


Abstract
Recently, a polyspecific organic cation transporter, hOCT1, was cloned from human liver. To date, limited studies examining the functional characteristics of the transporter have been performed. The purpose of the present study was to develop a mammalian expression system for hOCT1 and to characterize the interactions of various compounds with the cloned transporter. Lipofection was used to transiently transfect the hOCT1 plasmid DNA in a human cell line, HeLa. We tested the interaction of an array of organic cations and other compounds with hOCT1 by determining Ki values in inhibiting14C-tetraethylammonium (TEA) transport in the transfected cells. Transient expression of hOCT1 activity was observed between 24 and 72 hr post-transfection, with maximal expression at approximately 40 hr. TEA transport was temperature dependent and saturable withVmax and Kmvalues of 2.89 ± 0.448 nmol/mg protein/30 min and 229 ± 78.4 μM, respectively. 14C-TEA uptake in hOCT1 plasmid DNA-transfected HeLa cells was trans-stimulated by unlabeled TEA and 1-methyl-4-phenyl-pyridinium. Organic cations, including clonidine, quinine, quinidine and verapamil (0.1 mM), significantly inhibited 14C-TEA uptake, whereas the organic anion, p-aminohippuric acid (5 mM), did not. The neutral compounds, corticosterone (Ki, 7.0 μM) and midazolam (Ki, 3.7 μM) potently inhibited14C-TEA uptake. The Ki values of several compounds in interacting with hOCT1 differed substantially from the corresponding values for the rat organic cation transporter, rOCT1, and the human kidney-specific organic cation transporter, hOCT2, determined in previous studies. Transiently transfected HeLa cells represent a useful tool in studying the interactions and kinetics of organic cations and other xenobiotics with hOCT1 and in understanding the molecular events involved in organic cation transport.
Organic cations are positively charged amines under physiological pH. Drugs from a wide array of clinical classes including antihistamines (e.g., cimetidine), antiarrhythmics (e.g.,procainamide), skeletal muscle relaxants (e.g., vecuronium) and beta adrenoceptor blocking agents (e.g.,acebutolol), as well as endogenous bioactive amines such as dopamine, NMN and choline are organic cations. These polar molecules are transported across epithelia by carrier-mediated processes. Based on previous studies in isolated plasma membrane vesicles and intact tissue preparations, a three-step model for the secretory flux of organic cations across various epithelia has been proposed: an electrogenic, facilitated diffusion step at the basolateral membrane, intracellular sequestration, and a proton or an organic cation exchange mechanism at the apical side (Pritchard and Miller, 1993; Zhang et al., 1998).
The first organic cation transporter, rOCT1, was cloned in 1994 from rat kidney (Grundemann et al., 1994). Since then several organic cation transporters have been cloned from various species and tissues (Gorboulev et al., 1997; Grundemann et al., 1997; Ikumi et al., 1997, Okuda et al., 1996; Terashita et al., 1998; Zhang et al., 1997a, b). Recently, we and others cloned a human organic cation transporter, hOCT1, from liver by homology cloning methods (Gorboulevet al., 1997; Zhang et al., 1997b). Initial functional expression studies carried out in Xenopus laevisoocytes (Zhang et al., 1997b) demonstrated that hOCT1, similar to rOCT1, is polyspecific with respect to substrate selectivity. Namely, the prototype organic cations, TEA and MPP+, are its permeants (substrates). In addition, both small and bulkier organic cations as well as other compounds such as nucleosides and bile acids inhibit the transport of MPP+ mediated by hOCT1 (Zhang et al.,1997b). Northern blot analysis indicates that the mRNA transcripts of hOCT1 are expressed primarily in the liver; however, reverse transcriptase polymerase chain reaction analysis demonstrates that hOCT1 mRNA transcripts also are expressed in lower abundance in the human kidney and intestine as well as in other tissues (Gorboulevet al., 1997; Zhang et al., 1997b).
X. laevis oocytes have been used extensively in elucidating the functional characteristics of transport proteins (Giacominiet al., 1994; Gorboulev et al., 1997; Grundemannet al., 1994; Murer and Biber, 1997; Okuda et al., 1996; Sigel, 1990; Terashita et al., 1998; Zhanget al., 1997a, b). Although the oocyte expression system has important advantages for studying transporters, it also has major disadvantages which limit its use particularly for routine studies of drug transport and in high throughput screening of drugs. Notably, oocytes are subject to seasonal variability in their viability as well as in their protein translation function. mRNA which is very unstable needs to be transcribed in vitro before being injected into oocytes. Moreover, injecting mRNA into oocytes requires tedious micro-injection techniques and specialized equipment. In contrast, mammalian expression systems do not require the in vitrohandling of mRNA and are adapted more readily to routine use in drug screening processes. Recently, mammalian expression systems have been developed and used in expressing several transport proteins (Boyeret al., 1994; Cardarelli et al., 1995; Clark and Amara, 1994; Martel et al., 1996; Risso et al., 1996; Schaner et al., 1997; Varoqui et al., 1996). For organic cation transporters, the human embryonic kidney (HEK) 293 cell line has been used to study the function of rOCT1 (Martel et al., 1996) and OCT2p, a second member of the OCT family cloned from LLCPK1 cells (Grundemannet al., 1997).
