


Abstract
Many of the biological actions of progesterone are mediated through the progesterone receptor (PR), a nuclear transcription factor. Progesterone is well recognized to protect against seizures in animal models. Although this activity has been attributed to the progesterone metabolite allopregnanolone, a GABAA receptor-modulating neurosteroid with anticonvulsant properties, PRs could also play a role. Here, we used PR knockout (PRKO-/-) mice bearing a targeted deletion of the PR gene that eliminates both isoforms of the PR to investigate the contribution of the PR to the anticonvulsant activity of progesterone. The protective activity of progesterone was examined in female and male homozygous PRKO mice and isogenic wild-type controls in the pentylenetetrazol (PTZ), maximal electroshock, and amygdala-kindling seizure models. In all three models, the anticonvulsant potency of progesterone was undiminished in PRKO mice compared with control mice. On the contrary, there was a substantial increase in the anticonvulsant potency of progesterone in the PTZ and kindling models. The antiseizure activity of progesterone in PRKO mice was reversed by pretreatment with finasteride, a 5α-reductase inhibitor that blocks the metabolism of progesterone to allopregnanolone. Unlike progesterone, the neurosteroids allopregnanolone and allotetrahydrodeoxycorticosterone exhibited comparable anticonvulsant potency in PRKO and wild-type mice. The basis for the heightened progesterone responsiveness of PRKO mice is not attributable to pharmacokinetic factors, because the plasma allopregnanolone levels achieved after progesterone administration were not greater in the PRKO mice. These studies provide strong evidence that the PR is not required for the antiseizure effects of progesterone, which mainly occurs through its conversion to the neurosteroid allopregnanolone.
The progesterone receptor (PR), a member of the nuclear receptor superfamily of transcription factors, mediates many of the physiological effects of the sex steroid progesterone (Tsai and O'Malley, 1994). In progesterone-responsive target cells, progesterone binds to cytoplasmic PRs and the hormone-nuclear receptor complexes translocate to the cell nucleus where they activate or silence the transcription of downstream gene networks, thus effecting the physiological response of the target cell. The PR exists in two isoforms, PR-A and PR-B, that are transcribed from the same gene and are structurally identical except for a short amino acid extension located in the N-terminal region of PR-B (Mulac-Jericevic et al., 2000). The central nervous system is an important target for progesterone. The steroid acts in discrete regions of the female brain to regulate sexual receptivity, ovulation, pregnancy, and other aspects of reproduction through classical genomic actions mediated by PRs (Clarke and Sutherland, 1990; Lydon et al., 1995; Mani et al., 1997; Frye and Vongher, 2001). In addition, progesterone can indirectly affect central nervous system function independently of the PR through its reduced metabolite allopregnanolone (5α-pregnan-3α-ol-20-one), which acts as a potent positive modulator of GABAA receptors (Engel and Grant, 2001; Frye, 2001; Mellon et al., 2001; Lambert et al., 2003). Binding of allopregnanolone to synaptic and extrasynaptic GABAA receptors augments inhibitory chloride conductance, resulting in decreased neuronal excitability (Lambert et al., 2003; Stell et al., 2003). Among the diverse consequences of the enhancement of neuronal inhibition produced by allopregnanolone is protection against seizures in various animal models of epilepsy (Belelli et al., 1989; Kokate et al., 1996; Frye and Scalise, 2000; Reddy and Kulkarni, 2000; Reddy and Rogawski, 2001; Rogawski and Reddy, 2003).
Protection against epileptic seizures was one of the first identified central actions of exogenously administered progesterone (Selye, 1942; Craig, 1966), and it is believed that fluctuations in endogenous progesterone levels can affect seizure susceptibility in women with epilepsy (Bäcktröm et al., 1984; Herzog, 1999; Rogawski and Reddy, 2004). In recent years, numerous studies have confirmed the powerful anticonvulsant activity of progesterone in diverse animal seizure models (Landgren et al., 1978; Holmes and Weber, 1984; Reddy and Kulkarni, 1997; Mohammad et al., 1998; Kokate et al., 1999; Frye and Scalise, 2000). Because 5α-reductase isoenzymes catalyze the rate-limiting step in the conversion of progesterone to allopregnanolone, it has been possible to use 5α-reductase inhibitors to examine whether the anticonvulsant activity of progesterone is dependent upon conversion to reduced metabolites (Frye et al., 1998; Kokate et al., 1999). These studies, along with additional experiments using mice that lack the type I 5α-reductase isoenzyme (Frye et al., 2002), support the concept that 5α-reduced metabolites of progesterone, particularly allopregnanolone, are responsible for the seizure protection conferred by the parent hormone.
Nevertheless, the extent to which PRs could contribute to the anticonvulsant activity of progesterone has not been fully explored. Although PRs are expressed at high levels in the hypothalamus where they are believed to participate in the regulation of reproductive function, PRs are also found in the neocortex, hippocampus, amygdala, and limbic areas (Parsons et al., 1982; Hagihara et al., 1992; Kato et al., 1994; Alves et al., 2000). These extrahypothalamic brain regions are well recognized to participate in seizure generation and spread, and it is conceivable that cells expressing PRs in these regions could contribute to alterations in seizure susceptibility produced by progesterone. In fact, some authors have suggested that the effects of progesterone on seizure susceptibility involve interactions with PRs (Rupprecht et al., 1993; Edwards et al., 2001a).
In the present study, we sought to establish the role of the PR in the antiseizure actions of progesterone using PR knockout (PRKO) mice bearing a targeted null mutation of the PR gene that abrogates the function of both PR-A and PR-B subtypes (Lydon et al., 1995). Male and female embryos homozygous for the PR mutation develop normally to adulthood, except for some reproductive abnormalities (Lydon et al., 1995; Chappell et al., 1997, 1999). We evaluated the protective activity of progesterone in the pentylenetetrazol (PTZ), maximal electroshock (MES), and amygdala kindling seizure models in PRKO mice. Our results indicate that the PR is not required for the protective activity of progesterone in these models. In fact, we unexpectedly found that progesterone has enhanced antiseizure potency in both male and female mice that lack PRs.
Materials and Methods
Animals. Adult PRKO mice weighing 25 to 30 g were used in the study. The development of the PRKO mouse strain has been described previously (Lydon et al., 1995). A breeding colony was established at the North Carolina State University College of Veterinary Medicine and National Institute of Neurological Disorders and Stroke Veterinary Resource Program. The strain was maintained on a C57BL6/129SvEv hybrid background. Mice were housed separately four to a cage with free access to food and water. The animals were cared for in strict compliance with the guidelines outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Heterozygous PRKO+/- and homozygous PRKO-/- pups were obtained from crosses between heterozygous PRKO+/- females and homozygous PRKO-/- males. For PR genotyping, genomic DNA was isolated from tails and analyzed by polymerase chain reaction amplification (Lydon et al., 1995). All animal procedures were approved by the North Carolina State University and National Institute of Neurological Disorders and Stroke Animal Care and Use committees.
