


Abstract
Monomethylfumarate (MMF) is thought to be the bioactive ingredient of the drug Fumaderm (Biogen Idec, Cambridge, MA), licensed in Germany since 1994 for the treatment of moderate-to-severe psoriasis. Psoriasis is a common inflammatory hyperproliferative skin disorder that involves cross-talk between different cell types, including immune cells and keratinocytes. Psoriatic lesions are characterized by hyperproliferation, aberrant differentiation, and inflammation, with the psoriatic cytokine network maintained by communication between immune cells and keratinocytes. Recently, there is increasing evidence regarding the pivotal role of keratinocytes in mediating the disease process, and these cells can be regarded as safe therapeutic targets. From the data available on human subjects treated with Fumaderm, MMF is an effective antipsoriatic agent with known effects on immune cells. However, little is known about its direct effects on keratinocytes. We hypothesized that MMF has direct antiproliferative, prodifferentiative, and anti-inflammatory effects on keratinocytes. Indeed, MMF dose-dependently inhibited [3H]thymidine incorporation into DNA, indicating a direct antiproliferative action on keratinocytes. MMF significantly increased the protein level of keratin 10, the early keratinocyte differentiation marker, and the activity of transglutaminase, a late differentiation marker. These results are consistent with an ability of MMF to promote keratinocyte differentiation and inhibit proliferation, thereby improving psoriatic lesions. In 12-O-tetradecanoylphorbol-13-acetate (TPA)–induced keratinocytes, MMF significantly inhibited the expression of the proinflammatory cytokines, tumor necrosis factor-α (TNFα), interleukin-6, and interleukin-1α as well as the production of TNFα. Our results support the notion that MMF has direct antiproliferative, prodifferentiative, and anti-inflammatory effects on keratinocytes, highlighting its potential use as a multifactorial antipsoriatic agent.
Introduction
Fumaric acid esters (FAEs) have been known as antipsoriatic agents since 1959; however, their exact effects on different cell types as well as their mechanism of action are as yet unresolved. Fumaderm (Biogen Idec, Cambridge, MA) is a drug that has been licensed in Germany since 1994 and consists of a mixture of FAEs including dimethylfumarate (DMF) and three salts of monoethylfumarate (Rostami Yazdi and Mrowietz, 2008). FAEs have recently garnered interest in the United States, especially with various multicenter studies revealing the drug’s efficacy and limited safety concerns in most patients (Hodges et al., 2012). Although DMF is the main ingredient of the drug, DMF does not seem to be the active ingredient because it cannot be detected in the blood of patients, unlike monomethylfumarate (MMF), which has been detected in the blood for up to 36 hours after administration (Mrowietz et al., 1999).
Data have been published regarding the effects of FAEs (including MMF) on endothelial as well as immune cells; however, little is known about their effects on keratinocytes (Nibbering et al., 1993; Thio et al., 1994; Litjens et al., 2004b; Wallbrecht et al., 2011; Dehmel et al., 2014). Thus, MMF has known effects on immune cells, such as dendritic cells and granulocytes, which could possibly account in part for its antipsoriatic actions (Nibbering et al., 1993; Litjens et al., 2004b). In addition, FAEs (including MMF) exhibit inhibitory effects on the proliferation, differentiation, migration, and tube formation of endothelial cells (Arbiser, 2011; García-Caballero et al., 2011). However, the known effects of MMF on immune cells and endothelial cells may not adequately explain the superior results with Fumaderm in treating moderate to severe psoriasis, and which are accompanied by limited and nonserious side effects compared with other immune system–targeting drugs, such as biologic drugs (Mrowietz et al., 1998; Menter et al., 2008).
The skin maintains its barrier function through tight regulation of the proliferation and differentiation of keratinocytes, the primary cellular component of the epidermis. As keratinocytes begin to differentiate, they express early markers of keratinocyte differentiation, such as keratin 1 (K1) and keratin 10 (K10). Expression of K1 and K10, along with growth arrest, is considered the fundamental indicator of the switch from a proliferative basal phenotype to the postmitotic differentiating phenotype. As keratinocytes differentiate and express various differentiation-associated proteins, these proteins are cross-linked by the enzyme transglutaminase (TGase), which catalyzes the formation of the isopeptide bonds that maintain the insolubility of the stratum corneum and hence barrier integrity (Candi et al., 2005). In psoriasis, this differentiation process is dysregulated, and keratinocytes exhibit hyperproliferation and aberrant differentiation, which are two key hallmarks of psoriatic lesions (Roberson and Bowcock, 2010).
