


Abstract
Transport of new quinolone antibacterial agents (quinolones) at the blood-brain barrier (BBB) was studied in vitro by using immortalized rat brain capillary endothelial cells RBEC1, and in vivo by using the brain perfusion method in rats and multidrug-resistantmdr1a/1b gene-deficient mice. The permeability coefficient of grepafloxacin measured by brain perfusion was increased by an excess of unlabeled grepafloxacin, suggesting a participation of a saturable BBB efflux system. Uptake coefficients of [14C]grepafloxacin, [14C]sparfloxacin, and [14C]levofloxacin by RBEC1 cells at the steady state were increased in the presence of the unlabeled quinolones. The steady-state uptake of [14C]grepafloxacin was increased in the presence of various quinolones. Brain distributions of [14C]grepafloxacin and [14C]sparfloxacin evaluated in terms of the brain-to-plasma free concentration ratio inmdr1a/1b gene-deficient mice were significantly higher than those in wild-type mice, demonstrating an involvement of P-glycoprotein as the efflux transporter. Anionic compounds, including 4,4′-diisothiocyanatostilbene-2,2′-disulfonic acid (DIDS) and genistein, increased the steady-state uptake of [14C]grepafloxacin by RBEC1 cells. Because [14C]grepafloxacin was transported by multidrug resistance-associated protein (MRP), in MRP1-overexpressing cells and because RBEC1 and primary cultured brain capillary endothelial cells expressed MRP1, this protein may be an additional efflux transporter for quinolones. Furthermore, the permeability coefficient of [14C]grepafloxacin across the BBB was increased by DIDS or in the absence of bicarbonate ions in the brain perfusion method. DIDS or bicarbonate ion did not affect MRP1 function. Accordingly, the brain distribution of quinolones is restricted by the action of multiple efflux transporters, including P-glycoprotein, MRP1, and an unknown anion exchange transporter.
New quinolone antibacterial agents (quinolones) are well absorbed from the gastrointestinal tract and are distributed to tissues with high distribution volume (Sörgel et al., 1989). In the brain distribution of quinolones, lipophilicity may be an important factor (Jaehde et al., 1993), but some quinolone derivatives exhibit low distribution to the brain compared with other peripheral tissues (Murata et al., 1995; Turnidge, 1999). Because clinically used drugs, including quinolones, may exhibit central nervous system toxicity, it is important to elucidate the mechanisms of brain distribution of such drugs, especially quinolones, which may cause convulsion by competing with γ-aminobutyric acid binding to its receptor (Tsuji et al., 1988a,b). Several studies on brain distribution of quinolones have demonstrated that brain-to-blood elimination across the blood-cerebrospinal fluid barrier includes an active transport process (Jaehde et al., 1992; Ooie et al., 1996a,b). However, efflux across the blood-brain barrier (BBB) may be important for quinolones, rather than the transport across the blood-cerebrospinal fluid barrier (Ooie et al., 1997a). Furthermore, by using brain microdialysis method, it was demonstrated that the concentrations of several quinolones were significantly lower than those in serum and cerebrospinal fluid, demonstrating that they were effluxed out of brain across the BBB (Ooie et al., 1997b). We and others have already reported an involvement of the efflux transporter P-glycoprotein, which transfers drugs from the brain to the bloodstream, thereby decreasing the apparent distribution of various drugs in brain tissue (Tatsuta et al., 1992; Tsuji et al., 1992; Schinkel et al., 1994; Tamai and Tsuji, 1996). In addition to P-glycoprotein, other efflux systems, which transport anionic compounds, also may be active at the BBB (Deguchi et al., 1997; Kakee et al., 1997; Morrison et al., 1999). One such transporter is multidrug resistance-associated protein 1 (MRP1), which is thought to be expressed in the brain (Huai-Yun et al., 1998; Kusuhara et al., 1998;Regina et al., 1998). Because quinolones such as grepafloxacin and HSR-903 and their conjugated metabolites are transported by MRP2, it is likely that these quinolones also are transported by its isoform MRP1 at the BBB (Murata et al., 1998; Sasabe et al., 1998). Accordingly, such efflux transporters may explain low brain accumulation of quinolones.
We have recently found that the new quinolone antibacterial agent HSR-903 may be transported from the brain via multiple pathways, such as P-glycoprotein and anion transporters that are present at the brain capillary endothelial cells (BCECs) that form the BBB (Murata et al., 1999). However, the mechanisms other than P-glycoprotein have not been well elucidated. Grepafloxacin is another quinolone that exhibits similar pharmacokinetic behavior to HSR-903, including tissue distribution and elimination pathways. Accordingly, multiple transporters may be involved in the BBB transport of various quinolones.
In the present study, we examined the brain distribution of several quinolones by using various in vivo and in vitro methods to elucidate the mechanisms that are commonly involved in their efflux transport. The structures of the drugs investigated in this study are shown in Fig. 1.

Chemical structures of grepafloxacin (A), sparfloxacin (B), and levofloxacin (C).
Materials and Methods
Chemicals.