PKRO mice lack PR binding activity for the PR agonist promegestone (R5020) in uterine cytosolic extracts (Lydon et al., 1995), indicating that variant forms of the receptor with ligand-binding capacity are not expressed as has been found to occur in estrogen receptor knockout mice that lack functional estrogen receptor-α (Couse et al., 1995). In binding assays, both wild-type (WT) and heterozygous animals exhibited saturable R5020 binding (Lydon et al., 1995). Heterozygotes had approximately 50% of the maximal binding observed in wild-type mice. Our immunohistochemical studies have confirmed that PRKO mice do not express PR in brain (unpublished observations). Homozygous PRKO mice exhibit reproductive abnormalities, including lack of ovulation, estrus cyclicity, and gonadotropin in surges, but there were no significant differences in plasma progesterone or estrogen levels compared with wild-type animals (Chappel et al., 1997).
PTZ Seizure Test. The effects of progesterone and neurosteroids on seizure susceptibility were evaluated using the s.c. PTZ test (White et al., 1995). Thirty minutes after administration of various doses of progesterone (10–150 mg/kg i.p.) or 15 min after allopregnanolone (1–50 mg/kg s.c.), animals received a s.c. injection of PTZ (85 mg/kg) and were then observed for a 30-min period for clonic seizures. Animals that showed clonic seizures lasting longer than 5 s were scored positive for seizure occurrence. The PTZ can induce both clonic and tonic-clonic seizures. The forelimb clonus originates from forebrain structures, whereas tonic-clonic seizures, including tonic hindlimb extension (THE) are thought to originate from brainstem structures. Thus, we also evaluated the protective effect of progesterone against PTZ-induced THE in male and female PRKO mice. The incidence of THE was recorded in each group. Mice were euthanized within 1 min after meeting the criteria for seizures. If no clonic seizures and THE occurred during this time, animals were considered protected from seizures and promptly euthanized.
Amygdala Kindling. Electrode implantation and stimulation procedures for mouse amygdala kindling were performed as described previously (Rogawski et al., 2001; Reddy and Rogawski, 2002). Briefly, mice were anesthetized by intraperitoneal injection of a mixture of ketamine (100 mg/kg) and xylazine (10 mg/kg). A twisted bipolar stainless steel wire electrode (model MS303/1; Plastics One, Roanoke, VA) was stereotaxically implanted in the right amygdala complex (1.3 mm posterior and 3.0 mm lateral to bregma, and 4.6 mm below the dorsal surface of the skull) (Franklin and Paxinos, 1997) and anchored with dental acrylic to three jeweler's screws placed in the skull. A period of 7 to 10 days was allowed for recovery. The stimulation paradigm consisted of 1-ms duration, bipolar, square current pulses delivered at 60 Hz for 1 s using a kindling stimulator (custom made at National Institutes of Health Research Services Branch, Bethesda, MD). The afterdischarge threshold was determined by stimulating at 5-min intervals beginning with an intensity of 75 μA and increasing in steps of 50 μA until an afterdischarge of at least 5 s was obtained. Stimulation on subsequent days used a stimulation intensity 125% of the threshold value. Seizure activity after each stimulation was rated according to the criterion of Racine (1972) as modified for the mouse: stage 0, no response or behavior arrest; stage 1, chewing or head nodding; stage 2, chewing and head nodding; stage 3, forelimb clonus; stage 4, bilateral forelimb clonus and rearing; and stage 5, falling. The after-discharge was recorded from the amygdala electrode with a Grass CP511 AC electroencephalogram preamplifier (Astro-Med, West Warwick, RI) and stored in digital form using Axotape 2.02 (Axon Instruments, Foster City, CA). Kindling stimulation was delivered daily until stage 5 seizures were elicited on three consecutive days. Significant differences were observed in the rate of kindling development between female wild-type and PRKO mice (Reddy et al., 2001a). The PRKO mice exhibited delayed kindling development, but there was no significant difference in the expression of kindled seizures in fully kindled wild-type and PRKO mice. Thus, we chose fully kindled mice as models for progesterone testing. To examine the ability of progesterone to suppress the expression of kindled seizures, the kindled mice underwent a 3-day test protocol. On the 1st day they were verified to exhibit a stimulation-induced stage 5 seizure. They were then tested on day 2 after having received an i.p. progesterone injection 30 min before stimulation. The animals received control stimulations on day 3. The two control seizure scores before and after drug administration were averaged. At the end of the study, mice were anesthetized and perfused transcardially with paraformaldehyde for Nissl staining to verify the electrode placement.
Estimation of Allopregnanolone. Mice were anesthetized with an injection of ketamine (100 mg/kg)-xylazine (10 mg/kg) solution, and ∼1 ml carotid blood was collected in heparinized tubes. The plasma was separated by centrifugation at 12,000g for 10 min and stored at -20°C in 10-ml glass tubes containing 7.5% EDTA solution (68 μl). The concentration of allopregnanolone was quantified by liquid chromatography-mass spectrometry using a Hewlett Packard (Palo Alto, CA) liquid chromatograph (analytical column, Genesis C18, 4 μm, 3 × 30 mm; Jones Chromatography, Lakewood, CO) and Micromass Quattro II mass spectrometer (Reddy and Rogawski, 2000, 2002). Briefly, a 200-μl plasma sample was added to a tube containing evaporated internal standard (5β,3α-pregnane-21-ol-20-one). The steroid and internal standard were extracted with 4 ml of hexane. Each sample was analyzed using the atmospheric pressure chemical ionization technique under acidic conditions. A standard curve was plotted using pure allopregnanolone in methanol mixed with 0.2 ml of blank plasma. The detection limit of the assay was <0.5 ng/ml.
MES Seizure Test. Thirty minutes after intraperitoneal injection of progesterone, mice were subjected to a 50-mA, 0.2-s, 60-Hz electrical stimulus via corneal electrodes (5-mm-diameter stainless steel balls) wetted with normal saline using an MES stimulator (custom made at National Institutes of Health Research Services Branch, Bethesda, MD). Animals failing to show tonic hindlimb extension were scored as protected.
Sedation/Motor Impairment Test. Evaluation for sedation/motor impairment was carried out using an inverted screen test that determines an animal's ability to support its own body weight by grasping a grid (Coughenour et al., 1977). Thirty minutes after progesterone injection, mice were placed on a horizontally oriented grid (consisting of parallel 1.5-mm-diameter rods situated 1 cm apart), and the grid was inverted. Animals that fell from the grid within 1 min were scored as positive for sedation/motor impairment. Vehicle-treated mice never fell from the grid.
Drugs. Stock solutions of progesterone, finasteride, and other steroids for injection were made in 30% hydroxypropyl-β-cyclodextrin (β-cyclodextrin) in water, and additional dilutions were made using normal saline. By itself, β-cyclodextrin at concentrations as high as 50% failed to affect PTZ or kindled seizures. PTZ was dissolved in normal saline. Drug solutions were administered s.c. or i.p. in a volume equaling 1% of the animal's body weight. Finasteride was procured from Steraloids (Newport, RI). All other drugs were obtained from Sigma-Aldrich (St. Louis, MO). The doses of progesterone in the PTZ and MES models were based on our previously published study (Kokate et al., 1999). The doses of progesterone in the kindling model were based on a dose-response curve generated in a preliminary study in fully kindled mice. The doses chosen were comparable to those previously found to be effective in the kindling model in rats (Mohammad et al., 1998; Lonsdale and Burnham, 2003).
