


Abstract
Clozapine is the prototype atypical antipsychotic drug, producing little or no extrapyramidal side effects, while improving negative symptoms of psychosis. Clozapine's high affinity for serotonin receptors has been hypothesized to confer the unique antipsychotic properties of this drug. Recently, we demonstrated that both typical and atypical antipsychotic drugs are inverse agonists at constitutively active 5-hydroxytryptamine2A (5-HT2A) receptors. To determine whether inverse agonist activity at 5-HT2Creceptors plays a role in antipsychotic efficacy, typical and atypical antipsychotic drugs were tested for inhibition of basal inositol phosphate production in mammalian cells expressing rat or human 5-HT2C receptors. Atypical antipsychotic drugs (sertindole, clozapine, olanzapine, ziprasidone, risperidone, zotepine, tiospirone, fluperlapine, tenilapine) displayed potent inverse agonist activity at rat and human 5-HT2C receptors. Typical antipsychotic drugs (chlorpromazine, loxapine, thioridazine, prochlorperazine, perphenazine, mesoridazine, trifluperidol, fluphenazine, spiperone, haloperidol, pimozide, penfluridol, thiothixene) were devoid of inverse agonist activity, with the exception of loxapine. We review the evidence that loxapine has unique properties characteristic of both atypical and typical antipsychotic drugs. Several typical antipsychotic drugs (chlorpromazine, thioridazine, spiperone, thiothixene) displayed neutral antagonist activity by reversing clozapine inverse agonism. These data suggest that 5-HT2C inverse agonist activity is associated with atypical antipsychotic drugs with moderate to high affinity for 5-HT2C receptors, and imply that effects of atypical antipsychotic drugs on the 5-HT2C receptor may play a role in their unique clinical properties. These data also imply that dysfunction of brain 5-HT2C receptor systems may be one of the factors involved in the etiology of psychosis.
Clozapine is the prototype atypical antipsychotic. It is effective in treating schizophrenics refractory to classical antipsychotic drugs, produces fewer extrapyramidal side-effects, and appears to be more effective in reducing many symptoms of the schizophrenic syndrome (Meltzer and McGurk, 1999). Although it is clear that clozapine must have a unique site or site(s) of action in the brain, the precise mechanism by which clozapine achieves its superior effects has not been discerned. Clozapine produces agranulocytosis in approximately 1% of the patient population, and thus blood monitoring is recommended during treatment with clozapine. Therefore, a clozapine-like drug (atypical antipsychotic) without deleterious hematological effects is desirable. Furthermore, an insight into the mechanism of action of clozapine may add to our knowledge of the etiology of schizophrenia.
It is clear that the potency of the “typical” antipsychotic drugs is correlated with their affinity for the D2dopamine receptor subtype, based on radioligand binding studies and human data (Creese et al., 1976; Seeman et al., 1976). Clozapine possesses D2 dopamine receptor-blocking activity and also has been found to display high affinity for various types of 5-hydroxytryptamine (5-HT) receptors, most notably the 5-HT2A and 5-HT2C receptors (Meltzer et al., 1989; Roth et al., 1992). With the discovery of new dopamine receptor subtypes (D3, D4) and more 5-HT receptor subtypes (5-HT6, 5-HT7), the receptor pharmacology of clozapine has become more complex. In addition to having high affinity for D4 receptors (Seeman et al., 1997) and 5-HT6 and 5-HT7 receptors (Roth et al., 1994), clozapine also has high affinity for adrenergic, muscarinic, and histamine receptors (Roth et al., 1998; Meltzer and McGurk, 1999).
A popular working hypothesis on the mode of action of atypical antipsychotics is that their unique actions depend, in some as-yet-unspecified manner, on their interaction with brain 5-HT2A or 5-HT2C receptors, while also occupying brain D2 dopamine receptors. It has been reported that there is a significant correlation between the 5-HT2A/D2 receptor affinity ratio and atypical antipsychotic properties (Meltzer et al., 1989): these drugs bind to 5-HT2A receptors with higher affinity than to D2 receptors but occupy both receptor populations in the brain and appear to be the best candidates for atypical antipsychotic activity. In addition, several atypical antipsychotic drugs have higher affinity for 5-HT2C receptors than D2receptors (Roth et al., 1994). Thus the data support a role for 5-HT2A, 5-HT2C, or both receptors in the actions of atypical antipsychotic drugs. These types of drugs have been referred to as “SDI” drugs (serotonin, dopamine inhibitors). Clozapine is an excellent example of this: it has an affinity of 60 nM for the D2 receptor and 5 to 10 nM for 5-HT2A and 5-HT2Creceptors (Roth et al., 1994).