The goal of this study was to develop a mammalian cell expression system for hOCT1 and to elucidate the functional characteristics of the transporter in this cell line. Specifically, we determined the potency of various organic cations and other compounds in interacting with hOCT1. In addition, we examined the stereoselectivity of hOCT1 with the enantiomers of disopyramide and the diastereomers, quinine and quinidine. Finally, we determined whether the transporter can operate bidirectionally as an organic cation/organic cation exchanger. Our data represent the first demonstration of functional expression of a human organic cation transporter in a mammalian cell line (HeLa). A comparison of the data obtained in this study with data in the literature suggests that there are notable differences in the intrinsic function of hOCT1 and its rat homolog, rOCT1. Furthermore there are differences in the transport function of hOCT1 and a second human organic cation transporter, hOCT2 (Gorboulev et al., 1997). Transiently transfected HeLa cells represent a useful tool for the elucidation of the molecular mechanisms involved in the function of hOCT1.
Materials and Methods
Construction and isolation of plasmid DNA for transfection.
hOCT1 cDNA was obtained by reverse transcriptase polymerase chain reaction as described previously (Zhang et al., 1997b). After gel purification, the polymerase chain reaction products were ligated to the mammalian expression vector pTargeT (Promega, Madison, WI) with T4 DNA ligase followed by transformation into DH5α competent cells (Gibco, BRL, Gaithersburg, MD). The construct with the cDNA under the CMV promoter (sense orientation), pTargeT-hOCT1, was identified by restriction enzyme analysis and the sequence was confirmed by DNA sequencing (Biomolecular Resource Center, UCSF, CA). Empty vector of pTargeT was constructed by cutting the hOCT1 insert out of pTargeT-hOCT1 with EcoRI followed by gel purification and ligation. The construct of empty vector was confirmed by restriction enzyme analysis.
DNA for transfection was isolated with the QIAGEN Endo-free DNA isolation kit (QIAGEN Inc., Santa Clarita, CA). Several different DNA preparations were carried out for the study. DNA concentration ranged from 1.7 to 4.0 μg/μl as determined by UV spectroscopy. The DNA was stored in endotoxin-free TE buffer (QIAGEN) at −20°C until use.
HeLa cell culture and transfection.
HeLa cells were obtained from the UCSF Cell Culture Facility. Original stocks were from American Type Culture Collection (ATCC, Rockville, MD). Passages from 3 to 18 were used in the studies. The cells were grown at 37°C in a 5% CO2/95% air humidified atmosphere. The medium was Eagle’s minimum essential medium with Earle’s balanced salt supplemented with 2 mM glutamine, 100 IU/ml penicillin, 100 μg/ml streptomycin, 0.25 μg/ml fungizone and 10% (vol/vol) fetal bovine serum. The cells were maintained in Nunc cell culture flasks (Nalge Nunc International, Naperville, IL). The cells were seeded at a density of 1.8 × 105 cells/well in 12-well tissue culture plates (Corning Costar Corp, Cambridge, MA) 24 hr before transfection. The cells were transfected with a cationic liposome technique by LipofectAMINE (2 mg/ml, Gibco, BRL) observing a modified protocol from Gibco, BRL. For each well, 100 μl Opti-MEM media (Gibco, BRL) was incubated with 2 μg DNA and another 100 μl Opti-MEM media with the lipid (4–8 μl; 7 μl for most of the experiments). The two solutions then were mixed together and incubated for 30 min at room temperature. After incubation, 800 μl Opti-MEM media was added to the previous mixture. The final mixture (1 ml) was applied to each well after rinsing the cells with the Opti-MEM media once. The cells were exposed to the lipid-DNA complex for 18 hr before replacing the transfection media with the fresh standard culture media.
Uptake measurements.
The uptake studies were carried out between 24 and 72 hr post-transfection. Cells were incubated and washed with PBS once before the uptake studies. Subsequently, the cells were incubated at room temperature or 4°C (for temperature-dependence study) with 5 μM 14C-TEA (55 mCi/mmol, American Radiolabeled Chemicals, St. Louis, MO) in 0.5 ml of PBS. For Michaelis-Menten studies, various amounts of unlabeled TEA also were included in the reaction mixture. For inhibition and IC50 studies, various amounts of the tested compounds were included in the reaction mixture. Incubation was stopped by rinsing the cells once with 2 ml of ice-cold PBS and twice with 1 ml of ice-cold PBS buffer. After solubilizing the cells with 1 ml of 0.5% Triton X-100, 0.5 ml of sample was assayed by liquid scintillation counting (Beckman, Palo Alto, CA).