Data Analysis. ED50 values (the doses at which 50% of tested animals were protected from seizures or scored positive in the motor impairment test) with 95% confidence limits were determined by log-probit analysis using the Litchfield and Wilcoxon procedure. The significance of differences between quantal dose-response curves was assessed with the Litchfield and Wilcoxon χ2 test. Kindled seizure severity in groups of mice is expressed as the mean ± S.E.M. of the seizure stage score; differences between groups were compared with the nonparametric Kruskal-Wallis test followed by the Mann-Whitney U test. The mean afterdischarge duration values were compared with Dunnett's t test. Plasma allopregnanolone concentration data are expressed as the mean ± S.E.M. The significance of differences in the mean plasma concentrations was assessed by one-way analysis of variance, followed by Dunnett's t test. A logistic model was used to fit dose-response data and plots of seizure protection versus plasma allopregnanolone levels. In all tests, the criterion for statistical significance was p < 0.05. Statistical analyses were carried out with PHARM/PCS version 4.2 (Microcomputer Specialists, Philadelphia, PA). Unless otherwise stated, each steroid dose was tested on groups of at least six mice.
Results
Antiseizure Activity of Progesterone in PTZ Model. To test the hypothesis that the antiseizure activity of progesterone occurs independently of its actions via PR, we examined the ability of progesterone to protect against PTZ-induced clonic seizures in male and female PRKO mice. In wild-type littermates, progesterone (10–150 mg/kg i.p.) protected against PTZ (85 mg/kg s.c.)-induced seizures in a dose-dependent manner (Fig. 1). Progesterone also protected against PTZ-induced seizures in heterozygous littermates and homozygous PRKO mice. In PRKO mice, the dose-response curve for antiseizure activity of progesterone was significantly (p < 0.05) shifted in a parallel manner to the left from that of wild-type animals. There was no significant difference in the antiseizure potency of progesterone in heterozygous and wild-type animals. The ED50 values derived from the dose-response data are given in Table 1. The ED50 values for progesterone in female and male PRKO animals were significantly less than the corresponding ED50 values in female and male wild-type littermates, indicating an increase in the antiseizure potency of progesterone in knockout mice. We also compared the potencies of progesterone for protection against PTZ seizures in female and male mice bearing the PRKO allelles. There were significant gender-related differences (p < 0.05) in the antiseizure potency of progesterone in female and male homozygous, heterozygous, and wild-type animals (Fig. 1); in each case, female mice were more sensitive to progesterone than were male mice with the same PRKO allele copy number (Table 1). In addition, we evaluated the effect of progesterone against PTZ-induced THE in male and female PRKO mice. Progesterone (10–150 mg/kg) dose dependently protected mice against PTZ-induced THE when tested 30-min postadministration. There was no significant difference in the protective potency of progesterone in female wild-type [ED50, 26 mg/kg (CL 14–48 mg/kg)] and PRKO mice [ED50, 27 mg/kg (CL 15–47 mg/kg)]. However, the ED50 values were markedly higher in male wild-type [ED50, 32 mg/kg (CL 19–53 mg/kg)] and PRKO mice [ED50, 41 mg/kg (CL 26–65 mg/kg)] compared with females. Progesterone (25–200 mg/kg i.p.) was also evaluated for its effects as a sedative and on motor activity using the inverted screen test in PRKO mice. As was the case for seizure protection in the PTZ test, progesterone's potency in the inverted screen test was significantly (p < 0.05) higher in PRKO mice (Table 1). There was also a trend toward increased activity of progesterone in the inverted screen test in female animals of all genotypes, but the gender-related differences did not reach statistical significance.

A, dose-response curves for protection against PTZ-induced seizures by progesterone in female homozygous PRKO mice and their female heterozygous (Het) and WT littermates. B, dose-response curves for protection against PTZ-induced seizures by progesterone in male PRKO, Het, and WT mice. For both females and males, the curves for PRKO animals are significantly shifted toward the left of the WT curve, indicating a higher (∼2-fold) protective potency of progesterone in PRKO than in WT mice. Progesterone (10–150 mg/kg i.p.) was administered 30 min before the PTZ (85 mg/kg s.c.) seizure test. Each point represents data from six to eight animals. ED50 values are given in Table 1.
