, 0.3, or 1 mg of AN3485 (in 20 μl) were administrated to the same areas 30 minutes before and 15 minutes after PMA application. The thickness of right and left ears of each mouse was measured 6 hours after PMA stimulation. Ear edema was calculated by subtracting the thickness of the left ear from that of the right ear. Ears were then snap-frozen in a mixture of dry ice and ethanol and homogenized using Bullet Blender Homogenizer (Next Advance Inc., Averill Park, NY). The protein concentrations of ear homogenates were quantitated by Bradford method using Bio-Rad protein assay (Bio-Rad, Hercules, CA). Cytokines in ear homogenates were determined using BD Cytometric bead array (BD Biosciences, San Diego, CA) and normalized to protein concentration.
Topical Oxazolone-Induced Allergic Contact Dermatitis.
BALB/c female mice (20–30 g), five mice/group, were sensitized with 100 μl of 1.5% oxazolone (4-ethoxymethylene-2-phenyloxazol-5-one) in acetone to their shaved abdomen and 7 days later were challenged with 20 μl of 1% oxazolone at the anterior and posterior surface of the right ears (Evans et al., 1971). Twenty microliters of vehicle (ethanol:acetone, 1:1, vol/vol); 0.3 mg of dexamethasone (in 20 μl); 0.1, 0.3, or 1 mg of AN3485 (in 20 μl) were painted on the same areas of the right ears 30 minutes before and 15 minutes after oxazolone challenge. Ear edema was measured 24 hours after oxazolone challenge by subtracting the thickness of the left ear from that of the right ear. Ears were then snap-frozen and homogenized for cytokine determination as described above.
Toxicity Study.
Female CD-1 mice (20–30 g), five mice/group, were orally dosed once a day with 100 or 300 mg/kg of AN3485 in vehicle composed of 70% PEG400, 25% PG, and 5% DMSO for 7 days. Body weights and clinical observations were recorded daily. Blood samples were collected in K2EDTA-coated tubes 24 hours after the last dose by cardiac puncture for hematologic analysis, spleens were collected and weighed, and splenocytes were further isolated as described by Fantini et al. (2007) and stained with anti-CD3, anti-CD45R (B220), and anti-CD11b (BD Biosciences) for fluorescence-activated cell sorting analysis.
Collagen-Induced Arthritis.
DBA/1J male mice (6–7 weeks old) were immunized by subcutaneous injection with 50 μl of 2 mg/ml of bovine type II collagen emulsified with complete Freund’s adjuvant at the base of the tail and boosted on day 21 with 50 μl of 2 mg/ml of collagen emulsified with incomplete Freund’s adjuvant. Body weight, overall physical condition, and development of arthritis were assessed three times weekly, and clinical severity of arthritis in each paw was quantified based on a scale of 0–4 as described (Alonzi et al., 1998), and a cumulative disease score ranging from 0 to 16 was assigned for each mouse. When the cumulative disease score of a mouse reached 3.2, mice were randomly assigned to different groups with 10 mice per group (mice with cumulative disease scores <2 or >6 were excluded). AN3485 (twice a day), prednisolone (daily), or vehicle (twice a day) (70/25/5 PEG400/PG/DMSO solution) was orally administrated, and signs of arthritis and body weight were monitored once every other day for 20 days after dosing regimen. On day 20, the limbs were removed and immediately preserved in 10% formalin buffer, decalcified in 5% formic acid for 2–3 days, embedded in paraffin, sectioned at 8 μm, and stained with toluidine blue to highlight the joint cartilage. Histologic slides were scored for synovial inflammation, pannus, cartilage damage, and bone resorption, with all parameters on a scale of 0–5, where 0 indicated normal and 5 indicated severe as described by Bendele et al. (2000).
Statistical Analysis.
The data were presented as the mean ± standard deviation of the mean (S.D.). Differences between groups were analyzed using two-tailed unpaired Student’s t-test. Significance was considered when P < 0.05.
Results
AN3485 Inhibits LPS-Induced Cytokine Secretion In Vitro.
