


Abstract
We have examined the role of the human organic cation transporters [hOCTs and human novel organic cation transporter (hOCTN); SLC22A1–5] and apical multidrug and toxin extrusion (hMATE) in the cellular accumulation and cytotoxicity of platinum agents using the human embryonic kidney (HEK) 293 cells transiently transfected with the transporter cDNAs. Both the cytotoxicity and accumulation of cisplatin were enhanced by the expression of hOCT2 and weakly by hOCT1, and those of oxaliplatin were also enhanced by the expression of hOCT2 and weakly by hOCT3. The hOCT-mediated uptake of tetraethylammonium (TEA) was markedly decreased in the presence of cisplatin in a concentration-dependent manner. However, oxaliplatin showed almost no influence on the TEA uptakes in the HEK293 cells expressing hOCT1, hOCT2, and hOCT3. The hMATE1 and hMATE2-K, but not hOCTN1 and OCTN2, mediated the cellular accumulation of cisplatin and oxaliplatin without a marked release of lactate dehydrogenase. Oxaliplatin, but not cisplatin, markedly decreased the hMATE2-K-mediated TEA uptake. However, the inhibitory effect of cisplatin and oxaliplatin against the hMATE1-mediated TEA uptake was similar. The release of lactate dehydrogenase and the cellular accumulation of carboplatin and nedaplatin were not found in the HEK293 cells transiently expressing these seven organic cation transporters. These results indicate that cisplatin is a relatively good substrate of hOCT1, hOCT2, and hMATE1, and oxaliplatin is of hOCT2, hOCT3, hMATE1, and hMATE2-K. These transporters could play predominant roles in the tissue distribution and anticancer effects and/or adverse effects of platinum agent-based chemotherapy.
Four platinum-based anticancer drugs are currently registered for clinical use. cis-Diamminedichloroplatinum II (cisplatin) has been clinically used for over 30 years and continues to play an essential role in cancer chemotherapy against solid tumors of the prostate, bladder, colon, lung, testis, liver, and brain (Ho et al., 2003). However, severe nephrotoxicity limits its clinical application because it was reported that an increase in the serum creatinine concentration was observed in 41% of patients treated with a high dose of cisplatin (de Jongh et al., 2003). cis-Diammine(1,1-cyclobutanedicarboxylato)platinum II (carboplatin), trans-l-1,2-diaminocyclohexaneoxalatoplatinum II (oxaliplatin), and cis-diammineglycolatoplatinum (nedaplatin) are analogs of cisplatin and show a lowered nephrotoxicity compared with cisplatin. However, it is not clear why the nephrotoxicity of these analogs is low, although cisplatin induces severe nephrotoxicity. The chemical structures of platinum agents are shown in Fig. 1.
The mechanisms of cellular uptake and efflux of platinum agents are not fully understood. In our laboratory, the cisplatin uptake was reported to be mediated by specific transporter(s) in the renal tubular cells (Okuda et al., 1999). Recently, we reported that rat kidney-specific organic cation transporter 2 (rOCT2/Slc22a2) transported cisplatin and was responsible for cisplatin-induced nephrotoxicity (Yonezawa et al., 2005). Just after our report, hOCT2 was reported to transport cisplatin (Ciarimboli et al., 2005). However, it is not clear whether these analogs were transported by OCT2.
The organic ion transporter family (SLC22) consists of the organic anion transporter (OAT), OCT, and novel organic cation transporter (OCTN), which transport drugs, toxins and endogenous metabolites (Inui et al., 2000; Jonker and Schinkel, 2004). hOCT1 is expressed mainly in the liver, hOCT2 in the kidney (Gorboulev et al., 1997), and hOCT3 is predominantly expressed in the placenta and weakly in the intestine, heart, brain, and kidney (Kekuda et al., 1998). The substrate specificities of hOCT1, hOCT2, and hOCT3 have been considered to not be the same. hOCTN1 and hOCTN2 were expressed not only in the kidney but also in other tissues. They were expressed in the apical membrane of renal proximal tubules. Human multidrug and toxin extrusion 1 (hMATE1) was recently cloned and characterized (Otsuka et al., 2005). It was expressed in the liver, kidney, and skeletal muscle. Moreover, we cloned hMATE2-K, which is specifically expressed in the apical membrane of renal epithelial cells and transports cationic drugs (Masuda et al., 2006).

Chemical structures of four platinum agents.
Based on these backgrounds, we hypothesized that organic cation transporters could transport four platinum drugs and play an important role in the cellular accumulation and toxicity of platinum agents. In the present study, we examined whether the basolateral cation transporters hOCT1, hOCT2, and hOCT3, and the apical cation transporters, hMATE1, hMATE2-K, hOCTN1, and hOCTN2, transported platinum agents and affected the cytotoxicity induced by these drugs.