ED50 values (milligrams per kilogram) for progesterone seizure protection in the PTZ test and for sedation/motor impairment in the inverted screen test in male and female wild-type, heterozygous, and PRKO mice Numbers in parentheses are 95% confidence limits. Values were obtained from log-probit fits to the data presented in Fig. 1.
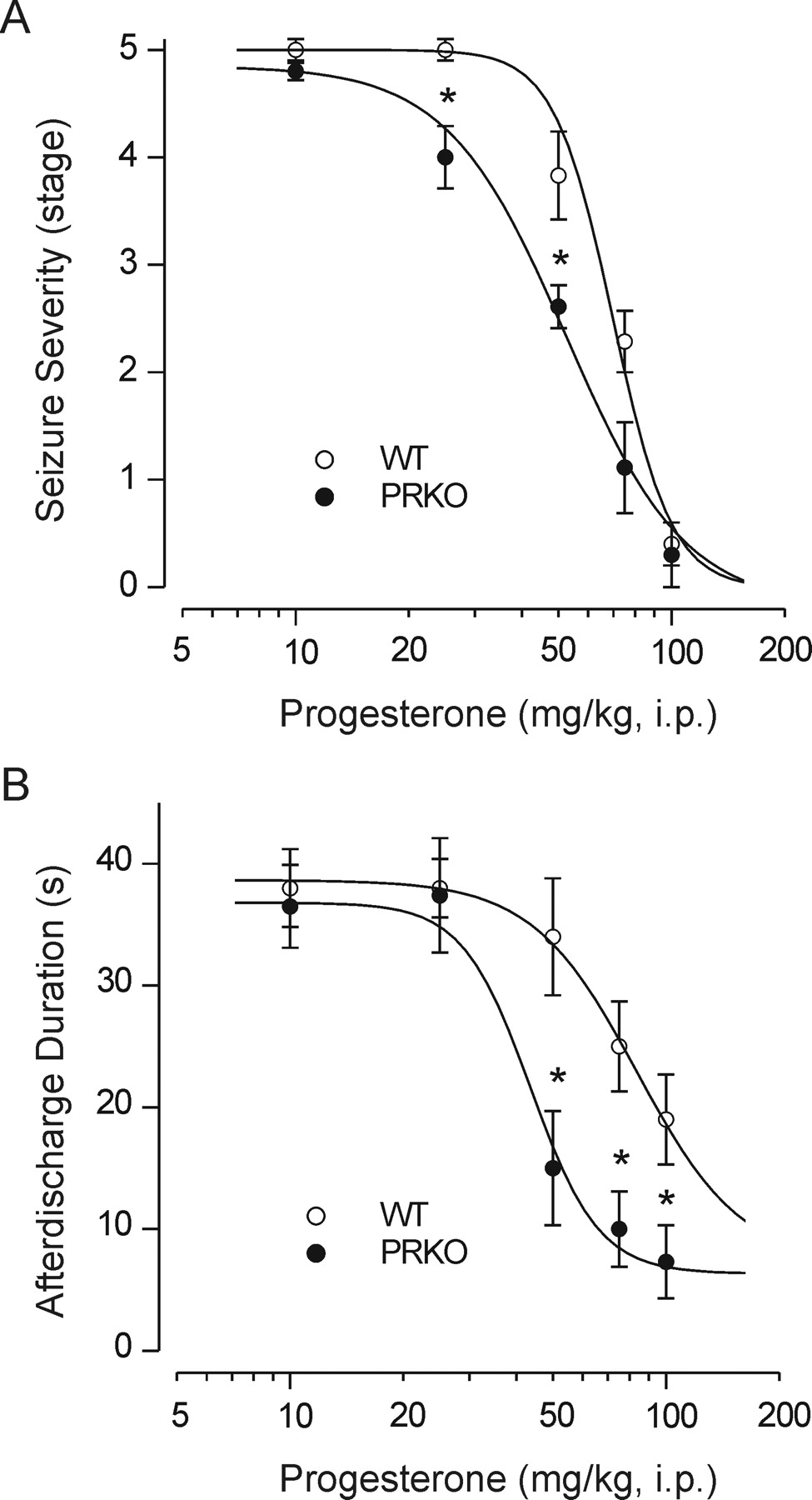
Antiseizure Activity of Progesterone in Kindling Model of Epilepsy. As shown in Fig. 2A, progesterone treatment (10–100 mg/kg i.p.) 30 min before electrical stimulation in fully kindled wild-type and PRKO animals produced a dose-dependent suppression of the behavioral seizure score, indicating that progesterone is protective in the amygdala kindling model. At the highest dose tested, behavioral seizures were nearly completely suppressed. As in the PTZ model, PRKO mice were significantly more sensitive to progesterone-induced inhibition of kindled seizures than were their wild-type littermate. Progesterone also reduced the afterdischarge duration in the kindled animals; as in the suppression of seizure stage, PRKO animals were more sensitive than were their wild-type littermates (Fig. 2B). On the day after progesterone treatment, all mice exhibited full-blown stage 5 seizures with normal duration afterdischarges, indicating that progesterone treatment suppresses the expression of kindled seizures but does not alter the kindled state (data not shown).

Inhibition of amygdala-kindled seizures by progesterone in female PRKO mice and their WT littermates. A, dose-response curves for progesterone (10–100 mg/kg i.p.)-induced suppression of behavioral seizure stage. The mean seizure severity scores for progesterone at 25 and 75 mg/kg is significantly less in PRKO than in WT mice, indicating that progesterone is more potent in PRKO mice (*, p < 0.05, Kruskal-Wallis test and Mann-Whitney U test). Mice were kindled as described under Materials and Methods and used when they exhibited fully kindled (stage 5) seizures on three consecutive days. Vehicle or progesterone was injected intraperitoneally 30 min before stimulation. Each point represents the mean ± S.E.M. (eight mice per group). B, dose-response curves for suppression of afterdischarge duration from the experiments in A. PRKO mice exhibit greater suppression than the WT controls. One-way ANOVA F (9, 69) = 8.98 (p < 0.001). *, p < 0.05 versus WT group (Dunnett's t test).
Antiseizure Activity of Progesterone in MES Seizure Test. Progesterone at high doses (50–200 mg/kg i.p.) exhibited dose-dependent protection in the MES seizure test in both female wild-type [ED50, 110 mg/kg (CL 85–143 mg/kg)] and PRKO mice [ED50, 113 mg/kg (CL 84–154 mg/kg)]. There was no statistically significant difference in the potencies in the two groups. The doses conferring protection in the MES test were associated with sedation and motor impairment (Table 1). However, muscle relaxation is unlikely to account for the protective activity as agents that produce motor impairment do not necessarily demonstrate positive activity in the MES test (White et al., 1995). Thus, progesterone-induced inhibition of THE in the MES test is a specific anti-convulsant effect and not secondary to sedation and motor impairment.
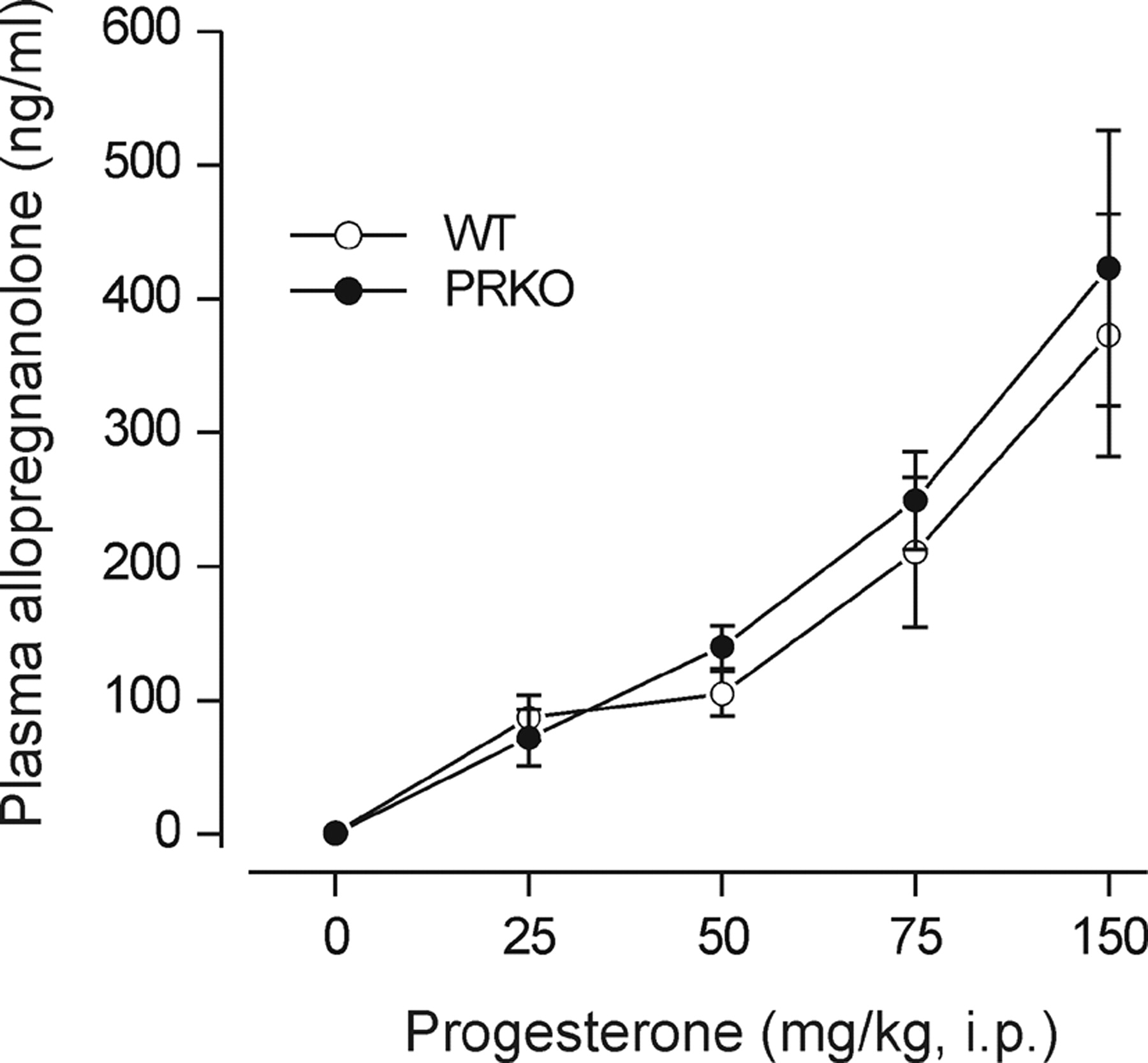
Allopregnanolone Plasma Levels: Correlation with Seizure Protection. Plasma concentrations of allopregnanolone were determined 30 min after the administration of various doses of progesterone (25–150 mg/kg i.p.) in wild-type and homozygous PRKO mice. In both groups of animals, mean plasma allopregnanolone concentrations increased in a dose-dependent manner with increasing progesterone dose (Fig. 3). There were no significant differences in the mean allopregnanolone plasma levels achieved between the genotypes at any of the progesterone doses. Figure 4 shows logistic fits to plots of seizure protection against plasma allopregnanolone concentration for female wild-type and PRKO mice in the PTZ and kindling models. These graphs were used to estimate the plasma concentrations of allopregnanolone producing 50% seizure protection (EC50) (see Fig. 4 legend). In both seizure models, the EC50 value in PRKO animals was less than that for wild-type mice, confirming the greater sensitivity in the PRKO animals.