In trans-stimulation studies, each well of cells was preincubated with either 0.5 ml PBS (control) or 0.5 ml PBS plus the indicated concentration of unlabeled compounds at 37°C for 1 hr. Cells were then rinsed with 1 ml of ice-cold PBS twice before the uptake studies.
Protein assay.
For each plate used in the uptake study, two wells were saved for protein analysis. Cells were washed with PBS buffer and then solubilized with 0.5 ml of 1 N NaOH. After 2 hr, the solution was neutralized with 0.5 ml of 1 N HCl. 100 μl of solubilized cells were used for the protein assay with the Bio-Rad reagent (Bio-Rad, Hercules, CA). Absorbance was read at 595 nm, and the amount of protein was calculated from the standard curve generated by use of the known amounts of bovine serum albumin as standard.
Data analysis.
Uptake values are presented as mean ± standard deviation (S.D.) or mean ± standard error (S.E.) as indicated in the figure legends. In each experiment, a minimum of two wells was used to generate each data point, and each experiment was repeated at least once. For Michaelis-Menten studies, data were fit to the equation V =Vmax[S]/(Km+ [S]) + Kns[S] by nonlinear regression with Kaleidagraph Version 3.0 (Abelbeck Software). V is the transport rate, [S] is the substrate concentration and Kns is rate constant of nonspecific uptake. Kns was calculated from the rate constant of uptake in cells transfected with empty vector. The IC50 was estimated by a sigmoidal inhibition model and was fit to the equationV = V0/(1 + (I/IC50)n) by nonlinear regression. V is the uptake of TEA in the presence of the inhibitor, V0 is the uptake of TEA in the absence of inhibitor, I is the inhibitor concentration and n is the slope. Data from nonlinear regression are presented as mean ± error. For each compound, theKi was calculated from the corresponding IC50 assuming a competitive inhibition; however, because the ratio of 14C-TEA (5 μM) concentration used in the inhibition studies to theKm of TEA (230 μM) was less than 5%,Ki is virtually identical with IC50 regardless of the inhibition model (e.g., competitive versus noncompetitive) (Cheng and Prusoff, 1973). Statistical analysis was carried out by comparing the treated to the controls from the same experiments by an unpaired Student’s t test (Primer of Biostatistics software, Version 3, written by Stanton A. Glantz, McGraw-Hill Companies, 1991), and a value of P < .05 was considered significant.
Materials.
All the media and buffers used to maintain the cells were obtained from the UCSF Cell Culture Facility unless otherwise indicated. All chemicals were obtained from Sigma (St. Louis, MO) and Fisher (Pittsburgh, PA) or as indicated.R-(−)-disopyramide and S-(+)-disopyramide were resolved as described previously to greater than 98% purity (Valdivieso et al., 1988). 14C-TEA (55 mCi/mmol) was purchased from American Radiolabeled Chemicals (St. Louis, MO). 3H-MPP+ (79.9 Ci/mmol) was purchased from DuPont NEN Research Products (Boston, MA), and 3H-cimetidine (23.0 Ci/mmol) was purchased from Amersham (Arlington Heights, IL).
Results
Initial characterization of hOCT1 expression in transiently transfected HeLa cells.
The lipid to DNA ratio has been shown previously to be important in the transient expression of the DNA products in transfected cells (Hawley-Nelson et al., 1993). Initial titration studies with 1 to 3 μg lipid/μg DNA resulted in no significant 14C-TEA uptake in the cells transfected with pTargeT-hOCT1 versus untransfected cells (data not shown). Significant uptake was observed in the transfected cells when the lipid-to-DNA ratio ranged between 4:1 and 8:1 (the highest lipid to DNA ratio tested). A lipid-to-DNA ratio of 7:1, 6.5:1 or 6:1, which resulted in the highest activity, was used in subsequent studies. Because the 14C-TEA uptake in mock-transfected cells (cells transfected with the empty vector) was not significantly different from that in the untransfected cells, in some experiments, untransfected cells were used as controls (see figure legends).
14C-TEA influx into HeLa cells transfected with hOCT1 plasmid DNA increased with time and reached an apparent plateau (at 90 min) that was approximately 5-fold that in HeLa cells transfected with empty vector. At 1, 5, 10, 30, 60 and 90 min, the respective uptake values in the hOCT1 plasmid DNA transfected cells were 8.45 ± 1.04, 20.9 ± 5.81, 24.2 ± 6.30, 56.3 ± 4.36, 77.6 ± 20.4 and 93.3 ± 30.3 pmol/mg protein. Uptake in the empty vector transfected cells also increased with time (data not shown). A plot of uptake versus time was linear to 30 min with a positive intercept (which was apparent even in plots during the first 5 min). Therefore, further studies were carried out between 25 and 30 min.