AN3485, a benzoxaborole with a boron atom incorporated into the five-membered ring of a six-five bicyclic system, inhibited LPS-induced TNF-α, IL-1β and IL-6 secretion from human PBMCs and isolated monocytes with IC50 values ranging from 18 to 103 nM after 24 hour exposure (Table 1). The effect of AN3485 on LPS-induced cytokines was time independent, because the IC50 values did not change when human PBMCs were stimulated for 4, 8, 12, 16, 20, and 24 hours (Supplemental Fig. 1). In comparison with AN3485, roflumilast (Daxas, Daliresp; Takeda Pharmaceuticals, Zurich, Switzerland), a PDE4 inhibitor, and SB203580 [4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl)-1H-imidazole] (Sigma-Aldrich, St. Louis, MO), a p38 MAPK inhibitor, inhibited LPS-induced TNF-α but had no effect on IL-6 release (Supplemental Table 1). The distinct pattern of cytokine blockade suggests that AN3485 may target a molecule different from PDE4 and p38 MAPK.
Inhibition of LPS-induced TNF-α, IL-1β and IL-6 secretion from PBMCs and isolated monocytes
Human PBMCs or monocytes were stimulated with 1 μg/ml of LPS for 24 hours in the presence of serial 10-fold diluted AN3485 with final concentration ranging from 0.1 nM to 100 μM. Cytokines in the culture supernatants were analyzed by using Cisbio HTFR cytokine determination kits. The data are the average of at least four independent experiments, mean ± S.D.
To ensure that the inhibition of cytokine secretion by AN3485 was not simply due to a cytotoxic effect on cells, the viability of LPS-stimulated PBMCs and monocytes was investigated by using MTS assay. More than 70% of cells survived after 24 hour exposure to 100 μM of AN3485, and this compound had no effect on cell viability when PBMCs were stimulated for 4 hours (Fig. 1), at which time AN3485 showed comparable IC50 values against LPS-induced TNF-α, IL-1β, and IL-6 (Supplemental Fig. 1). These results indicated that the inhibitory capability of AN3485 was unlikely due to a cytotoxic effect, instead, it inhibited TLR4-mediated cytokine synthesis or secretion.
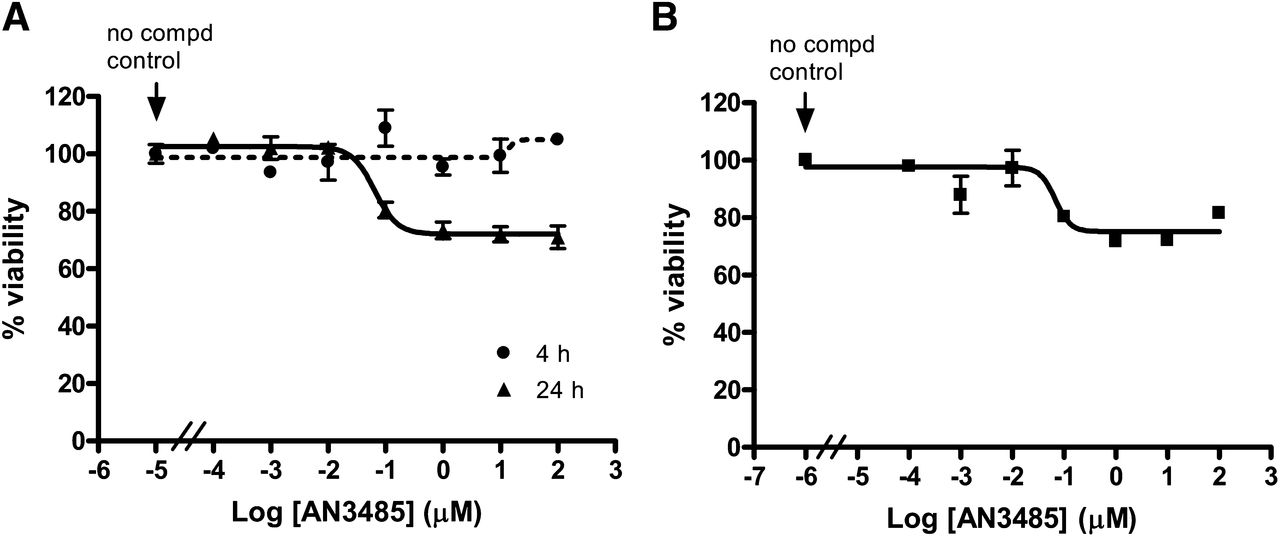
Effect of AN3485 on human PBMCs and monocytes viability. Human PBMCs (A) and monocytes (B) were stimulated with LPS in the presence of AN3485 for indicated time periods. The cell viability was determined by further incubation of the cells with MTS solution as described in Materials and Methods.
AN3485 Reduces LPS-Induced Cytokine Transcription.