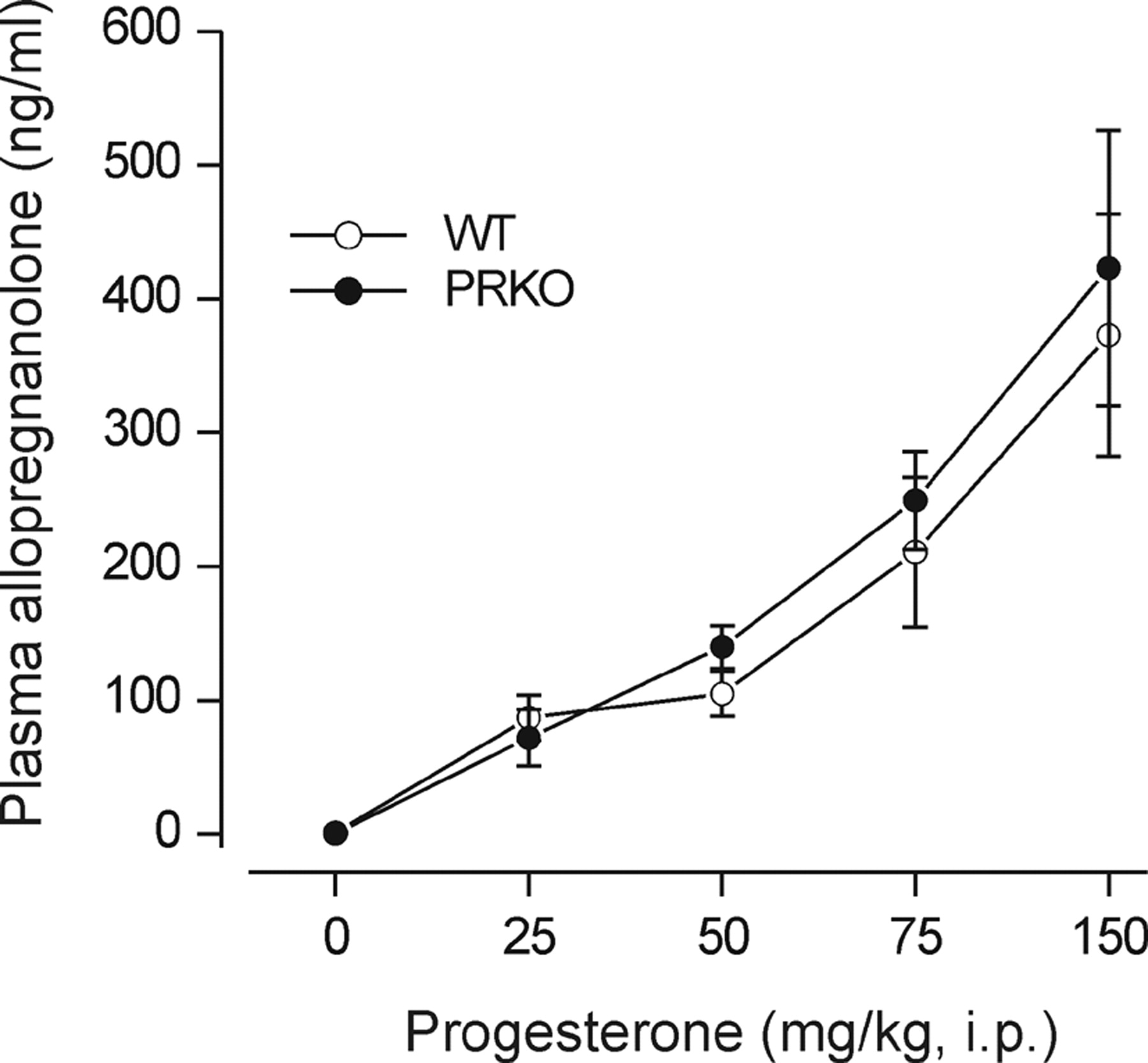
Plasma allopregnanolone levels in female homozygous PRKO mice and wild-type littermates after progesterone administration. Animals were injected with vehicle or various doses of progesterone (25–150 mg/kg). Thirty minutes later, plasma samples were collected for allopregnanolone determination by liquid chromatography-mass spectrometry. Plasma allopregnanolone levels for corresponding progesterone doses between PRKO and wild-type animals were not significantly different, but they were significantly elevated from vehicle control (Dunnett's t test). Data points represent the mean ± S.E.M. (four to seven mice per group).