[14C]Grepafloxacin, [quinoline-2-14C]OPC-106 (81.2 μCi/mg); [14C]levofloxacin, (S)-(−)-[pyridone ring-2-14C]ofloxacin (53 mCi/mmol); and grepafloxacin were kindly supplied by Otsuka Seiyaku Co., Ltd. (Tokyo, Japan). Other unlabeled quinolones were obtained from Hokuriku Seiyaku Co., Ltd. (Fukui, Japan). Sparfloxacin, [2-14C]- (9 mCi/mmol), and methotrexate, disodium salt, [3′,5′,7-3H(N)]- (30.3 Ci/mmol), were purchased from Moravek Biochemicals, Inc. (Costa Mesa, CA). [6,6′(n)-3H]Sucrose was purchased from Amersham Pharmacia Biotech (Buckinghamshire, UK). All other reagents were commercial products of reagent grade.
In Situ Brain Perfusion Study.
All animal experiments were performed according to the Guidelines for the Care and Use of Laboratory Animals at the Takara-machi Campus of Kanazawa University and were approved by the Committee of the Ethics of Animal Experimentation of Kanazawa University, Takara-machi Campus. Male Sprague-Dawley rats (210–300 g) were purchased from Charles River Japan, Inc. (Kanagawa, Japan). Brain perfusion was performed by the method reported previously (Takasato et al., 1984). In brief, the rats were anesthetized and the right carotid artery was catheterized with polyethylene tubing (SP-10) filled with sodium heparin (100 I.U./ml). The perfusate (bicarbonate-buffered physiological saline, 122 mM NaCl, 3 mM KCl, 25 mM NaHCO3, 1.2 mM MgSO4, 0.4 mM K2HPO4, 1.4 mM CaCl2, 10 mM d-glucose, and 10 mM HEPES, pH 7.4) containing [14C]grepafloxacin and [3H]sucrose, which was used as a brain intravascular volume marker, was perfused through the catheter at the rate of 4.98 ml/min by an infusion pump (Harvard Apparatus, South Natick, MA). At the end of a 30-s perfusion, the rat was decapitated, and the right cerebral hemisphere was dissected from the perfused brain and weighed. The perfused right cerebral hemisphere was solubilized in Soluene-350 (Packard, Meriden, CT) at 60°C for 3 h. The radioactivity was measured by liquid scintillation counting. In vivo BBB permeability (μl/min/g of brain) was calculated as described previously (Tamai et al., 1995) after correcting for the remaining intravascular grepafloxacin, estimated from the apparent brain uptake of [3H]sucrose.
In Vivo Brain Distribution Study in Mice.
FVB/NJ andmdr1a/1b gene-deficient mice were purchased from Jackson Laboratories (Bar Harbor, ME) and Taconic Farms, Inc. (Germantown, NY), respectively. 14C-Labeled quinolones (1 μCi/head) dissolved in saline solution in a total volume of 50 μl were i.v. injected. At 2 h after dosing, the mice were sacrificed under ether anesthesia and immediately dissected. The whole brain was isolated, weighed, and solubilized in Soluene-350 at 60°C for 3 h. Plasma and brain-associated radioactivity were measured by liquid scintillation counting. Plasma unbound fraction was determined by ultrafiltration of plasma by using a Microcon YM-10 (Millipore Co., Bedford, MA).
Transport Study in Immortalized Rat BCECs (Rat Brain Capillary Endothelial Cells, RBEC1).
Immortalized rat BCECs (RBEC1) were established by transfection of recombinant plasmids containing origin-defective simian virus 40 gene, SVori-8-16, into primary cultured rat BCECs as described previously (Kido et al., 2000). RBEC1 cells were cultivated on multiwell dishes and cultured at 37°C in a 5% CO2, 95% air atmosphere. When the cells reached confluence, they were washed three times with 1 ml of incubation solution (122 mM NaCl, 3 mM KCl, 25 mM NaHCO3, 1.2 mM MgSO4, 0.4 mM K2HPO4, 1.4 mM CaCl2, 10 mM d-glucose, and 10 mM HEPES, pH 7.4) and preincubated at 37°C for 20 min. After the preincubation, test solution (0.25 ml) containing a radiolabeled test compound was added to initiate uptake. The cells were incubated at 37°C for the designated time, and then washed three times with 1 ml of ice-cold incubation solution to terminate the uptake. The cells were solubilized with 1 N NaOH for 120 min, and then neutralized with HCl, and the radioactivity was determined. Cellular protein content was measured by the method of Lowry et al. (1951) by using BSA as a standard. Net uptake was expressed as the cell-to-medium ratio (μl/mg of protein) obtained by dividing the uptake amount by the concentration of substrate in the incubation medium. To prepare NaHCO3-free medium, NaHCO3was replaced with sodium gluconate.
Transport Studies in HL60 and MDR HL60/ADM Cells.
