


Abstract
Duloxetine, a selective but balanced serotonergic and noradrenergic reuptake inhibitor, was evaluated in the acute nociceptive pain models of tail flick and hot plate in mice and in the persistent and/or inflammatory pain models of acetic acid-induced writhing in mice, carrageenan-induced thermal hyperalgesia and mechanical allodynia in rats, and capsaicin-induced mechanical allodynia in rats. In acute pain models, duloxetine had no significant effect on response latency in the mouse tail-flick test but produced modest increases in response latencies in the mouse hot plate test. Morphine produced dose-related analgesic effects in both the mouse tail-flick and hot plate tests. In models of inflammatory and/or persistent pain, duloxetine, morphine, and ibuprofen produced dose-related decreases in acetic acid-induced writhing in mice. Duloxetine, ibuprofen, and gabapentin also produced dose-dependent reversals of both thermal hyperalgesia and mechanical allodynia produced by carrageenan in rats. In addition, both duloxetine and morphine produced a significant reduction of capsaicin-induced mechanical allodynia in rats. Duloxetine and gabapentin were without substantial effect on the Rotorod test in mice, whereas morphine and ibuprofen produced a significant impairment. Our data indicate that duloxetine may be efficacious in the treatment of persistent and/or inflammatory pain states at doses that have modest or no effect on acute nociception or motor performance.
Descending pain inhibitory circuits contribute to the supraspinal control of spinal transmission of nociceptive information. Descending inhibitory circuits include not only the neuronal connections between the ventrolateral periaqueductal gray and rostral ventral medulla but also the dorsolateral pontine tegmentum, areas that in turn project to the spinal cord dorsal horn lamina (e.g., for review, see Fields and Basbaum, 1994). The rostral ventral medulla is a primary source of serotonergic input to the spinal cord, whereas the dorsolateral pontine tegmentum (particularly the A7 and A5 adrenergic cell groups) is a source of noradrenergic input to the spinal cord. Accordingly, the serotonergic and noradrenergic systems have been implicated in the descending inhibitory control of pain in humans and animals (e.g., Fields et al., 1991). For example, the analgesia produced by the injection of morphine into the periaqueductal gray was antagonized only by a combination of serotonergic and noradrenergic antagonists administered intrathecally (Yaksh, 1979). More recently, antidepressant drugs with serotonergic or noradrenergic reuptake inhibition have been reported to produce varying degrees of pain relief in several persistent, or chronic, pain syndromes in humans, including diabetic neuropathy (Max et al., 1992; Sindrup and Jensen, 1999), postherpetic neuralgia (e.g., Kishore-Kumar et al., 1990; Max et al., 1988), and fibromyalgia (Goldenberg et al., 1996; Rani et al., 1996). In addition, antidepressant drugs that inhibit serotonin and/or norepinephrine reuptake also have been reported to produce modest analgesic efficacy in neuropathic and inflammatory pain models in animals (e.g., Jett et al., 1997; Sawynok and Reid, 2001). However, the effects of simultaneously inhibiting both serotonergic and noradrenergic reuptake in models of acute nociception as well as persistent hyperalgesia and allodynia are not well studied.
Persistent pain states may arise from a number of etiologies, including trauma, disease states, chronic inflammation, amputation, or metabolic disturbances, which alter the processing or transmission of nociceptive and non-nociceptive stimuli in pain pathways. Although the underlying pathophysiology of persistent pain is not yet fully understood, several factors have been demonstrated to be important, including central sensitization as a result of excessive afferent stimulation, loss of inhibitory neurons in the spinal cord, and reorganization of primary afferent projections within the dorsal horn of the spinal cord (Woolf and Doubell, 1994). Central sensitization is characterized by altered responsiveness of dorsal horn and/or thalamic neurons, expansion of receptive fields, and plasticity of neuronal connections (see e.g., Devor and Wall, 1978; Woolf et al., 1994), and manifests as hyperalgesia (an increased sensitivity to noxious stimuli) and/or allodynia (a nocifensive response to previously non-noxious stimuli) (Woolf and Doubell, 1994). Hypofunctioning of the descending serotonergic and/or noradrenergic inhibitory circuits may lead to excessive afferent stimulation and thereby contribute to the development of central sensitization in persistent pain states (e.g., Fields and Basbaum, 1994; Traub, 1997).
Duloxetine is a balanced serotonergic and noradrenergic reuptake inhibitor with efficacy in the treatment of major depressive disorder (e.g., Detke et al., 2002). Duloxetine is approximately equipotent in inhibiting the reuptake of serotonin and norepinephrine as determined by inhibition of ex vivo uptake in rat hypothalamic synaptosomes with ED50 values of 12 ± 1.6 and 14.5 ± 2.3 mg/kg orally in rats, respectively (Wong et al., 1993). Moreover, duloxetine has little or no affinity for serotonin, norepinephrine, acetylcholine, dopamine, histamine, or opioid receptors nor ion channels including sodium channels (Wong et al., 1988, 1993; Fuller et al., 1994; Bymaster et al., 2001). Duloxetine is effective in reversing mechanical allodynia in nerve injury models of neuropathic pain and models of persistent pain in rats (Iyengar et al., 2004). Full efficacy occurs at doses that do not impair performance on the Rotorod test in rats, suggesting that duloxetine is nonsedating. The purpose of the present studies was to evaluate the analgesic efficacy of this balanced serotonergic and noradrenergic reuptake inhibitor in acute nociceptive pain models as well as in inflammatory and persistent pain models in mice and rats. Accordingly, dose-response curves were determined for duloxetine in the mouse tail-flick and hot plate tests of acute nociception as well as in the inflammatory pain tests of acetic acid-induced writhing in mice and carrageenan-induced thermal hyperalgesia and mechanical allodynia in rats, as well as in capsaicin-induced mechanical allodynia in rats. For purposes of comparison, dose-response curves were also determined for morphine in the tail-flick, hot plate, writhing, and capsaicin tests, for ibuprofen in the writhing and carrageenan tests, and for gabapentin in the carrageenan test. The effects of duloxetine on motor performance were also compared with those of morphine, gabapentin, and ibuprofen in the Rotorod test in mice.
