


Abstract
Synthetic cannabinoids (CBs) [naphthalen-1-yl-(1-pentylindol-3-yl) methanone (JWH-018) and naphthalen-1-yl-(1-butylindol-3-yl) methanone (JWH-073)] are marketed, sold, and used as alternatives to cannabis. Synthetic CBs appear to have effects similar to those of Δ9-tetrahydrocannabinol (Δ9-THC), the drug primarily responsible for the behavioral effects of cannabis. However, synthetic CB products produce atypical effects (e.g., hypertension, seizures, and panic attacks). One potential explanation for atypical effects is CB1 receptor agonist efficacy, which is reportedly higher for JWH-018 and JWH-073 compared with Δ9-THC. The goal of this study was to test a prediction from receptor theory that tolerance/cross-tolerance (i.e., resulting from daily Δ9-THC treatment) is greater for a low-efficacy agonist compared with a high-efficacy agonist. Rhesus monkeys discriminated 0.1 mg/kg Δ9-THC i.v. from vehicle, and sensitivity to CB1 agonists was determined before and after 3 and 14 days of Δ9-THC treatment (1 mg/kg per day s.c.). (1R,3R,4R)-3-[2-Hydroxy-4-(1,1-dimethylheptyl) phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol (CP-55,940), a prototype high-efficacy CB1 receptor agonist, JWH-018, and JWH-073 substituted for the discriminative stimulus effects of Δ9-THC. Three days of Δ9-THC treatment produced less tolerance/cross-tolerance than 14 days of Δ9-THC treatment. Three days of Δ9-THC did not result in cross-tolerance to CP-55,940, JWH-073, and JWH-018; in contrast, as reported previously, 3 days of Δ9-THC treatment decreased sensitivity to Δ9-THC 3-fold. Fourteen days of Δ9-THC decreased sensitivity to Δ9-THC, CP-55,940, JWH-018, and JWH-073 9.2-fold, 3.6-fold, 4.3-fold, and 5.6-fold, respectively. The greater loss of sensitivity to Δ9-THC relative to CP-55,940 and JWH-018 suggests that differences in CB1 receptor agonist efficacy are important in vivo and might underlie differences in the dependence liability and adverse effects of synthetic CBs versus cannabis.
Introduction
Recreational use of herbal mixtures under various brand names, such as “Spice,” has increased because of several factors: the herbal mixtures contain synthetic cannabinoids (CBs), the products are readily available, and there is no common method available to detect synthetic CBs in urine (Auwärter et al., 2009; Vardakou et al., 2010). Many of these products are sold via the internet or in so-called “headshops” and smoked despite warnings that these products are not intended for human consumption. The products are mixtures (blends) reported to contain plant/herbal ingredients such as Pedicularis densiflora, Nymphaea caerulea, Leonotis leonurus, Leonurus sibiricus, Canavalia maritima, or Zornia latifolia without mentioning any synthetic additives. Chemical analysis revealed that the psychoactive effects of the herbal mixture were due to added synthetic CBs and not Δ9-tetrahydrocannabinol (Δ9-THC). Two synthetic CBs very often present in marketed products are JWH-018 [naphthalen-1-yl-(1-pentylindol-3-yl) methanone] and JWH-073 [naphthalen-1-yl-(1-butylindol-3-yl) methanone] (Lindigkeit et al., 2009; Dresen et al., 2010; Uchiyama et al., 2010).
JWH-018 and JWH-073 are members of the JWH aminoalkylindole family and, similar to Δ9-THC, bind to and activate CB receptors in the central nervous system. Compared with cannabis use, synthetic CB use appears to result in a higher prevalence of adverse effects such as hypertension, seizures, and panic attacks that often require immediate medical care (Müller et al., 2010; Vearrier and Osterhoudt, 2010; Schneir et al., 2011; Simmons et al., 2011). Given that relatively large doses of Δ9-THC can produce anxiety (Carbuto et al., 2012), anxiety resulting from use of synthetic CBs is expected. However, the extent to which Δ9-THC and synthetic CBs differ qualitatively or quantitatively in producing anxiogenic effects has not been established. In a published study using the same Δ9-THC discrimination assay as that used here, JWH-018 and JWH-073 substituted for the discriminative stimulus effects of Δ9-THC (Ginsburg et al., 2012). In addition to being qualitatively similar, quantitative (Schild) analysis suggested that the discriminative stimulus effects of all three agonists were mediated by the same CB1 receptors (Ginsburg et al., 2012). Other in vivo effects of JWH-018 and JWH-073, which include hypoactivity, immobility, antinociception, and hypothermia in mice (Wiley et al., 1998), also are consistent with a CB1 receptor agonist profile.
