


Abstract
In this study, the aorta vasorelaxant, coronary calcitonin gene-related peptide (CGRP) releasing, and atrial contractility effects of glyceryl nonivamide (GLNVA) were investigated in guinea pigs. In the isolated thoracic aorta, although GLNVA (0.01–50 μM) concentration dependently induced endothelium-independent relaxations and relaxed phenylephrine-(1.0 μM) induced contractions, it failed to relax 80 mM KCl-induced contractions. The GLNVA (1.0 μM) relaxation response in the aorta was significantly inhibited by tetraethylammonium (2.5–10 mM) or ouabain (5.0 μM) and was attenuated by increased extracellular potassium gradient (10–30 mM). Glibenclamide (0.01–10 μM) dose dependently antagonized the GLNVA relaxant effect. In the isolated perfused guinea pig heart, GLNVA (0.1–10 μM) increased CGRP-like immunoreactivity outflow from coronary circulation in a concentration-dependent manner. In the isolated right and left guinea pig atria, GLNVA (0.01–10 μM) produced concentration-dependent positive inotropic and chronotropic effects, but these effects were inhibited by pretreatments with ruthenium red (1.0 μM), capsazepine (10 μM), human calcitonin-gene-related peptide (CGRP8–37) (1.0 μM) and sensory neuron denervation, respectively. Based on these findings, we suggest that CGRP may be released by GLNVA from cardiovascular sensory neuron, and it then activates CGRP receptors on the coronary artery and atrium. The GLNVA-induced vasorelaxant effect in the vascular smooth muscle of the aorta is due to CGRP release associated K+ channel opening, and this effect eliminates capsaicin-derived excitability-associated K+ channel blocking activities.
Capsaicin, a pungent natural product of red pepper, is known to possess a wide spectrum of biological activities (Holzer, 1991). It has been used to activate sensory C-fiber to release several neuropeptides, including substance P, neurokinin A and CGRP (Bucsics and Lembeck, 1981; Franco-Cerecedaet al., 1988; Manzini et al., 1989). Furthermore, capsaicin has also been found to have desensitizing and antinociceptive benefits with its long-term local administration (Holzer, 1991;O’Neill, 1991; Stretton et al., 1992). Franco-Cereceda and Lundberg, (1985) reported that capsaicin, similar to CGRP, caused positive inotropic and chronotropic effects on guinea pig right atria. This cardiotonic effect of capsaicin may be secondary to the release of endogenous CGRP from local sensory nerve endings within the heart (Franco-Cereceda et al., 1988). In the cardiac and sensory neuron system, the stimulated release of endogenous CGRP by capsaicin is not generally associated with the positive chronotropic and inotropic activities necessary for the treatment of cardiac dysfunction. However, CGRP has been shown to improve cardiac performance in patients with congestive heart failure (Gennari et al., 1990). CGRP has also been proven to enhance adenylate cyclase activity, resulting in an elevation of cyclic AMP content that then increases calcium influx and potassium permeability (Ohmura et al., 1990).
Recently, because capsaicin reduces ischemic ventricular tachycardia and fibrillation in the isolated perfused hearts of rat and guinea pig, it was suggested that capsaicin has antiischemic and antiarrhythmic activity associated with its Ca++ channel inhibition (D’Alonzo et al., 1995). However, capsaicin may have a second direct action on cardiac myocyte and prolong the action potential associated with the inhibition of K+ current (Castle, 1992). Capsaicin also inhibits the contractile tension of isolated rat papillary muscle (Yamato et al., 1996b).
Regarding blood vessels, acute application of capsaicin in vitro has produced a vascular smooth muscle relaxation and a delayed smooth muscle contraction. It was suggested that the contractile response is most likely due to direct effects on vascular smooth muscle, although the relaxation response is attributed to release of CGRP (Franco-Cereceda et al., 1988; Lo et al., 1995). Our electrophysiological study of capsaicin in isolated and cultured vascular smooth muscle cells of rat aorta has indicated that capsaicin might cause relaxation of aorta smooth muscle and inhibition of voltage-dependent L-type Ca++ channels, whereas capsaicin-induced inhibition of voltage-dependent K+ channel might produce an increase in cell excitability (Lo et al., 1995). In our study, vascular contractile effects of capsaicin on aorta smooth muscle is proposed to be due to an increase in cell excitability associated with K+ channel blocking activity.
In addition to its positive inotropic effects, the vasodilatory effects of capsaicin and CGRP may be beneficial in the treatment of cardiovascular dysfunction. Up to date analogues of capsaicin have rarely been used to compare CGRP releasing, antiarrhythmic effects, antiischemic and vasorelaxant activities that are derived from capsaicin. Capsinolol, another capsaicin derivative previously evaluated as a β-adrenoceptor blocker with a CGRP releasing property, may share many of the beneficial effects of capsaicin and released CGRP (Chen et al., 1996). We have hoped to find some other analogues of capsaicin that cause the release of CGRP, that did not block the K+ channel, but had the same vasorelaxant activity as capsaicin. GLNVA, a chemically modified product of capsaicin, synthesized and tested in vivo previously, was thus further tested in vitro in our study to ascertain whether it had these properties.