Materials and Methods
Subjects. Male CF1 mice (Harlan, Indianapolis, IN) weighing between 18 and 30 g and male Sprague-Dawley rats (Harlan) weighing 70 to 90 g for the carrageenan experiments and 290 to 325 g for the capsaicin experiments were used. Mice were housed in groups of up to 17 per cage and rats in groups of up to 6 per cage in a large colony room on a 12-h light/dark cycle (lights on 6:00 AM), with food and water provided ad libitum. Each animal was used only once. Test sessions were performed between 8:00 AM and 6:00 PM. All treatment or dose groups consisted of 5 to 12 mice or rats. All experiments were conducted in accordance with the National Institutes of Health regulations of animal care covered in “Principles of Laboratory Animal Care,” National Institutes of Health publication 85-23 and were approved by the Institutional Animal Care and Use Committee.
Rotorod Test. The effects of duloxetine and morphine on motor performance were evaluated using a Rotorod. All mice were given two initial training trials of 120 s, approximately 10 min apart, to maintain posture on a Rotorod (model 7650; Ugo Basile, Comerio, Italy), 3 cm in diameter, and rotating at a constant 17 revolutions/min. After the initial training trials, a baseline trial of 120 s was conducted, followed by administration of vehicle, a dose of duloxetine i.p. or p.o., or a dose of morphine s.c., a dose of ibuprofen p.o., or a dose of gabapentin p.o., and 30 min after i.p. or s.c. administration or 60 min after p.o. administration, a 120-s test trial was conducted. The time each animal remained on the Rotorod was recorded; animals not falling off the Rotorod were given a maximum score of 120 s.
Tail-Flick Test in Mice. Groups of five mice had their tails individually immersed in a water bath maintained at 55°C, and the latency to removal was measured. A baseline response latency was determined, followed by administration of vehicle or a dose of drug either s.c. or i.p., and 30 min later, a postinjection latency was obtained. If an animal did not flick its tail within 10 s, it was removed and assigned a response time of 10 s. For each animal, the % maximum possible effect (%MPE) was calculated using the following formula: [(postdrug latency - predrug latency)/(10-s predrug latency)] × 100.
Hot Plate Test in Mice. Groups of five mice were individually placed on a hot plate maintained at 55°C and the latency to lick the front or hind paws was recorded for each mouse. A baseline response latency was determined, vehicle or a dose of drug administered s.c. or i.p., and 30 min later, a postinjection latency was obtained. Animals not responding within 30 s were removed and assigned a score of 30 s. For each animal, %MPE was calculated using the following formula: [(postdrug latency - predrug latency)/(30-s predrug latency)] × 100.
Writhing Test in Mice. Separate groups of 5 or 10 mice were administered vehicle or a dose of drug s.c. or p.o. followed 25 (s.c.) or 55 (p.o.) min later by an i.p. injection of 0.55% acetic acid. Each mouse was then placed in an individual clear plastic observation chamber, and the total number of writhes made by each mouse was counted between 5 and 10 min after acetic acid administration (30–35 s.c. or 60–65 p.o. min after vehicle or dose of drug). For the time course study, a separate group of 10 mice was used for each time point, specifically 2, 4, 8, 18, or 24 h postdrug treatment, under the following test procedure: each group of mice was administered vehicle or a 30 mg/kg dose of duloxetine p.o., then 5 min prior to testing, each mouse was injected i.p. with a 0.55% acetic acid solution, and the number of writhes was counted between 5 and 10 min after acetic acid administration. For both the dose response and time course studies, data are expressed as the mean number of writhes during the 5-min observation period.
Carrageenan-Induced Thermal Hyperalgesia and Mechanical Allodynia. Groups of 6 to 12 rats were injected s.c. with λ-carrageenan (100 μl of a 1.5% solution) into the plantar surface of the right hind paw at time 0 followed 90 min later by a p.o. or i.p. injection of vehicle or a dose of drug. Withdrawal latencies to mechanical and thermal stimuli were determined after approximately an additional 30 and 40 min, respectively. Withdrawal latencies to a nociceptive thermal stimulus were assessed using a modification of the methods of Hargreaves et al. (1988). Each rat was placed in a Plexiglas cubicle with a glass floor through which an infrared photobeam was shown onto the plantar surface of the hind paws and the latency to withdrawal from the thermal stimulus was determined. The intensity of the infrared photobeam from the plantar reflex device (Plantar Test, Ugo Basile) was adjusted to produce a mean response latency in untreated rats of approximately 12 to 15 s. The response latency was determined using a timer linked to the photodiode motion sensors in the plantar reflex device. Response latency was defined as the time from the onset of exposure to the infrared photobeam to the cessation of the photobeam when the photodiode motion sensors detected the withdrawal response of the paw of the rat. Response to the thermal stimulus was reported as the withdrawal latency differences between the treated and untreated paws in seconds and was calculated using the following formula: withdrawal latency of the carrageenan-treated paw - the withdrawal latency of the untreated paw. Mechanical allodynia was evaluated with a calibrated series of von Frey filaments using a modification of the methods of Chaplan et al. (1994). Briefly, rats were placed in clear plastic cages (17.5 × 15 × 15 cm) fitted with wire mesh flooring and allowed to acclimate for approximately 5 min; the withdrawal thresholds were determined by the up and down method (Chaplan et al., 1994) by applying each filament to the midplantar surface of each hind paw in a perpendicular fashion at 5, 10, and 15 mm from the primary injection site and depressed slowly (4–5 s) until bending occurred and the maximum force of the fiber was exerted. Any paw withdrawal response in the 5-, 10-, or 15-mm region outside the primary site of injection was scored as a response to the filament.