5-HT2A and 5-HT2C receptors have been implicated in the control of cognition and are widely distributed throughout the brain (Titeler et al., 1988; Hoffman and Mezey, 1989; Kennett et al., 1994; Jakab and Goldman-Rakic, 1998; Marek and Aghajanian, 1998; Millan et al., 1998). Both 5-HT2A and 5-HT2C receptors are linked to the stimulation of intracellular inositol phosphate (IP) levels through G-protein-mediated mechanisms (Sanders-Bush and Conn, 1986; Barker et al., 1991). Constitutively active mutant (CAM) forms of 5-HT2A and 5-HT2C receptors have been produced by site-directed mutagenesis (Herrick-Davis et al., 1997; Egan et al., 1998). Drugs that were previously classified as competitive antagonists, based on their properties in blocking agonist stimulation of native 5-HT2A and 5-HT2C receptors, were found to demonstrate inverse agonist activity at CAM 5-HT2A and 5-HT2C receptors. Both typical and atypical antipsychotic drugs behave as inverse agonists at the CAM C322K 5-HT2A receptor (Egan et al., 1998), while preliminary studies indicate that atypical antipsychotic drugs are inverse agonists at the CAM S312K 5-HT2C receptor (Herrick-Davis et al., 1998). The preliminary studies were performed using a recombinant cell line expressing a CAM form of the rat 5-HT2C receptor. Recently, it was demonstrated that 5-HT2C receptors undergo RNA editing to produce multiple isoforms (Burns et al., 1997) and that the human 5-HT2C receptor displays significant basal activity in its native, unedited (5-HT2C-INI) form (Herrick-Davis et al., 1999, Niswender et al., 1999). Therefore, the present study was designed to evaluate typical and atypical antipsychotic drugs for inverse agonist activity at native human 5-HT2C-INI receptors expressed in a mammalian cell line.
Experimental Procedures
Materials.
COS-7 cells were purchased from the American Type Culture Collection. Chemicals and reagents were purchased as follows: [3H]mesulergine (Amersham Pharmacia Biotech, Piscataway, NJ); myo-[3H]inositol (NEN Life Science Products, Boston, MA); 5-HT, lithium chloride, pargyline, fetal bovine serum, and all buffers for IP assays (Sigma Chemical Co., St. Louis, MO); lipofectAMINE and cell culture medium (Life Technologies, Inc., Gaithersburg, MD); anion exchange resin and columns (Bio-Rad, Richmond, CA); Ecoscint cocktail (National Diagnostics, Manville, NJ). Antipsychotic drugs were provided by the National Institute of Mental Health psychoactive drug screening program, purchased from Research Biochemicals International (Natick, MA), or donated by the manufacturer (sertindole: Lundbeck, Copenhagen, Denmark; ziprasidone: Pfizer, New York, NY). Human 5-HT2C-INI cDNA in pCMV2 was provided generously by Dr. Elaine Sanders-Bush, Departments of Pharmacology and Psychiatry and the Center for Molecular Neuroscience, Vanderbilt University (Nashville, TN). Rat S312K 5-HT2C-VSI was created by site-directed mutagenesis as described previously (Herrick-Davis et al., 1997).
Cell Culture and Transfection.
COS-7 cells were grown in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum in a humidified incubator with 5% CO2 at 37°C. Twenty-four hours before transfection, cells were seeded at 105 cells/well in 24-well cluster plates for IP assays and for [3H]mesulergine binding studies performed in parallel to monitor receptor expression. Cells were transfected with the rat or human 5-HT2C receptor by combining 2 μl of lipofectAMINE with 0.5 μg of plasmid DNA in 400 μl of serum-free DMEM and added to each well for 5 h at 37°C/5% CO2. For radioligand binding studies, COS-7 cells were seeded at 80% confluence in 100-mm dishes and transfected with 5 μg of plasmid DNA, 20 μl of lipofectAMINE in 4 ml of serum-free DMEM for 5 h at 37°C/5% CO2. After transfection, cells were returned to complete culture medium for 48 h before membrane preparation for radioligand binding studies.
IP Production Assays.
IP production was measured as previously described (Herrick-Davis et al., 1999). In brief, 24 h after transfection COS-7 cells were washed with PBS and labeled overnight with 0.5 μCi/well ofmyo-[3H]inositol in inositol-free/serum-free DMEM at 37°C/5% CO2. After labeling, cells were washed with PBS and preincubated in inositol-free/serum-free DMEM with 10 mM LiCl and 10 μM pargyline (assay medium) for 10 min. Antipsychotic drugs were added during the 10-min preincubation. 5-HT, or assay medium alone, was added to each well and incubation continued for an additional 35 min to determine basal activity. Assay medium was removed and cells were lysed in 200 μl of stop solution (1 M KOH/18 mM sodium borate/3.8 mM EDTA) and neutralized by adding 200 μl of 7.5% HCl. The contents of each well were extracted with 3 volumes of chloroform:methanol (1:2, v/v) and centrifuged 5 min at 10,000g, and the upper layer was loaded onto a 1-ml AG1-X8 resin (100–200 mesh) column. Columns were washed with 10 ml of 5 mM myo-inositol and 10 ml of 5 mM sodium borate/60 mM sodium formate. Total [3H]IPs were eluted with 3 ml of 0.1 M formic acid/1 M ammonium formate. Radioactivity was measured by liquid scintillation counting in Ecoscint cocktail.