LPS-induced cytokines are regulated at both transcriptional (Economou et al., 1989; Yao et al., 1997; Xu et al., 2001) and post-transcriptional (Han et al., 1990; Lai et al., 1999; MacKenzie et al., 2002) levels. To elucidate whether AN3485 modulates the mRNA levels of these cytokines, the mRNAs of TNF-α, IL-1β, and IL-6 in LPS-stimulated monocytes were quantitated. AN3485 concentration-dependently reduced the amounts of TNF-α, IL-1β and IL-6 mRNA in response to LPS stimulation, and at 3 hours after LPS stimulation, 0.1 μM of the compound decreased these three cytokine mRNAs by 68%, 63%, and 88%, respectively (Fig. 2A). Correspondingly, AN3485 significantly inhibited the secretion of TNF-α and IL-6 proteins in cell culture supernatant (Fig. 2B). Although IL-1β mRNA was induced as early as 1 hour upon LPS stimulation, and was 5- and 21-fold more abundant than IL-6 and TNF-α mRNA at the 3 hour time point, IL-1β was undetectable in the culture supernatant even after 3 hour stimulation. The absence of secreted IL-1β was expected because the secretion of IL-1β requires a rate-limiting second step to cleave pro-IL-1β (Netea et al., 2009; Carta et al., 2011).

AN3485 inhibits LPS-induced cytokines at mRNA and protein levels. Freshly isolated human monocytes from individual donor’s buffy coat were stimulated with LPS in the presence of AN3485 for indicated time periods. The mRNA of a cytokine was quantitated by qPCR and normalized to the level of GAPDH (A), and cytokines in the culture supernatant were determined (B). Each value represents the mean ± S.D. of triplicates. Statistical significance compared with untreated control (*P < 0.05; **P < 0.005; ***P < 0.001).
AN3485 Reduces TLR2-, TLR3-, and TLR5-Mediated Cytokine Production.
To understand whether AN3485 also functions in other TLR-mediated pathways, the effect of AN3485 on zymosan (TLR2)-, Poly I:C (TLR3)-, and flagellin (TLR5)-induced cytokine release was evaluated. In comparison with the effect on LPS stimulation, AN3485 had similar potency against Poly I:C- and zymosan-induced TNF-α, IL-1β, and IL-6 synthesis in human PBMCs with IC50 values ranging from 28 to 580 nM (Table 2), and the IC50 values did not show differences with 4 and 24 hour activation (data not shown). AN3485 also prevented flagellin-induced TNF-α and IL-6 release from human PBMCs with IC50 values of 190 and 80 nM when cells were stimulated for 4 hours.
AN3485 inhibits zymosan- and Poly I:C-induced cytokine production
Human PBMCs were stimulated with zymosan and Poly I:C for 24 hours in the presence of 10-fold serial dilution of AN3485 with final concentration ranging from 0.1 nM to 100 μM. Cytokines in the culture supernatants were quantitated by using Cisbio HTRF cytokine determination kits. The data are average of three or more independent experiments, mean ± S.D.
AN3485 Reduces TLR2-, TLR3-, and TLR5-Mediated Cytokine Transcription.
Effects of AN3485 on the mRNA levels of TNF-α, IL-1β, and IL-6 were also observed when human monocytes were stimulated with zymosan, Poly I:C, or flagellin. Induction of TNF-α and IL-6 mRNA in response to zymosan was relatively slower than that in response to LPS, Poly I:C, and flagellin, such that IL-6 mRNA was barely induced by zymosan even by 3 hours. Transcription of TNF-α in response to flagellin was rapid, but its mRNA level fell off quickly (Fig. 3A). After 3 hour stimulation, 1 μM of AN3485 decreased zymosan- and Poly I:C-induced TNF-α mRNA by 85% and 76%; zymosan-, Poly I:C-, and flagellin-induced IL-1β mRNA by 80%, 52%, and 84%; and Poly I:C- and flagellin-induced IL-6 mRNA by 97% and 97% (Fig. 3A). As a consequence, the effect on transcription was translated to the inhibition of TNF-α and IL-6 proteins released to the culture supernatant (Fig. 3B). Together, these results indicated that AN3485 was able to inhibit TLR2-, TLR3-, and TLR5-mediated cytokine release by effects on the transcription of these cytokines.