Human leukemic HL60/ADM cells that overexpress MRP1 (Wada et al., 1999) and the control parent HL60 cells were cultured in RPMI-1640 medium, supplemented with 10% heat-inactivated fetal calf serum, 2 mM glutamine, 100 U/ml penicillin, and 5 μg/ml gentamicin at 37°C under 95% air, 5% CO2. The cultured cells were suspended in incubation solution as described above, and centrifuged at 700g for 5 min. The resultant pellets were suspended in incubation solution. The cell suspension was preincubated for 20 min and then mixed with incubation solution containing a radiolabeled test compound to initiate uptake. At appropriate times, the reaction was terminated by separating the cells from the medium by means of a centrifugal filtration technique. The radioactivities of the supernatant and the cell pellet were determined. Cellular protein content was determined according to the method of Bradford by using a Bio-Rad protein assay kit (Bradford, 1976). Net uptake of substrate was expressed as the cell-to-medium ratio (μl/mg of protein), as described above.
Detection of MRP1 by Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Method.
Primary cultured rat BCECs were isolated from cerebrum of male Wistar rats (SLC, Shizuoka, Japan), 4 weeks old, as described previously (Ichikawa et al., 1996). Total RNA was isolated from primary cultured BCECs and RBEC1 cells by means of the standard method with acid guanidinium-phenol-chloroform. Poly(A)+ RNA was purified by using Oligotex dT 30 (Takara, Ootsu, Japan). RT was performed at 42°C for 1 h with 1 μg of Poly(A)+ RNA, 0.05 μg of random primer, 0.125 μg of RNase Guard (Life Technologies, Inc., Gaithersburg, MD), and 100 U of Super Script II (Life Technologies, Inc.) in a 50-μl reaction volume. For PCR amplification, the primer sequences and the expected size of PCR product for rat MRP1 were as follows: sense 5′-CTGGCTTGGTGTGAACTGAT-3′, antisense 5′-AGGCTCTGGCTTGGCTCTAT-3′, 394 base pairs. PCR amplification of cDNA was done at 94°C for 30 s, 62°C for 90 s, and 72°C for 90 s through a total of 30 cycles. The PCR products were separated by electrophoresis in 1.5% agarose and visualized under UV light in the presence of ethidium bromide.
Results
BBB Permeability of Grepafloxacin in Rats by Brain Perfusion Method.
The BBB permeability of grepafloxacin was determined by using an in situ brain perfusion technique in rats and the results are shown in Table 1. The BBB permeability coefficient of [14C]grepafloxacin at a tracer concentration (0.78 μM) was 11.9 μl/min/g of brain after subtraction of the value of [3H]sucrose, which was used as a BBB-impermeable brain vascular marker. The obtained value is close to that of sucrose (26.0 μl/min/g of brain), demonstrating a low permeability in spite of a sufficient lipophilicity (octanol/water partition coefficient of 3.7 at neutral pH). When unlabeled grepafloxacin was added to the perfusion solution at 2 mM, the permeability