Materials and Methods
Cell Culture and Transfection. HEK293 cells (CRL-1573; American Type Culture Collection, Manassas, VA) were cultured in complete medium consisting of Dulbecco's modified Eagle's medium (Sigma Chemical Co., St. Louis, MO) with 10% fetal bovine serum (Whittaker Bioproducts Inc., St. Louis, MO) in an atmosphere of 5% CO2-95% air at 37°C.
For a transient expression system, pCMV6-XL4 plasmid vector DNA (OriGene Technologies, Rockville, MO), pcDNA3.1(+) plasmid vector DNA (Invitrogen, Carlsbad, CA), or pBK-CMV plasmid vector DNA (Stratagene, La Jolla, CA) containing hOCT1, hOCT2, hOCT3, hMATE1, hMATE2-K, hOCTN1, hOCTN2, hOAT1, and hOAT3 cDNA was purified using Midi-V100 Ultrapure Plasmid Extraction Systems (Viogene, Sunnyvale, CA). The day before transfection, HEK293 cells were seeded onto poly-d-lysine-coated 24-well plates at a density of 2.0 × 105 cells per well. The cells were transfected with 800 ng of plasmid DNA per well in a combination of empty vector and transporter cDNA using 2 μl of Lipofectamine 2000 (Invitrogen) per well according to the manufacturer's instructions. The amount of transporter cDNA was 100 ng except in the experiment examining the transporter cDNA-dependence. Forty-eight hours after the transfection, the cells were used for the experiments.
Uptake Experiment. Cellular uptake of tetraethylammonium (TEA; 88.8 MBq/mmol; PerkinElmer Life Analytical Sciences, Boston, MA), [3H]1-methyl-4-phenylpyridium acetate ([3H]MPP; 2.7 TBq/mmol; PerkinElmer), p-[14C]aminohippurate ([14C]PAH; 1.9 GBq/mmol; PerkinElmer), and [6,7-3H(N)]esterone-3-sulfate, ammonium salt ([3H]ES; 2.1 TBq/mmol, PerkinElmer) was measured with HEK293 cells grown on poly-d-lysine-coated 24-well plates. The composition of the incubation buffer was as follows: 145 mM NaCl, 3 mM KCl, 1 mM CaCl2, 0.5 mM MgCl2, 5 mM d-glucose, and 5 mM HEPES, pH 7.4 or pH 8.4 adjusted with NaOH. Experimental procedures were performed as described previously (Urakami et al., 2004).
To measure the cellular accumulation of the platinum agents, HEK293 cells were seeded on poly-d-lysine-coated 24-well plates. Cells were incubated in Dulbecco's modified Eagle's medium with 10% fetal bovine serum containing cisplatin (Sigma), carboplatin (Sigma), oxaliplatin (a gift from Yakult Co. Ltd., Tokyo, Japan), and nedaplatin (LKT Laboratories, Inc., St. Paul, MN). After the incubation, the monolayers were rapidly washed twice with ice-cold incubation buffer containing 3% bovine serum albumin (Nacalai Tesque Inc., Kyoto, Japan) and then washed three times with ice-cold incubation buffer. The cells were solubilized in 0.5 N NaOH, and the amount of platinum was determined using inductively coupled plasma-mass spectrometry (ICP-MS) by the Pharmacokinetics and Bioanalysis Center, Shin Nippon Biomedical Laboratories, Ltd. (Wakayama, Japan). The protein content of the cell monolayers solubilized in 0.5 N NaOH was determined with a Bio-Rad protein assay kit (Bio-Rad Laboratories, Richmond, CA).
Measurement of Released Lactate Dehydrogenase Activity. The cytotoxicity of the platinum agents was measured with HEK293 cells seeded on poly-d-lysine-coated 24-well plates. Cells were incubated with the medium containing platinum agent with or without cimetidine (Nacalai) for 2 h. After the removal of the medium, a drug-free medium was added. After incubation for 24 h, the medium was collected, and the lactate dehydrogenase (LDH) activity in the medium was measured using an LDH cytotoxicity detection kit (Takara Bio Inc., Shiga, Japan), according to the manufacturer's instructions. Cytotoxicity was evaluated by measuring the LDH activity in the medium. The total LDH activity was defined as the LDH activity in the medium containing 1% Triton X-100. LDH release (percentage) represents (LDH activity – LDH activity of control)/(total LDH activity – LDH activity of control) × 100.
Statistical Analysis. Data are expressed as means ± S.E.M. Data were analyzed statistically using the unpaired Student's t test. Multiple comparisons were performed with Dunnett's two-tailed test after a one-way ANOVA. Probability values of less than 0.05 were considered statistically significant.