AN3485 inhibits zymosan-, Poly I:C-, and flagellin-induced cytokines at mRNA and protein levels. Freshly isolated human monocytes were stimulated with zymosan, Poly I:C, or flagellin in the presence of AN3485 for indicated time periods. The mRNA of a cytokine was quantitated by qPCR and normalized to the level of GAPDH (A), and cytokines in the culture supernatant were determined (B). Each value represents the mean ± S.D. of triplicates. Statistical significance compared with untreated control (*P < 0.05; **P < 0.005; ***P < 0.001).
AN3485 is Inactive Against Kinases in TLR Signaling Pathways.
The possibility that AN3485 might act as an inhibitor of a kinase in TLR pathways was addressed by in vitro kinase assay. The activities of 34 kinases involved in TLR signaling (Gaestel et al., 2009) were tested. AN3485 (30 μM) had no effect on the activities of IRAK1/4, TAK1, TBK1, MAP3K8, MEK1/2, MKK4/6, p38α/β/γ/δ, MAPKAPK2/3/5, ERK1/2, IKKα/β/ε, AKT1/2/3, PI3Kα/β/γ/δ, JNK1/2/3, PKCα/β1, and PKA. Therefore, AN3485 apparently does not act as a kinase inhibitor by blocking TLR-mediated TNF-α, IL-1β, and IL-6 production.
AN3485 Shows Activity Against T Cell Receptor-Mediated Cytokine Secretion.
The activity of AN3485 against TCR-mediated (T-cell receptor) cytokine production was investigated. AN3485 prevented anti-CD3/CD28-induced IL-2, IFN-γ, TNF-α, IL-4, and IL-13 secretion from purified human T cells with IC50 values of 1.07, 2.37, 2.45, 1.94, and 12.3 μM, respectively, indicating that this activity is less potent than that against TLR-mediated cytokine production in monocytes. With concentrations up to 100 μM, AN3485 did not show effect on the viability of resting and activated T cells after 48 hour exposure. Additionally, with 3 day incubation, the IC50 of AN3485 on the viability of Jurkat (a human T-cell line) was >30 μM.
Pharmacokinetics of AN3485.
The pharmacokinetic profile of AN3485 was assessed in CD-1 mice. The plasma clearance of the compound was 2210 ml/h/kg following i.v. injection at 5 mg/kg. T1/2 and AUC0–inf with oral administration of 10 mg/kg were 2.8 h and 3.05 h*μg/ml, respectively, with optimal oral bioavailability of 59%. The in vitro plasma protein binding for AN3485 was 76.2% in mice as determined by using a rapid equilibrium dialysis (RED) device and quantitated by liquid chromatography-tandem mass spectrometry. These data indicated that adequate trough concentration of AN3485 would be achieved by dosing twice a day for a long-term efficacy study.
Oral Administration of AN3485 Suppresses Cytokine Elaboration in Response to LPS In Vivo.
Preliminary studies showed that intraperitoneal injection of 10 μg/kg of LPS induced significant amounts of TNF-α and IL-6 in mouse plasma with peak at 1 hour post-administration, which was consistent with that reported previously (Ruggiero et al., 1993; Gozzi et al., 1999). Pretreatment with AN3485 dose-dependently decreased LPS-induced plasma levels of TNF-α and IL-6 in mice, with ED50 and ED90 values of approximately 8 and 30 mg/kg (Fig. 4). The plasma levels of AN3485 with 8 and 30 mg/kg dosages at 2 hours after administration were approximately 0.7 and 2.3 μM, which corresponded to 167 [0.7 μM × (100%–76.2%) = 0.167 μM] and 547 nM [2.3 μM × (100%–76.2%) = 0.547 μM] of free compound in the plasma as calculated based on the protein binding of 76.2% for AN3485. The concentrations of free AN3485 in plasma correlated well with the in vitro IC50 values and IC90 values of LPS-induced TNF-α (IC50 = 103 nM; IC90 = 1000 nM) and IL-6 (IC50 = 72 nM; IC90 = 500 nM), suggesting that likely the same mechanism was effected in vitro and in vivo. The ability of oral AN3485 against LPS-induced cytokine production in vivo supports its potential application in inflammatory diseases associated with these cytokines.

Oral administration of AN3485 inhibits LPS-induced TNF-α and IL-6 in mice. AN3485 in plasma after 2 hours of administration is indicated in blue. Each value represents the mean ± S.D. of 10 mice each group.
In Vivo Toxicity of AN3485.