GLNVA is a nonpungent and antinociceptive capsaicin derivative (Chenet al., 1992). At high doses, we found it to have a blood pressure lowering effect, which is different from capsaicin, and to induce triphasic blood pressure and heart rate changes in rats (Yehet al., 1993; Lo et al., 1994). The depressor effect of GLNVA was halted with an intrathecal injection of substance P antagonist [d-Pro2,d-Trp7,9]-SP (Yeh et al., 1993). The aim of our study was to ascertain whether GLNVA displayed similar CGRP releasing, K+ channel opening and enhanced vasorelaxant activities to capsaicin to provide the same potential capsaicin-derived benefits in the cardiovascular system, without the excitability-associated K+ channel blocking activities of capsaicin.
Methods
Vasorelaxation effect in isolated guinea pig aorta.
Guinea pigs weighing 200 to 300 g were anesthetized with ether before they were killed. The aorta was quickly removed, cleaned of adhering fat and connective tissue, and cut into 3 to 4 mm wide transverse rings. Then, the tissue was mounted at 1.0 g resting tension on stainless steel hooks in 10 ml organ baths and bathed at 37°C in a PSS containing (in millimolar): NaCl, 140.0; KCl, 4.6; CaCl2, 1.5; MgCl2, 1.0; glucose, 10.0; and 4-(2-hydroxyethyl)-1-piperazine-ethane-sulfonic acid, 5.0. The pH was adjusted to 7.25 with 0.1 N NaOH, aerated with a mixture of 95% O2 and 5% CO2. One end of the preparation was fixed at the bottom of the bath, and the other end was connected with a hook to the level of a force displacement transducer (Ugo Basile, model 7004, Italy). The tension was recorded on-line on an AT High Speed Videography unit (Coulbourn Instruments, Lehigh Valley, PA) that was connected to a personal computer with the use of an interface card and built-in software ACODAS (Wu et al., 1991; Lo et al., 1995). The tissues were always allowed to equilibrate for 90 min before experiments were begun. The endothelium of rat aorta was stripped by pulling a thread back and forth through the vessel lumen, confirmed later by the inability of carbachol (1.0 μM) to induce relaxation.
Ca++ channel activity on aorta smooth muscle.
In the test for the possible involvement of Ca++ blocking activity, according to the method of Meisheri et al. (1987,1988), such Ca++ mobilizers as PE, high K+solution and Ca++ channel blocker were added to the bath, respectively. Briefly, a control contraction was first produced by exposing rat aorta to PE (1 μM). Fifteen min after determination of the maximal response to PE, the tissue was washed with PSS five times and allowed to return to resting tension for 1 hr before exposure to either PE (1.0 μM) or high K+ solution (20–80 mM KCl). The tissues were pretreated with 20 μM phentolamine for 15 min before exposure to high K+ solution to prevent the effect of endogenously released norepinephrine. After achieving a plateau in the contractions (15–20 min), GLNVA (1.0 μM) was added to the bath to observe its effect on relaxation activity. A contrast experiment was also carried out with D600 (1.0 μM), a calcium antagonist, to observe its effect on relaxation activity. High K+-PSS containing 80 mM KCl was prepared by equimolar replacement of NaCl with KCl. Intermediate high-K+ solutions containing 25 or 30 mM KCl were prepared by proportional mixing of 80 mM K+-PSS and normal PSS. In cases of 20 mM K+-PSS, extra KCl was added to the normal PSS without lowering the NaCl content.
K+ channel opening activity on aorta smooth muscle.
In the test for the extent of involvement of K+ channel opening activity, K+ gradient, K+ channel blocker and Na+-K+-ATPase inhibitor were added to the bath, respectively. Briefly, the isolated aorta was first contracted with PE (1.0 μM) as described above. Before their second exposure to PE, the tissues were exposed for 20 min to PSS that contained increasing concentrations of KCl (10–25 mM). GLNVA (1.0 μM) was then added at the plateau of PE contraction. Phentolamine was not used in these experiments as the tissues were exposed to PE after K+treatment.
The effects of tetraethylammonium (TEA, a nonspecific calcium-activated potassium channel inhibitor) and ouabain (a Na+-K+-ATPase inhibitor) on GLNVA relaxation of PE contraction were also obtained in normal PSS and K+-free PSS. As described above, between the two PE contractions the tissues were exposed for 1 hr to PSS containing TEA (2.5, 5.0, 7.5 and 10 mM) or ouabain (5.0 μM). We carried out two experiments in which we examined the effects of ouabain (5.0 μM) and TEA (10 mM) under the condition of K+-free PSS. The experimental protocol remained the same as described above with the exception that, before the second PE contraction, the tissues were exposed to K+-free PSS for 1 hr with or without the additional treatments of ouabain or TEA. K+-free PSS contained no KCl, and correspondingly NaCl was increased to 144.6 mM. The rest of the ingredients in K+-free PSS remained the same. In these experiments, forskolin (1.0 μM) was used to examine the activity of vessel smooth muscle.