CB1 receptors are G-protein-coupled and are densely localized in brain areas such as the hippocampus, basal ganglia, cortex, amygdala, and cerebellum (Herkenham et al., 1991; Gifford et al., 1999) and in lesser amounts in adipose tissue, skeletal muscle, and liver (Lindborg et al., 2010; Wu et al., 2011). CB1 receptor agonists vary in the maximum to which they stimulate G-protein-coupled signaling (agonist efficacy), as indexed by G-protein activation assessed with guanosine 5′-O-(3-[35S]thio)triphosphate (GTPγS) binding in rodent brain tissue. JWH-018 and JWH-073 were reported to have high efficacy, i.e., similar to (1R,3R,4R)-3-[2-hydroxy-4-(1,1-dimethylheptyl) phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol (CP-55,940) and approximately 5-fold greater than Δ9-THC (Brents et al., 2011, 2012). However, despite low efficacy, Δ9-THC often produces the same maximal effect in vivo as that obtained with high-efficacy CB agonists, perhaps because of a large number of CB1 receptors (i.e., spare receptors) in the central nervous system (Gifford et al., 1999). CP-55,940, JWH-018, and JWH-073 fully substituted for the discriminative stimulus effects of Δ9-THC in rhesus monkeys (McMahon, 2006; Ginsburg et al., 2012). Given that animals trained to Δ9-THC are presumably discriminating low-efficacy CB1 receptor agonism, full substitution with higher efficacy agonists is to be expected.
One strategy for assessing relative efficacy among agonists is to decrease receptor function through chronic agonist treatment. Chronic Δ9-THC results in CB1 receptor down-regulation and desensitization and tolerance to the effects of Δ9-THC on locomotor activity, rectal temperature, and nociception in mice; however, there is less cross-tolerance to CP-55,940, 4,5-dihydro-2-methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo[3,2,1-i,j]quinolin-6-one (WIN-55,212-2), and anandamide (Fan et al., 1994; Falenski et al., 210). The greater magnitude of change in sensitivity to Δ9-THC compared with CP-55,940, WIN-55,212-2, and anandamide under identical conditions of CB treatment is consistent with receptor theory (Kenakin, 1981) inasmuch as anandamide, WIN-55,212-2, and CP-55,940 have higher efficacy than Δ9-THC (Breivogel and Childers, 2000; Childers, 2006). Relative to effects on locomotor activity, rectal temperature, and nociception, the discriminative stimulus effects of Δ9-THC are pharmacologically selective (Balster and Prescott, 1992). With the discrimination assay used here, there is strong evidence that the same, presumably CB1, receptors mediate the effects of various agonists. Moreover, tolerance develops to the discriminative stimulus effects of Δ9-THC, as described previously (Wiley et al., 1993; McMahon, 2011). However, relationships between tolerance/cross-tolerance to discriminative stimulus effects and efficacy among agonists such as JWH-018 and JWH-073 have not been established.
In the current study, rhesus monkeys received repeated Δ9-THC treatment. In particular, rhesus monkeys discriminated Δ9-THC (0.1 mg/kg i.v.) from vehicle and sensitivity to the discriminative stimulus effects of Δ9-THC and the Δ9-THC-like discriminative stimulus effect of CP-55,940 (a prototypical high-efficacy agonist), JWH-018, and JWH-073 was assessed before and after 3 or 14 days of Δ9-THC treatment (1 mg/kg/day s.c.). Overall, this study tested the hypothesis that the magnitude of tolerance to Δ9-THC, a lower efficacy agonist, is greater than cross-tolerance to higher efficacy agonists.
Materials and Methods
Subjects.
Two female and three male adult rhesus monkeys (Macaca mulatta) discriminated Δ9-THC from vehicle. Monkeys were housed individually on a 14-h light/10-h dark schedule and maintained at 95% free-feeding weight (range, 5.8–10.4 kg) with a diet consisting of primate chow (High Protein Monkey Diet; Harlan Teklad, Madison, WI), fresh fruit, and peanuts; water was provided in the home cage. Monkeys received CBs and non-CBs in previous studies (McMahon, 2011; Ginsburg et al., 2012). Monkeys were maintained in accordance with the Institutional Animal Care and Use Committee, The University of Texas Health Science Center at San Antonio, and the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, 2011).
Surgery.
Monkeys were anesthetized with ketamine (10 mg/kg i.m.) followed by isoflurane (1.5–3.0% inhaled via facemask). A catheter (heparin-coated polyurethane; o.d. = 1.68 mm; i.d. = 1.02 mm; Instech Laboratories, Plymouth Meeting, PA) was inserted into a subclavian or femoral vein and secured to the vessel with suture silk (coated Vicryl; Ethicon Inc., Somerville, NJ). The catheter extended from the vessel to the midscapular region of the back and was attached to a vascular access port located subcutaneously (Mida-cbas-c50; Instech Laboratories).
Apparatus.
Monkeys were seated in chairs (model R001; Primate Products, Miami, FL) that provided restraint. Chairs were placed in ventilated, sound-attenuating chambers equipped with two levers; a light was positioned above each lever. Feet were placed in shoes containing brass electrodes to which a brief electric stimulus (3 mA, 250 ms) could be delivered from an a.c. generator. The chambers were connected to a computer with an interface (MED Associates, St. Albans, VT); experimental events were controlled and recorded with Med-PC software (MED Associates).
Drug Discrimination Training.