Results
Cytotoxicity of Cisplatin in HEK293 Cells Transiently Expressing hOCT2. We examined the expression level-dependent effect of hOCT2 on the cisplatin-induced cytotoxicity. When HEK293 cells transfected with 50 to 800 ng of hOCT2 cDNA per well were treated with 500 μM cisplatin for 2 h and subsequently cultured in normal medium for 24 h, the release of LDH into the culture medium was increased in a hOCT2 expression-dependent manner (Fig. 2A). Based on these results, we decided that cells were transfected with 100 ng of transporter cDNA. Moreover, cimetidine inhibited the cytotoxicity induced by 500 μM cisplatin in a concentration-dependent manner with an IC50 value of 109.3 ± 4.4 μM (mean ± S.E.M. of three separate experiments) in hOCT2-expressing cells (Fig. 2B).

Role of hOCT2 expression in the cisplatin-induced cytotoxicity. A, HEK293 cells were transfected with each amount of hOCT2 cDNA and vector plasmid added to 800 ng using 2 μl of Lipofectamine 2000. The cells were exposed to 500 μM cisplatin in the medium for 2 h and then incubated in normal medium for 24 h. LDH released into the medium was measured. B, HEK293 cells were transfected with each amount of 100 ng of hOCT2 cDNA and 700 ng of vector plasmid. The cells were incubated in the medium containing 500 μM cisplatin with 0 to 1000 μM cimetidine for 2 h and then incubated in normal medium for 24 h. LDH released into the medium was measured. Each point represents the mean ± S.E.M. of three wells.
Effects of Expression of Basolateral Organic Ion Transporters, hOCT1, hOCT2, hOCT3, hOAT1, and hOAT3, on the Cisplatin-Induced Cytotoxicity in HEK293 Cells. We examined the effect of the basolateral organic ion transporter expression on cisplatin-induced cytotoxicity. When the cells transiently expressing hOCT1, hOCT2, and hOCT3 were treated with 50 to 1000 μM cisplatin for 2 h, the cisplatin-induced cytotoxicity was strongly enhanced by the expression of hOCT2, weakly by the expression of hOCT1, but not by hOCT3 (Fig. 3A). When the cells were transfected with 100 or 800 ng of hOAT1 or hOAT3 cDNA, the cytotoxicity of hOAT-expressing cells was not induced by 500 μM cisplatin (Fig. 3D). In addition, these transporter activities were confirmed by the uptake of their typical substrates, TEA, MPP, PAH, and ES, as shown in Fig. 3, B, C, E, and F.
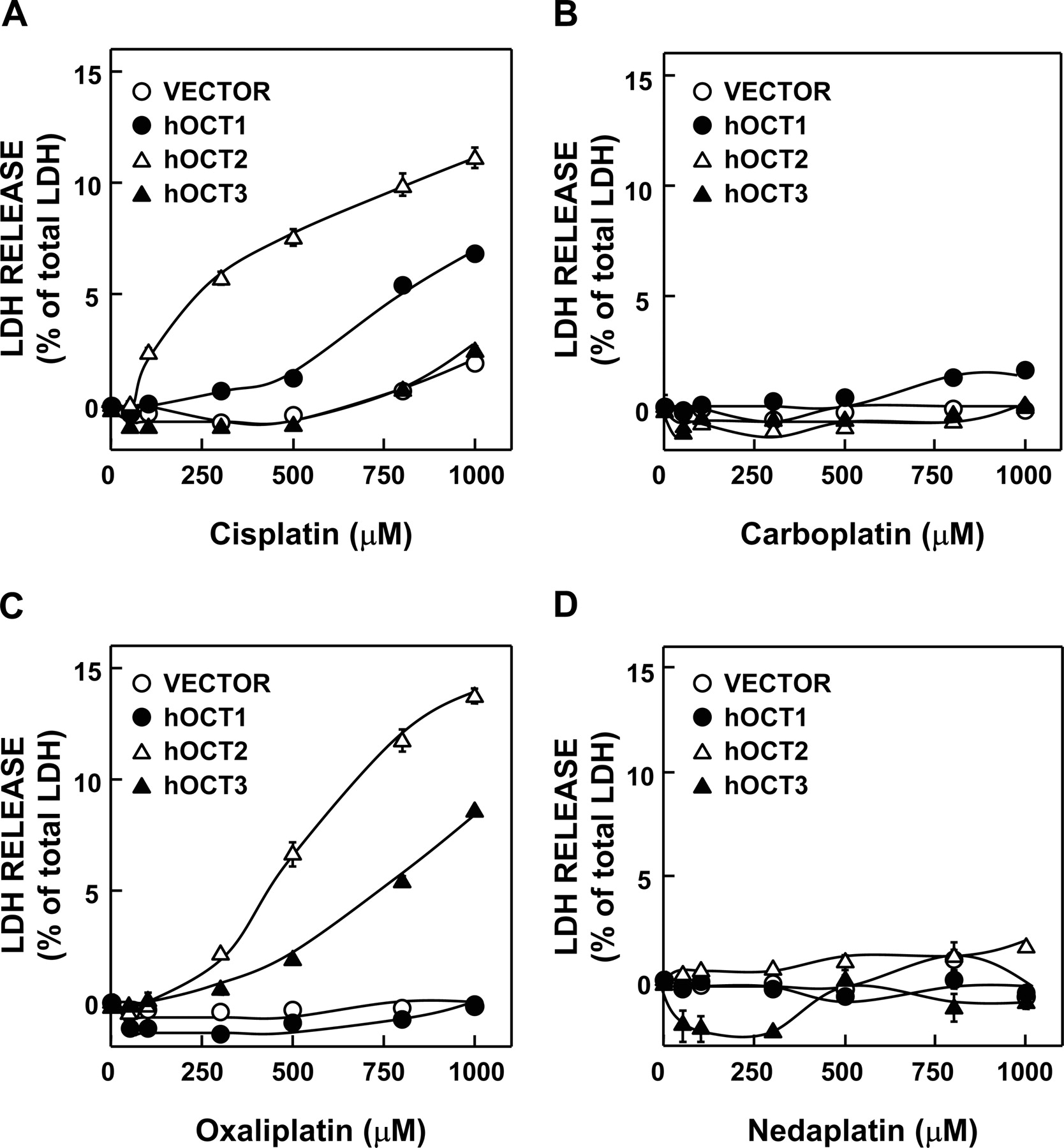