coefficient was increased 6.2-fold, whereas nonspecific BBB permeability was not affected in the presence of such a high concentration of grepafloxacin, as assessed in terms of the [3H]sucrose permeability coefficient. The effect of unlabeled grepafloxacin may be ascribed to the decrease of efflux by saturation of the specific efflux mechanism, the increase of influx by allosteric effect, or others. However, the observed low brain distribution suggests a significance of an efflux transport from brain to blood across the BBB.
Effect of unlabeled grepafloxacin, DIDS, and bicarbonate ion on BBB permeability of [14C]grepafloxacin and [3H]sucrose by in situ brain perfusion method
Uptake of Quinolones by RBEC1 Cells.
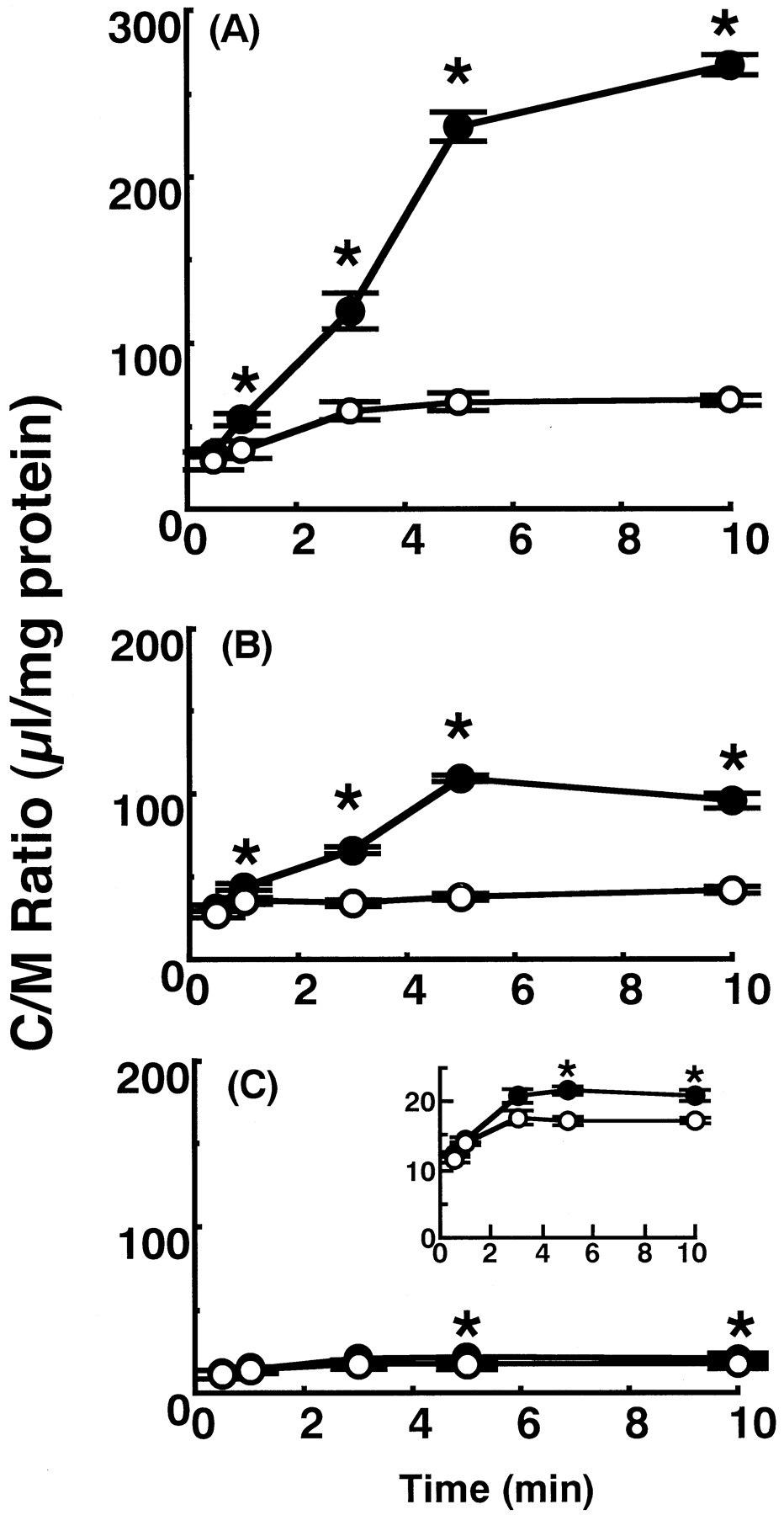
The participation of specialized transport mechanisms was suggested by the in situ brain perfusion study in Table 1 and it is important to clarify the mechanisms involved. However, brain perfusion technique is not useful for the mechanistic characterization, therefore, the in vitro BBB transport experiments were performed in the subsequent studies. As in the in vitro method, immortalized BCECs established in our laboratory were used (Kido et al., 2000). Furthermore, it is important to elucidate the structure-transport relationship among quinolones. So, various quinolones were used as the substrates and/or competitive agents to the transport of grepafloxacin. Figure2 shows the time course of uptake for three quinolones, [14C]grepafloxacin, [14C]sparfloxacin, and [14C]levofloxacin, by immortalized RBEC1 cells. Their concentrations were 1.37, 4, and 0.7 μM, respectively, based on the available specific radioactivity of each compound. The uptake of each quinolone reached a steady state within 5 min and in each case, the steady-state uptake was increased by the addition of the unlabeled quinolone at 5 mM, whereas the initial uptake rate was not affected. The highest increase on addition of unlabeled compound was observed with grepafloxacin. The increase in the uptake of sparfloxacin was intermediate and the extent of increase was slight but significant in the case of levofloxacin. The observed increase of the steady-state uptake represents the participation of efflux transporters in RBEC1 cells, although the direct effect on the influx of grepafloxacin cannot be excluded.
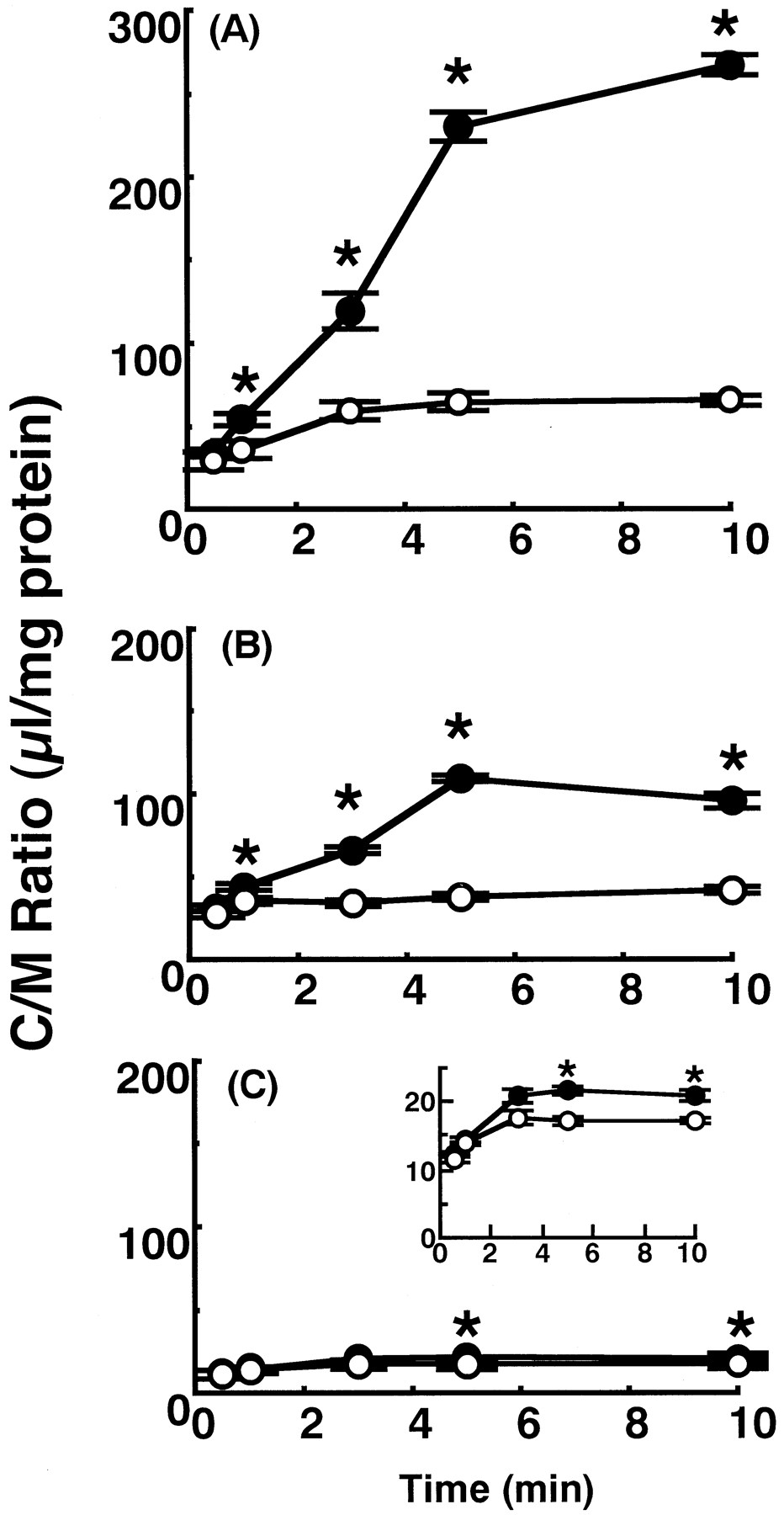
Time courses of uptake of radiolabeled quinolones by RBEC1 cells. RBEC1 cells were preincubated at 37°C for 20 min. Uptake of 14C-labeled quinolones was measured at pH 7.4 and 37°C in the absence (○) or presence (●) of 5 mM unlabeled compound. Each value represents the mean ± S.E. of three to four experiments. A, [14C]grepafloxacin (1.37 μM). B, [14C]sparfloxacin (4 μM). C, [14C]levofloxacin (0.7 μM). The inset of C magnified the effect of unlabeled levofloxacin. ∗, significantly different from the control (P < .05).