Monkeys discriminated Δ9-THC (0.1 mg/kg i.v.) from vehicle [1 part absolute ethanol, 1 part Emulphor-620 (Rhodia Inc., Cranbury, NJ), and 18 parts saline] while responding under a fixed ratio (FR) 5 schedule of stimulus-shock termination. The experimental sessions were divided into consecutive 10-min multiple cycles; each cycle began with a 5-min timeout. Responses during the timeout had no programmed consequence. The timeout was followed by a 5-min schedule of stimulus-shock termination, the beginning of which was signaled by illumination of red lights. Five consecutive responses on the correct lever extinguished the red lights, prevented delivery of an electric stimulus, and initiated a 30-s timeout. Otherwise, an electric stimulus was delivered every 40 s. Responding on the incorrect lever reset the response requirement on the correct lever. Determination of correct levers varied among monkeys (e.g., left lever associated with 0.1 mg/kg Δ9-THC and right lever associated with vehicle) and remained the same for that monkey for the duration of the study.
Training sessions consisted of a minimum of three and a maximum of six cycles. Δ9-THC training consisted of administration of Δ9-THC (0.1 mg/kg) within the first minute of three cycles; vehicle or sham (dull pressure applied to the skin overlying the vascular access port) was administered within the first minute of subsequent cycles. Vehicle training involved administration of vehicle within the first minute of a cycle followed by vehicle or sham in subsequent cycles. Zero to three vehicle-training cycles immediately preceded three Δ9-THC training cycles. Completion of the FR on the correct lever was required for reinforcement during each training cycle. Monkeys had previously satisfied the criteria for testing, i.e., at least 80% of the total responses occurred on the correct lever, and fewer than five responses occurred on the incorrect lever before completion of the first FR on the correct lever within a cycle for all cycles during five consecutive or six of seven training sessions. Tests were conducted after performance for consecutive training sessions, including both vehicle and drug training sessions, satisfied the test criteria. The order of training with drug or vehicle was varied nonsystematically.
Drug Discrimination Testing.
During test sessions, five consecutive responses on either lever postponed the shock schedule. Dose-effect functions for CB agonists were determined by administering vehicle in the first cycle followed by doses increasing by 0.5 log unit in subsequent cycles. The dose-effect function included ineffective doses (i.e., doses producing responses predominantly on the vehicle lever) up to doses that produced more than 80% of responses on the Δ9-THC lever. The day after a dose-effect determination, vehicle-test sessions were conducted on consecutive days for 3 or 14 days by administering vehicle in the first cycle followed by sham in a second and third cycle; monkeys received Δ9-THC (1 mg/kg s.c.) for 3 or 14 days or vehicle for 14 days after each of these sessions. On the day after 3 or 14 days of treatment, a test session was conducted by administering vehicle in the first cycle followed by cumulative doses of the same test drug administered on the day before that particular treatment. After the 3-day treatment regimen, cumulative drug dose-effect tests were conducted every other day until sensitivity was not different from the initial dose-effect functions, as determined for individual monkeys. Vehicle-test sessions consisting of three cycles were conducted on intervening days. After the 14-day treatment regimen, cumulative dose-effect tests were conducted once every 7 days until sensitivity was not different from the initial dose-effect function, as determined for individual monkeys. Experimental sessions were not conducted on the 7 days between testing with cumulative doses of drug.
Drugs were studied within subjects, and only one test drug was studied per Δ9-THC treatment episode (i.e., 3 or 14 days). Successive episodes of daily Δ9-THC treatment (i.e., the number of days between the last day of treatment of one episode and the first day of treatment of the next episode) were separated by at least 30 days. Before the initial dose-effect tests before treatment, performance for consecutive training sessions, including both vehicle and drug training sessions, satisfied the test criteria. Vehicle and Δ9-THC training sessions preceding these test sessions varied nonsystematically.
Sensitivity to CP-55,940 was determined before and after once daily Δ9-THC (1 mg/kg) treatment for 3 days in one female and 3 males, and results were compared with those reported previously for Δ9-THC under the same experimental conditions (McMahon, 2011). Then, sensitivity to JWH-018 and JWH-073 was determined in nonsystematic order among these four monkeys.
Two males and two females were tested with Δ9-THC, CP-55,940, JWH-018, and JWH-073 before and after 14 days of Δ9-THC treatment. First, sensitivity to Δ9-THC and CP-55,940 was determined in counter-balanced order. Monkeys then received 14 days of vehicle treatment. Consequently, sensitivity to JWH-018 and JWH-073 was determined in counter-balanced order.
Drugs.
Δ9-THC (200 mg/ml in absolute ethanol; The Research Technology Branch, National Institute on Drug Abuse, Rockville, MD), CP-55,940 (Tocris Bioscience, Ellisville, MO), JWH-018 (IU Chemical Holding Co., Ltd., Shanghai, China), and JWH-073 (Research Chemical Supplier, Scottsdale, AZ) were dissolved in a mixture of 1 part absolute ethanol, 1 part Emulphor-620 (Rhodia Inc.), and 18 parts physiologic saline and were administered intravenously in a volume of 0.1 ml/kg. Doses were expressed as the weight of the forms listed above in milligrams per kilogram of body weight.
Data Analyses.