Effects of hOCT1, hOCT2, and hOCT3 Expression on Platinum Agent-Induced Cytotoxicity in HEK293 Cells. The effects of hOCT1, hOCT2, and hOCT3 expression on the platinum agent-induced cytotoxicity were simultaneously examined using the same passage cells in Fig. 4. When the cells transfected with 100 ng of hOCT1, hOCT2, and hOCT3 were treated with 50 to 1000 μM cisplatin, the highest LDH release in hOCT2-expressing cells among these OCT-expressing cells was observed. Moreover, hOCT1 also enhanced the cisplatin-induced cytotoxicity at higher concentrations, but not hOCT3 (Fig. 4A). The reproducibility (Figs. 3A and 4A) was confirmed in these transfectants, and then we compared the susceptibility of these transfectants to analogs of cisplatin. Oxaliplatin also induced cytotoxicity in HEK293 cells expressing hOCT2 and hOCT3 (Fig. 4C). The treatment with carboplatin and nedaplatin (50–1000 μM) did not affect the LDH release in hOCT1-, hOCT2-, and hOCT3-expressing cells (Fig. 4, B and D).
Transport of Platinum Agents by HEK293 Cells Transiently Expressing Basolateral Organic Cation Transporters, hOCT1, hOCT2, and hOCT3. We compared the accumulations of platinum when HEK293 cells transiently expressing hOCT1, hOCT2, and hOCT3 were treated with the medium containing platinum agents for 1 h. The accumulation of platinum after the treatment with cisplatin was increased in a dose-dependent manner and was much higher in hOCT2-expressing cells and weakly higher in hOCT1-expressing cells (Fig. 5A). When cells were treated with oxaliplatin, it was much higher in hOCT2-expressing cells and slightly in hOCT3-expressing cells (Fig. 5C). The accumulation of carboplatin and nedaplatin was not increased by the expression of hOCT1–3 (Fig. 5, B and D).
Inhibitory Effects of Cisplatin and Oxaliplatin on the [14C]TEA Uptake by HEK293 Cells Expressing Basolateral Organic Cation Transporters, hOCT1, hOCT2, and hOCT3. We compared the affinities of cisplatin and oxaliplatin with hOCT1, hOCT2, and hOCT3 at a concentration range between 50 and 5000 μM because of their solubility. The uptake of [14C]TEA by hOCT1- and hOCT2-expressing cells was partially inhibited by cisplatin but not oxaliplatin at a concentration range between 50 and 5000 μM (Fig. 6, A and B). The inhibitory effect of cisplatin on the uptake of [14C]TEA by hOCT3-expressing cells was stronger than that of oxaliplatin (Fig. 6C).
Effects of the Expression of Apical Organic Cation Transporters, hMATE1, hMATE2-K, hOCTN1, and hOCTN2, on the Cisplatin-Induced Cytotoxicity in HEK293 Cells. When the cells transiently expressing hMATE1, hMATE2-K, hOCTN1, and hOCTN2 were treated with 50 to 1000 μM cisplatin for 2 h, the expression of hMATE1, hMATE2-K, hOCTN1, or hOCTN2 did not affect the cisplatin-induced cytotoxicity (Fig. 7, A and B). In addition, these transporter activities were confirmed by the up-take of [14C]TEA as shown in Fig. 7, C and D.