Effect of ATP-sensitive K+ channel blocker on isolated guinea pig aorta.
In the test for possible ATP-sensitive K+ channel opening activity of GLNVA, glibenclamide (0.01–10 μM), a specific blocker of ATP-dependent K+channels, was added to the bath for 45 min before the experiment to block possible K+ channel opening activity of GLNVA.
Cardiosensory functioning in isolated guinea pig atrium.
In the test for possible sensory C-fiber activating and the CGRP-releasing activities of GLNVA in the atria, guinea pigs weighing 200 to 300 g were anesthetized with ether and the right and left atria were quickly dissected and mounted in a 10-ml organ bath at a passive force of 0.5 × g. The atria were perfused with Tyrode’s solution containing (in millimolar): NaCl, 137.0; KCl, 5.4; MgCl2, 1.05; NaHCO3, 11.9; NaH2PO4, 0.42; glucose, 5.6 and CaCl2, 1.8. The solution was aerated with a mixture of 95% O2 and 5% CO2 that was kept at 37°C. Mechanical activity was measured using a force displacement transducer (Grass, Quincy, MA, Model FTO3). The frequency of contraction was measured on a separate channel by a tachometer (Coulbourn Instruments, model S77-26, Allentown, PA) and recorded on a High-Speed Videograph unit (Coulbourn Instruments, AT, L19-69, Allentown, PA). The preparations were allowed to equilibrate for 1 hr before the administration of drugs. The left atrium was stimulated at a frequency of 1 HZ by rectangular pulses of 1 msec duration at supramaximal intensity with an isolated stimulator (Grass, model S11, Quincy, MA).
Sensory denervation.
To stop the sensory neuron from functioning and releasing CGRP, capsaicin (50 mg/kg/day) was administered s.c. for 2 wks to induce denervation of the sensory neuron in guinea pigs before the experiments. This may cause an almost total loss of SP- and CGRP-immunoreactive nerves within the heart (Lundberget al., 1985). Terbutaline (50 μg/kg, s.c.) and theophylline (100 mg/kg, i.p.) were given to ensure respiratory function. Ketamine (50 mg/kg, i.m.) was given as an analgesic during the administration of capsaicin.
Sensory neuron activation and CGRP release.
To examine the sensory neuron activating and the CGRP releasing activities of GLNVA in the isolated guinea pig atrium, after the addition of either a competitive capsaicin receptor antagonist capsazepine (1.0 μM), a cation channel blocker ruthenium red (1.0 μM), a CGRP antagonist hCGRP8–37 (1.0 μM) or a β-adrenoceptor blocker atenolol for 30 min, GLNVA was added to the respective baths to induce atrial contraction.
Immunoreactivity of released CGRP from coronary circulation.
Guinea pigs weighing 200 to 300 g were anesthetized with ether. After rapid cannulation of the aorta, 100 IE heparin in 5 ml of Tyrode’s solution were injected into the coronary circulation. The heart was then excised and perfused according to the Langenodorff technique (Broadley, 1979) with a pressure of 70 cm H2O. The solution was warmed to 37°C and gassed with 95% O2and 5% CO2 immediately before entering the coronary circulation. Perfusate fractions (5 min) were collected on ice to give a final concentration of acetic acid of about 0.2 M. The perfusate samples were desalted, using SEP-PAK C18 cartridges (Wu et al., 1983), lyophilized and redissolved in 0.5 ml of appropriate buffer to determine CGRP-LI by radioimmunoassay with an antiserum raised against human CGRP-α (which cross-reacts equally well with rat CGRP-α; RAS 6009) (Peninsula, CA) (Chen et al., 1996).
Drugs and chemicals.
Glyceryl nonivamide was synthesized in our institution. Capsaicin, human CGRP (8–37), ruthenium red, ketamine, phenylephrine, ouabain, TEA chloride, glibenclamide and D600 were purchased from Sigma Chemical Co. (St. Louis, MO). Other chemicals used were of reagent grade. All drugs were dissolved in Tyrode’s solution immediately before the experiments, except capsaicin and GLNVA that were first dissolved in 10% ethanol and 10% Tween 80 to a concentration of 1.0 mM and further diluted in Tyrode’s solution. Capsazepine was dissolved in dimethylsulfoxide to a concentration of 10 mM and then further diluted in 5% ethanol and 5% Tween 80.
Data analysis.
All values and figures are expressed as mean ± S.E. Statistical differences were evaluated by Student’st test in unpaired samples and by paired t test in paired samples. When evaluating multiple values, the Bonferroni correction was used. P < .05 were considered to be significant. Analysis of the data and plotting of the figures were done with the aid of software (SigmaStat and SigmaPlot, version 5.0, Jandel; PHARM/PCS, version 4.2, MCS, CM, CA) run on an IBM PC-AT computer.
Results
Vasorelaxation effect in the isolated guinea pig aorta.
The intact and endothelium-denuded guinea pig thoracic aorta contracted by PE (1.0 μM) were relaxed by GLNVA (1.0 μM) (fig.1A-a,b). In rat endothelium-denuded aortic rings, GLNVA relaxed PE-induced contractions in a concentration-dependent manner (fig. 1B). However, as shown in the tracing of figure 1A-c, GLNVA produced only slight relaxation in high-K+-induced contraction, and these preparations could be relaxed by D600, a calcium antagonist.