Keratinocytes are also considered a major source of a wide spectrum of proinflammatory mediators (Alappatt et al., 2000; Gröne, 2002), including cytokines, such as interleukin (IL)-1, IL-6, IL-7, IL-8, IL-10, IL-12, IL-15, IL-18, and IL-20 as well as tumor necrosis factor-α (TNFα) (Gröne, 2002), and chemokines, such as CXCL8–CXCL11 and CCL20 (Nestle et al., 2009). Cytokines and chemotactic factors produced by keratinocytes can initiate an inflammatory process in response to local irritation or injury. Thus, agents that have direct antiproliferative, prodifferentiative and anti-inflammatory effects on keratinocytes could serve as ideal antipsoriatic agents. In addition, keratinocytes possess cytokine receptors, so they are also targets for some cytokines. Indeed, certain cytokines can activate keratinocytes and lead to hyperproliferation and dysregulated differentiation and further induce the production of additional cytokines by these activated keratinocytes (Gröne, 2002). Accordingly, since there is little known about the actions of MMF on keratinocytes, we hypothesized that MMF exerts direct antiproliferative, prodifferentiative, and anti-inflammatory effects on epidermal keratinocytes. These effects may provide MMF an advantage over other current antipsoriatic agents that may have no or minimal effects on the regulation of keratinocyte growth and differentiation.
Data from this study demonstrate that MMF dose-dependently inhibited keratinocyte proliferation. We showed for the first time that MMF had prodifferentiative effects, enhancing the protein expression of K10 and TGase enzyme activity. Furthermore, we showed that MMF significantly inhibited the mRNA expression of TNFα, IL-6, and IL-1α in 12-O-tetradecanolylphorbol 13-acetate (TPA)–induced keratinocytes as well as TNFα secretion from these cells. These direct effects of MMF on keratinocytes, together with its known effects on immune cells and endothelial cells, may explain its superior clinical results and suggest that MMF is likely a multifactorial antipsoriatic agent.
Materials and Methods
Commercial keratinocyte serum-free medium (K-SFM) and the appropriate supplements were obtained from Gibco/Invitrogen (Grand Island, NY). Keratinocyte Basal Medium (KBM-Gold) was from Lonza, Inc. (Walkersville, MD). MMF was from Sigma-Aldrich (St. Louis, MO). [3H]Thymidine was from Moravek Biochemicals, Inc. (Brea, CA) and Ecolite scintillation fluid was from MP Biomedicals (Solon, OH). Protein assay products and mini-protean TGX precast gels were obtained from Bio-Rad (Hercules, CA). Total RNA was reverse transcribed to cDNA using iScript Reverse Transcription Supermix from Bio-Rad, and RNA purification kits were from 5 PRIME (Gaithersburg, MD). IL-1α, IL-6, and TNFα TaqMan probes were from Life Technologies (Grand Island, NY). Primary rabbit anti-K10 antibody was from Covance (Princeton, NJ). Primary monoclonal mouse anti–β-actin antibody was from Sigma-Aldrich. Secondary antibodies used for Western blot analysis were as follows: AlexaFluor IRDye 800–conjugated goat anti-rabbit IgG and IRDye 680–conjugated goat anti-mouse IgG secondary antibody from LI-COR (Lincoln, NE).
Primary Mouse Keratinocyte Culture.
Mouse epidermal keratinocytes were isolated from newborn male and female ICR CD1 outbred mice and cultured as previously described (Griner et al., 1999). Briefly, skins were harvested and floated on 0.25% trypsin at 4°C overnight prior to a brief incubation at 37°C. The epidermis and dermis were then mechanically separated and the keratinocytes were scraped from the underside of the epidermis. Cells were collected by centrifugation and seeded overnight in dialyzed serum-containing plating medium as described previously (Ernest Dodd et al., 2005). After incubation overnight, the medium was then replaced with K-SFM (50 µM CaCl2). The medium was replaced every other day and the cells were cultured until they reached the desired confluence for experimentation.
Normal Human Epidermal Keratinocyte Culture.
Adult normal human male and female epidermal keratinocytes (NHEK catalog no. 192627) were obtained from Lonza, Inc. and were subcultured according to the supplier’s instructions. Cells were plated in six-well plates and cultured in commercially available Keratinocyte Basal Medium (KBM-Gold medium) from Lonza, Inc. This medium contains a calcium concentration of 14.7 mg/l (99.986 µM) in the form of calcium chloride dihydrate and is supplemented with hydrocortisone, transferrin, epinephrine, gentamicin sulfate, amphotericin-B, bovine pituitary extract, recombinant human epidermal growth factor, and insulin. Supplements were supplied separately in quantities specified by the manufacturer and were added to the fresh medium before use. The medium was replaced every other day, and the cells were cultured until they reached the desired confluence for experimentation.
Measurement of Cell Proliferation by [3H]Thymidine Incorporation Assay.
Primary cultures of mouse keratinocytes or normal human keratinocytes (60–70% confluent) were treated with the indicated concentrations of MMF in K-SFM or complete KBM-Gold medium, respectively, for 24 hours. Control samples contained no MMF. Cells were then labeled with 1 µCi/ml [3H]thymidine for an additional 1 hour. Cells were washed twice with phosphate-buffered saline lacking divalent cations and the reactions terminated with ice-cold 5% trichloroacetic acid. Cells were washed sequentially with 5% trichloroacetic acid followed by deionized water and were solubilized in 0.3 M NaOH. An aliquot of this NaOH extract was counted in Ecolite scintillation fluid using a scintillation spectrometer (Beckman Coulter, Inc., Brea, CA).