Table 2 (left) shows the effect of quinolone antibacterial agents on the steady-state (5 min) uptake of [14C]grepafloxacin. The uptake of [14C]grepafloxacin at 1.4 μM was increased greatly in the presence of unlabeled grepafloxacin; moderately by sparfloxacin, norfloxacin, and enoxacin; and slightly by nalidixic acid, whereas levofloxacin and ofloxacin had no significant effect at 2 mM. So, there appears to be a common efflux mechanism for most of the quinolone derivatives, although ofloxacin and levofloxacin, which is the S-isomer of racemic ofloxacin, may have low or no affinity for the efflux transporter.
Effect of various quinolones on steady-state uptake of [14C]grepafloxacin by RBEC1
Table 3 shows the effect of various anionic compounds on the steady-state uptake of [14C]grepafloxacin by RBEC1 cells. The uptake of [14C]grepafloxacin was increased in the presence of the anion exchange inhibitor 4,4′-diisothiocyanatostilbene-2,2′-disulfonic acid (DIDS, 1 mM); the anion transport inhibitors genistein (0.2 mM), indomethacin (0.1 mM), sulfobromophthalein (BSP, 0.1 mM), and sulfinpyrazone (1 mM); the anionic drug methotrexate (2 mM); and the physiological anion taurocholic acid (2 mM) in the presence of bicarbonate ion in the uptake medium. These results suggested that an anion transporter may be involved in the efflux of grepafloxacin and other quinolones in RBEC1 cells.
Effect of various compounds on steady-state uptake of [14C]grepafloxacin by RBEC1
Effect of Bicarbonate Ion on Uptake of Quinolones by RBEC1 Cells.
Figure 3 shows the effect of external bicarbonate ion on the uptake of [14C]grepafloxacin (1.4 μM) by RBEC1 cells. When bicarbonate ion (25 mM) in the uptake medium was replaced by gluconate ion, the steady-state uptake was significantly increased, although no such effect was observed over the initial 1 min. When DIDS was included in the uptake medium at a concentration of 1 mM, the uptake of [14C]grepafloxacin was significantly increased in the presence of bicarbonate ion, whereas no effect of DIDS was observed in the absence of bicarbonate ion. So, the results suggested that DIDS-sensitive efflux can be ascribed to grepafloxacin/bicarbonate ion exchange transport. Table 2 (right) shows the effect of several quinolones on the steady-state uptake of [14C]grepafloxacin by RBEC1 cells in the absence of bicarbonate ion. The extent of increment of uptake of [14C]grepafloxacin in the presence of each quinolone was decreased in the absence of bicarbonate ion compared with that in the presence of bicarbonate ion. Similarly, the increments by anionic compounds such as BSP, genistein, and DIDS also were decreased to 119 ± 4.09, 102 ± 3.04, and 101 ± 4.94% of the control, respectively, in the absence of bicarbonate ion compared with the results in the presence of bicarbonate ion (Table 3). Only grepafloxacin (2 mM) and BSP (0.1 mM) showed a significant increase of [14C]grepafloxacin uptake in the absence of bicarbonate ion.