A, Typical recordings of inhibitory effects of GLNVA (1.0 μM) on PE (1.0 μM) and 80 mM K+-induced maximal contraction in the isolated guinea pig aorta. GLNVA relaxed PE contraction (a and b), but did not relax high K+contraction (c). D600 relaxed the high-K+ contraction. E(+): with endothelium; E (−): without endothelium. B, Concentration-response curve of GLNVA on endothelium denuded guinea pig thoracic aorta. Data are expressed as the percentage of the respective first PE (1.0 μM) contraction (n = 8).
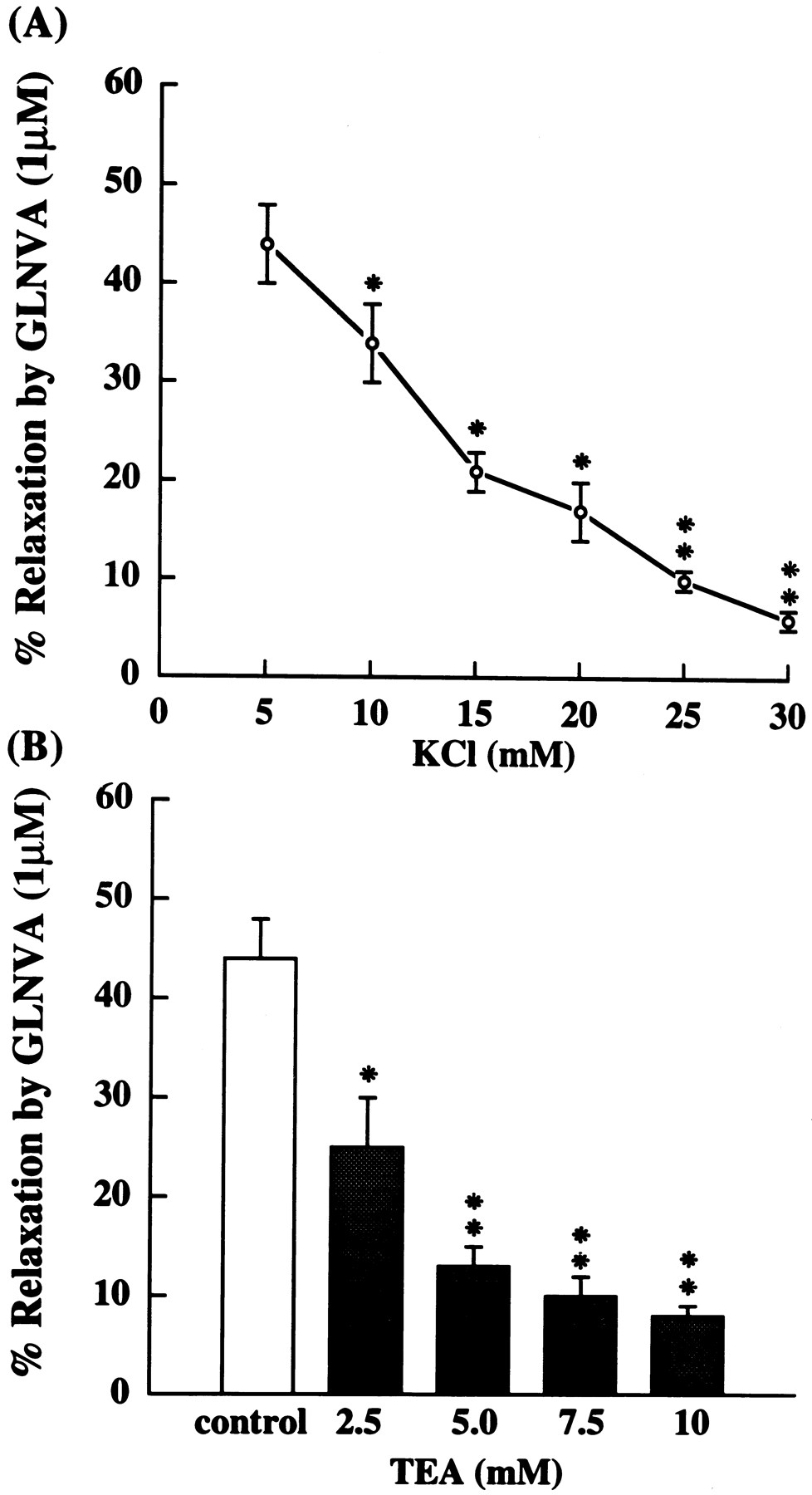
Because GLNVA could not relax high-K+-induced contractions, we investigated whether the K+ gradient and TEA, a potassium channel blocker, were significantly involved in the GLNVA-induced relaxation effect. Increasing extracellular K+ (5–30 mM) was found to inhibit the relaxation effect of GLNVA (fig. 2A) and pretreatment with TEA (2.5–10 mM) for 1 hr also inhibited the relaxation effect of GLNVA in a concentration-dependent manner (fig. 2B). Increasing extracellular K+ (10, 15 and 20 mM) also was found to cause an increase in the contraction produced by PE. However, under the conditions of normal PSS (containing 4.6 mM K+), 25 or 30 mM K+, the second PE contraction was not significantly greater than the first control PE contraction. Under these conditions, relaxations could still be obtained with other vasodilators, such as D600 (5.0 μM).