Discrimination data were expressed as a percentage of responses on the drug lever out of the total number of responses on both the drug and vehicle levers. Rate of responding on both levers (i.e., drug and vehicle) was calculated as responses per second excluding responses during timeouts. Rate of responding during a test was expressed as the percentage of the control response rate for individual animals. The control was defined as the average response rate for all cycles during the five previous vehicle training sessions excluding sessions during which the test criteria were not satisfied. Discrimination and rate data were averaged among subjects and plotted as a function of dose.
To estimate the ED50 value, or dose producing 50% responding on the drug lever, individual dose-response data were analyzed with linear regression (Prism version 5.0 for Windows; GraphPad Software Inc., San Diego, CA). The analyses included doses spanning the linear portion of the dose-response curve, and a common, best-fitting slope was used for further analyses (Kenakin, 1997). Doses corresponding to the 50% level of effect (ED50 value), potency ratios, and their 95% confidence limits were calculated by parallel line analyses of data from individual subjects (Tallarida, 2000). Potencies were considered significantly different when the 95% confidence limits of the potency ratio did not include 1.
To compare the magnitude of tolerance and cross-tolerance between drugs and the two treatment durations (3 versus 14 days), a dose ratio (ED50 after treatment/ED50 before treatment) was calculated for individual subjects, and significant differences were assessed with paired t tests (P < 0.05). The effects of drugs on response rate were examined with linear regression; a significant dose-dependent decrease in responding would be evidenced by the slope being significantly different from 0 (P < 0.05).
Results
Discriminative Stimulus Effects of Δ9-THC before and Immediately after 14 Days of Vehicle Treatment
In monkeys discriminating Δ9-THC, the training drug dose-dependently increased the percentage of responses on the Δ9-THC lever; at the training dose (0.1 mg/kg i.v.), the percentage responding on the Δ9-THC lever was 96% (Fig. 1, top, left, circles). After administration of vehicle, the percentage of responses on the Δ9-THC lever was 0% (Fig. 1, top, left, circles above VEH). Fourteen days of treatment with vehicle did not significantly decrease the potency of Δ9-THC to produce discriminative stimulus effects (P > 0.05; Fig. 1, left). ED50 values (95% confidence limits) before and after vehicle treatment were 0.042 (0.026–0.066) and 0.036 (0.020–0.066), respectively. Up to the training dose, Δ9-THC did not significantly modify response rate during the tests conducted before and after 14 days of vehicle treatment (P > 0.05).
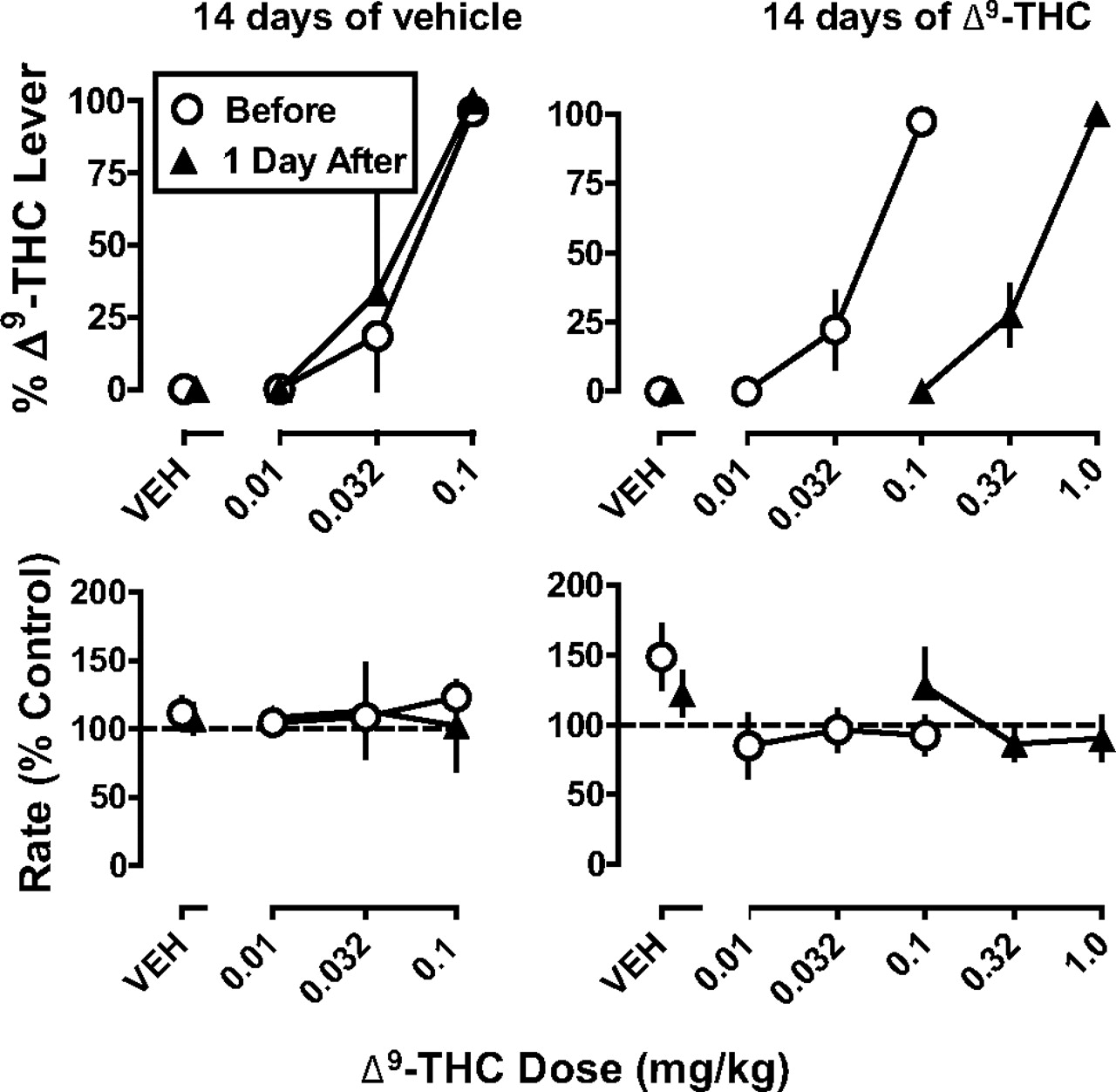
Discriminative stimulus effects of Δ9-THC in rhesus monkeys before and after 14 days of vehicle (left) or 1 mg/kg per day of Δ9-THC (right). Abscissae: vehicle (VEH) or dose administered intravenously of Δ9-THC in milligrams per kilogram of body weight. Ordinates: mean (±S.E.M.) percentage of responding on the Δ9-THC lever (top) and rate of responding expressed as a percentage of control (bottom).