Transport of Platinum Agents by HEK293 Cells Transiently Expressing Apical Organic Cation Transporters, hMATE1, hMATE2-K, hOCTN1, and hOCTN2. The cellular accumulation of platinum was determined in HEK293 cells expressing hMATE or hOCTN when the cells were treated with 500 μM platinum agents for 1 h. The accumulation of cisplatin was enhanced by hMATE1 more than hMATE2-K. On the other hand, the accumulation of oxaliplatin was enhanced by hMATE2-K more than hMATE1. These transporters could transport cisplatin and oxaliplatin in culture medium without an artificial pH gradient (Fig. 8A). hOCTN1 or hOCTN2 did not stimulate the transport of platinum agents (Fig. 8B).

Effect of basolateral organic ion transporters expression on the cisplatin-induced cytotoxicity. A, HEK293 cells were transfected with empty vector (open circle), hOCT1 (closed circle), hOCT2 (open triangle), or hOCT3 (closed triangle). The cells were treated with the medium containing 50 to 1000 μM cisplatin for 2 h and then incubated in normal medium for 24 h. D, HEK293 cells were transfected with empty vector, hOAT1, and hOAT3 (100 or 800 ng/well). The cells were treated with the medium containing 500 μM cisplatin for 2 h and then incubated in normal medium for 24 h. LDH released into the medium was measured. Uptake of typical substrates, TEA, MPP, PAH, and ES, by HEK293 cells expressing hOCT1, hOCT2, hOCT3, hOAT1, and hOAT3 is shown. HEK293 cells were transfected with empty vector, hOCT1–3 (B and C), hOAT1 (E), or hOAT3 (F). The cells were incubated with the incubation buffer containing 50 μM [14C]TEA (B), 13.7 nM [3H]MPP (C), 5 μM [14C]PAH (E), or 20 nM [3H]ES (F) at pH 7.4 for various periods at 37°C. The amount of substrates in the cells was determined by measuring the radioactivity of solubilized cells. Each point and each bar represent the mean ± S.E.M. of three wells.
Inhibitory Effects of Cisplatin and Oxaliplatin on [14C]TEA Uptake by HEK293 Cells Expressing hMATE1 and hMATE2-K. To examine the uptake of [14C]TEA by hMATE1- and hMATE2-K-expressing cells, the cells were preincubated in the buffer containing ammonium chloride (30 mM) to make an artificial pH-gradient and then were incubated in the buffer, pH 7.4, containing 50 μM [14C]TEA with or without cisplatin or oxaliplatin for 2 min. Cisplatin and oxaliplatin similarly inhibited the uptake of [14C]TEA by hMATE1-expressing cells (Fig. 9A). On the other hand, cisplatin did not inhibit the uptake of [14C]TEA by HEK293 cells expressing hMATE2-K, although oxaliplatin did (Fig. 9B).
Discussion
We previously reported that rOCT2 was responsible for the cisplatin-induced renal tubular toxicity (Yonezawa et al., 2005). In that report, the expression of rOCT2 enhanced the cisplatin-induced cytotoxicity in HEK293 cells, and rOCT2 played a role in the tissue distribution of cisplatin and the cisplatin-induced renal tubular toxicity in the rat. In the present study, we progressively indicated that hOCT2 also abundantly transported oxaliplatin as well as cisplatin, but not carboplatin and nedaplatin (Figs. 4 and 5). In addition, cisplatin and oxaliplatin were transported by hOCT1 and hOCT3, respectively. These results suggested that the basolateral hOCTs are the determinants for the tissue distribution of platinum agents from the circulation. Moreover, apical transporter hMATE1 and hMATE2-K also mediated the transport of cisplatin and oxaliplatin (Fig. 8A). This is the first report indicating that cisplatin was transported by hOCT1, hMATE1 and hMATE2-K, as well as hOCT2, and oxaliplatin was transported by hOCT2, hOCT3, hMATE1, and hMATE2-K.

Effect of hOCT1–3 expression on the platinum agent-induced cytotoxicity. HEK293 cells were transfected with empty vector (open circle), hOCT1 (closed circle), hOCT2 (open triangle), or hOCT3 (closed triangle). The cells were treated with the medium containing 50 to 1000 μM cisplatin (A), carboplatin (B), oxaliplatin (C), and nedaplatin (D) for 2 h and then incubated in normal medium for 24 h. LDH released into the medium was measured. Each point represents the mean ± S.E.M. of three wells.

Uptake of platinum agents by HEK293 cells expressing hOCT1, hOCT2, and hOCT3. HEK293 cells were transfected with empty vector (open circle), hOCT1 (closed circle), hOCT2 (open triangle), or hOCT3 (closed triangle). The cells were treated with the medium containing 100, 500, and 1000 μM cisplatin (A), carboplatin (B), oxaliplatin (C), and nedaplatin (D) for 1 h. After washing, the cells were solubilized in 0.5 N NaOH, and the amount of platinum was determined by ICP-MS. Each point represents the mean ± S.E.M. of four wells.