Capsaicin-Induced Allodynia. Groups of six rats were injected s.c. or i.p. with vehicle or a dose of drug 15 min before capsaicin (30 μg in 25 μl) was injected into the plantar surface of the right hind paw. Ten minutes after the injection of capsaicin, mechanical allodynia was evaluated with a calibrated series of von Frey filaments using the modification of the up-and-down method of Chaplan et al. (1994) as described above.
Drugs. Duloxetine [(+)-N-methyl-3-(1-naphthalenyloxy)-3-(2-thienyl) propamine HCl] (Eli Lilly & Co., Indianapolis, IN), morphine sulfate (Sigma/RBI, Natick, MA), and λ-carrageenan (Sigma-Aldrich, St. Louis, MO) were dissolved in double deionized water. Ibuprofen (Sigma-Aldrich) was suspended in 5% acacia and double deionized water. Doses refer to the form of the drug listed. All drugs were administered s.c., i.p., or p.o. by gavage in a volume of 1.0 ml/kg in rats and 10 ml/kg in mice. A 0.55% acetic acid solution was prepared by adding 0.55 ml of acetic acid to double deionized water in a final volume of 100 ml (Fisher Scientific Co., Pittsburgh, PA). The injection volume of 0.55% acetic acid was 0.01 ml/g i.p. Capsaicin (Sigma-Aldrich) was prepared as a 3-mg/2.5-ml solution, dissolved in olive oil (Sigma-Aldrich) and sonicated for 25 min in a 45°C water bath.
Statistical Analysis. Data were expressed as means ± S.E.M. ED50 values were determined using a four-parameter logistic equation with the top or bottom fixed (DeLean et al., 1978). The ED50 was defined as the dose that produced 50% of the maximum possible effect (50% MPE). Treatment groups were compared with appropriate control groups using one-way analysis of variance and Dunnett's t test. Statistical analyses were performed using JMP version 4.04 statistical software (SAS Institute, Cary, NC). A probability of p ≤ 0.05 was taken as the level of statistical significance.
Results
Duloxetine had no significant effect on gross motor performance, as measured by the Rotorod test, after administration of 1.0 to 30.0 mg/kg i.p. or p.o. (Table 1). Similarly, gabapentin had no effect on the Rotorod over the dose range of 3 to 100 mg/kg p.o. (Table 1). In contrast, morphine decreased motor performance on the Rotorod over the dose range of 1.0 to 30.0 mg/kg when administered s.c., with a significant decrease after the 10 mg/kg dose (Table 1). Ibuprofen was without effect on the Rotorod over the dose range of 30 to 300 mg/kg p.o.; however, a dose of 1000 mg/kg p.o. produced an impairment (Table 1).
Lack of effects of duloxetine on motor performance as measured on the Rotorod Mice were administered the indicated dose of duloxetine i.p. or p.o., morphine s.c., ibuprofen p.o., or gabapentin p.o., then tested on the Rotorod for 30 min (following i.p. or s.c. administration) or 60 min (following p.o. administration) later. Each value represents the mean ± S.E.M. of 10 mice.
In the 55°C tail-flick test, duloxetine was without significant effect over the dose range of 1 to 30 mg/kg i.p. (Fig. 1). In comparison, morphine (1.0–30 mg/kg, s.c.) produced significant, dose-related antinociceptive effects at 3, 10, and 30 mg/kg (Fig. 1). The ED50 (±95% CL) value for morphine was 3.2 (2.5–4.4) mg/kg.

Duloxetine (i.p.) had no effect in mice in the tail-flick test using a 55°C water bath, in comparison with the dose-related antinociception produced by morphine (s.c.). Each point represents the mean of five mice, and each mouse was used only once. The predrug baseline latencies were 2.50 ± 0.11 and 2.24 ± 0.16 s in the duloxetine and morphine groups, respectively. Points above Veh represent the effects of vehicle alone. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, dose of drug in milligrams per kilogram. Ordinate, %MPE (see Materials and Methods). *, p < 0.05 versus Veh, Dunnett's t test.