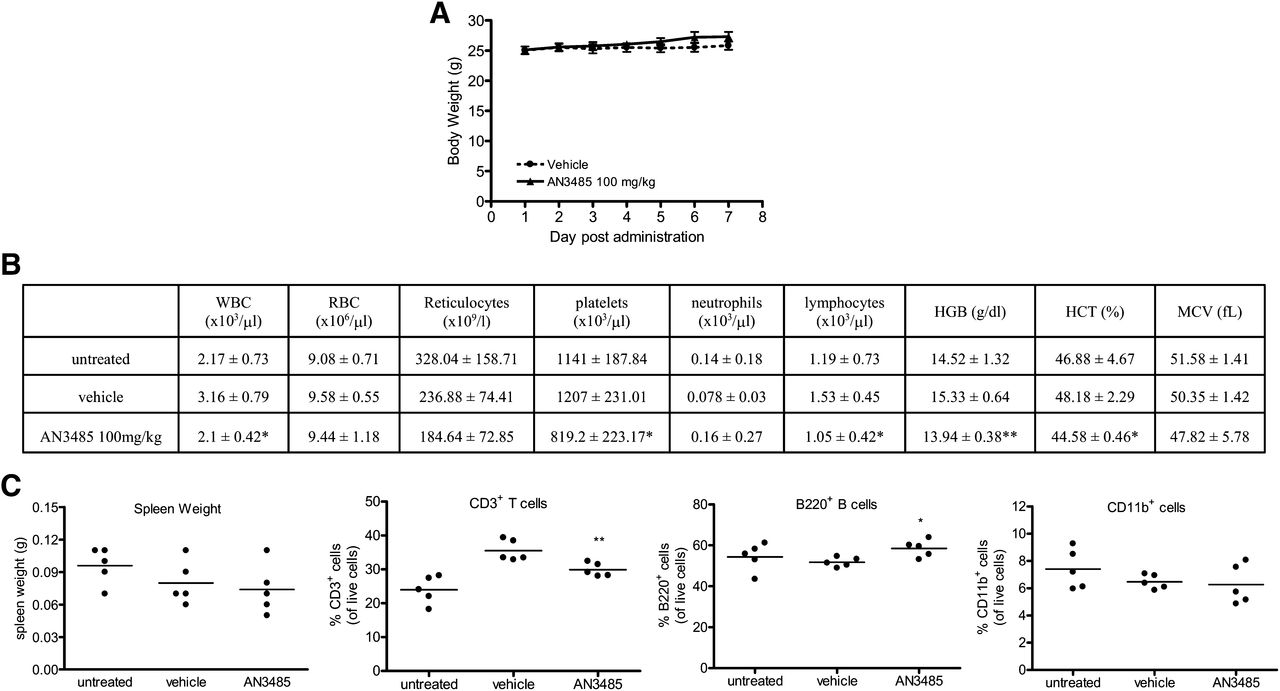
The toxicity of AN3485 was evaluated in preparation for long-term efficacy studies. CD-1 mice were orally dosed once a day for 7 consecutive days with 100 or 300 mg/kg of AN3485 or vehicle. Clinical signs, including ataxia, rough coat, ptosis, and lethargy and 13–18% decrease in body weight gains between days 1 and 5 were observed in the 300 mg/kg group. No clinical signs and decrease in body weight gains were noted in 100 mg/kg group (Fig. 5A). In comparison with vehicle-treated group, mice with 100 mg/kg of AN3485 had decreases in total white blood cells, lymphocytes, and platelets and lower hemoglobin levels and hematocrits in blood (Fig. 5B). However, untreated mice also showed lower values for each of these compared with vehicle-treated controls, and there were no statistical differences of these parameters between untreated mice and the 100 mg/kg AN3485-treated group. Hence, the effects of AN3485 at 100 mg/kg were not clinically significant. fluorescence-activated cell sorting analysis demonstrated that the major immune cells in the spleen, including T cells (CD3+), B cells (B220+), and monocytes/macrophages (CD11b+), were not significantly altered in mice treated with 100 mg/kg of AN3485 compared with the untreated group, although there was a decrease of T cells and an increase of B cells in AN3485-treated mice compared with the vehicle-treated group (Fig. 5C). Spleen weights were not statistically different among these three groups of mice (Fig. 5C). In conclusion, oral administration of AN3485 for 7 consecutive days did not result in any signs of overt toxicity at dose of 100 mg/kg/day, which is considered to be the no observable adverse effect level.