Radioligand Binding.
Membranes were prepared by scraping a confluent 100-mm dish of transfected COS-7 cells into 20 ml of 50 mM Tris-HCl/5 mM MgSO4/0.5 mM EDTA, pH 7.4 (assay buffer) and centrifugation at 10,000g for 30 min. Membranes were resuspended in 20 ml of assay buffer, homogenized, and centrifuged again. After resuspension in 15 ml of assay buffer, 0.5-ml membrane aliquots were added to each assay tube containing 1 nM [3H]mesulergine and varying concentrations of competing drug in a final volume of 1 ml. Mianserin (10 μM) was used to define nonspecific binding. Samples were incubated at 37°C for 30 min, filtered through glass fiber filters (presoaked in 0.3% polyethylenamine) on a Brandel cell harvester, and counted in Ecoscint cocktail in a liquid scintillation counter (Beckman, Berkeley, CA) at 40% efficiency.
Data Analyses.
Data analyses were performed using Prism software (GraphPad, San Diego, CA). The Cheng/Prusoff equation was used to calculate Ki values from IC50 values.
Results
The present study was performed to determine whether antipsychotic drugs are inverse agonists at the 5-HT2C receptor and to determine whether this is a specific feature of atypical antipsychotic drugs, not shared by typical antipsychotic drugs. A preliminary study was performed using a stable cell line expressing CAM (S312K) rat 5-HT2C-VSI receptors (Fig.1). IP production was measured in NIH-3T3 cells labeled with myo-[3H]inositol and challenged with 1 μM atypical or 10 μM typical antipsychotic drug. Four atypical antipsychotic drugs (clozapine, olanzapine, ziprasidone, risperidone) displayed significant inverse agonist activity at 1 μM. However, four typical antipsychotic drugs (haloperidol, chlorpromazine, thioridazine, perphenazine) were devoid of inverse agonist activity at 10 μM.

Inhibition of rat 5-HT2Creceptor basal activity by antipsychotic drugs. [3H]IP production was measured in NIH-3T3 cells stably expressing a constitutively active mutant (S312K) rat 5-HT2C-VSI receptor at 5 pmol/mg of protein. Cells were treated with 1 μM atypical or 10 μM typical antipsychotic drug. IC50 values for clozapine and risperidone were 132 ± 9.7 nM and 207 ± 41 nM, respectively. Data represent the mean ± S.E. of three independent experiments. *P < .01 versus basal.