Effect of DIDS and bicarbonate ions on [14C]grepafloxacin uptake by RBEC1 cells. A, RBEC1 cells were preincubated at 37°C for 20 min in the presence (○) or absence of HCO3− by replacing HCO3− with gluconate ions (●) and the uptake of [14C]grepafloxacin (1.37 μM) was measured at pH 7.4 and 37°C. Each value represents the mean ± S.E. of four experiments. ∗, significantly different from the control shown by open circles (P < .05). B, uptake of [14C]grepafloxacin (1.37 μM) by RBEC1 cells was measured at 5 min in the presence or absence of DIDS and HCO3−. Each condition of the uptake was shown in the lower part of the figure. ∗, significantly different from the control study, which was shown by open column (P < .05).
Effect of Bicarbonate Ion and DIDS on Permeability Coefficient of Grepafloxacin by In Situ Brain Perfusion Method.
The effects of bicarbonate ion and DIDS on the in vivo BBB transport of [14C]grepafloxacin were examined by in situ brain perfusion method. As shown in Table 1, when bicarbonate ion was replaced with gluconate in the perfusion solution, the permeability coefficient of [14C]grepafloxacin (0.78 μM) was increased significantly to 31.6 μl/min/g of brain. In the presence of bicarbonate ion, DIDS (1 mM) also significantly increased the permeability coefficient of [14C]grepafloxacin to 24.7 μl/min/g of brain, whereas no further increase of the permeability coefficient of [14C]grepafloxacin by DIDS was observed in the absence of bicarbonate ion (39.3 μl/min/g of brain). So, the effects of DIDS and bicarbonate ion may be mediated by the same mechanism.
Brain Distribution of Quinolones in mdr1a/1bGene-Deficient Mice.
To evaluate the involvement of the multidrug efflux transporter P-glycoprotein in the transport of the quinolones, the brain distributions of [14C]grepafloxacin, [14C]sparfloxacin, and [14C]levofloxacin were measured by using P-glycoprotein-deficient mdr1a/1b-gene(−/−) mice and compared with those in wild FVB mice. Two hours after i.v. bolus administration of a tracer amount of quinolone into mice, the brain-to-plasma free concentration ratio (Kp,f) was determined (Table4). The Kp,f values inmdr1a/1b(−/−) mice were increased 3- and 4-fold for grepafloxacin and sparfloxacin, respectively, compared with those in FVB mice, whereas no significant change in the Kp,f for levofloxacin was observed. No significant change was observed in the plasma unbound fraction of each quinolone between wild and mdr1a/1bgene-deficient mice (data not shown). Thus, the change could be directly ascribed to the effect of P-glycoprotein-mediated efflux transport. Herein, because the total radioactivity was measured to evaluate Kp,f, the observation may partially include the effect of metabolic products of quinolones, although the extent of metabolites should be much less than that of the parent compounds in plasma (Sasabe et al., 1998).
Comparison of brain distribution of various quinolones inmdr1a/1b ( +/+ ) and mdr1a/1b(−/−) mice
Uptake of Grepafloxacin by MRP1-Overexpressing HL60/ADM and Parental HL60 Cells.
The possible involvement of MRP1, which has been suggested to be expressed at the BBB, in the brain efflux transport of quinolones was examined by using MRP1-overexpressing HL60/ADM cells. The uptake of [14C]grepafloxacin (1.4 μM) by HL60/ADM cells at the steady state (30 min) was significantly less (117 ± 4.9 μl/mg of protein) than that by parental drug-sensitive HL60 cells (190 ± 3.8 μl/mg of protein). Furthermore, uptake of [14C]grepafloxacin by HL60/ADM cells was not increased by replacing bicarbonate ion with gluconate ion, and was reduced in the presence of 1 mM DIDS. These observations demonstrate that grepafloxacin is a substrate for human MRP1, but the MRP1-mediated transport of grepafloxacin is not DIDS or bicarbonate ion sensitive.
Detection of MRP1 Expression in RBEC1 Cells by RT-PCR Method.
To confirm the involvement of MRP1, we examined the expression of MRP1 in RBEC1 and primary cultured BCECs by the RT-PCR method. As shown in Fig. 4, the PCR product that corresponds to MRP1 was observed in both the immortalized RBEC1 cells and primary cultured BCECs. Furthermore, we measured the uptake of an MRP1 substrate, [3H]methotrexate (Heijn et al., 1997), by RBEC1 cells (data not shown). The uptake of [3H]methotrexate was increased by depleting ATP by preincubating the cells with metabolic inhibitors from 30.6 ± 2.1 μl/mg of protein (n = 4) to 54.3 ± 1.8 μl/mg of protein at 60 min. So, RBEC1 cells may be useful to evaluate MRP1-mediated transport of drugs. However, RBEC1 cells were not adequate to examine P-glycoprotein-mediated transport because it was very difficult to measure the increase in uptake of [14C]grepafloxacin or [3H]cyclosporin A, a typical substrate of P-glycoprotein, in response to P-glycoprotein inhibitors such as unlabeled cyclosporin A, quinidine, or ATP depletors (data not shown).