The transient expression of hOCT1-mediated14C-TEA uptake in the HeLa cells was determined at various hours post-transfection (24–72 hr). At 24 hr,14C-TEA uptake was significantly greater (around 3-fold) in the pTargeT-hOCT1-transfected cells than in the cells transfected with the empty vector. At 41 hr post-transfection,14C-TEA uptake was approximately enhanced 5-fold of that in empty vector-transfected cells (fig.1). At 65 hr post-transfection,14C-TEA uptake in the pTargeT-hOCT1-transfected cells declined to 2-fold of that in the mock-transfected cells (fig.1). For further studies, uptake was measured between 24 and 48 hr after transfection.

Transient expression of hOCT1 in HeLa cells over time. After transfection of pTargeT-hOCT1 (•) and empty vector (○) in HeLa cells, the 30-min uptake of 14C-TEA was measured at 24, 41 and 65 hr post-transfection. Data represent mean ± S.D. of duplicate determinations from one representative experiment.
To determine whether 14C-TEA uptake in pTargeT-hOCT1 transfected HeLa cells is temperature dependent, an important characteristic of carrier-mediated processes,14C-TEA uptake at various time points was measured at 22°C and 4°C in HeLa cells transfected with pTargeT-hOCT1 and empty vector. Figure 2showed that 14C-TEA uptake in pTargeT-hOCT1-transfected cells was significantly higher at 22°C than that at 4°C.

Temperature dependence of 14C-TEA transport in pTargeT-hOCT1 (dark solid bars and horizontal hatched bars) and empty vector (vertical hatched bars and open bars)-transfected HeLa cells. Uptake of 14C-TEA (5 μM) was measured at various time points at 22°C (dark bars and vertical hatched bars) and 4°C (horizontal bars and open bars) 42 hr post-transfection. Data represent mean ± S.D. of duplicate determinations from one representative experiment.
Permeant studies.
The rate of 14C-TEA uptake in the pTargeT-hOCT1-transfected cells was saturable, whereas the uptake rate in the empty vector-transfected cells was linear across the same concentration range (fig. 3). The Vmax and Kmof TEA transport were 2.89 ± 0.448 nmol/mg protein/30 min and 229 ± 78.4 μM, respectively; and theKns, the rate constant for the linear process, determined from TEA uptake in the empty vector-transfected cells was 2.46 pmol/mg protein/30 min/μM.

Concentration dependence of 14C-TEA transport in pTargeT-hOCT1 (•) and empty vector (○)-transfected HeLa cells. 14C-TEA transport (at 30 min) was measured in increasing concentrations of TEA. The solid and dotted lines represent the estimated overall and nonsaturable transport, respectively. Data represent mean ± S.D. of duplicate determinations from one representative experiment.
3H-MPP+ (35 nM labeled and 1 μM unlabeled) uptake in the hOCT1-transfected cells was high but not significantly different from that in the mock-transfected cells (34.0 ± 1.49 vs. 26.2 ± 2.15 pmol/mg protein at 5 min and 50.3 ± 3.96 vs. 44.0 ± 3.46 pmol/mg protein at 30 min). In comparison with the endogenous uptake of TEA, there was a high endogenous uptake of MPP+ in HeLa cells (42.6 pmol/mg protein/μM for MPP+uptake versus 2.12 pmol/mg protein/μM for TEA uptake at 30 min). This high endogenous uptake may have obscured detection of the expressed transporter when MPP+ was used as the radiolabeled ligand.
Cimetidine, an antihistamine, has been used as a model compound for organic cation transport in some studies. During a 60-min period,3H-cimetidine (24 nM labeled plus 5 μM unlabeled) uptake in HeLa cells expressing hOCT1 was enhanced only slightly more than in the mock-transfected cells, which suggests that under the experimental conditions, cimetidine is not translocated appreciably by hOCT1 (fig. 4). The endogenous uptake of cimetidine was moderate (6.84 vs. 2.12 pmol/mg protein/μM for cimetidine vs. TEA uptake at 30 min).

Time course of 3H-cimetidine (5 μM) uptake in the pTargeT-hOCT1-transfected (•) and empty vector-transfected (○) HeLa cells. Data represent mean ± S.D. of duplicate determinations from one representative experiment.
Inhibition studies.