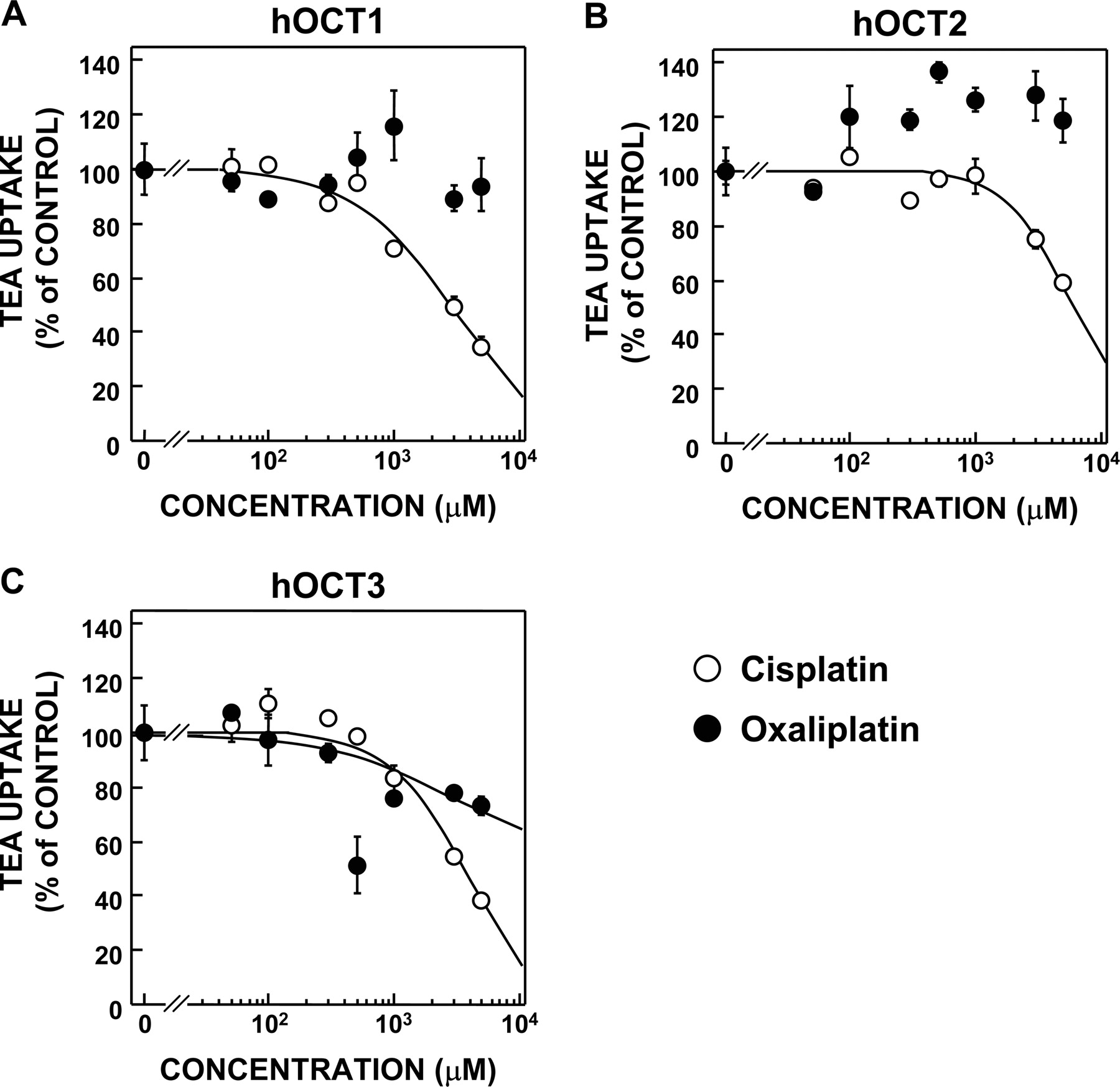
Inhibitory effects of cisplatin and oxaliplatin on the uptake of [14C]TEA by HEK293 cells expressing hOCT1 (A), hOCT2 (B), and hOCT3 (C). The cells were preincubated in the incubation buffer for 10 min and then were incubated in the buffer containing 50 μM [14C]TEA for 2 min with or without cisplatin and oxaliplatin at various concentrations. The amount of [14C]TEA in the cells was determined by measuring the radioactivity of solubilized cells. Each point represents the mean ± S.E.M. of three wells.