Discriminative Stimulus Effects of Drugs before and Immediately after Δ9-THC Treatment
Induction of Tolerance to Δ9-THC.
Δ9-THC dose-dependently increased responding on the Δ9-THC lever, from 0% at a dose of 0.01 mg/kg to 97% at the training dose (0.1 mg/kg) (Fig. 1, top, right, circles). In contrast, vehicle produced 0% of the responses on the Δ9-THC lever (Fig. 1, top, right, circles above VEH). Fourteen days of treatment with Δ9-THC (1 mg/kg s.c.) produced a significant decrease in the potency of Δ9-THC to produce discriminative stimulus effects (Fig. 1, top, right) and increased the ED50 9.2-fold (Table 1). Sensitivity to Δ9-THC (i.e., the ED50 value) after 14 days of Δ9-THC treatment was no longer significantly different from control 17 days after treatment was discontinued (Table 1). Up to a dose of 1 mg/kg, Δ9-THC did not significantly modify response rate during the tests conducted before and after 14 days of Δ9-THC treatment (P > 0.05).
ED50 values and 95% confidence limits determined before and after Δ9-THC treatment (1 mg/kg per day s.c. for up to 14 days) in monkeys discriminating Δ9-THC
Cross-Tolerance from Δ9-THC to CP-55,940, JWH-073, and JWH-018
CP-55,940 (Fig. 2), JWH-073 (Fig. 3), and JWH-018 (Fig. 4) dose-dependently increased responding on the Δ9-THC lever; the ED50 values are shown in Table 1. When the dose-response curves were redetermined after 3 days of Δ9-THC (1 mg/kg) treatment, the potencies of CP-55,940 (Fig. 2, left), JWH-073 (Fig. 3, left), and JWH-018 (Fig. 4, left) were not significantly different from those of the respective controls (P > 0.05; Table 1). In contrast, 14 days of treatment with Δ9-THC (1 mg/kg s.c.) produced a significant decrease in the potency of CP-55,940 (Fig. 2, right), JWH-073 (Fig. 3, right), and JWH-018 (Fig. 4, right) to produce discriminative stimulus effects. Sensitivity was decreased 3.6-fold for CP-55,940, 4.3-fold for JWH-018, and 5.6-fold for JWH-073 (Table 1). The development of cross-tolerance to CP-55,940, JWH-073, and JWH-018 was time-dependent and reversible; the ED50 value was no longer significantly different from those of the respective controls 9 days after treatment was discontinued (Table 1). There was no significant relationship between dose of CP-55,940 (up to 0.032 mg/kg) and JWH-018 (up to 0.32 mg/kg) and rate of responding. JWH-073, at a dose of 1 mg/kg, tended to decrease response rate after 14 days of treatment.

Discriminative stimulus effects of CP-55,940 in rhesus monkeys before and after 3 (left) or 14 days (right) of Δ9-THC (1 mg/kg per day s.c.). Abscissae: vehicle (VEH) or dose administered intravenously of CP-55,940 in milligrams per kilogram of body weight. Ordinates: mean (±S.E.M.) percentage of responding on the Δ9-THC lever (top) and rate of responding expressed as a percentage of control (bottom).

Discriminative stimulus effects of JWH-073 in rhesus monkeys before and after 3 (left) or 14 days (right) of Δ9-THC (1 mg/kg per day s.c.). Abscissae: vehicle (VEH) or dose administered intravenously of JWH-073 in milligrams per kilogram of body weight. Ordinates: mean (±S.E.M.) percentage of responding on the Δ9-THC lever (top) and rate of responding expressed as a percentage of control (bottom). The rate data for the dose 1 mg/kg were averaged among two subjects.