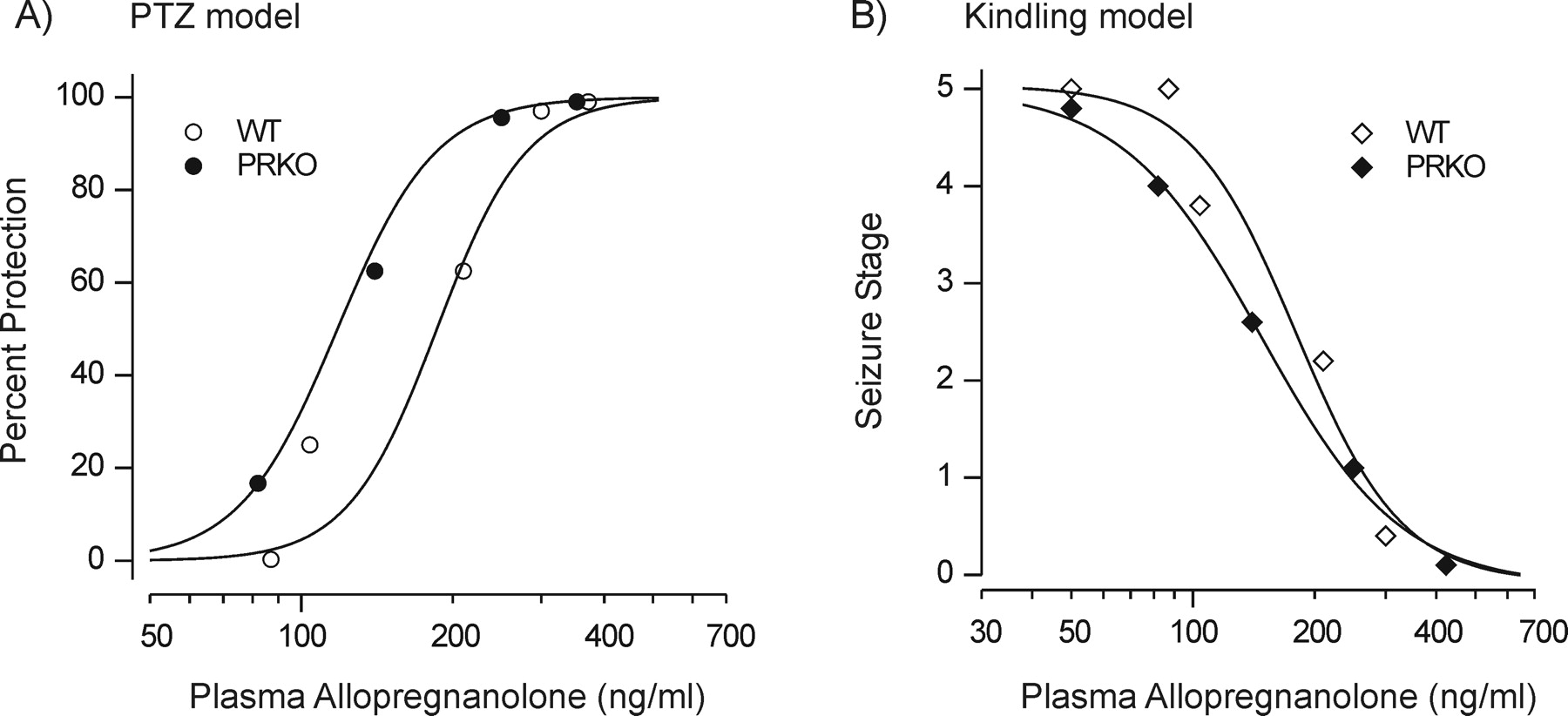
Correlation between plasma allopregnanolone concentrations and seizure protection in the PTZ and kindling models. A, plasma allopregnanolone levels after various doses of progesterone (from Fig. 3) in WT and PRKO female mice were plotted against PTZ seizure protection values from Fig. 1A. The data points were fit with a logistic function (wild-type EC50 = 190 ng/ml; PRKO EC50 = 125 ng/ml). B, similar analysis from the kindling experiment of Fig. 2A (wild-type EC50 = 180 ng/ml; PRKO EC50 = 150 ng/ml).
Finasteride Reverses the Antiseizure Activity of Progesterone: Correlation with Allopregnanolone Levels. To confirm that the antiseizure activity of progesterone in PRKO mice is due to its conversion to the neurosteroid allopregnanolone, PRKO mice and wild-type controls were pretreated with the 5α-reductase inhibitor finasteride (Azzolina et al., 1997), which has previously been shown to block the seizure protection conferred by progesterone (Kokate et al., 1999). As shown in Fig. 5, finasteride (5–200 mg/kg i.p.) produced a dose-dependent inhibition of the protective activity of progesterone (100 mg/kg i.p.) in the PTZ test in both female PRKO mice and female wild-type controls. At 200 mg/kg finasteride, there was almost complete inhibition of the anticonvulsant activity of progesterone in both groups. The ED50 value for finasteride inhibition of the anticonvulsant activity of progesterone in PRKO mice was 69 mg/kg (95% CL 35–136 mg/kg). We also determined the influence of finasteride on the protective effects of progesterone in the amygdala kindling model. Pretreatment with finasteride (50 mg/kg) significantly reversed the effect of progesterone (75 mg/kg) on kindled seizure score (Fig. 6A). In these experiments, measurements of plasma allopregnanolone concentrations (Fig. 6B) confirmed that finasteride significantly decreased (∼60% in wild-type mice; ∼51% in PRKO mice) the allopregnanolone levels. In contrast to the PTZ and kindling models, finasteride (100 mg/kg) did not affect the protective activity of progesterone (200 mg/kg) in the MES test in PRKO mice (five of five mice were protected from MES seizures).

Finasteride (5–200 mg/kg i.p.) inhibition of the protective activity of progesterone (100 mg/kg i.p.) in the PTZ seizure model in female PRKO and WT mice. Animals were pretreated with finasteride 60 min before the progesterone injection and challenged with PTZ 30 min later. The fractional protection obtained in the absence of finasteride was taken as 100% for calculating percentage of inhibition by finasteride (wild-type, 76%; PRKO, 98%). Each point represents data from seven to nine mice.

Finasteride inhibition of the protective activity of progesterone in the kindling model. A, fully kindled female wild-type and PRKO mice received an injection of progesterone (75 mg/kg i.p.) followed 30 min later by kindling stimulation. Some animals were pretreated with finasteride (50 mg/kg i.p.) 60 min before the progesterone, which significantly increased the seizure stage in both groups. (The increase represents a reversal of the progesterone-induced reduction as seen by comparison with the data presented in Fig. 3.) B, plasma allopregnanolone levels 30 min after progesterone (75 mg/kg i.p.). Some animals were pretreated with finasteride (50 mg/kg i.p) 60 min before progesterone. Each bar represents mean ± S.E.M. (six to eight mice per group). #, p < 0.05 versus progesterone alone (Mann-Whitney U test); *, p < 0.05 versus progesterone alone (Dunnett's t test).