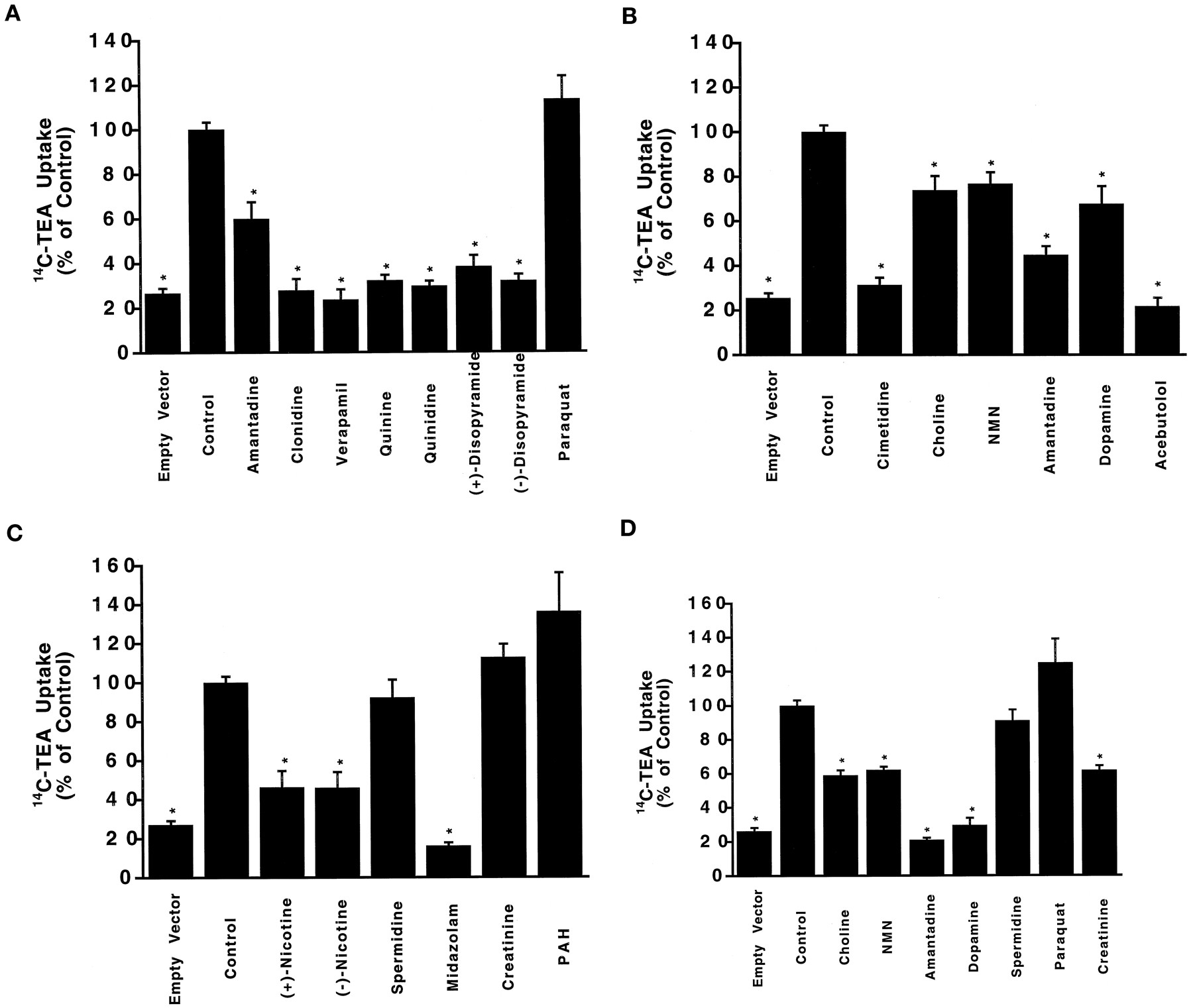
To characterize the function of hOCT1 further, we studied 14C-TEA uptake in the presence of various compounds at concentrations of 0.1 mM, 0.5 mM or 5 mM (fig. 5, A–D). At 0.1 mM, the organic cations, amantadine, clonidine, verapamil, quinine, quinidine,S-(+)-disopyramide and R-(−)-disopyramide significantly inhibited 14C-TEA uptake (fig. 5A, P < .05). In contrast, paraquat, dopamine and its precursors (l-dopa and d-dopa) (not shown) did not significantly affect the uptake of 14C-TEA. At 0.5 mM, cimetidine, choline, NMN, amantadine, dopamine, acebutolol, (+)-nicotine, (−)-nicotine and midazolam significantly inhibited14C-TEA uptake (fig. 5, B and C, P < .05), whereas spermidine (polyamine), creatinine (zwitterion) and the organic anion, PAH, did not produce any inhibition (fig. 5C). At high concentrations (5 mM), paraquat, spermidine, l-dopa,d-dopa and PAH (data not shown) did not produce significant inhibition, whereas creatinine did (fig. 5D, P < .05). These data indicate that hOCT1 is broadly selective for organic cations and some other compounds, but not the organic anion, PAH, nor the multivalent cations, paraquat and spermidine.