Detection of MRP1 expression by the RT-PCR method in RBEC1 (RBEC1) and primary cultured BCECs (primary). Total RNA from cultured cells was subjected to RT-PCR by using primer pairs specific for the rat MRP1 gene. The RT-PCR products were analyzed by agarose gel electrophoresis. Bands were stained with ethidium bromide and visualized under UV light. DNA size marker is shown in left lane.
Discussion
In the present study, we examined the mechanism involved in the limited distribution of several quinolones, especially grepafloxacin as the typical model derivative, into the brain, by using in vitro and in vivo experimental methods. First, the involvement of efflux transport of grepafloxacin and other quinolones was examined in vivo by using a brain perfusion method and in vitro by using an immortalized cultured cell line RBEC1 derived from RBECs (Kido et al., 2000). The permeability coefficient of [14C]grepafloxacin in vivo was low (11.9 μl/min/g of brain) at tracer concentration (0.78 μM), whereas it was increased to 74.3 μl/min/g of brain by the addition of 2 mM unlabeled grepafloxacin without any effect on the vascular volume, which was assessed by measuring the apparent [3H]sucrose permeability. The increase of permeability coefficient in the presence of 2 mM grepafloxacin (6.2-fold) may be explained by the saturation of efflux transport from the BCECs, although the involvement of allosteric effect and/or trans-stimulation on the influx of radiolabeled grepafloxacin by the unlabeled compound cannot be completely excluded. A similar nonlinear increase in BBB permeability was observed with quinolone antibacterial agent, HSR-903 at higher concentrations of 20 and 40 mM (Murata et al., 1999). Furthermore, efflux transport activity also was demonstrated in in vitro uptake studies with RBEC1 cells. Namely, the initial uptakes of grepafloxacin, sparfloxacin, and levofloxacin were not affected by the addition of excess concentrations of unlabeled compound, whereas their steady-state uptakes were significantly increased; the maximal effect was seen with grepafloxacin and the smallest effect with levofloxacin. Sparfloxacin exhibited an intermediate but significant increase. These results strongly indicate that efflux transport limits the brain distribution. Because the uptake of grepafloxacin by RBEC1 was increased in the presence of various quinolones, the saturable efflux transporter may act commonly on the quinolones.
Second, we examined the mechanisms involved in the efflux of the quinolones from the brain. Because the participation of P-glycoprotein in the brain efflux of HSR-903 has already been demonstrated by usingmdr1a(−/−) mice (Murata et al., 1999), we used the same method for three quinolones to demonstrate directly the involvement of P-glycoprotein as an efflux transporter at the BBB. However,mdr1a(−/−) mice were suggested to have increased expression of mdr1b gene (Schinkel et al., 1994) and a cultured cell model of the mouse BBB, MBEC4 cell line expressesmdr1b but not mdr1a gene (Tatsuta et al., 1992). Therefore, in the present study we used mdr1a/1b-deficient mice. The Kp,f values were significantly higher than those in wild FVB mice for grepafloxacin and sparfloxacin, but not for levofloxacin. Because no significant change in the plasma protein binding between wild FVB and mdr1a/1b gene-deficient mice was observed for any derivative, the increased Kp,f values can be ascribed to decreased efflux transport activity owing to the loss of P-glycoprotein. The absence of a significant effect on the Kp,f values of levofloxacin may be explained by both the lower affinity of this drug for P-glycoprotein and its low passive membrane permeability due to its hydrophilic nature compared with the other derivatives (log P octanol/water = −0.62 for levofloxacin, 0.57 for grepafloxacin, and −0.31 for sparfloxacin).