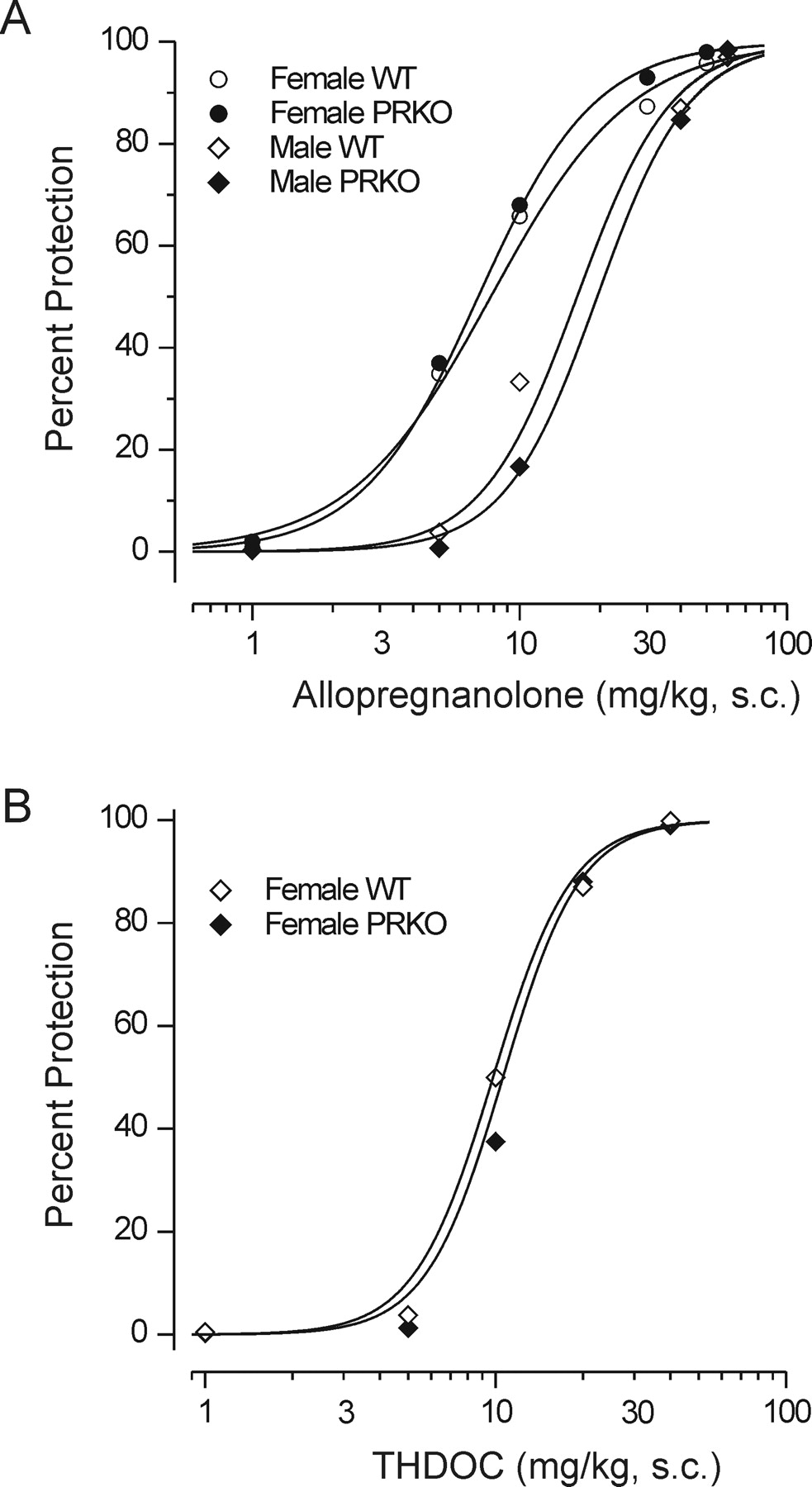
Anticonvulsant Activity of Neurosteroids. The neurosteroids allopregnanolone and allotetrahydrodeoxycorticosterone (THDOC) were also evaluated for protective activity in the PTZ test. As illustrated in Fig. 7, both neurosteroids protected PRKO mice and wild-type controls in a dose-dependent manner with similar potencies in the two genotypes. For both genotypes, the dose-response curve for allopregnanolone in male animals was shifted to the right of that for females, indicating reduced potency in the males. The differences between males and females did not reach statistical significance by the Litchfield and Wilcoxon χ2 test; however, there were significant differences in the percentage of protection values at 5 and 10 mg/kg for both genotypes. The ED50 values obtained from the dose-response data of Fig. 7 are presented in Table 2. Pretreatment with finasteride (200 mg/kg i.p.) had no effect on the anticonvulsant activity of allopregnanolone (30 mg/kg s.c.) in wild-type and PRKO mice (five of five mice were protected from seizure occurrence).

Protective activity of neurosteroids allopregnanolone and THDOC against PTZ-induced seizures. Dose-response curves for allopregnanolone and THDOC were similar in female WT and PRKO mice. Male animals exhibited reduced sensitivity to allopregnanolone compared with females at the 5- and 10-mg/kg doses (Table 2). Allopregnanolone (1–50 mg/kg s.c.) and THDOC (1–40 mg/kg s.c.) were administered 15 min before injection of PTZ (85 mg/kg s.c.). Each point represents data from six to eight mice.
Anticonvulsant ED50 values (milligrams per kilogram) of neurosteroids in the PTZ test in wild-type and PRKO mice Numbers in parentheses are 95% confidence limits. Values were obtained from log-probit fits to the data presented in Fig. 7.
Discussion
The key observation in this study is that the anticonvulsant activity of progesterone in several seizure models is undiminished in PRKO mice, which lack both forms of the PR, thus definitively proving that the PR is not required for the anticonvulsant activity of progesterone in these models. Although our study is conclusive on this point, there are certain caveats that must be considered in regard to the interpretation of the findings. First, the results apply only to the seizure models used in this study. Moreover, because the relationship of these models to human epilepsies is not well defined, it cannot be concluded that PRs do not participate in the regulation of seizure susceptibility in the clinical setting. Second, the relevance of the doses of progesterone that were used in this study in relation to those required to occupy brain PRs is uncertain. To obtain anticonvulsant effects, it was necessary to use doses of progesterone of 10 to 25 mg/kg and higher. Doses within this same range have been demonstrated to produce circulating levels of progestins similar to those in proestrous and are often used when examining the behavioral actions of progesterone believed to be mediated through genomic actions (Frye and Vongher, 2001). Nevertheless, we did not verify the extent to which these doses induce genomic effects in brain. Interestingly, however, it has recently been shown that doses of progesterone similar to those used here result in the rapid transcriptional induction of 5α-reductase type II in mouse brain (Matsui et al., 2002), demonstrating that genomic effects do occur with comparable doses.
Previous studies have indicated that 5α-reduced metabolites of progesterone, predominantly allopregnanolone, account for the anticonvulsant activity of the parent hormone in both animals and humans (Frye et al., 1998, 2002; Kokate et al., 1999; Herzog and Frye, 2003). In accordance with this concept, the 5α-reductase inhibitor finasteride reversed the anticonvulsant effects of progesterone in PRKO mice in the PTZ and kindling models. These results confirm prior reports demonstrating that finasteride inhibits the protective activity of progesterone in several seizures models in genetically normal mice and rats (Frye et al., 1998; Kokate et al., 1999). 5α-Reductase exists in two isoenzymes, designated type I and type II (Celotti et al., 1997), and in rodents finasteride inhibits both forms (Azzolina et al., 1997). After exogenously administered progesterone or in states of high endogenous progesterone, finasteride treatment is associated with a substantial reduction in circulating and brain levels of 5α-reduced progesterone metabolites, including allopregnanolone (Frye et al., 1998; Frye and Bayon, 1998; Reddy et al., 2001b). However, under ordinary conditions, finasteride does not affect the propensity for seizures in either the PTZ or kindling models, and it also does not affect the anticonvulsant activity of allopregnanolone, indicating that it does not influence seizure susceptibility apart from its actions on 5α-reductase (Kokate et al., 1999; Reddy and Rogawski, 2000, 2002). Therefore, our results support the conclusion that the anticonvulsant activity of progesterone in the PTZ and kindling models results not from effects on the PR but rather depend on the conversion of progesterone to 5α-reduced metabolites, predominantly allopregnanolone, which are known to act through modulatory effects on GABAA receptors (Lambert et al., 2003; Rogawski and Reddy, 2004).