A, Influence of extracellular K+ gradient on GLNVA-induced relaxation of endothelium denuded guinea pig thoracic aorta contracted by 1.0 μM PE. The percentage of relaxation by GLNVA at 10, 15, 20, 25 and 30 mM KCl was significantly (*P < .05, **P < .01) different from the control condition at 4.6 mM KCl (n = 8). B, TEA at 2.5, 5.0, 7.5 and 10 mM all significantly (*P < .05, **P < .01) inhibited GLNVA-induced relaxation of PE contraction. The tissues were exposed to TEA for 1 hr before PE contraction (n = 8).
Using ouabain (5.0 μM), the Na+-K+-ATPase inhibitor, to reduce the K+ gradient across the membrane by inhibition of the Na+-K+-ATPase pump, we found that ouabain (5.0 μM) could also inhibit the GLNVA relaxation effect in normal PSS (fig. 3B). In K+-free PSS, although the relaxation effect of GLNVA was the same as normal PSS inhibited by TEA (fig. 3, A and C), ouabain did not completely inhibit the relaxation effect (fig. 3D). Under these conditions, relaxation could still be obtained with other vasodilators, such as forskolin (1.0 μM, fig. 3, A and B).

Influence of TEA (10 mM) or ouabain (5.0 μM) on GLNVA-(1.0 μM) induced endothelium denuded guinea pig thoracic aorta relaxation under normal PSS or K+-free PSS condition. GLNVA-induced relaxation was inhibited by TEA (A) and Ouabain (B) under normal PSS condition. However, in the K+-free PSS condition, GLNVA-induced relaxation was almost abolished by TEA (C) but not completely by ouabain (D).
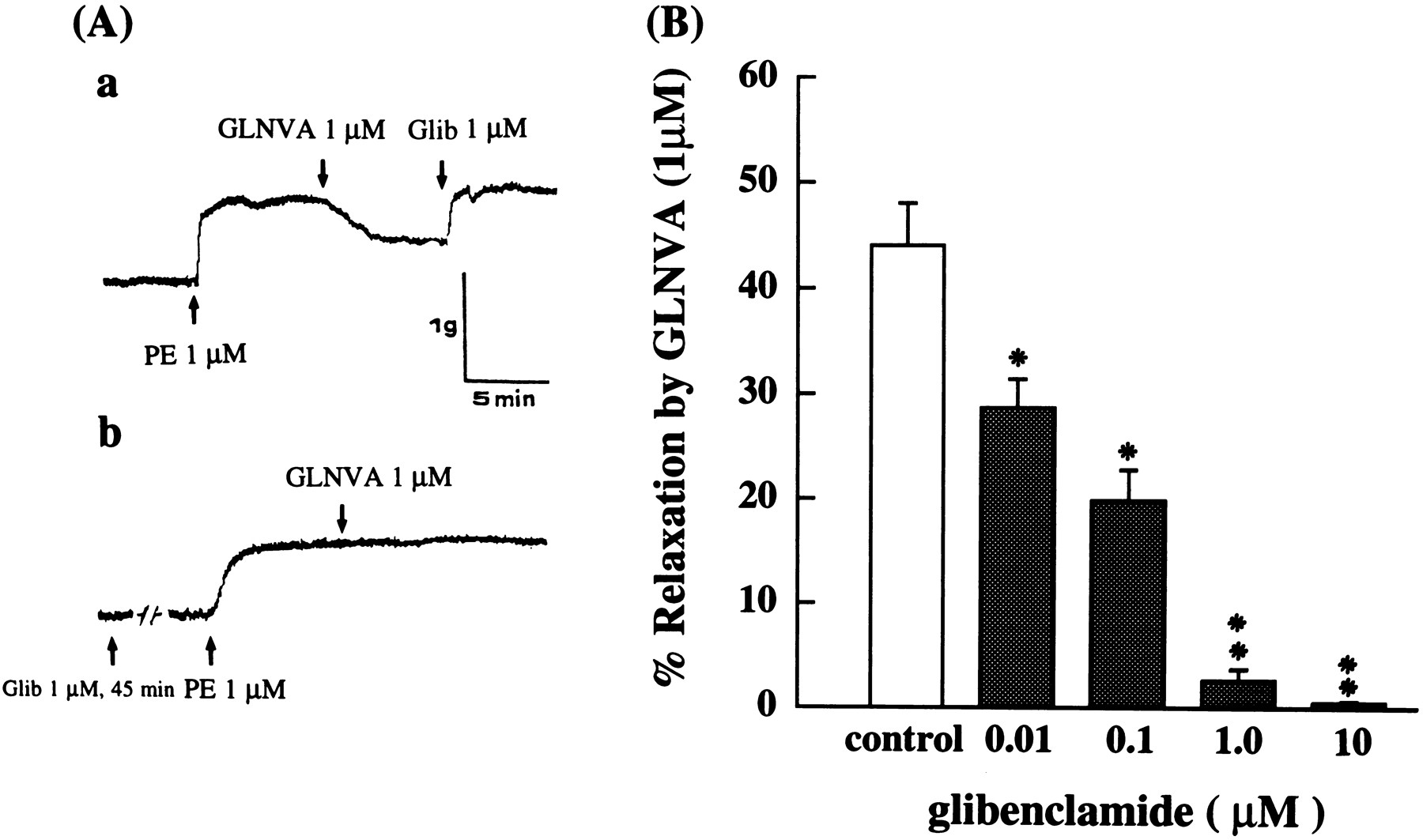
To investigate whether the relaxation effect of GLNVA involved the opening of ATP-sensitive K+ channel, the effects of its blocker glibenclamide were investigated. As shown in fig.4A-a, after GLNVA produced a relaxation response in PE-induced contraction, application of glibenclamide (1.0 μM) induced contraction. In contrast to these findings, the glibenclamide (1.0 μM), an ATP-sensitive K+ channel blocker, did block the relaxation response of GLNVA (1.0 μM) (fig. 4A-b), and glibenclamide (0.01–10 μM) dose dependently antagonized the relaxant effect of GLNVA (fig. 4B).