Inhibition of 14C-TEA uptake. The uptake of 5 μM 14C-TEA (at 25 min) was measured in the presence of 0.1 mM (A), 0.5 mM (B and C) and 5 mM (D) of the given compounds in pTargeT-hOCT1-transfected HeLa cells. Controls represent uptake of 14C-TEA in the HeLa cells transfected with pTargeT-hOCT1 in the absence of inhibitors. Uptake of14C-TEA in the empty vector-transfected cells in the absence of inhibitors is shown as well (indicated as Empty Vector). Data represent the mean ± S.E. (n = 4–22) obtained from 2 to 10 separate experiments. * P < .05.
To elucidate the functional properties of hOCT1 expressed in different expression systems, Ki values of the organic cations TEA, decynium-22 and vecuronium were compared with those obtained previously in the oocyte expression system (table1) (Zhang et al., 1997b). In the transfected cells, vecuronium had a somewhat higherKi (232 μM vs. 120 μM), whereas decynium-22 had a slightly lower Ki(2.7 μM vs. 4.4 μM).
Comparison of Ki values of organic cations determined from experiments conducted in X. laevis oocytes or transfected HeLa cells
IC50 values of selected compounds in inhibiting14C-TEA uptake mediated by hOCT1 were determined by nonlinear regression analysis. Ki values were calculated from the IC50 values assuming a competitive model; however, because the substrate concentration used in the experiment is low compared with the Km(i.e., 5 μM vs. 229 μM),Ki values will be virtually identical with the IC50 values despite the mechanism of inhibition (e.g., competitive or noncompetitive) (Cheng and Prusoff, 1973). Ki values were compared with those determined previously for rOCT1 and hOCT2 (see table 2) (Gorboulev et al., 1997; Grundemann et al., 1994,1997). Notable differences were observed in theKi values of some compounds in interacting with hOCT1 and rOCT1, which suggests that there may be important interspecies differences in the intrinsic function of the transporters. Furthermore, with the exception of desipramine,Ki values generally were lower for hOCT2 than for hOCT1.
Ki of various compounds in inhibiting TEA uptake mediated by hOCT1, rOCT1 and hOCT2
To test whether there are stereoselective interactions of organic cations with hOCT1, the IC50 values of pairs of isomers were determined and compared. The IC50 for quinine was 23.4 ± 6.94 μM, and for quinidine was 17.9 ± 4.69 μM (fig. 6A). The IC50 of S-(+)-disopyramide was 29.9 ± 8.50 μM, and of R-(−)-disopyramide, it was 15.4 ± 11.0 μM (fig. 6B) (P < .05).

Concentration-dependent inhibition of14C-TEA uptake by stereoisomers in HeLa cells transfected with pTargeT-hOCT1. Initial rates of transport were determined in the presence of quinidine (A, •), quinine (A, ⋄),S-(+)-disopyramide (B, •) andR-(−)-disopyramide (B, ⋄). Data represent mean ± S.D. of duplicate determinations from one representative experiment.
Trans-stimulation studies.
To determine whether hOCT1 can transport organic cations in both directions,trans-stimulation studies were performed. As shown in figure7, after preincubation of the pTargeT-hOCT1-transfected HeLa cells with TEA (2 mM) or MPP+ (0.5 mM) for 1 hr at 37°C,14C-TEA uptake was enhanced significantly (P < .05). The trans-stimulation also could becis-inhibited more than 50% with 0.5 mM unlabeled TEA in the reaction media (data not shown). In contrast, preincubation of cells with 2 mM cimetidine did not result in a significant change in14C-TEA uptake and preincubation of cells with 50 μM decynium-22 resulted in a significant decrease (apparent “trans-inhibition”) of 14C-TEA uptake (P < .05).

Trans-stimulation of14C-TEA uptake in pTargeT-hOCT1-transfected HeLa cells. The uptake (at 25 min) of 14C-TEA (5 μM) was measured after 60 min incubation of the untransfected cells with PBS, and pTargeT-hOCT1-transfected cells with PBS (control) or PBS containing 2 mM TEA, 0.5 mM MPP+, 2 mM cimetidine or 50 μM decynium-22, respectively, at 37°C. Data represent the mean ± S.E. (n = 4) obtained from two separate experiments. * P < .05.
Discussion
The present study demonstrates that hOCT1 can be expressed functionally in a transiently transfected mammalian cell expression system (HeLa cells) by a lipofection method. Maximal TEA transport activity occurred between 36 and 48 hr (fig. 1) after transfection and was observed when the lipid-to-DNA ratio was between 6:1 and 8:1 (data not shown). In this system, TEA transport was temperature dependent and was saturable with kinetic values similar to those obtained in X. laevis oocytes injected with the cRNA of hOCT1 (Zhang et al., 1997b) (Ki, 161 μM in HeLa cells vs. 163 μM in oocytes). In addition, less than 2-fold differences in the Ki values of decynium-22 and vecuronium were observed in cRNA-injected oocytes (Zhang et al., 1997b) and transfected HeLa cells (table 1), which suggests that hOCT1 has similar functional characteristics in the two expression systems.