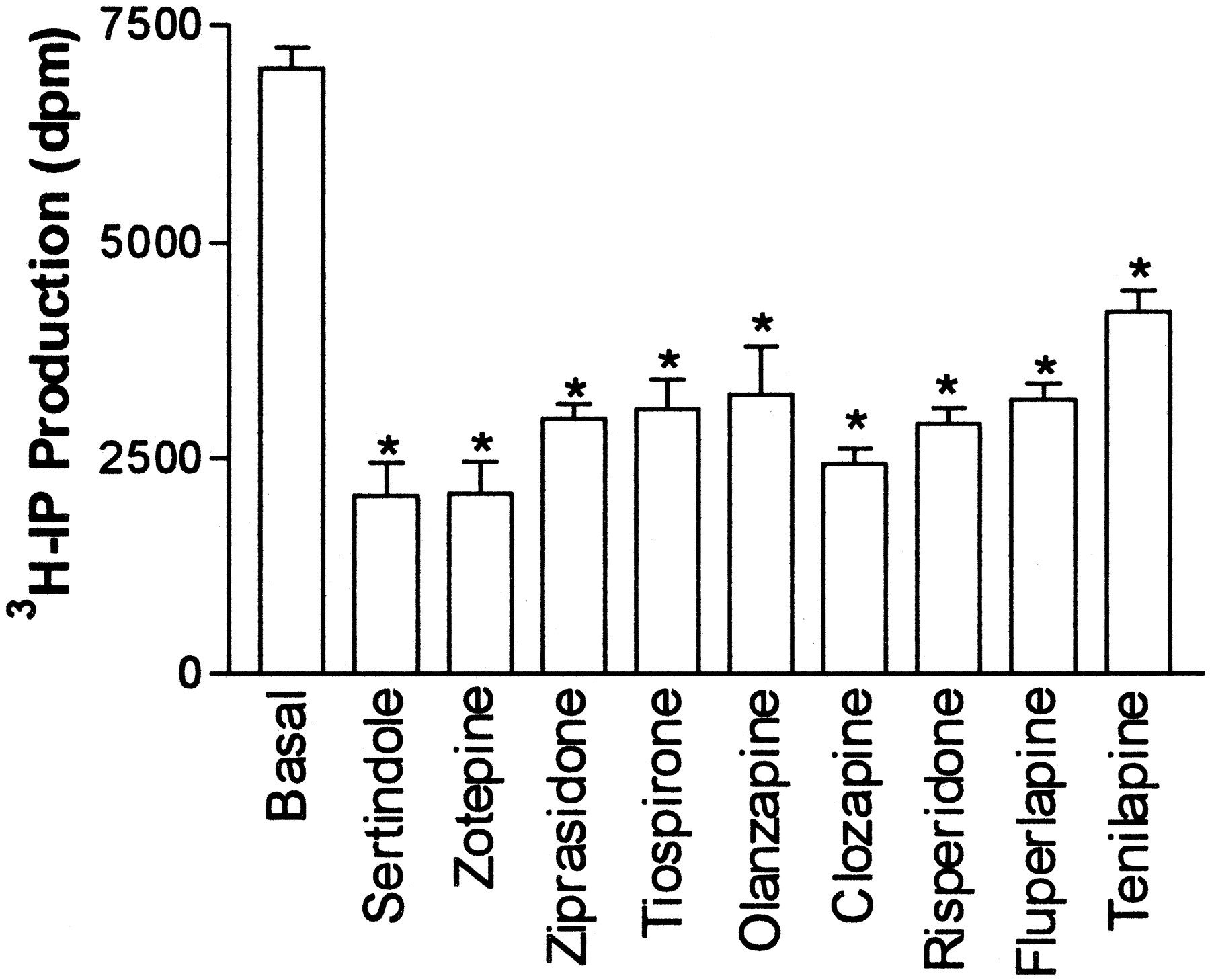
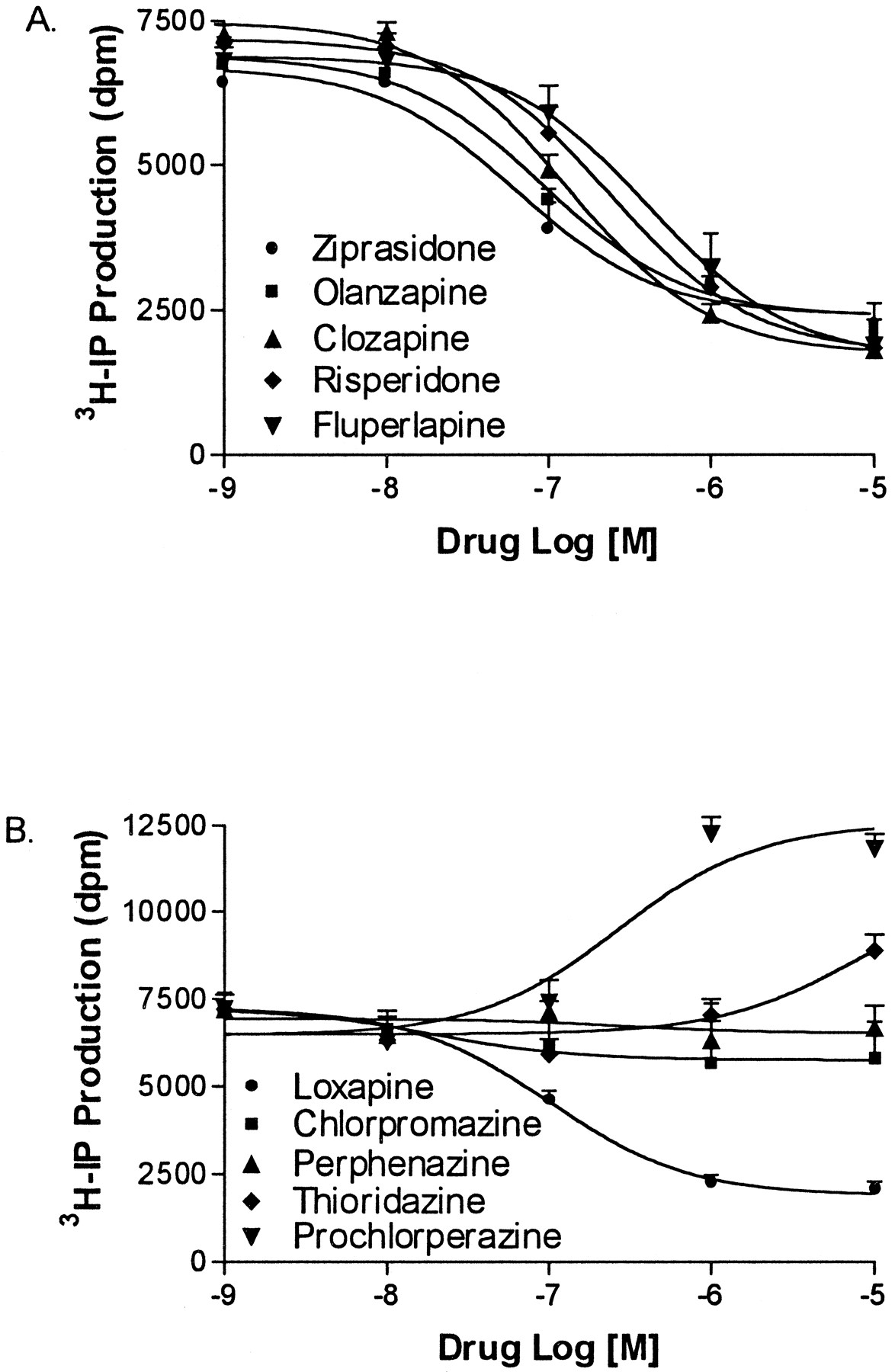
Based on these results, a more extensive study was performed to evaluate nine atypical (sertindole, clozapine, zotepine, ziprasidone, tiospirone, olanzapine, risperidone, fluperlapine, tenilapine) and 13 typical (chlorpromazine, loxapine, thioridazine, prochlorperazine, perphenazine, mesoridazine, trifluperazine, fluphenazine, spiperone, haloperidol, pimozide, penfluridol, thiothixene) antipsychotic drugs for inverse agonist activity at human 5-HT2C-INIreceptors. The atypical antipsychotic drugs used in this study were chosen because they have been reported to display high affinity (Ki < 100 nM) for rat 5-HT2C receptors (Roth et al., 1992, 1994). Figure 2 shows the basal level of IP formation produced by native, human 5-HT2C-INIreceptors expressed in COS-7 cells. Cells transfected with vector only (without 5-HT2C cDNA) were used to measure the amount of IP produced by COS-7 cells in the absence of 5-HT2C receptor expression. Basal IP production stimulated by 5-HT2C receptors was measured by subtracting the amount of IP produced in cells transfected with vector. All of the atypical antipsychotic drugs produced >50% reduction in 5-HT2C-INI receptor basal activity when tested at 1 μM (Fig. 2). Inverse agonist concentration-response curves for atypical antipsychotic drugs (ziprasidone, olanzapine, clozapine, risperidone, fluperlapine) are shown in Fig.3A.

Inhibition of human 5-HT2Creceptor basal activity by atypical antipsychotic drugs. [3H]IP production was measured in COS-7 cells expressing human 5-HT2C-INI receptors at 2.7 pmol/mg of protein. Cells were treated with or without 1 μM atypical antipsychotic. Basal activity was determined by subtracting [3H]IP produced in cells transfected with vector only. Data represent the mean ± S.E. of three independent experiments. *P < .01 versus basal.