Seven-day toxicity study of oral AN3485. Mice were either untreated or orally administrated once a day with 100 mg/kg of AN3485 or vehicle for 7 days. Body weights were recorded (A); hematology of blood collected on day 8 was assessed (B); and spleen weights, percentage of CD3+, B220+, and CD11b+ cells in spleens on day 8 were analyzed (C). The data represent the mean ± S.D. of 5 mice each group. Statistical significance compared with vehicle group (*P < 0.05; **P < 0.005).
Topical Application of AN3485 Reduces PMA-Induced Acute Contact Dermatitis in Mice.
PMA-treated mice show local increases in expression of IL-1β and IL-6 (DeVry et al., 2005) as well as in vascular permeability (Lee et al., 1994) and have been used as an experimental model of irritant contact dermatitis. Topical treatment with 0.3 mg of dexamethasone reduced mice ear swelling by 64% 6 hours after PMA stimulation. Topical AN3485 showed comparable effects: 0.1, 0.3, and 1 mg of AN3485 reduced ear edema by 21, 76, and 90%, respectively (Fig. 6A). Significant amounts of IL-1β and IL-6 were detected in mouse ear homogenate 6 hours after PMA challenge (Fig. 6B). Topical 0.1, 0.3, and 1 mg of AN3485 inhibited PMA-induced IL-6 by 65, 94, and 96% and IL-1β by 70, 84, and 91%, whereas 0.3 mg of dexamethasone blocked IL-6 and IL-1β by 70% and 54% (Fig. 6B).

Topical application of AN3485 reduces PMA-induced contact dermatitis. Mice ear edema (A) and cytokines in ears (B) were determined 6 hours after PMA stimulation. Each value represents the mean ± S.D. of 5 mice each group. Dex, dexamethasone. Statistical significance compared with vehicle group (*P < 0.05; **P < 0.005; ***P < 0.001).
Topical AN3485 Reduces Oxazolone-Induced Delayed-Type Hypersensitivity in Mice.
The effect of AN3485 on oxazolone-induced delayed-type hypersensitivity, a model of allergic contact dermatitis, was investigated. Topical application of 0.1, 0.3, and 1 mg of AN3485 reduced ear swelling by 58%, 92%, and 100% 24 hours after oxazolone challenge, whereas 0.3 mg of dexamethasone suppressed ear edema by 98% (Fig. 7A). Challenge with oxazolone also induced significant amounts of IL-1β, IL-6, TNF-α, and IL-4 in mice ears (Fig. 7B), but no IL-2, IL-13, and IFN-γ were detectable. Topical treatment with 0.1, 0.3, and 1 mg of AN3485 inhibited oxazolone-induced IL-1β by 16%, 43%, and 75%; IL-6 by 16%, 62%, and 83%; TNF-α by 2%, 26%, and 49%; and IL-4 by 14%, 49%, and 72% (Fig. 7B). Dexamethasone, 0.3 mg/ear, suppressed oxazolone-induced IL-1β, IL-6, TNF-α, and IL-4 by 96%, 96%, 64%, and 84% (Fig. 7B). Taken together, the effect of AN3485 on PMA- and oxazolone-induced ear edema and cytokine production in mice ears indicated that this compound could penetrate through skin to block skin inflammation and supports the topical application of AN3485 in cutaneous inflammatory disorders such as psoriasis and atopic dermatitis.

Topical AN3485 suppresses oxazolone-induced delayed-type hypersensitivity. Mice ear swelling (A) and cytokines in ears (B) were determined 24 hours after oxazolone challenge. Each value represents the mean ± S.D. of 5 mice each group. Statistical significance compared with vehicle group (*P < 0.05; **P < 0.005; ***P < 0.001).
AN3485 Inhibits Collagen-Induced Arthritis in Mice.
To examine the ability of AN3485 to suppress profound chronic inflammation, we examined its effects in collagen-induced arthritis (CIA), a murine model of rheumatoid arthritis (RA). In the CIA model and in RA patients, the synovial membrane is infiltrated by immune cells including macrophages and T cells, which produce inflammatory mediators leading to pain, inflammatory cell infiltration, swelling, and cartilage and bone erosion (Stuart et al., 1988). Mice given 35 mg/kg of AN3485 twice a day showed a dramatically curtailed rate of disease progression: the cumulative disease score of the vehicle-treated group soared 9.3 during the 20-day period, whereas the AN3485-treated group only increased 3.4, a 63% inhibition of the disease development (Fig. 8A). The amelioration of arthritis by AN3485 was also confirmed by histologic evaluation of the joints collected on day 20. Mice treated with AN3485 showed less severity of inflammation and bone erosion in joints of the limbs (Fig. 8B), with 67%, 85%, 81%, and 85% inhibition of synovial inflammation, pannus formation, cartilage damage, and bone resorption (Fig. 8C). The dosage of AN3485 was well tolerated: mice with AN3485 treatment showed no adverse events and had similar body weight gain to the vehicle-treated counterparts.