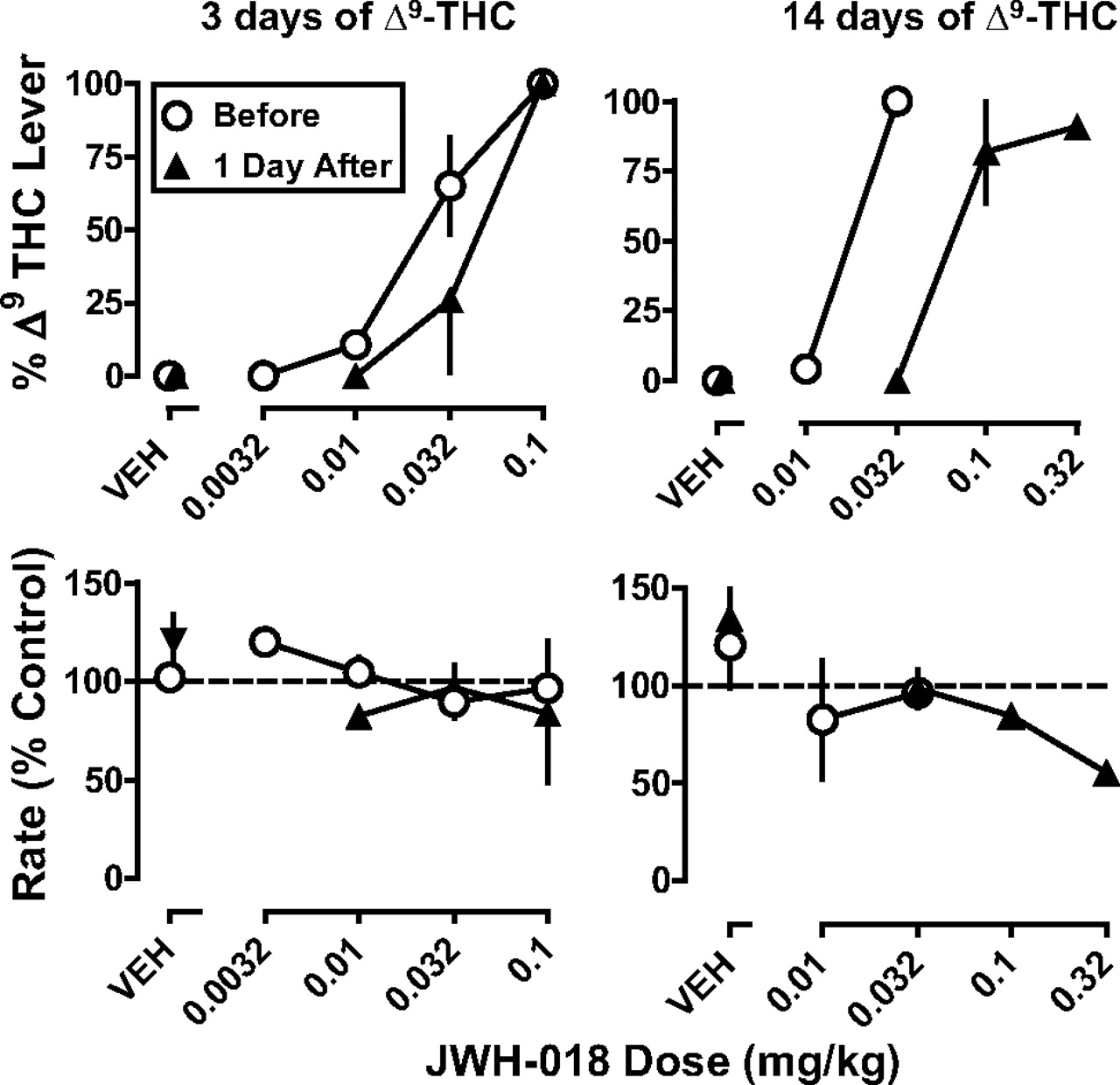
Discriminative stimulus effects of JWH-018 in rhesus monkeys before and after 3 (left) or 14 days (right) of Δ9-THC (1 mg/kg per day s.c.). Abscissae: vehicle (VEH) or dose administered intravenously of JWH-018 in milligrams per kilogram of body weight. Ordinates: mean (±S.E.M.) percentage of responding on the Δ9-THC lever (top) and rate of responding expressed as a percentage of control (bottom).
Comparing the Magnitude of Tolerance and Cross-Tolerance among CB1 Agonists
Figure 5 shows the mean dose ratio (i.e., tolerance/cross tolerance) calculated for individual monkeys as a function of test drug and duration of Δ9-THC treatment. T tests demonstrated that tolerance/cross-tolerance for every test drug was greater after 14 days of Δ9-THC treatment than after 3 days of Δ9-THC treatment (Fig. 5; P < 0.05). Even though significant tolerance to Δ9-THC developed after 3 days of treatment, the mean dose ratio (2.8-fold) was not significantly different from those of CP-55,940 (1.4-fold), JWH-073 (1.6-fold), and JWH-018 (1.8). On the other hand, after 14 days of Δ9-THC treatment, tolerance to Δ9-THC was significantly greater compared with cross-tolerance to CP-55,940 (P < 0.01) and JWH-018 (P < 0.05) but not to JWH-073 (P = 0.13).