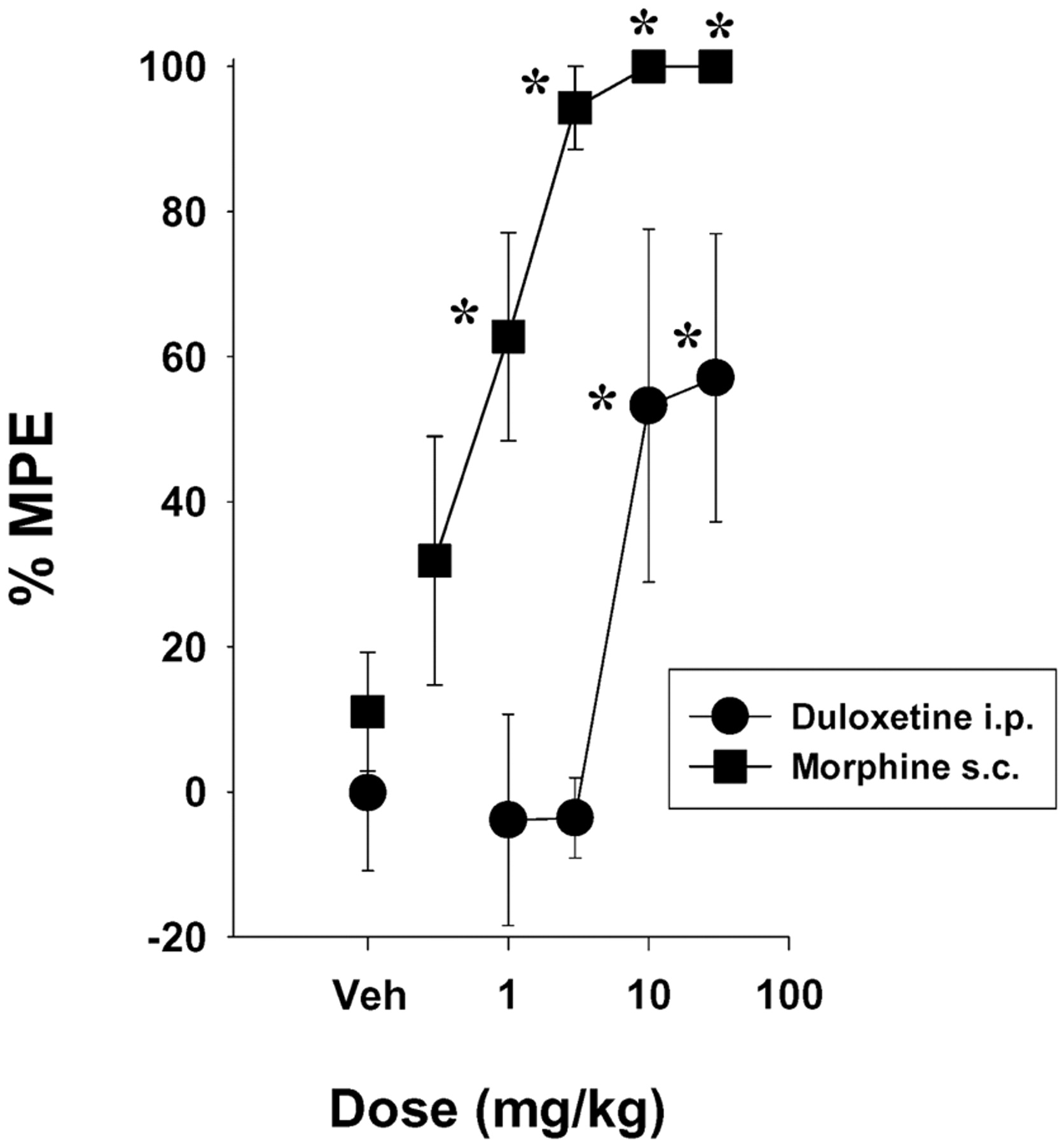
In the 55°C hot plate test, duloxetine increased response latencies to approximately 55 to 60% MPE after doses of 10 and 30 mg/kg i.p., whereas morphine produced significant increases in response latency after 1 mg/kg s.c. (Fig. 2) and approximately 100% MPE after doses of 3, 10, and 30 mg/kg s.c. (Fig. 2). The ED50 (±95% CL) values were 16.4 (7.2–37.7) mg/kg for duloxetine and 0.58 (0.31–2.1) mg/kg for morphine.
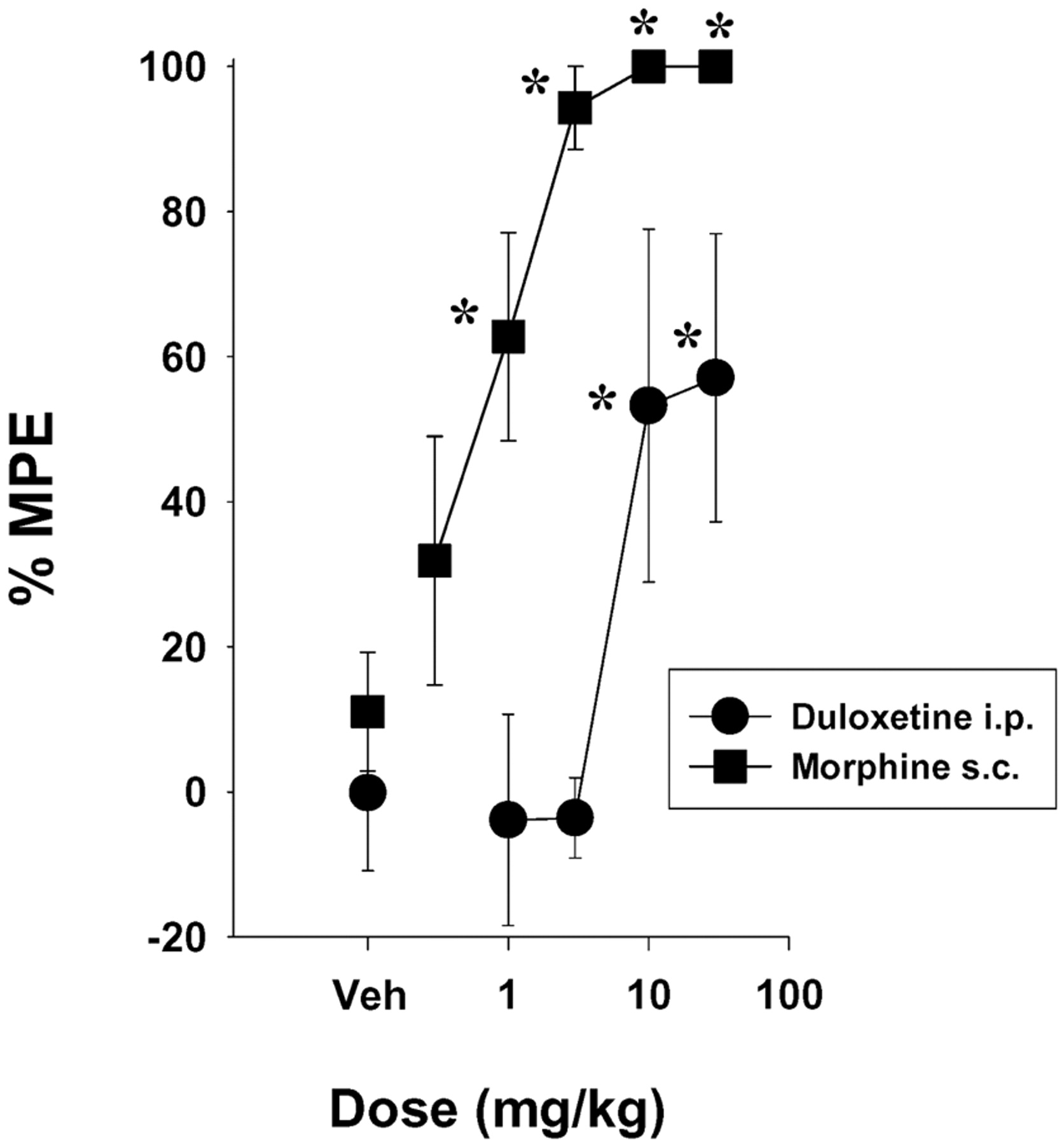
Dose-related antinociceptive effects of duloxetine (i.p.) and morphine (s.c.) in mice in the 55°C hot plate test. Each point represents the mean of five mice, and each mouse was used only once. The predrug baseline latencies were 9.7 ± 0.6 and 9.1 ± 0.6 s in the duloxetine and morphine groups, respectively. Points above Veh represent the effects of vehicle alone. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, dose of drug in milligrams per kilogram. Ordinate, %MPE. *, p < 0.05 versus Veh, Dunnett's t test.