Effects of Glibenclamide (Glib, 1.0 μM) on GLNVA-(1.0 μM) induced relaxation of PE contraction on endothelium denuded guinea pig thoracic aorta. A-a, Glib reverses GLNVA-induced relaxation of PE contraction. A-b, The GLNVA-induced relaxation on PE-induced contraction is inhibited with preincubation with glibenclamide. B, Glibenclamide (0.01–10 μM) dose dependently antagonized the relaxant effect of GLNVA.
Cardiosensory effects in isolated guinea pig atrium.
GLNVA (0.01, 0.1, 1.0 and 10 μM; n = 7) caused concentration-dependent positive inotropic (fig. 5A) and chronotropic (fig. 5B) effects in the isolated spontaneously beating guinea pig right atrium and electrically driven left atrium. EC50 values of 0.56 ± 0.04 and 150.59 ± 6.62 μM were calculated for inotropic and chronotropic effects of right atrium, respectively. An EC50 value of 0.88 ± 0.05 μM was calculated for inotropic effect of left atrium.

Concentration-dependent effect of GLNVA-induced increasing percentage change of resting tension (A) and resting frequency (B) on guinea pig right and left atria, n = 7.
Pretreatment with ruthenium red (1.0 μM), a nonselective cation blocker of capsaicin receptor, for 30 min in the organ bath was found to block the cardiac capsaicin’s effect on the heart (Franco-Cerecedaet al., 1991). As shown in figure 6, A, B, and C, the effect of GLNVA (1.0 μM) on the right atrium was markedly depressed and the tension change of left atrium was also depressed.

Effects of GLNVA (1.0 μM) on guinea right (A and B) and left (C) atria after different pretreatments with ruthenium red (1.0 μM), human CGRP8–37 (1.0 μM), capsazepine (1.0 μM), capsaicin pretreatment (guinea pig was pretreated with 50 mg/kg, s.c. for 2 wk) or atenolol (1.0 μM) in separate groups. The effects of GLNVA on both atria are significantly inhibited (*P < .05) after pretreatment with ruthenium red or human CGRP8–37and completely abolished (**P < .01) by capsazepine or capsaicin pretreatment, n = 7.
To investigate, whether CGRP was involved in the stimulatory effect, animals were given a large dose of capsaicin (50 mg/kg, s.c.) for 2 wk to produce sensory denervation. This pretreatment causes an almost total loss of sensory SP- and CGRP-immunoreactive neuron within the heart, so responses to GLNVA of isolated right and left atrium were completely abolished. Capsazepine is a selective blocker of the capsaicin receptor (Franco-Cereceda and Lundberg, 1992). As shown in figure 6, the effects of GLNVA on the right and left atria were completely inhibited by capsazepine (10 μM) pretreatment. Even in pretreatment with human CGRP (8–37) (1.0 μM), a CGRP antagonist, the atrial effect of GLNVA was also markedly depressed. However, atenolol (1.0 μM) had no significant effect on the GLNVA-induced inotropic and chronotropic effects.
Immunoreactivity of CGRP from coronary circulation.
The stimulatory effects of GLNVA on the release of CGRP-LI from the isolated perfused guinea pig heart is shown in figure7A. In comparison with the control group, GLNVA (0.1, 1.0 and 10 μM) perfused for 20 min increased the release of CGRP-LI concentration dependently. In the desensitized and isolated guinea pig heart, CGRP-LI was not increased by GLNVA (1.0 μM) (fig. 7B).

A, Concentration-dependent effect of GLNVA on the outflow of CGRP-LI in the isolated guinea pig heart, n= 6. B, Effect of GLNVA on the outflow of CGRP-LI is significantly inhibited (**P < .01) in the isolated guinea pig heart from capsaicin pretreated guinea pig, n = 8.