A major goal of this study was to determine the functional characteristics of hOCT1. In particular, the substrate selectivity of the transporter was examined. Paraquat and spermidine are multivalent organic cations. Our data demonstrated that neither compound interacted with hOCT1 at 5 mM, which suggests that the transporter does not play a role in the uptake of these compounds in human epithelia. These data are consistent with data in the literature which demonstrate that paraquat has a distinct transport pathway from TEA in the peritubular membrane of the rabbit kidney (Groves et al., 1995). The data also suggest that the polyamine, spermidine, is not a substrate of hOCT1, consistent with data in the literature which demonstrate unique transporters for polyamines (Sokol and Gates, 1990). Spermidine previously was shown to induce inward currents in oocytes expressing the rat homolog, rOCT1 (Busch et al., 1996), which indicates that spermidine may be translocated by rOCT1. The data suggest that there are interspecies differences in the function of the OCT1 transporters. However, recent studies demonstrated that a compound (e.g., quinine) could induce large inward currents without being translocated (Nagel, et al., 1997). Further studies are needed to determine whether spermidine is a substrate for rOCT1. Although spermidine and paraquat did not interact with hOCT1, the multivalent organic cation, vecuronium interacts in micromolar concentrations (Ki = 232 μM), which suggests that the nature of the hydrophobic moiety may play a role in the potency of interaction of multivalent organic cations with hOCT1.
We determined the effect of the neutral molecules, corticosterone and midazolam, on the transport of 14C-TEAvia hOCT1. Previously, corticosterone was thought to enter the cell via simple diffusion to gain access to its sites of action. However it is becoming increasingly clear that transporters may be involved in the entry of corticosterone into cells. For example, corticosterone has been shown to interact with both organic cation and organic anion transport in the microperfused kidney (Ullrich et al., 1993), and recently, has been shown to interact specifically with the rat liver organic anion transporter (oatp) (Bossuyt et al., 1996; Kanai et al., 1996), and the rat organic cation transporter, rOCT1 (Grundemann et al., 1994, 1997). The present study demonstrated that corticosterone is a potent inhibitor of TEA uptake mediated by hOCT1. Further studies are needed to determine whether corticosterone is an actual substrate or whether it inhibits, but is not translocated by hOCT1.
The neutral compound, midazolam, is a cytochrome P450 3A (CYP3A) substrate with a Km of approximately 10 μM for 1′-hydroxymidazolam formation (Wrighton and Ring, 1994). TheKi of midazolam in inhibiting TEA transport by hOCT1 is in the same range of its Km of metabolism by CYP3A. Because hOCT1 is expressed primarily in the liver, which is the major site for the metabolism of midazolam, it is possible that transport rate limits the metabolism of midazolam. Collectively, these data indicate that hOCT1 is polyspecific not only for organic cations but also for various neutral compounds. The positive charge(s) of a molecule may not be the only structural requirement for interaction with hOCT1 as previously hypothesized for OCT transporters. The hydrophobic moiety also may play a role in the potency of interaction of a chemical entity with hOCT1. Further structure-function relationship studies are needed to clarify this.
Numerous xenobiotics are chiral and contain one or more asymmetric carbon atoms, which results in two or more enantiomeric forms. There are many examples of differences in pharmacological activity between enantiomeric compounds (Blaschke and Giacomini, 1987; Drayer, 1986;Levy and Boddy, 1991). Stereoselective metabolism of xenobiotics has been well studied and documented (Drayer, 1986; Levy and Boddy, 1991). In this study, we examined the potency of the interaction of various isomers in inhibiting TEA uptake in hOCT1-transfected cells. Within the two pairs tested, the diastereoisomers, quinine and quinidine, had no significant differences in IC50 values (23.4 ± 6.94 μM vs. 17.9 ± 4.69 μM) whereasS-(+)-disopyramide was approximately 2-fold less potent thanR-(−)-disopyramide (29.9 ± 8.50 μM vs. 15.4 ± 11.0 μM). These data suggest that hOCT1 is not highly stereoselective, which is consistent with the polyspecificity of the transporter.