In studies with genetically normal animals, progesterone only exhibited protection in the MES test at high doses, and this action was not influenced by finasteride, leading to the conclusion that progesterone does not act through conversion to allopregnanolone in this model (Spiegel and Wycis, 1945; Kokate et al., 1999). We confirmed these results in the present study, and because progesterone had equivalent activity in wild-type and PRKO animals, we can eliminate the PR as a possible mediator. Moreover, this is in agreement with the general insensitivity of MES seizure model to agents that enhance GABA-mediated inhibition (Rogawski, 2002). The precise mechanism of action of progesterone in the MES model is unknown. It will be of interest to determine whether high doses of progesterone interact directly with ion channels other than GABAA receptors to confer protection.
Because allopregnanolone and the deoxycorticosterone-derived neurosteroid THDOC, which modulates GABAA receptors like allopregnanolone (Kokate et al., 1994; Reddy, 2003a), had similar anticonvulsant potencies in wild-type and PRKO mice, we conclude that the PR does not contribute to the protective effects of the neurosteroids. In fact, in contrast to progesterone, allopregnanolone and THDOC bind poorly to the PR (Iswari et al., 1986). However, it has been proposed that these neurosteroids can be oxidized intracellularly to 5α-pregnane metabolites that could potentially activate the PR (Rupprecht et al., 1993). Our results with PRKO mice indicate that this potential mechanism is not relevant in regard to the anticonvulsant effects of allopregnanolone and THDOC.
We unexpectedly observed that PRKO mice were more sensitive to the anticonvulsant effects of progesterone in the PTZ and the kindling models than wild-type or heterozygous animals. PRKO animals were also more sensitive to the sedative actions of progesterone. However, as noted, PRKO mice were not more sensitive to the anticonvulsant activity of allopregnanolone. This indicates that the enhanced potency is not attributable to alterations in target sensitivity in the absence of exogenous progesterone. To investigate the possibility that the difference results from alterations in the absorption, distribution, or metabolism of progesterone causing greater allopregnanolone levels in the PRKO mice, we determined the plasma allopregnanolone concentrations achieved with various progesterone doses. At no dose did PRKO mice exhibit significantly increased plasma allopregnanolone levels, demonstrating that pharmacokinetic factors cannot account for the altered progesterone sensitivity. We also note that a previous study found little or no differences in the plasma concentrations of progesterone between PRKO and wild-type mice (Chappell et al., 1997), indicating that the heightened sensitivity to progesterone in PRKO mice is not due to elevated endogenous progesterone levels in these animals. Measurements of plasma allopregnanolone levels allowed us to directly correlate the progesterone-induced elevation in plasma concentrations with seizure protection. There was a close correlation between the plasma allopregnanolone concentrations achieved with various progesterone doses and the seizure protection obtained in both seizure models. Progesterone was found to protect against PTZ and kindled seizures in PRKO mice at plasma allopregnanolone concentrations that were lower than those in wild-type mice. For example, in wild-type and PRKO mice, the estimated plasma concentrations producing 50% seizure protection (EC50) in the PTZ test were 190 and 125 ng/ml, respectively. These observations indicate that the enhanced progesterone responsiveness of PRKO mice is not related to pharmacokinetic factors. Rather, progesterone acting via the PR may cause reduced sensitivity to progesterone-derived allopregnanolone in animals that express PR (wild-type and heterozygous animals), but this mechanism is presumably inoperative in animals lacking PR, thus accounting for their enhanced sensitivity. We have previously reported that PRKO mice have a higher threshold for PTZ convulsions and exhibit delayed kindling development (Reddy et al., 2001a), indicating that these animals have a reduced susceptibility to seizures. The way in which the PR regulates responsiveness to the anticonvulsant effects of neurosteroids and alters seizure susceptibility is unknown. However, our recent analyses of gene expression profiles in the hippocampi of PRKO mice using DNA microarray techniques and Western blotting have identified changes in glutamate receptor subunits that could be relevant (M. Bevers, D. C. Castaneda, D. S. Reddy, and M. A. Rogawski, unpublished data). Moreover, real-time TaqMan PCR analysis revealed significant progesterone-induced alterations in GABAA receptor subunit expression in the hippocampus (Reddy, 2003b), indicating that PR could participate in GABAA receptor transcriptional regulation.
It has previously been observed that female mice and rats are more sensitive to the central nervous system actions of progesterone than are males (Holmes and Weber, 1984; Finn and Gee, 1994; Mohammad et al., 1998; Reddy and Kulkarni, 1999). Our finding that wild-type, heterozygous, and PRKO female mice are more sensitive to the anticonvulsant effects of progesterone than males (Table 1) confirms this result. Moreover, the fact that female PRKO animals also exhibit greater progesterone sensitivity demonstrates that known differences in the density or distribution of the PR in the female and male brain (Rainbow et al., 1982) are not responsible for the sexual dimorphism. Because the brain levels of enzymes involved in conversion of progesterone to allopregnanolone are similar in both sexes (Mensah-Nyagan et al., 1999), the enhanced potency of progesterone in female mice is also unlikely to be due to differences in local progesterone metabolism. In fact, we found that allopregnanolone itself was also a more potent anticonvulsant in females than in males (Table 2). This is unlikely to be due to differences in the sensitivity of GABAA receptors to allopregnanolone because, if anything, GABAA receptors in female rodent limbic brain regions are less sensitive to allopregnanolone than in the male (Wilson and Biscardi, 1997). However, hormonal differences between females and males might be responsible. For example, testosterone and related androgens have been demonstrated to decrease allopregnanolone's ability to protect against seizures (Edwards et al., 2001b), which could contribute to the reduced potency in males. Alternatively, there could be gender differences in the bioavailability or pharmacokinetics of progesterone and allopregnanolone.
In conclusion, our results with PRKO mice provide strong evidence that the PR is not required for the antiseizure effects of progesterone in any of the models examined in this study. In the PTZ and kindling models, the anticonvulsant activity of progesterone likely resides in its metabolic conversion to allopregnanolone. This is consistent with the well recognized sensitivity of these seizure models to agents that enhance GABA-mediated inhibition. However, the situation is different for the MES test. As expected by the lack of activity of GABA-modulating agents in this model, other (and as yet poorly understood) properties of the steroid seem to play a role.
Acknowledgments
We thank Drs. Shun-ichi Yamaguchi (National Institute of Neurological Disorders and Stroke) and John P. Lydon (Baylor College of Medicine) for expert assistance.
Footnotes
-
This study was supported in part by National Institutes of Health Grants HD 07857/07495 (to B.W.O.) and by a North Carolina State College of Veterinary Medicine grant (to D.S.R.).
-
DOI: 10.1124/jpet.104.065268.
-
ABBREVIATIONS: PR, progesterone receptor; PRKO, progesterone receptor knockout; PTZ, pentylenetetrazol; MES, maximal electroshock; THE, tonic hindlimb extension; CL, confidence limits; THDOC, allotetrahydrodeoxycorticosterone; R5020, promegestone; WT, wild type.
- Received January 15, 2004.
- Accepted February 23, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}