Oral administration of AN3485 attenuates collagen-induced arthritis in mice. Disease development (cumulative disease score) was assessed three times a week (A), histologic evaluation of joints was performed on day 20 after drug regimen with toluidine blue staining (B), and histologic scores of synovial inflammation, pannus, cartilage damage, and bone resorption were quantitated and found to be statistically different from vehicle group (P < 0.001 for all of these parameters) (C). W indicates wrist, and the arrows indicate some of the affected joints with significant cartilage and bone erosion and tissue swelling (magnification 16×).
Discussion
Excessive cytokine production contributes to the pathogenesis of a number of inflammatory and autoimmune diseases. Biologics targeting TNF-α, IL-6, IL-1, and IL-23 have shown beneficial effects in rheumatoid arthritis, psoriasis, and Crohn’s disease (Cohen, et al., 2002; Leonardi, et al., 2003; Kopf et al., 2010). However, due to incomplete responses in some patients and side effects of current available drugs, there is an unmet medical need for new immunosuppressive drugs. An orally or topically bioavailable small molecule capable of suppressing multiple critical cytokines, such as AN3485 described herein, may be a good approach for autoimmune and inflammatory diseases.
We demonstrated in this paper that AN3485 inhibited TLR-mediated TNF-α, IL-1β, and IL-6 secretion with submicromolar IC50 values, and the effect was transcriptionally mediated, because 100 nM of AN3485 could inhibit LPS-induced upregulation of the mRNA levels of these three cytokines by 68%, 63%, and 88%, respectively. AN3485 at pharmacologically effective concentrations had mild effects on the viability of monocytes, suggesting cytotoxicity is unlikely the mechanism contributing to cytokine blockade.
Eleven TLRs, which recognize a wide range of exogenous PAMPs and endogenous DAMPs, have been identified in human. Upon engagement with their cognate ligands, TLRs either homo- or heterodimerize to promote intracellular TIR-TIR interaction and recruit adaptor molecules to the complex (Jin and Lee, 2008). TLR3 recruits TRIF to induce TRIF-dependent response, whereas the other TLRs can interact with MyD88 to trigger MyD88-dependent signaling. The formation of TLR complex triggers the autophosphorylation of IRAK4, which further leads to the signal cascade mediated by a number of kinases and eventually results in the transcription of inflammatory mediators (Han and Ulevitch, 2005; Gaestel, et al., 2009; Kawai and Akira, 2010). Crystal structures of the extracellular domains of TLR2/TLR1 (Jin et al., 2007), TLR4/MD-2 (Kim et al., 2007; Park et al., 2009), TLR3 (Liu et al., 2008), and TLR5 (Yoon et al., 2012) with their ligands reveal that although these receptors differ in ligand recognition, they all form a common tail-to-tail “m”-shaped dimer, suggesting the possible conservation of the “m”-like structure among all TLR complexes. In this study, we demonstrated that AN3485 not only inhibited TLR4-mediated TNF-α, IL-1β, and IL-6 production, but also blocked TLR2-, TLR3-, and TLR5-mediated cytokine synthesis at both mRNA and protein levels. Given the differences in the ligands and adaptor molecules associated with these TLR complexes, it is unlikely that AN3485 acts by depleting a TLR ligand or interacting directly with an adaptor molecule in the complex. Instead, AN3485 may bind to the conserved “m”-like structure of the complexes or target a molecule shared by the TLR pathways that leads to the transcription of the cytokines.
A number of kinases are activated upon the binding of a TLR agonist and act as essential signaling components among the TLR pathways (Gaestel et al., 2009). In vitro kinase assays indicated that AN3485 was inactive against 34 kinases involved in TLR pathways. Further work to decipher the effect of the compound on TLR-mediated signaling pathways in human monocytes is in progress, and the results will help to understand the mechanism of action of the molecule.