Potency ratios of Δ9-THC, JWH-073, JWH-018, and CP-55,940 after 3 and 14 days of treatment with Δ9-THC. The potency ratios were determined as the ED50 after treatment divided by the ED50 before treatment. #, significantly different from the same drug after 14 days of Δ9-THC treatment. &, significantly different from Δ9-THC after 14 days of Δ9-THC treatment.
Discussion
In rhesus monkeys discriminating Δ9-THC (0.1 mg/kg i.v.) from vehicle, once-daily treatment with Δ9-THC (1 mg/kg s.c.) resulted in a loss of sensitivity to the discriminative stimulus effects of Δ9-THC and other CB1 receptor agonists. The magnitude of change in sensitivity was positively correlated with treatment duration. Loss of sensitivity was significantly greater after 14 days than 3 days of Δ9-THC treatment. The magnitude of change also varied as a function of test drug. As reported previously, 3 days of Δ9-THC (1 mg/kg s.c.) significantly decreased sensitivity to Δ9-THC; however, in the current study, 3 days of Δ9-THC did not modify sensitivity to CP-55,940, JWH-018, and JWH-073. Fourteen days of treatment further decreased sensitivity to Δ9-THC and resulted in a loss of sensitivity to CP-55,940, JWH-018, and JWH-073. However, the magnitude of change for CP-55,940 and JWH-018 was significantly less than that for Δ9-THC. Taken together, the data collected here and in a previous study (McMahon, 2011) demonstrate that tolerance and cross-tolerance develop to the discriminative stimulus effects of CBs and that the magnitude is inversely related to the CB1 receptor agonist efficacy of the drugs being tested for tolerance and cross-tolerance. In particular, there was a 2.6-fold difference in the magnitude of tolerance/cross-tolerance between Δ9-THC and CP-55,940 after 14 days of Δ9-THC treatment, compared with a ∼5-fold difference in efficacy as indexed by maximal stimulation of GTPγS binding in brain (Brents et al., 2011). JWH-018 and CP-55,940 did not differ from each other with respect to the magnitude of cross-tolerance or maximal stimulation of G proteins. However, cross-tolerance to JWH-073 was somewhat less than that to CP-55,940, consistent with JWH-073 having slightly lower efficacy than CP-55,940 (Brents et al., 2012).
Receptor theory predicts that a loss of receptor function confers a greater loss of sensitivity to low-efficacy agonists, as evidenced by decreased potency and perhaps maximal effect, compared with higher efficacy agonists (Kenakin, 1997). Differential tolerance and cross-tolerance that develop to μ-opioid agonists provide a striking confirmation of this prediction. Under identical conditions of chronic treatment with morphine or another μ agonist, greater tolerance and cross-tolerance develop to low-efficacy μ agonists compared with high-efficacy μ agonists (Stevens and Yaksh, 1989; Walker et al., 1997). A similar relationship is observed between drug sensitivity and spare μ-opioid receptor reserve. The magnitude of tolerance that develops to morphine is inversely related to spare receptor number (Porreca and Burks, 1983; Chavkin and Goldstein, 1984). CB and opioid receptors are G-protein-coupled, and agonists from both classes vary considerably in agonist efficacy, as evidenced by maximal stimulation of G proteins. Here, the importance of relative efficacy to in vivo effects was examined by measuring tolerance to Δ9-THC and cross-tolerance to higher efficacy CB1 receptor agonists including synthetic CBs with abuse potential.
A previous study (McMahon, 2011) in rhesus monkeys discriminating Δ9-THC (i.e., the assay used here) demonstrated that 3 days of Δ9-THC treatment produced moderate tolerance (3-fold) to the discriminative stimulus effects of Δ9-THC without cross-tolerance to CP-55,940 and WIN-55,212-2. Given that CP-55,940 and WIN-55,212-2 have higher CB1 receptor agonist efficacy than Δ9-THC, as evidenced by differences in maximal stimulation of GTPγS binding in brain (Breivogel and Childers, 2000), differences in tolerance and cross-tolerance to behavioral effects in rhesus monkeys appeared to be due to corresponding differences in CB1 receptor agonist efficacy. Here, Δ9-THC treatment duration was increased to examine relationships between treatment duration and tolerance, as well as the extent to which cross-tolerance develops to discriminative stimulus effects. JWH-018 and JWH-073 were of interest given the increased abuse of these and other synthetic CBs. The current study replicated the results of the previous study, i.e., 3 days of Δ9-THC treatment did not result in cross-tolerance to CP-55,940 (McMahon, 2011), and extended them to JWH-018 and JWH-073, which also were not susceptible to cross-tolerance after 3 days of Δ9-THC treatment. Increasing Δ9-THC treatment to 14 days resulted in 9.4-fold tolerance and significant, albeit less, cross-tolerance to JWH-018 (4.3-fold) and CP-55,940 (3.6-fold).