Log dose-response curves for inhibition of 5-HT2C receptor basal activity by antipsychotic drugs. Human 5-HT2C-INI receptor basal activity was measured after treatment with increasing concentrations of antipsychotic drug. Basal activity was determined by subtracting [3H]IP produced in cells transfected with vector only. A, atypical antipsychotic drugs. IC50 values are given in Table 1. B, typical antipsychotic drugs. Loxapine IC50 = 94 ± 1.4 nM. Prochlorperazine EC50 = 270 ± 2.8 nM. Data represent the mean ± S.E. of three independent experiments.
Sertindole, olanzapine, zotepine, ziprasidone, and tiospirone were the most potent inhibitors of basal activity with inverse agonist IC50 values <100 nM (Table1). At 10 μM, the atypical antipsychotic drugs inhibited approximately 70% of the basal activity, with the exception of tiospirone (59%) and tenilapine (61%). The binding affinities for atypical antipsychotic drugs at the human 5-HT2C-INI receptor are given in Table 1. Sertindole, zotepine, ziprasidone, tiospirone, olanzapine, and clozapine all displayed Ki values ≤10 nM for [3H]mesulergine binding, while risperidone had a Ki value of 24 nM.
Atypical antipsychotic binding affinity and inverse agonist potency at human 5-HT2C-INI receptors
In the next series of experiments, typical antipsychotic drugs were tested for inverse agonist activity at human 5-HT2C-INI receptors. Concentration-response curves for five moderate to high affinity typical antipsychotic drugs are shown in Fig. 3B. Only loxapine demonstrated inverse agonist activity (IC50 = 94 ± 4.1 nM). Chlorpromazine, thioridazine, prochlorperazine, and perphenazine were devoid of inverse agonist activity at concentrations ranging from 1 nM to 10 μM. Prochlorperazine behaved as a partial agonist with an EC50 of 270 nM (Fig. 3B) and intrinsic activity of 0.8 compared with 20 nM 5-HT (data not shown). Eight additional typical antipsychotic drugs were tested for inverse agonist activity at a single concentration of 10 μM (Fig.4). None of these drugs displayed inverse agonist activity.