The dose- and time-related effects of duloxetine were evaluated in the acetic acid-induced writhing test in mice, a visceral inflammatory pain test. Mice treated with an injection of 0.55% acetic acid solution alone exhibited an average of approximately 15 writhes over a 5-min interval (see points above Veh in Fig. 3). Duloxetine p.o. produced a significant, dose-related inhibition of acetic acid-induced writhing in mice (Fig. 3) and with a duration of action of approximately 8 to 18 h (Fig. 4). Morphine and ibuprofen also both produced dose-related reductions in acetic acid-induced writhing (Fig. 3). The rank order of potencies and ED50 (95% CL) values in milligrams per kilogram of the compounds in reversing acetic acid-induced writhing were morphine [0.31 (0.17–0.59)] > duloxetine [11.8 (6.4–22.8)] > ibuprofen [95 (44–190)].

Dose-related inhibition of acetic acid-induced writhing by duloxetine (p.o.), morphine (s.c.), and ibuprofen (p.o.) in mice. Points above Veh represent the effects of vehicle alone. Each point represents the mean of 10 or 5 (morphine) mice, and each mouse was used only once. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, dose of drug in milligrams per kilogram. Ordinate, number of writhes in 5 min. *, p < 0.05 versus Veh, Dunnett's t test.

Time-related effects of duloxetine (30 mg/kg) or vehicle administered orally in preventing acetic acid-induced writhing in mice. Each point represents the mean of 10 mice, and each mouse was used only once. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, time in hours postduloxetine (30 mg/kg p.o.). Ordinate, number of writhes in 5 min. *, p < 0.05 versus Veh group at the same time point, Student's t test.
The effects of duloxetine were further evaluated in reversing carrageenan-induced thermal hyperalgesia and mechanical allodynia, a model of persistent inflammatory pain. When administered i.p. 90 min after carrageenan, duloxetine produced a dose-related reversal of carrageenan-induced thermal hyperalgesia, which was significant at the 10 and 30 mg/kg doses, and mechanical allodynia, also significant at the 10 and 30 mg/kg doses (Fig. 5, left and right, respectively). In comparison, ibuprofen and gabapentin also reversed carrageenan-induced thermal hyperalgesia, and these effects were significant after doses of 10 to 100 mg/kg p.o. for gabapentin and 1000 mg/kg p.o. ibuprofen (Fig. 5, left). Ibuprofen and gabapentin also reversed carrageenan-induced mechanical allodynia, with significant reversals after doses of 10 to 100 mg/kg p.o. for gabapentin and doses of 300 and 1000 mg/kg p.o. for ibuprofen (Fig. 5, right). The rank order of potencies and ED50 values in milligrams per kilogram of the compounds in reversing carrageenan-induced thermal hyperalgesia (Fig. 5, left) were duloxetine [4.6 (1.8–10.7)] ≥ gabapentin [4.4 (3.0–6.4)] > ibuprofen [300 (300–300)]. In comparison, the rank order of potencies and ED50 values in milligrams per kilogram of the compounds tested in reversing carrageenan-induced mechanical allodynia (Fig. 5, right) were duloxetine [10.7 (9.1–12.6)] > gabapentin [29.3 (21.6–39.7)] > ibuprofen [121 (58–791].