Effect of apical organic cation transporters expression on cisplatin-induced cytotoxicity. A, HEK293 cells were transfected with empty vector (open circle), hMATE1 (closed circle), or hMATE2-K (open triangle). B, HEK293 cells were transfected with empty vector (open circle), hOCTN1 (closed circle), and hOCTN2 (open triangle). The cells were treated with the medium containing 50 to 1000 μM cisplatin for 2 h and then incubated in normal medium for 24 h. LDH released into the medium was measured. The uptake of typical substrates by HEK293 cells expressing hMATE1, hMATE2-K, hOCTN1, and hOCTN2. HEK293 cells were transfected with empty vector, hMATE1, or hMATE2-K (C) and hOCTN1 or hOCTN2 (D). The cells were incubated with the incubation buffer at pH 8.4 (C) or 7.4 (D) containing 50 μM [14C]TEA for 10 min at 37°C. The amount of substrates in the cells was determined by measuring the radioactivity of solubilized cells. Each point and each bar represent the mean ± S.E.M. of three wells.
Cisplatin is commonly used against hepatocellular carcinoma, administered from a catheter in the hepatic artery. The systemic blood concentration of cisplatin after its administration from the hepatic artery was not different from that after i.v. administration. Thus, the probability of the adverse effect was not different despite the advantages regarding antitumor efficacy (Lo et al., 2002). It was reported that hepatic OCT1 was expressed in rat diethylnitrosamine-induced hepatocarcinoma cells and differentiated Fao cells, a hepatoma cell line (Lecureur et al., 1998). Interestingly, the cytotoxicity of cisplatin was also enhanced by hOCT1 at a relatively higher concentration (Figs. 3A and 4A). Therefore, hOCT1 was suggested to play an important role in the hepatic uptake of cisplatin.

Uptake of platinum agents by HEK293 cells expressing hMATE1, hMATE2-K, hOCTN1, and hOCTN2. A, HEK293 cells were transfected with empty vector (open bar), hMATE1 (gray bar), and hMATE2-K (black bar). B, HEK293 cells were transfected with empty vector (open bar), hOCTN1 (gray bar), and hOCTN2 (black bar). The cells were treated with the medium containing 500 μM platinum agents for 1 h. After washing, the cells were solubilized in 0.5 N NaOH, and the amount of platinum was determined by ICP-MS. Each bar represents the mean ± S.E.M. of four wells. **, P < 0.01, significantly different from vector-transfected cells.

Inhibitory effects of cisplatin and oxaliplatin on the uptake of [14C]TEA by HEK293 cells expressing hMATE1 (A) and hMATE2-K (B). The cells were preincubated in the incubation buffer containing 30 mM ammonium chloride for 20 min and then were incubated in the buffer containing 50 μM [14C]TEA for 2 min with or without cisplatin and oxaliplatin at various concentrations. The amount of [14C]TEA in the cells was determined by measuring the radioactivity of solubilized cells. Each point represents the mean ± S.E.M. of three wells.
Oxaliplatin has a different spectrum of activity and the different mechanisms of action and resistance from those of cisplatin and carboplatin (Raymond et al., 2002; Fuertes et al., 2003; Wang and Lippard, 2005). Carboplatin, oxaliplatin, and nedaplatin are low nephrotoxic agents despite their good efficacy. Therefore, it had been assumed that they would not be transported by hOCT2. Expectedly, carboplatin and nedaplatin were not transported by hOCT2 (Figs. 4, B and D, and 5, B and D). Surprisingly, oxaliplatin was transported by hOCT2 (Figs. 4C and 5C), although oxaliplatin did not induce nephrotoxicity (Raymond et al., 2002). In addition, oxaliplatin was transported by hMATE1 and hMATE2-K (Fig. 8A). The nephrotoxic pattern of oxaliplatin observed in the renal slice with collapsed and nonperfused lumens resembled to that of cisplatin (Kanou et al., 2004). It is likely that oxaliplatin would be accumulated in the renal slice without efflux systems, thereby exacerbating its toxicity. Based on these findings and the present results, it is presumed that the basolateral hOCT2 is an influx transporter as the inducer of oxaliplatin-induced toxicity, and the apical hMATE1 and hMATE2-K are efflux transporters as a means to protect cells. Therefore, transcellular transport and cellular toxicity of oxaliplatin should be further examined using polarized double transfectants with both basolateral OCT and apical MATE transporters.
hOCT3 is the organic cation transporter predominantly expressed in the placenta and widely expressed in tissues, including the kidney and intestine (Kekuda et al., 1998). The substrate specificity and pharmacokinetic role of hOCT3 are not fully understood. We identified oxaliplatin as a hOCT3 substrate, but not cisplatin (Fig. 5, A and C). In addition, there is a previous report that oxaliplatin had a more potent cytotoxicity than cisplatin against colon cancer cells, although the molecular mechanism was not clear (Ducreux et al., 1998). Because hOCT3 was reported to be expressed in the intestine (Kekuda et al., 1998), it was supposed that hOCT3 expression would contribute at least in part to the oxaliplatin sensitivity in colon cancer cells.