Effect of typical antipsychotic drugs on 5-HT2C receptor basal activity. [3H]IP production was measured in COS-7 cells expressing human 5-HT2C-INI receptors. Cells were treated with or without 10 μM typical antipsychotic. Basal activity was determined by subtracting [3H]IP produced in cells transfected with vector only. Data represent the mean ± S.E. of three independent experiments. *P < .01 versus basal.
Radioligand binding studies were performed to ensure that the typical antipsychotic drugs were capable of binding to 5-HT2C receptors at the concentrations tested in the inverse agonist assay. Table 2 listsKi values for inhibition of 1 nM [3H]mesulergine binding to human 5-HT2C-INI receptors. Chlorpromazine, loxapine, and thioridazine had the highest affinities withKi values of 6.1, 17, and 46 nM, respectively. Five of the typical antipsychotic drugs had moderate to high affinity (Ki < 150 nM) for human 5-HT2C-INI receptors. With the exception of haloperidol, the rest of the drugs displayedKi values ranging from 150 nM to 2 μM. These results indicate that the typical antipsychotic drugs were occupying >90% of the available receptors when tested at 10 μM in the inverse agonist assay.
Binding affinity of typical antipsychotic drugs at human 5-HT2C-INI receptors
To confirm these results, an additional experiment was performed to demonstrate functional antagonism. Chlorpromazine, thioridazine, spiperone, and thiothixene were tested for the ability to block 5-HT-stimulated IP production in COS-7 cells transfected with human 5-HT2C-INI receptors (Fig.5). Transfected cells were pretreated for 10 min with 10 μM typical antipsychotic drug, followed by a 35-min challenge with 20 nM 5-HT. 5-HT stimulated IP production 2-fold over basal levels. All four drugs produced >50% inhibition of 5-HT-stimulated IP production.

Inhibition of 5-HT-stimulated [3H]IP production by typical antipsychotic drugs. COS-7 cells expressing human 5-HT2C-INIreceptors were pretreated ± 10 μM drug for 10 min followed by a 35-min challenge with 20 nM 5-HT ± 10 μM drug. Basal activity was measured subtracting [3H]IP produced in cells transfected with vector only. Data represent the mean ± S.E. of three independent experiments.+P < .01 versus basal; *P < .01 versus 5-HT.
Because the previous experiments failed to detect inverse agonist activity of typical antipsychotic drugs, yet demonstrated functional antagonism of 5-HT-stimulated IP production, additional experiments were performed to determine whether typical antipsychotics behave as neutral antagonists at human 5-HT2C-INIreceptors. Five typical antipsychotic drugs (representing four different chemical classes of drugs used in this study) were tested for the ability to reverse clozapine-induced inhibition of 5-HT2C receptor basal activity (Fig.6). Transfected cells were pretreated with 10 μM typical antipsychotic drug followed by challenge with 200 nM clozapine. Chlorpromazine, thioridazine, and thiothixene reversed clozapine inverse agonism by returning IP production back to basal levels. Pimozide produced >60% inhibition, and spiperone produced >35% inhibition of clozapine inverse agonism.

Inhibition of clozapine inverse agonism by typical antipsychotic drugs. COS-7 cells expressing human 5-HT2C-INI receptors were pretreated ± 10 μM typical antipsychotic drug for 10 min followed by a 35-min challenge with 200 nM clozapine ± 10 μM typical antipsychotic drug. Basal activity was measured by subtracting [3H]IP produced in cells transfected with vector only. % inhibition of clozapine inverse agonism: chlorpromazine, 85 ± 5.0; thioridazine, 100 ± 4.3; spiperone, 37 ± 1.4; pimozide, 63 ± 6.8; thiothixene, 100 ± 4.4. Data represent the mean ± S.E. of three independent experiments. +P < .01 versus basal; *P < .01 versus clozapine.