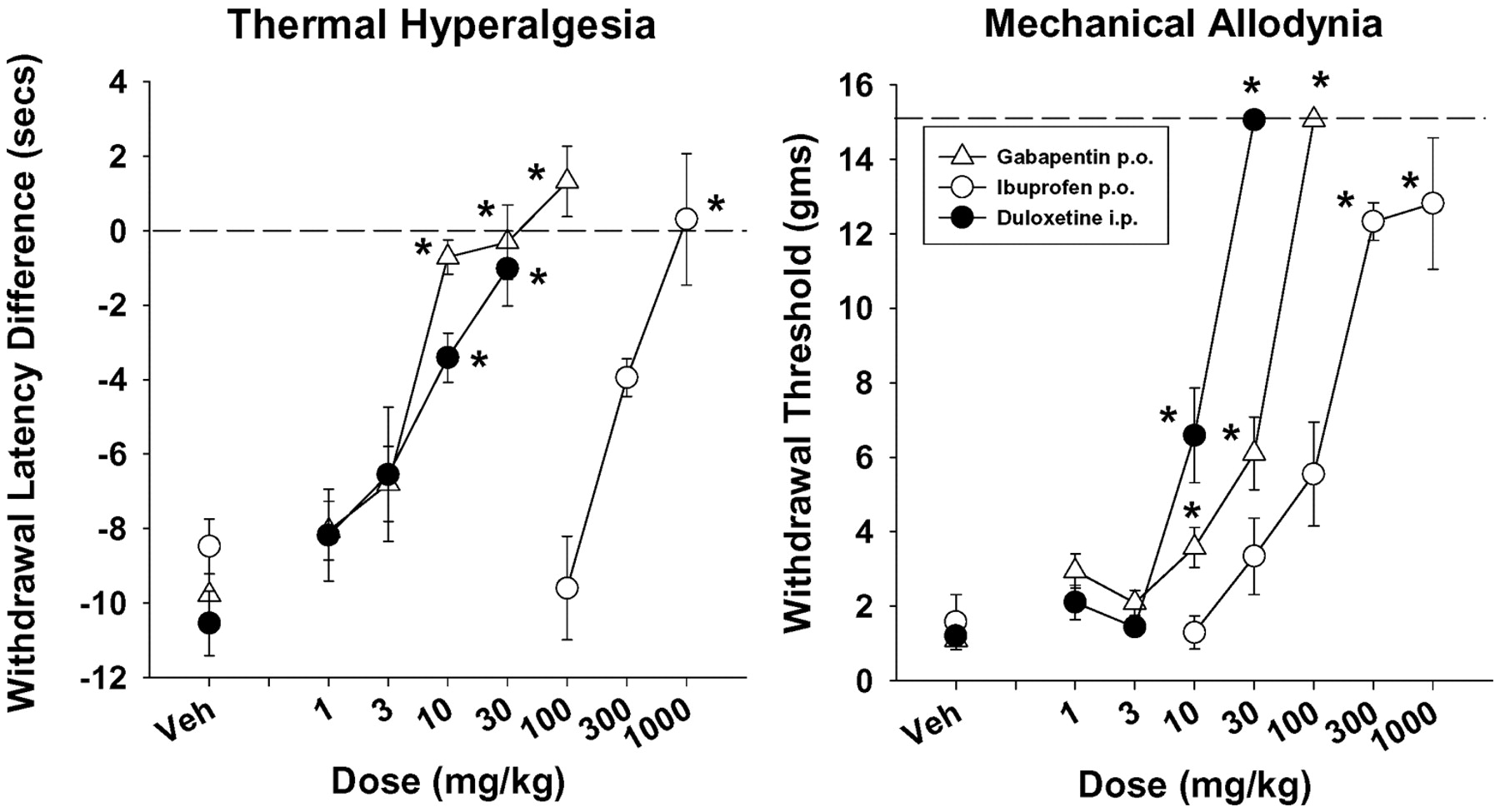
Dose-related effects of duloxetine (i.p.), gabapentin (p.o.), and ibuprofen (p.o.) in reversing carrageenan-induced thermal (left) and mechanical (right) hyperalgesia in rats. Each point represents the mean of 6 (morphine) or 12 rats, and each rat was used only once. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, dose of drug in milligrams per kilogram. Ordinate, left, withdrawal latency difference in seconds to a thermal stimulus; right, withdrawal threshold in grams to a mechanical stimulus. *, p < 0.05 versus Veh, Dunnett's t test.
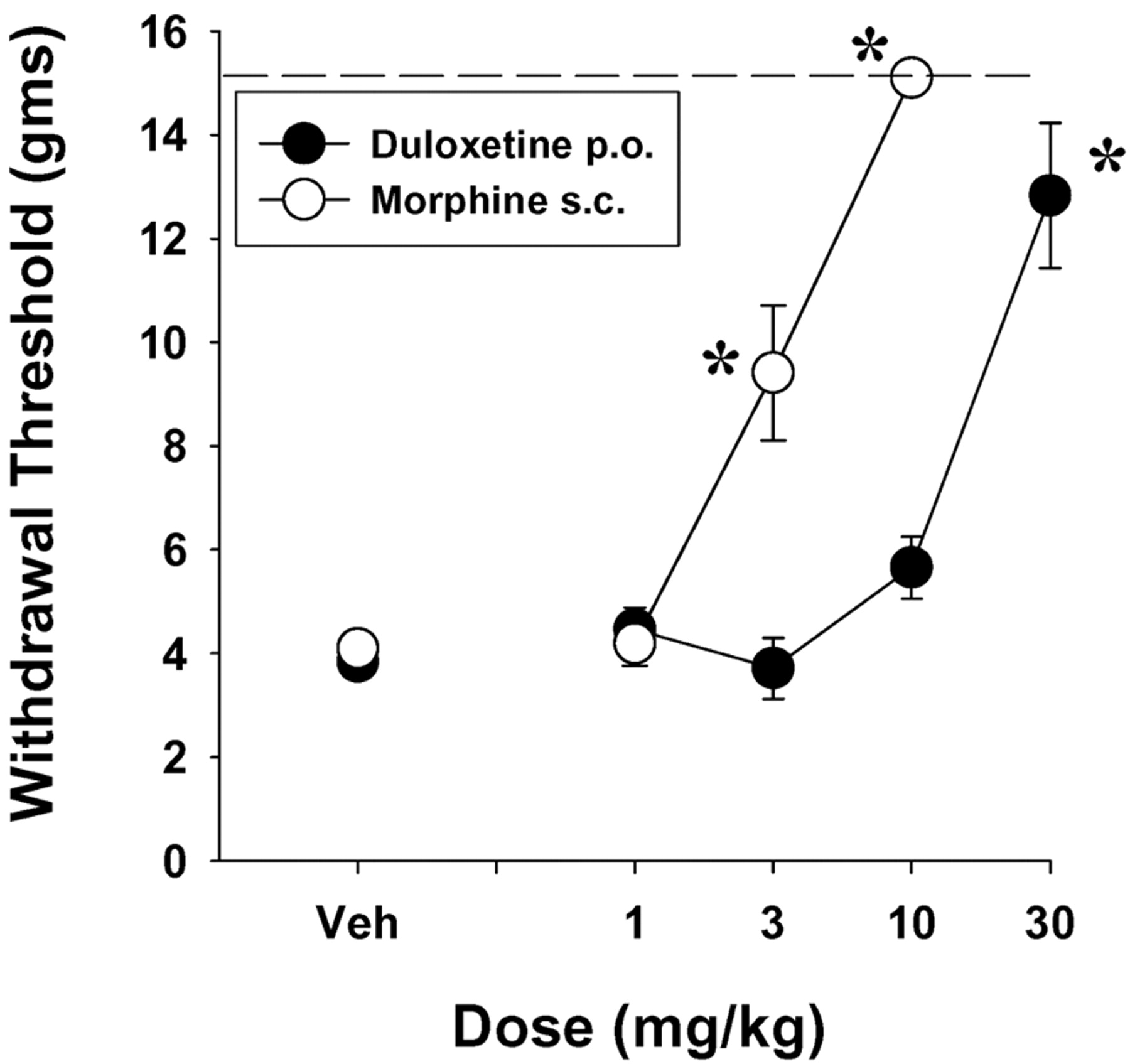
The effects of duloxetine were also evaluated on capsaicin-induced mechanical allodynia in rats. Duloxetine, administered 15 min before capsaicin, produced a significant prevention of capsaicin-induced mechanical allodynia (Fig. 6) that was statistically significant after a dose of 30 mg/kg p.o. and had an ED50 (95% CL mg/kg) value of 9.2 (3.1–27.2). Morphine also produced a significant dose-dependent prevention of capsaicin-induced mechanical allodynia that was significant after doses of 3 and 10 mg/kg s.c. and with an ED50 (95% CL mg/kg) value of 1.9 (1.3–2.9) (Fig. 6).

Dose-related effects of duloxetine (i.p.) and morphine (s.c.) in reversing capsaicin-induced mechanical hyperalgesia in rats. Each point represents the mean of six rats, and each rat was used only once. Vertical lines represent ±S.E.M. and are absent when less than the size of the point. Abscissa, dose of drug in milligrams per kilogram. Ordinate, withdrawal threshold in grams to a mechanical stimulus. *, p < 0.05 versus Veh, Dunnett's t test.
Discussion
In the present studies, duloxetine, a balanced serotonergic and noradrenergic reuptake inhibitor, was efficacious in reversing and/or preventing not only acetic acid-induced writhing but also carrageenan- and capsaicin-induced hyperalgesia and allodynia at doses that had limited effects on acute nociception in the tail-flick and hot plate tests and little impairment of normal motor function as assessed on the Rotorod. Moreover, duloxetine was approximately equiefficacious to morphine, gabapentin, and ibuprofen in the carrageenan and/or capsaicin tests and was generally more potent than either gabapentin or ibuprofen. Taken together, the present findings suggest that duloxetine, a balanced serotonergic and noradrenergic reuptake inhibitor, may be clinically efficacious in the treatment of inflammatory and persistent pain syndromes in humans.
Nociceptive information is processed and integrated peripherally as well as at spinal and supraspinal levels within the central nervous system. In the present study, duloxetine had no effect in the mouse tail-flick test, a spinally integrated nociceptive reflex, and only modest effects at the highest doses tested in the mouse hot plate test, a complex response that is supraspinally integrated. Thus, the present data suggest that duloxetine was without effect on the processing of acute noxious stimuli at the level of the spinal cord