Discussion
The clinical applications of capsaicin as an antinociceptive agent are sharply limited by the vagus reflex as well as the untoward bradycardia and apnea accompanying its intravenous administration (Donnerer and Lembeck, 1982). Those untoward effects of capsaicin are attributed to autonomic and sensory stimulation (Lo et al., 1994). Capsaicin-sensitive sensory neurons present in the mammalian heart (Lundberg et al., 1985; Saito et al., 1987) can release transmitters such as CGRP, neurokinin A and substance P (Maggi and Meli, 1987; Franco-Cereceda et al., 1988). It was reported that CGRP produces an inotropic effect, mediated by elevated cyclic AMP in rat atrium, and increase Ca++ currents in frog atrial cells (Ohmura et al., 1990). Capsaicin, similar to CGRP, also produced positive inotropic and chronotropic effects in the isolated guinea pig atrium (Franco-Cereceda and Lundberg, 1988). Because CGRP has been found to be present in the nerve fibers of sinoatrial node and right atrium in high density (Saito et al., 1986; Sugiyama et al., 1989), it may be released by capsaicin for the most part in the atrium and to a lesser degree in the ventricle. We assumed that because the positive inotropic actions of capsaicin and CGRP are primarily found at the atrial myocardium, it would therefore be of no value in treating congestive heart failure in which positive inotropic action on the ventricular myocardium is more favorable. However, CGRP has been found to improve cardiac performance in patients with congestive heart failure (Gennari et al., 1990). Recently, in the isolated rat heart, capsaicin was further reported to have antiischemic and antiarrhythmic effects (D’Alonzoet al., 1995). Our synthesized capsinolol also revealed CGRP releasing and β-adrenoceptor blocking activities in a previous study (Chen et al., 1996). In addition to their positive inotropic effect, such analogues of capsaicin as GLNVA with CGRP-releasing properties may be of value in their vasorelaxing benefits in the treatment of ischemic cardiovascular dysfunction. We have shown that our synthesized GLNVA has no significant vagus reflex response but retains the hypotensive cardiovascular effect of capsaicin (Yehet al., 1993; Lo et al., 1994). GLNVA appears to act in vivo more exclusively than capsaicin by stimulating peripheral perivascular small diameter C-fiber sensory nerves (Yehet al., 1993). However, the mechanism of vasorelaxation obtained from GLNVA remains unclear. In our study, we further examined the vasorelaxant and cardiac effects of GLNVA in in vitroand tried to provide some clues to its mechanisms.
CGRP, substance P, and capsaicin can all cause coronary vasodilation (Duckles, 1986; Vallotton et al., 1989; Nelson et al., 1990; Lo et al., 1995), but substance P-induced relaxation is subject to rapid tachyphylaxis and does not occur after removal of the endothelium (Furchgott, 1984; Duckles, 1986). The relaxation effects of CGRP and capsaicin are endothelium independent (Duckles, 1986; Vallotton et al., 1989; Lo et al., 1995). CGRP involved in the coronary vasodilatory effects of capsaicin is partly made effective by activating K+ channel (Franco-Cereceda et al., 1987). The effect of capsaicin on the ionic currents recently examined in A7r5 vascular smooth muscle cell suggest that inhibition of the voltage-depended L-type Ca2+ channel is also involved in the capsaicin-induced relaxation of vascular smooth muscle, whereas capsaicin-induced inhibition of voltage-dependent K+ channel might produce an increase in cell excitability (Lo et al., 1995). Therefore, the development of capsaicin analogues with less potency for block of Ik in vascular smooth muscle could potentially reduce their excitatory effects.
In our study, we have found that GLNVA causes an endothelium-independent vasorelaxant effect. In comparison with capsaicin (Lo et al., 1995), this provides considerable evidence of the important role of K+ permeability in the vascular smooth muscle relaxing action of GLNVA. Except in solutions of 80 mM K+, GLNVA causes relaxation of PE-induced contraction. As shown in figure 2A, although the extracellular K+ concentration was increased to reduce the K+gradient across the membrane, the ability of GLNVA to relax PE contraction was attenuated. TEA has been shown to cause depolarization (Haeusler and Thorens, 1980; Harder and Waters, 1984) and to block K+ current in isolated smooth muscle membrane patches (Benha et al., 1985). In this study, pretreatment with TEA (2.5–10 mM) also significantly inhibited GLNVA-induced relaxation (fig. 2B). TEA was able to block GLNVA relaxation of PE-induced contraction under normal PSS and zero K+-PSS. To abolish the K+ gradient effect via inhibition of Na+-K+-ATPase, the use of ouabain also inhibited GLNVA relaxation under normal PSS, but attenuated the inhibited effect under zero K+-PSS. Under zero K+-PSS condition, the K+ gradient across the membrane is increased toward infinity (outside K+, 0 mM; inside, ∼150 mM; Jones, 1980). Thus, the loss of K+gradient due to the action of ouabain probably remained incomplete under this condition (Meisheri et al., 1988), and GLNVA could use the existing enhanced K+ gradient to produce relaxation under the conduction of zero K+. The inhibition of the voltage-dependent L-type Ca++ channel is involved in the capsaicin-induced relaxation of the vascular smooth muscle; however, capsaicin-induced inhibition of voltage-dependent K+ channels might produce an increase in cell excitability (Lo et al., 1995). Therefore, these results suggest that the mechanism of GLNVA on vascular smooth muscle is significantly different from capsaicin. Because GLNVA has been found to induce CGRP-LI release from the coronary circulation, the vasodilatory effect of GLNVA may be caused by the release of CGRP, which has been previously described as a potent vasodilator (Brain et al., 1985). In our study, the relaxation induced by GLNVA could be blocked by ouabain, a Na+-K+ATPase inhibitor, and also by glibenclamide, an ATP-sensitive K+ channel blocker. These results indicate that ATP-sensitive K+ channel opening plays an important role in the relaxation response of GLNVA and is in contrast to capsaicin-induced blocking of voltage dependent K+ channel-associated cell excitability (Lo et al., 1995). The activation of ATP-sensitive K+channels by GLNVA may be a result of CGRP release which has been mentioned by Nelson et al. 1990), and thus both action pathways together make up substantial parts of vascular smooth muscle relaxation.