Ki values of various compounds in inhibiting TEA or MPP+ uptake mediated by hOCT1 (Gorboulev et al., 1997; Zhang et al., 1997b) and rOCT1 (Grundemann et al., 1994, 1997; Martel, 1996) (i.e., between species) are listed in table2. Ki values of TEA, MPP+, desipramine and clonidine in interacting with hOCT1 and rOCT1 are similar (within 3-fold). In contrast, most of the other organic cations studied, including decynium-22, procainamide, NMN, quinine and vecuronium, have much higher Ki values (more than 3-fold) for hOCT1 than for rOCT1. These data indicate that in comparison with rats, humans possess an organic cation transporter with a lower affinity for most compounds, which suggests that in human epithelial cells organic cations translocated primarily by OCT1 transporters will be eliminated more slowly than in rats. For example, it has been shown previously that the organic cation vecuronium is transported at a considerably slower rate in human hepatocytes than in rat hepatocytes (Sandkeret al., 1994). Higher Ki values of vecuronium for hOCT1 than for rOCT1 may explain in part its slower transport rate in humans. The steroid, corticosterone, however, has a higher affinity for hOCT1 than for rOCT1, which may suggest that cholesterol compounds will have more potent interaction with organic cation transport in humans than in rats. However, further studies are needed to determine whether the interacting compounds are actual substrates of OCT1 transporters and not simply inhibitors. Moreover, it will be essential to study the site-specific localization of the OCT1 and the relative amounts of the transporters expressed in various tissues.
Comparisons were made between Ki values of various compounds in interacting with the organ-specific transporters in humans, hOCT1 (expressed primarily in liver) (Gorboulev et al., 1997; Zhang et al., 1997b) and hOCT2 (expressed primarily in kidney) (Gorboulev et al., 1997) (table 2). hOCT2 apparently has a higher affinity for most of the organic cations studied except for desipramine, which seems to have lower affinity for hOCT2 than for hOCT1. Thus, it appears that the kidney-specific organic cation transporter, hOCT2, more potently interacts with potential substrates than the liver-specific transporter, hOCT1.
Previous studies have shown that the transport of organic cationsvia hOCT1 can be driven by a favorable (inside negative) electrical potential difference (Zhang et al., 1997b). However, it is not known whether transport of organic cations may also be driven by the exchange or countertransport of other organic cations. Our data demonstrate that 14C-TEA transport can be driven by the countertransport of unlabeled TEA (fig. 7), which suggests that hOCT1 may operate as an organic cation/organic cation exchanger. In addition, the ability of a compound totrans-stimulate the transport of14C-TEA mediated by hOCT1 indicates that the compound also is translocated by the transporter. The finding that MPP+ trans-stimulated14C-TEA transport is consistent with previous studies in oocytes which demonstrated that MPP+is a substrate of hOCT1 (Zhang et al., 1997b). However, because of the high background uptake of3H-MPP+ in the empty vector-transfected cells, an enhanced uptake of3H-MPP+ was not observed in the hOCT1-transfected cells. The finding that cimetidine did nottrans-stimulate TEA uptake, consistent with results from permeant studies, indicates that cimetidine is not a good permeant of hOCT1 under the experimental conditions (fig. 4). In contrast, decynium-22 was shown to “trans-inhibit” TEA uptake, which suggests that decynium-22 binds tightly to the transporter and is not washed off during the experimental procedures. Alternatively, hOCT1 loaded with decynium-22 cycles more slowly than the unloaded transporter. Collectively, these data suggest that hOCT1 is a uniporter which can translocate organic cations in both directions. When radiolabeled compounds are not available, trans-stimulation studies provide an alternative way of determining whether a chemical entity is a substrate for the transporter.
In summary, hOCT1 has been transiently expressed in a mammalian cell line, HeLa. The fundamental properties of TEA transport in this system are similar to those previously described in X. laevisoocytes. Using this expression system, studies were conducted which demonstrated the selectivity of the transporter for a variety of substrates. Differences in the Ki values of certain compounds in interacting with the rat and the human organic cation transporters, and the human liver-specific versuskidney-specific organic cation transporters were observed. This study provides the first information about the molecular basis for the differences in organic cation transport observed among species and organs. The development of this mammalian expression system will facilitate the study of drug interactions and transport in vitro and the development of drugs that specifically target cells that express hOCT1.
Footnotes
-
Send reprint requests to: Kathleen M. Giacomini, Ph.D., Department of Biopharmaceutical Sciences, Box 0446, University of California, San Francisco, San Francisco, CA 94143-0446.
-
↵1 This study was supported by grants from the National Institutes of Health (GM-36780 and GM-57656).
-
↵2 Supported in part by the UCSF Chancellor’s Research Fellowship.
- Abbreviations:
- OCT
- organic cation transporter
- TEA
- tetraethylammonium
- MPP+
- 1-methyl-4-phenylpyridinium
- NMN
- N1-methylnicotinamide
- PAH
- p-aminohippuric acid
- dopa
- 3, 4-dihydroxyphenylalanine
- PBS
- phosphate-buffered saline
- Received November 20, 1997.
- Accepted March 6, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}