Discussion
The mechanisms responsible for the superior clinical effects of atypical antipsychotic drugs such as clozapine are not known. Popular hypotheses involve the ability of these drugs to inhibit serotonergic systems in the frontal cortex through blockade of serotonin receptors (Lieberman et al., 1998; Meltzer, 1999). Although atypical antipsychotic drugs display high affinity for 5-HT2A, 5-HT2C, 5-HT6, and 5-HT7 receptors (Roth et al., 1994), most of the research has focused on the role of 5-HT2A and/or 5-HT2Creceptors. These receptors are expressed in the frontal cortex and have been demonstrated to alter cortical function on stimulation with agonists or blockade with antagonists (Teitler et al., 1988; Hoffman and Mezey, 1989; Kennett et al., 1994; Jakab and Goldman-Rakic, 1998;Marek and Aghajanian, 1998; Millan et al., 1998). Recently we demonstrated that 5-HT2A and 5-HT2C receptors are rendered constitutively active by site-specific mutagenesis, involving the alteration of one critical amino acid (Herrick-Davis et al., 1997; Egan et al., 1998). Antipsychotic drugs, which were previously classified as competitive antagonists at these receptors, were found to be inverse agonists at the 5-HT2A receptor: both typical and atypical antipsychotic drugs reversed the constitutive activity of the CAM C322K 5-HT2A receptor (Egan et al., 1998). In contrast to the 5-HT2A receptor, the 5-HT2C receptor exists in multiple isoforms due to RNA editing (Burns et al., 1997). The different isoforms of the native receptor display different levels of constitutive activity (Herrick-Davis et al., 1999) and are widely distributed throughout the brain (Niswender et al., 1999). These results suggest that constitutive activity of native 5-HT2C receptors may occur in vivo.
The production of CAM forms of G-protein-coupled receptors (GPCR), as well as the observation of constitutive activity in native forms of GPCR, has resulted in a revised model for the molecular events coupling GPCR to their respective GTP-binding proteins (Samama et al., 1993). In the revised model the receptor exists in equilibrium between an inactive and active conformation. The observation of constitutive activity in native forms of GPCR is consistent with the revised model where the equilibrium is shifted to favor the activated state in the absence of activating ligand. In addition, the revised model predicts the existence of inverse agonist activity of drugs, a property not predicted by the classical model. Inverse agonists shift the equilibrium of GPCR to favor the inactive form of the receptor. Many drugs thought to be classical antagonists (neutral antagonists) display this property when a GPCR is stabilized in a CAM form by mutagenesis or when the native form of the receptor displays constitutive activity. In theory, drugs displaying inverse agonist activity at a receptor may have different effects from neutral antagonists at the same receptor. Several examples of diseases produced by constitutive activation of GPCR have been discovered (Parma et al., 1993; Rao et al., 1994). Inverse agonists for these receptors represent possible therapeutic avenues.
In the present study, we investigated the effects of antipsychotic drugs at rat and human 5-HT2C receptors to determine whether there is a relationship between the classification of the antipsychotic drug (atypical versus typical) and inverse agonist activity at 5-HT2C receptors. As shown in Figs. 1and 2, atypical antipsychotic drugs are robust inverse agonists at both rat and human 5-HT2C receptors. The results with the native human 5-HT2C-INI receptor are especially intriguing. Given that 5-HT2C receptor isoforms are constitutively active (Herrick-Davis et al., 1999) and are widely distributed throughout the brain (Fitzgerald et al., 1999;Niswender et al., 1999), 5-HT2C inverse agonists may produce unique functional effects in vivo.
Many of the clinically proven atypical antipsychotic drugs developed to date have been demonstrated to have high affinity for 5-HT2C receptors. However, previous studies have reported a poor correlation between 5-HT2Creceptor affinity and atypical antipsychotic drug classification (Roth et al., 1992). These results were based on the observation that two typical antipsychotic drugs, chlorpromazine and thioridazine, have high affinity for 5-HT2C receptors, whereas two putative atypical antipsychotic drugs, amperozide and melperone, have low affinity for 5-HT2C receptors. Although amperozide has a 5-HT2A/D2affinity ratio predictive of atypical antipsychotic activity, clinical trails demonstrating atypical antipsychotic activity of amperozide and melperone are lacking. However, if these drugs are proven to be atypical antipsychotics in clinical trials, it is unlikely that their mechanism of action would include 5-HT2Creceptor-mediated mechanisms.
Although high affinity for 5-HT2C receptors in and of itself may not be a good predictor of atypical versus typical antipsychotic activity, the results of our study demonstrate that all of the atypical antipsychotic drugs tested display inverse agonist activity. Seven of these drugs had Kivalues <50 nM for 5-HT2C-INI receptors and two drugs had Ki values between 50 and 200 nM. With the exception of loxapine, typical antipsychotic drugs were devoid of inverse agonist activity, even though two drugs (chlorpromazine, thioridazine) had high affinity (Ki < 50 nM) and three drugs (prochlorperazine, perphenazine, mesoridazine) hadKi values between 100 and 200 nM for 5-HT2C-INI receptors. Several typical antipsychotic drugs (chlorpromazine, thioridazine, spiperone, pimozide, thiothixene) were identified as neutral antagonists based on their ability to block 5-HT-stimulated PI hydrolysis and to inhibit clozapine's inverse agonist activity.