and had only modest effect at the supraspinal level in mice. That duloxetine might have peripheral effects on the modulation of acute nociceptive stimuli seems unlikely but could not be determined from the present results. Previous studies have reported that direct acting serotonergic and noradrenergic receptor agonists such as MK212 and quipazine (e.g., Yaksh and Wilson, 1979) and the α2-adrenergic receptor agonists clonidine and oxymetazoline (e.g., Yaksh, 1979; Womer and Shannon, 2000) produced dose-related acute antinociceptive effects in the tail-flick test when given intrathecally or systemically. Taken together, the data suggest that duloxetine may not increase extracellular serotonin and norepinephrine sufficiently to stimulate the relevant serotonergic and/or noradrenergic receptors that modulate acute nociception, particularly at the spinal cord level.
The efficacy of duloxetine was also evaluated in acetic acid-induced writhing in mice, which may be considered a model of visceral inflammatory pain, and carrageenan-induced thermal hyperalgesia and mechanical allodynia in rats, additional models of inflammation-induced central sensitization and pain. Both acetic acid-induced writhing and carrageenan-induced hyperalgesia and allodynia are readily reversed by nonsteroidal anti-inflammatory drugs such as ibuprofen as well as by opioid agonists such as morphine. In the present studies, duloxetine inhibited acetic acid-induced writhing in mice and reversed carrageenan-induced thermal hyperalgesia and mechanical allodynia in rats. Thus, duloxetine was efficacious in reducing inflammation-related pain and central sensitization in the present studies. Moreover, duloxetine was equiefficacious to both the nonsteroidal anti-inflammatory drug ibuprofen and the opioid receptor agonist morphine in the writhing and carrageenan models, as well as equiefficacious to gabapentin in the carrageenan model. Our findings are in agreement with those of Singh et al. (2001) who showed that the selective serotonergic reuptake inhibitor fluoxetine produced a dose-dependent antinociceptive effect in the acetic acid-induced writhing test in mice. In the rat carrageenan model, our findings are also consistent with previous reports that the selective serotonin reuptake inhibitors clomipramine and fluoxetine as well as the noradrenergic reuptake inhibitor amitriptyline produced at least modest antihyperalgesia (Bianchi et al., 1995; Eisenach and Gebhart, 1995). Previous studies suggest that chronic inflammatory pain directly suppresses descending serotonergic and noradrenergic pathways (Traub, 1997). Thus, one possible mechanism for the efficacy of duloxetine in the acetic acid and carrageenan models may be through an enhancement of serotonergic and noradrenergic descending inhibitory pathways that modulate central sensitization. Further studies are needed to more clearly establish the role of serotonergic and noradrenergic systems in the prevention and reversal produced by duloxetine of hyperalgesia and central sensitization.