hMATE1, hMATE2-K, hOCTN1, and hOCTN2 are expressed in the apical membrane of renal proximal tubules. In this study, cisplatin and oxaliplatin were weakly but significantly transported by hMATE1 and hMATE2-K, but not carboplatin and nedaplatin (Fig. 8A). In contrast, hOCTN1 or hOCTN2 did not transport four platinum agents (Fig. 8B). Because the H+-gradient stimulated the hMATE1- (Otsuka et al., 2005), hMATE2-K- (Masuda et al., 2006), and rMATE1- (Terada et al., 2006) mediated transport of cationic compounds, the artificial H+-gradient is necessary as the driving force for the substrate transport. Intracellular acidification by ammonium chloride markedly stimulated the hMATE1-, hMATE2-K-, and rMATE1-mediated uptake of [14C]TEA. As shown in Fig. 8A, the cells were incubated in the culture medium containing platinum agents without an artificial H+-gradient because of the stability of platinum agents, as described under Materials and Methods. It is possible that the transport activities of hMATE1 and hMATE2-K for cisplatin and oxaliplatin are much higher at the renal apical membrane of proximal tubules with an oppositely directed H+-gradient in the physiological condition.
The clearance of cisplatin was reported to exceed the glomerular filtration rate, suggesting that cisplatin was secreted across the renal tubular cells (Jacobs et al., 1980). In addition, it was reported that cisplatin competitively inhibited the TEA uptake in the basolateral and brush border membrane vesicles (Williams and Hottendorf, 1985), implying that cisplatin was also transported by the organic cation transport system in the basolateral and brush border membrane. In the present study, cisplatin was transported by hOCT2, hMATE1, and hMATE2-K (Figs. 5A and 8A). In addition, the IC50 of cisplatin against [14C]TEA uptake by hMATE1-expressing cells was lower than by hMATE2-K-expressing cells (Fig. 9, A and B). It was indicated that cisplatin was a good substrate for hOCT2 and hMATE1 but a low-affinity substrate for hMATE2-K. Therefore, it was suggested that the tubular secretion of cisplatin was explained as follows. Basolateral uptake of cisplatin was mediated by hOCT2 and apical secretion was mediated mainly by hMATE1 and slightly by hMATE2-K. Moreover, carboplatin was reported to not be excreted by the tubular secretion (Sorensen et al., 1992). This report corresponded to our results that carboplatin was not transported by organic cation transporters (Figs. 5B and 8, A and B). Oxaliplatin was transported by hOCT2, hMATE1, and hMATE2-K, but not nedaplatin (Figs. 5C and 8A). The inhibitory effects of oxaliplatin on the hMATE1- and hMATE2-K-mediated transport of [14C]TEA were similar (Fig. 9, A and B). These results suggested that oxaliplatin was secreted across the renal epithelial cells via hOCT2, hMATE1, and hMATE2-K, although nedaplatin was not.
In conclusion, we clearly indicated that cisplatin was transported greatly by hOCT2 and weakly by hOCT1 and that oxaliplatin was transported potently by hOCT2 and weakly by hOCT3. Moreover, cisplatin and oxaliplatin were revealed to be preferentially transported by hMATE1 and hMATE2-K, respectively. These basolateral and apical transporters could play a predominant role in the tissue distribution and tubular secretion of platinum agents, being responsible for anticancer effects and/or adverse effects. Therefore, these transporters may be the molecular targets for the prevention from the resistant and adverse effects in platinum-based chemotherapy.
Footnotes
-
This work was supported in part by a Grant-in-Aid for Research on Advanced Medical Technology from the Ministry of Health, Labor, and Welfare of Japan; by a Japan Health Science Foundation “Research on Health Sciences Focusing on Drug Innovation”; by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Culture, and Sports of Japan; and by the 21st Century Center of Excellence program “Knowledge Information Infrastructure for Genome Science.” A.Y. was supported as a Research Assistant by the 21st Century Center of Excellence program “Knowledge Information Infrastructure for Genome Science.”
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.110346.
-
ABBREVIATIONS: r, rat; OCT, organic cation transporter; h, human; OAT, organic anion transporter; OCTN, novel organic cation transporter; MATE, multidrug and toxin extrusion; HEK, human embryonic kidney; TEA, tetraethylammonium; MPP, 1-methyl-4-phenylpyridium; PAH, p-aminohippurate; ES, [6,7-(N)]esterone-3-sulfate; ICP-MS, inductively coupled plasma-mass spectrometry; LDH, lactate dehydrogenase.
- Received July 4, 2006.
- Accepted August 14, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}