The immortalized cell line RBEC1 weakly expresses P-glycoprotein, but the expression was too low to allow detection of functional P-glycoprotein because of the down-regulation by cultivation (our unpublished observation). We studied the effects of cyclosporin A and quinidine, as inhibitors of P-glycoprotein, on the steady-state uptake of grepafloxacin by RBEC1 cells, but no significant increase was observed (our unpublished observation). Similarly, uptake of [3H]cyclosporin A was not increased in the presence of unlabeled cyclosporin A or metabolic inhibitors (data not shown), although an increase in the uptake of [3H]cyclosporin A in the presence of these inhibitors was observed in primary cultured bovine BCECs (Sakata et al., 1994). Therefore, it was difficult to confirm the functional involvement of P-glycoprotein by using RBEC1 cells. However, increased steady-state uptake of grepafloxacin was observed in the presence of several anionic compounds, i.e., DIDS, genistein, indomethacin, BSP, sulfinpyrazone, methotrexate, and taurocholic acid (Table 3). This result suggested that an anion-sensitive transporter might be involved in the efflux transport of grepafloxacin in the brain-to-blood efflux direction. We have previously postulated the exchange transport of HSR-903 and bicarbonate ion in primary cultured bovine BCECs (Murata et al., 1999). In that study, addition of bicarbonate ion to the uptake medium decreased the uptake and this could not be explained by the function of P-glycoprotein. In the present study, a similar effect of bicarbonate ion was observed in RBEC1 cells. So, to examine the possible involvement of an efflux transporter other than P-glycoprotein physiologically, we examined the effect of DIDS and bicarbonate ion on the brain distribution of grepafloxacin by the in situ brain perfusion method in rats (Table 1). Addition of DIDS increased the permeability coefficient of grepafloxacin from 11.9 to 24.7 μl/min/g of brain. When bicarbonate ion was replaced with gluconate ion, an increase to 31.6 μl/min/g of brain was observed. No further increase by DIDS in the absence of bicarbonate ion was observed (permeability coefficient of 39.3 μl/min/g of brain). However, addition of 2 mM unlabeled grepafloxacin increased the value to 74.3 μl/min/g of brain. These results suggest that a bicarbonate-dependent and DIDS-sensitive efflux transporter is involved in the efflux of grepafloxacin and unlabeled grepafloxacin inhibited both the anion exchanger and P-glycoprotein-mediated efflux, and probably also MRP, as discussed below. Although DIDS or bicarbonate ion may increase the external or intracellular pH, no significant change in the uptake of [14C]grepafloxacin was observed in RBEC1 cells up to pH 8.0 (data not shown). So, the alteration in the uptake of grepafloxacin by DIDS or bicarbonate ion should be ascribed to a specific effect on the efflux transport but not to a change of pH. This is the first in vivo demonstration of anion-sensitive efflux transport of quinolones at the BBB.
MRP may be expressed at the BBB and transport anionic compounds out of brain (Huai-Yun et al., 1998; Regina et al., 1998). Glucuronidated grepafloxacin and HSR-903 were transported by an MRP isoform, MRP2 (canalicular multispecific organic anion transporter), across the hepatic bile canalicular membranes and the intact forms of the agents also were weakly transported (Murata et al., 1998; Sasabe et al., 1998). So, MRP at the BBB could be another molecular candidate for efflux transport of the quinolones. Expression of mRNA of MRP1 in brain endothelial cells has been reported (Huai-Yun et al., 1998; Regina et al., 1998). So, we examined whether grepafloxacin could be a substrate for MRP1 by using an MRP1-overexpressing cell line, HL60/ADM, by comparing the transport characteristics with its parental cell line HL60 as the control (Wada et al., 1999). Uptake of [14C]grepafloxacin by HL60/ADM cells was significantly less than that by the parent HL60 cells (61.6 ± 2.5% of HL60 cells). Furthermore, uptake of a substrate for MRP1, [3H]vincristine, by RBEC1 cells was increased by metabolic inhibitors and genistein pretreatment (119 ± 0.7% and 143 ± 0.3% of control, respectively), suggesting an involvement of ATP-dependent and anion-sensitive transport. Expression of MRP1 in RBEC1 cells as well as primary cultured rat BCECs was confirmed by the RT-PCR method (Fig. 4). These results suggested that the anion-sensitive efflux transporter for grepafloxacin at the BBB might be MRP1, although other ATP-dependent anion transporters than MRP1, if any, cannot be excluded. Herein, DIDS or bicarbonate ions did not increase the uptake of [14C]grepafloxacin by HL60 or HL60/ADM cells. Accordingly, it is thought that MRP1 may be involved as an efflux transporter for the quinolones, and that a DIDS and bicarbonate ion-sensitive transporter, different from MRP1, also operates at the BBB. So, multiple transporters seem to be functional in the efflux of GPFX from the brain.
In conclusion, in the present study, we demonstrated that the brain distribution of several quinolones is restricted by the operation of multiple efflux transporters, including P-glycoprotein, a DIDS- and bicarbonate ion-sensitive anion exchange transporter and by MRP1.
Footnotes
-
Send reprint requests to: Prof. Akira Tsuji, Department of Pharmacobio-dynamics, Faculty of Pharmaceutical Sciences, Kanazawa University, 13-1 Takara-machi, Kanazawa 920-0934, Japan. E-mail:tsuji{at}kenroku.kanazawa-u.ac.jp
-
↵1 This study was supported in part by a grant-in-aid for scientific research from the Ministry of Education, Science, Sports and Culture and by the Japan Research Foundation for Clinical Pharmacology, Japan.
- Abbreviations:
- BBB
- blood-brain barrier
- MDR
- multidrug-resistant
- MRP1
- multidrug resistance associated protein 1
- BCEC
- brain capillary endothelial cell
- RBEC
- rat brain capillary endothelial cell
- RT-PCR
- reverse transcription-polymerase chain reaction
- DIDS
- 4,4′-diisothiocyanatostilbene-2,2′-disulfonic acid
- BSP
- sulfobromophthalein
- Kp,f
- tissue-to-plasma free concentration ratio
- Received March 16, 2000.
- Accepted May 31, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}