In addition to blockade of TLR-mediated cytokine production, AN3485 also had some effect on TCR-mediated IL-2, IFN-γ, TNF-α, IL-4, and IL-13 synthesis. The potency of AN3485 against anti-CD3/CD28-induced TNF-α secretion from human T cells was 32-fold less than that against LPS-induced TNF-α release from monocytes, suggesting stronger inhibition of TLR signaling by AN3485. Although the activity against TCR-mediated cytokine secretion was relatively impotent, it may still contribute to the in vivo efficacy of AN3485 in the models of collagen-induced arthritis and oxazolone-induced delayed-type hypersensitivity, in which pathogenesis involves T cell activation.
Toxicity study indicated that mice orally dosed with 100 mg/kg/day of AN3485 for 7 days did not show clinical signs and had no significant changes in blood components and immune cell populations in spleens. Oral administration of 35 mg/kg of AN3485 twice a day for 20 days was tolerable and had no effect on mice body weight gain. These results coincide with the in vitro effect on cell viability, suggesting minimal effect of AN3485 on cell survival.
AN3485 reduced the profound inflammation of collagen-induced arthritis, a model that resembles human rheumatoid arthritis. AN3485 suppressed all elements of the joint damage including pannus formation, bone erosion, invasion of inflammatory cells, and cartilage damage by 67–85% and attenuated the severity of established arthritis by 63% on day 20. The efficacy is comparable to that of anti-TNF-α biologics infliximab (Remicade; Janssen Biotech, Horsham, PA) and etanercept (Enbrel; Amgen, Thousand Oaks, CA) (∼70%) (Joosten et al., 1999; Zalevsky et al., 2007) and a JAK1/2 inhibitor INCB028050 (Baricitinib; Incyte, Wilmington, DE) (∼60%) (Fridman et al., 2010), which demonstrated beneficial effects in patients with active rheumatoid arthritis. The potency of AN3485 in CIA model makes it promising for the application in rheumatoid arthritis.
In conclusion, AN3485, a novel benzoxaborole, inhibits TLR-mediated pro-inflammatory cytokine synthesis in a transcription-dependent manner. Topical application of AN3485 reduced PMA- and oxazolone-induced skin inflammation. Oral AN3485 alleviated the chronic inflammation associated with collagen-induced arthritis in mice. The potent anti-inflammatory activity of AN3485 in vitro and in disease models indicates its potential therapeutic application in acute and chronic inflammatory diseases, such as rheumatoid arthritis, psoriasis, and atopic dermatitis.
Authorship Contributions
Participated in research design: Dong, Jarnagin, Freund.
Conducted experiments: Dong, Sexton, Gertrudes, Martin, Virtucio, Chen, Fan, Feng.
Performed data analysis: Dong, Chen, Wu, Bu, Liu.
Contributed new reagents or analytic tools: Akama.
Wrote or contributed to the writing of the manuscript: Dong, Jarnagin, Freund.
Footnotes
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- AN2690
- 5-fluorobenzo[c][1,2]oxaborol-1(3H)-ol
- AN2728
- 4-(1-hydroxy-1,3-dihydrobenzo[c][1,2]oxaborol-5-yloxy)benzonitrile
- AN3365
- (s)-3-(aminomethyl)-7-(3-hydroxypropoxy)benzo[c][1,2]oxaborol-1(3H)-ol
- AN3485
- 6-[4-(aminomethyl)-2-chlorophenoxyl]benzo[c][1,2]oxaborol-1(3H)-ol
- DAMP
- damage-associated molecular pattern
- DMSO
- dimethylsulfoxide
- ED90
- 90% inhibitory dose
- FACS
- fluorescence-activated cell sorting
- IC50
- 50% inhibitory concentration
- IL
- interleukin
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- LPS
- lipopolysaccharide
- MTS
- (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxylmethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium)
- PAMP
- pathogen-associated molecular pattern
- PDE4
- human phosphodiesterase-4
- PEG400
- polyethylene glycol-400
- PG
- propylene glycol
- PMA
- phorbol 12-myristate 13-acetate
- PK
- pharmacokinetics
- SB203580
- 4-(4-fluorophenyl)-2-(4-methylsulfinylphenyl)-5-(4-pyridyl)-1H-imidazole
- TCR
- T-cell receptor
- TIR
- Toll-interleukin 1 receptor
- TLR
- Toll-like receptor
- TNF-α
- tumor necrosis factor
- Received September 20, 2012.
- Accepted November 27, 2012.
- Copyright © 2013 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}