The pattern of tolerance and cross-tolerance obtained in this study is highly consistent with a loss of CB1 receptor function and differences in CB1 receptor agonist efficacy. Tolerance to Δ9-THC in rodents is accompanied by a marked decrease in CB1 receptor number and desensitization (Sim et al., 1996; Breivogel et al., 1999; Selley et al., 2004; Falenski et al., 2010), i.e., uncoupling to G proteins. Tolerance to the discriminative stimulus effects of Δ9-THC in rhesus monkeys is consistent with CB1 receptor down-regulation and desensitization. Moreover, the predictable relationship between the magnitude of tolerance and cross-tolerance and the CB1 receptor agonist efficacy of the test drugs is further consistent with loss of CB1 receptor function. Taken together, estimates of in vitro efficacy reported previously (Brents et al., 2011) and the current results in nonhuman primates showing greater tolerance to a low-efficacy agonist (Δ9-THC) compared with cross-tolerance to higher efficacy agonists (JWH-018 and CP-55,940) suggest that human cannabis users (relative to nonusers) are unexpectedly more sensitive to synthetic CBs than to cannabis. The extent to which this might underlie adverse effects reported after use of herbal blends containing synthetic CBs warrants further investigation.
Drug discrimination involves repeated training; when training is decreased or suspended, there is the potential for the training drug to lose the control it exerts over choice behavior. To examine a loss of control by the drug, often referred to as stimulus control, monkeys received 14 days of vehicle treatment. Afterward, there was no change in sensitivity to the discriminative stimulus effects of Δ9-THC. Similar results were obtained in rats (Wiley et al., 1993): tolerance to the discriminative stimulus effects of Δ9-THC was not accompanied by a change in the potency of Δ9-THC after vehicle treatment in the absence of discrimination training. Each time there was a decrease in potency in the current study, cessation of Δ9-THC treatment resulted in a time-related return to control sensitivity despite the absence of any training. This is further evidence of tolerance and cross-tolerance without loss of stimulus control. The time required for loss or reversal of tolerance/cross-tolerance after cessation of Δ9-THC treatment was greater for Δ9-THC (e.g., more than 9 days) compared with CP-55,940, JWH-018, and JWH-073 (e.g., 9 days or less), a pattern consistent with differences in CB1 receptor agonist efficacy.
Differences in tolerance and cross-tolerance could result from differences among agonists in binding selectivity for two receptor types that mediate a common effect of the agonists. Positive GABAA receptor modulators acting at distinct receptor sites and sharing discriminative stimulus effects provide a striking example. Treatment with a positive GABAA receptor modulator acting at benzodiazepine sites (i.e., benzodiazepines) results in tolerance to other benzodiazepines but not cross-tolerance to positive GABAA modulators acting at nonbenzodiazepine sites (e.g., neuroactive steroid or barbiturate sites; Gerak et al.; 2008). However, there is no evidence that a difference in binding affinity at multiple receptor types accounts for the current results. Although Δ9-THC and the other drugs tested here are CB2 receptor agonists, CB2 receptor antagonists do not antagonize the discriminative stimulus effects of Δ9-THC (McMahon, 2006). Moreover, quantitative (Schild) analyses of antagonism of all four agonists used in the current study are highly consistent with the same rimonabant-sensitive CB1 receptors mediating their discriminative stimulus effects (McMahon, 2006; Ginsburg et al., 2012). The Δ9-THC discrimination assay used in the current study appears to be selectively mediated by CB1 receptor agonism.
In summary, the present results strongly suggest that JWH-018 and CP-55,940 have higher efficacy than Δ9-THC in vivo, whereas it appears that JWH-073 has efficacy intermediate to that of Δ9-THC and JWH-018. The higher efficacy of some synthetic CBs found in herbal blends used instead of or in addition to cannabis could be of particular concern inasmuch as abuse, dependence, and other adverse effects stemming from use of those products, as described previously (Zimmermann et al., 2009; Müller et al., 2010; Vearrier and Osterhoudt, 2010; Schneir et al., 2011; Simmons et al., 2011), are positively related to CB1 receptor agonist efficacy.
Authorship Contributions
Participated in research design: Hruba, Ginsburg, and McMahon.
Conducted experiments: Hruba.
Performed data analysis: Hruba and McMahon.
Wrote or contributed to the writing of the manuscript: Hruba and McMahon.
Acknowledgments
We thank D. Schulze for technical assistance.
Footnotes
This work was supported by the National Institutes of Health National Institute on Drug Abuse [Grant DA19222].
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
ABBREVIATIONS:
- CB
- cannabinoid
- Δ9-THC
- Δ9-tetrahydrocannabinol
- JWH-018
- naphthalen-1-yl-(1-pentylindol-3-yl) methanone
- JWH-073
- naphthalen-1-yl-(1-butylindol-3-yl) methanone
- WIN-55,212-2
- 4,5-dihydro-2-methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo[3,2,1-i,j]quinolin-6-one
- GTPγS
- guanosine 5′-O-(3-[35S]thio)triphosphate
- FR
- fixed ratio
- CP-55,940
- (1R,3R,4R)-3-[2-hydroxy-4-(1,1-dimethylheptyl) phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol.
- Received May 15, 2012.
- Accepted June 19, 2012.
- Copyright © 2012 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}