Another model of central sensitization involves the selective stimulation of C-fiber primary afferents through the subcutaneous administration of capsaicin. Capsaicin selectively activates vanilloid-1 receptors located on C-fiber primary neurons (Davis et al., 2000). The afferent barrage produced by capsaicin produces sensitization of spinothalamic tract neurons (Simone et al., 1991; Dougherty and Willis, 1992) resulting in thermal hyperalgesia and mechanical allodynia in animals (Gilchrist et al., 1996) as well as humans (LaMotte et al., 1991; Torebjörk et al., 1992). In the present study, pretreatment with duloxetine produced a dose-related inhibition of capsaicin-induced mechanical allodynia, and duloxetine was equiefficacious to morphine, although slightly less potent. The antihyperalgesic effects of duloxetine in preventing capsaicin-induced allodynia represent the first demonstration that a dual serotonergic and noradrenergic reuptake blocker can reduce the C-fiber-mediated hyperalgesia such as that produced by capsaicin. Clinical trials using the tricyclic antidepressants desipramine (Wallace et al., 2002) and amitriptyline (Eisenach et al., 1997) reported no significant effects on capsaicin-induced pain, flare response, or secondary hyperalgesia. However, desipramine and amitriptyline are relatively selective norepinephrine reuptake inhibitors and also have dose-limiting anticholinergic side effects (e.g., Max, 1995). Together with the present results, the lack of effects of desipramine and amitriptyline on capsaicin-induced hyperalgesia and allodynia in humans suggests that inhibition of reuptake of both serotonin and norepinephrine may be required for efficacy in preventing hyperalgesia and allodynia produced by capsaicin-induced stimulation of C-fibers. In addition, the possibility that the anticholinergic effects of desipramine and amitriptyline may have limited the efficacy or the dose that could be administered cannot be ruled out. Thus, further preclinical and clinical research is needed to determine the role of the serotonergic and noradrenergic systems in the mechanism of action of duloxetine in preventing capsaicin-induced hyperalgesia and allodynia.
Although antidepressant drugs with serotonergic, noradrenergic, or mixed reuptake inhibition are effective in the treatment of chronic pain syndromes in humans (for review, see Onghena and Van Houdenhove, 1992; Lynch, 2001), the use of most tricyclic antidepressants is often limited by numerous side effects including sedation, orthostatic hypotension, and cardiac conduction abnormalities, in part due to the affinity of these drugs for muscarinic cholinergic, α-1 adrenergic, and other receptors (e.g., Max, 1995). In a recent clinical trial (Detke et al., 2002), duloxetine was well tolerated in patients with major depressive disorder and was without evidence of anticholinergic effects, consistent with the dual reuptake inhibition and lack of affinity for acetylcholine, norepinephrine, histamine, dopamine, and serotonin receptors demonstrated in preclinical studies (Wong et al., 1988, 1993; Fuller et al., 1994). Furthermore, initial support for the potential importance of both the serotonergic and noradrenergic systems working in concert is provided by Goldenberg et al. (1996), who reported that the combination of fluoxetine and amitriptyline was more effective in relieving fibromyalgia pain than was either drug alone. Taken together, the preclinical and clinical data suggest that the balanced inhibition of serotonergic and noradrenergic reuptake and lack of anticholinergic effects of duloxetine may provide superior clinical efficacy and safety in the treatment of chronic/persistent pain syndromes.
As noted above, one possible explanation for the efficacy of duloxetine in reversing and/or preventing persistent pain states in rodents is that duloxetine may indirectly enhance the tone of descending serotonergic and noradrenergic inhibitory projections from the midbrain to the dorsal horn of the spinal cord, thereby decreasing transmission of nociceptive information in the dorsal horn and potentially supraspinally as well. Thus, in persistent pain states, serotonergic and noradrenergic mechanisms involved in descending inhibition may be compromised or hypofunctional. In this context, duloxetine may increase serotonergic and noradrenergic neuro-transmission in descending inhibitory pathways sufficiently to dampen processing of stimuli in pain pathways, leading to substantial pain relief.
Finally, it may be noteworthy that the same serotonergic and noradrenergic nuclei implicated in descending inhibition of pain pathways also send ascending projections to areas of the brain that are implicated in depression because there is a high degree of comorbidity of pain and depression (e.g., Max, 1995). It is tempting to speculate that if these nuclei are hypofunctional, then the hypofunctionality of ascending projections of these nuclei to higher brain regions may manifest as depression, whereas the hypofunctionality of descending projections of these nuclei to the spinal cord may manifest as persistent pain, thus potentially explaining the high comorbidity of these two disorders. In addition, enhancement of ascending serotonergic and noradrenergic pathways by duloxetine may provide pain relief by reducing negative affective components of clinical pain states.
In summary, duloxetine, a balanced serotonergic and noradrenergic reuptake inhibitor, produced antihyperalgesic and antiallodynic efficacy in several rodent models of visceral, inflammatory, and C-fiber-mediated pain at doses that had little effect on acute nociception or gross motor performance. Moreover, in these models, duloxetine was equiefficacious to morphine, gabapentin, and ibuprofen, examples of three classes of drugs currently used in the treatment of persistent pain states. Thus, the present data indicate that duloxetine may also be clinically efficacious in the treatment of persistent pain.
Acknowledgments
We thank Brian Eastwood, who provided expert statistical assistance and advice.
Footnotes
- Received August 10, 2004.
- Accepted October 14, 2004.
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
doi:10.1124/jpet.104.075960.
ABBREVIATIONS: MPE, maximum possible effect; CL, confidence limits.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}