Capsaicin has been reported to evoke an increase in contractile rate and tension of the isolated right atrium simultaneously with an enhanced outflow of CGRP-LI released from the atrium; ruthenium red and capsazepine further inhibit capsaicin and resiniferatoxin-induced excitation of cardiac sensory nerves as revealed by inhibition of both CGRP-LI release and the cardiostimulatory action of capsaicin (Franco-Cereceda et al., 1991). Similar to capsaicin, GLNVA was found to increase the contractility of isolated guinea pig atrium. The role of CGRP in this effect was thus taken into consideration. To verify that cardiostimulating effect of GLNVA was the same as capsaicin mediated via the release of a CGRP-like peptide, the effects of different pretreatments on GLNVA-induced action and CGRP releasing effect of GLNVA were investigated. As shown in our study, the cardiosensory stimulatory effects of GLNVA are also abolished after pretreatments with ruthenium red, capsazepine and further sensory denervation with long-term application of a large dose of capsaicin, during which capsaicin sensitive-cardiac sensory neurons were denervated (Papka et al., 1984). Moreover, GLNVA induces a concentration-dependent increase in CGRP-LI outflow. Although less potent than is found in capsaicin, this effect disappeared in sensory denervated animals as described above. Based on these results, it is suggested that GLNVA can induce a capsaicin-like cardiostimulating effect mediated via the release of CGRP.
In conclusion, these results indicate that the positive inotropic and chronotropic effects of GLNVA were mediated via CGRP release from sensory neuron endings in the heart, and the vasorelaxant effect was mediated via CGRP release-associated potassium channel opening activity and eliminated capsaicin-derived cell excitability in blood vessel smooth muscles. When compared to the effects of capsaicin (Lo et al., 1995), in this study, GLNVA’s CGRP-releasing effect is the same as capsaicin’s, but GLNVA’s vasorelaxant effect is notably better. Instead of the eliminated K+ channel blocking activity of capsaicin, the K+ channel opening activity is predominant in GLNVA and plays an important role in enhancing its vasorelaxant effect. Although the authors are not trying to identify GLNVA with potential clinical utility as a positive inotropic agent, this vasorelaxant activity resulting from GLNVA and its associated CGRP release significantly points to the possibility of the use of some other relatively nonpungent capsaicin analogues in the future for the treatment of ischemic cardiovascular dysfunctions. Otherwise, dissection of the relaxation mechanism for GLNVA might also have benefited from experiments after depletion of sarcoplasmic reticulum Ca++ stores. Actually, capsaicin did not inhibit the function of intracellular Ca++ store in rat cardiac muscles and its action in blood vessels is still not clearly understood (Yamatoet al., 1996a). Presently, an electrophysiological study of GLNVA on K+ and Ca++ channels of smooth muscle is now being performed in our laboratory.
Acknowledgment
The authors thank Ms. Li-May Ann for excellent technical assistance.
Footnotes
-
Send reprint requests to: Dr. Ing-Jun Chen, Department of Pharmacology, Kaohsiung Medical College, Kaohsiung, Taiwan 807, R.O.C.
-
↵1 This work was supported by research Grants NSC 82-0412-B-037-093 and NSC 86-2314-B-037-086 from the National Science Council of the Executive Yuan, Taiwan, R.O.C.
- Abbreviations:
- CGRP-LI
- calcitonin-gene related peptide-like immunoreactivity
- EGTA
- ethylene glycol bis (β-aminoethyl ether) N, N′-tetraacetic acid
- Glib
- glibenclamide
- GLNVA
- glyceryl nonivamide
- PE
- phenylephrine
- PSS
- physiological salt solution
- TEA
- tetraethylammonium
- AMP
- adenosine monophosphate
- Received June 25, 1996.
- Accepted December 24, 1996.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}