Several studies have called into question the classification of loxapine as a “typical” antipsychotic (Glazer, 1999; Meltzer and Jayathilake, 1999). A commonly used definition of an “atypical” antipsychotic is one that produces minimal extrapyramidal symptoms at doses producing effective antipsychotic activity (Meltzer and McGurk, 1999). By this definition, loxapine can't be classified as atypical, because it produces extrapyramidal symptoms. However, loxapine is different from all of the other typical antipsychotics tested in the present study. Loxapine has a 4-fold higher affinity for 5-HT2A receptors than D2 receptors, more reminiscent of atypical antipsychotic drugs (Roth et al., 1994). Loxapine has been reported to improve negative symptoms of schizophrenia (Glazer 1999; Meltzer and Jayathilake, 1999). Loxapine has equally high affinity for 5-HT2C and D2 receptors, whereas other typical antipsychotic drugs have 10- to 100-fold higher affinity for D2than 5-HT2C receptors (Roth et al., 1994). Loxapine is a 5-HT2C receptor inverse agonist, whereas other typical antipsychotics are neutral antagonists (Fig. 4).
Several studies have demonstrated the functional significance of 5-HT2C inverse agonism on second messenger production and receptor expression in recombinant and primary cell lines (Barker et al., 1994; Kuoppamaki et al., 1994; Westphal and Sanders-Bush, 1994; Palvimaki et al., 1998). In choroid plexus cells, chronic treatment with inverse agonist causes 5-HT2C receptor down-regulation, whereas neutral antagonists have no effect (Barker et al., 1994). In vivo, chronic treatment with the 5-HT2C inverse agonist clozapine has been demonstrated to decrease 5-HT2C receptor expression and 5-HT-mediated PI hydrolysis (Kuoppamaki et al., 1994). These results demonstrate that 5-HT2C inverse agonists alter 5-HT2C receptor function and that inverse agonists may differ in function from neutral antagonists. This is an important consideration in light of the present study demonstrating that antipsychotic drugs can be classified as 5-HT2C inverse agonists or neutral antagonists, and this corresponds with previous antipsychotic drug classification as atypical or typical (Meltzer et al., 1989; Roth et al., 1992).
Atypical antipsychotics have been reported to be more effective in treating the negative symptoms of schizophrenia and improving cognitive function than typical antipsychotic drugs (Meltzer and McGurk, 1999). It has been suggested that dopamine deficits in the prefrontal cortex may contribute to the negative symptoms of schizophrenia (Davis et al., 1991; Weinberger and Lipska, 1995). Several studies have demonstrated that atypical antipsychotic drugs preferentially enhance dopamine release in the prefrontal cortex and suggest that this is related to the ability of these drugs to improve cognitive function in patients with schizophrenia (Moghaddam and Bunney, 1990; Kuroki et al., 1999;Youngren et al., 1999). Recent studies have shown that 5-HT2C receptors can modulate the basal firing rate of dopamine neurons and regulate dopamine release. In anesthetized rats, the 5-HT2C receptor antagonist SB-242084 (0.16–0.64 mg/kg, i.v.) caused a dose-dependent increase in the basal firing rate of dopamine neurons in the ventral tegmental area, but not in the substantia nigra (Di Matteo et al., 1999). In a separate study, the 5-HT2C receptor agonist RO 60-0175 (2.5 mg/kg, s.c.) decreased dopamine dialysate levels, while the antagonist SB-242084 (10 mg/kg, i.p.) increased dopamine dialysate levels in the prefrontal cortex of freely moving rats (Millan et al., 1998). In the present study, we demonstrate that 5-HT2C inverse agonism is a property of atypical antipsychotic drugs. Based on these results we suggest that 5-HT2C receptor inverse agonism functions to increase prefrontal cortical dopamine release, and this may represent a possible mechanism through which atypical antipsychotic drugs produce beneficial effects in improving negative symptoms in patients with schizophrenia.
It is unrealistic to expect that all atypical antipsychotic drugs will have one common mechanism of action because of their widely varied receptor binding profiles. For the drugs included in the present study, 5-HT2C receptor inverse agonism should be investigated as a possible mechanism of action. Additional studies are required to test the hypothesis that 5-HT2Creceptor inverse agonism plays a role in the clinical efficacy of atypical antipsychotics.
Footnotes
-
Send reprint requests to: Katharine Herrick-Davis, Center for Neuropharmacology and Neuroscience, MC-136, Albany Medical College, 47 New Scotland Ave., Albany, NY 12208. E-mail: daviskh{at}mail.amc.edu
-
↵1 Supported by United State Public Health Services Grants MH-57019 (to K.H.-D.) and MH-56650 (to M.T.).
- Abbreviations:
- 5-HT
- 5-hydroxytryptamine
- CAM
- constitutively active mutant
- GPCR
- G-protein-coupled receptor
- DMEM
- Dulbecco's modified Eagle's medium
- IP
- inositol phosphate
- Received April 21, 2000.
- Accepted June 29, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}