Measurement of Cellular TGase Activity.
Keratinocytes (80–90% confluent) were treated with the indicated concentrations of MMF prepared in K-SFM or complete KBM-Gold medium. Control samples contained no MMF. Cells were harvested in homogenization buffer containing 0.1 M Tris-acetate (pH 8.5), 0.2 mM EDTA, 20 µM 4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride, 2 µg/ml aprotonin, 2 µM leupeptin, and 1 µM pepstatin, and TGase activity was assayed according to the technique of Folk and Chung (1985) with minor modifications as described in our previous publication (Griner et al., 1999).
Western Blot Analysis.
After treatment, cultured keratinocytes were solubilized with 330 µl warm lysis buffer containing 0.1875 M Tris (pH 8.5), 3% SDS, 1.5 mM EDTA, and water. Sample buffer containing 30% glycerol, 1% bromophenol blue, 15% β-mercaptoethanol, and 54% water was added to each cell lysate to constitute Laemmli buffer (Laemmli, 1970). Lysates were then briefly boiled and stored at −20°C. Equal volumes of protein were separated on 8–10% SDS gels and transferred to Immobilon-P membranes followed by blocking for 1 hour and incubation with primary antibodies overnight at 4°C. Membranes were then washed with Tris-buffered saline with 0.1% Tween and incubated with secondary antibodies for an hour at room temperature. Membranes were visualized using an infrared Odyssey imaging system (LI-COR Biosciences) and immunoreactive bands were quantified with the Odyssey software.
Quantitative Real-Time Polymerase Chain Reaction.
Primary mouse keratinocytes (80–90% confluent) were treated with different concentrations of MMF for 24 hours. At 22 hours, keratinocytes were treated for 2 hours with TPA (100 nM) to induce a proinflammatory response in keratinocytes (for a total of 24 hours of exposure to MMF). Total RNA was extracted using PerfectPure RNA tissue kits (5 PRIME, Inc.) as per the manufacturer’s protocol. The quality and quantity of total RNA were assayed using a NanoDrop instrument (NanoDrop Technologies, Wilmington, DE). The iScript cDNA synthesis kit (Bio-Rad) was used to reverse transcribe equal quantities of total RNA (1 μg) following the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using an ABI Step-One Plus Fast Real-Time PCR system (Applied Biosystems, Grand Island, NY) according to the manufacturer’s instructions with the recommended parameters. Each cDNA sample (5 µl; 125 ng) was added to 10 µl Fast Reagent Master Mix, 4 µl DNase-RNase free water and 1 µl TaqMan for a final volume of 20 µl and loaded into each well of a 96-well plate. Negative controls containing water instead of cDNA were performed to ensure purity of all reagents. Relative gene expression was calculated by the ΔΔCt method with glyceraldehyde-3-phosphate dehydrogenase used for normalization. The results were expressed as the fold difference in gene expression relative to the endogenous gene and compared with control samples. Values were then expressed as “fold regulation” as described in Yuan et al. (2006).
TNFα Protein Expression by Enzyme-Linked Immunosorbent Assay.
Mouse and human TNFα enzyme-linked immunosorbent assay (ELISA) kits from BD Biosciences (San Jose, CA) were used. Near-confluent mouse and human keratinocytes were treated with different concentrations of MMF (0, 300, 500, and 750 μM, and 1 mM) for 24 hours. Primary mouse and normal human keratinocytes were cotreated with or without 100 nM TPA for the final 2 hours or 8 hours, respectively (for a total of 24 hours of exposure to MMF). Cell culture supernatants were collected and stored at −80°C and subjected to a single freeze-thaw cycle. Reagents and standards were prepared according to the manufacturer’s instructions. Briefly, an aliquot of 50 μl of each sample was placed in each well. An aliquot of 50 μl ELISA diluent was added, and the plate was incubated for 2 hours at room temperature followed by aspiration of the contents of each well and washing five times with at least 300 μl of washing buffer. After the last wash, the plate was blotted on absorbent paper to remove any residual buffer. An aliquot of 100 μl of detection antibody was added to each well and the plate was incubated for 1 hour at room temperature. Aspiration of liquid and washing was performed as above. An aliquot of 100 μl enzyme working reagent (only for the mouse kit) was added to each well and incubated for 30 minutes at room temperature. A final aspiration/washing procedure was performed followed by the addition of 100 μl TMB One-Step Substrate Reagent and incubation for 30 minutes. Finally, an aliquot of 50 μl stop solution was added to each well and the optical density and protein content was detected in each sample (well) within 30 minutes, at a wavelength of 450 nm and corrected at 570 nm.
Statistical Analysis.
All experiments were performed independently and repeated a minimum of three times in duplicate. The values were statistically analyzed by one-way analysis of variance, with a Student–Newman–Keuls post hoc test, using Prism software (Graph Pad Software Inc., San Diego, CA) with statistical significance assigned at P < 0.05. All quantified data are expressed in the form of bar graphs with values representing means ± S.E.M.
Results
Inhibition of Keratinocyte Proliferation with Different Concentrations of MMF.
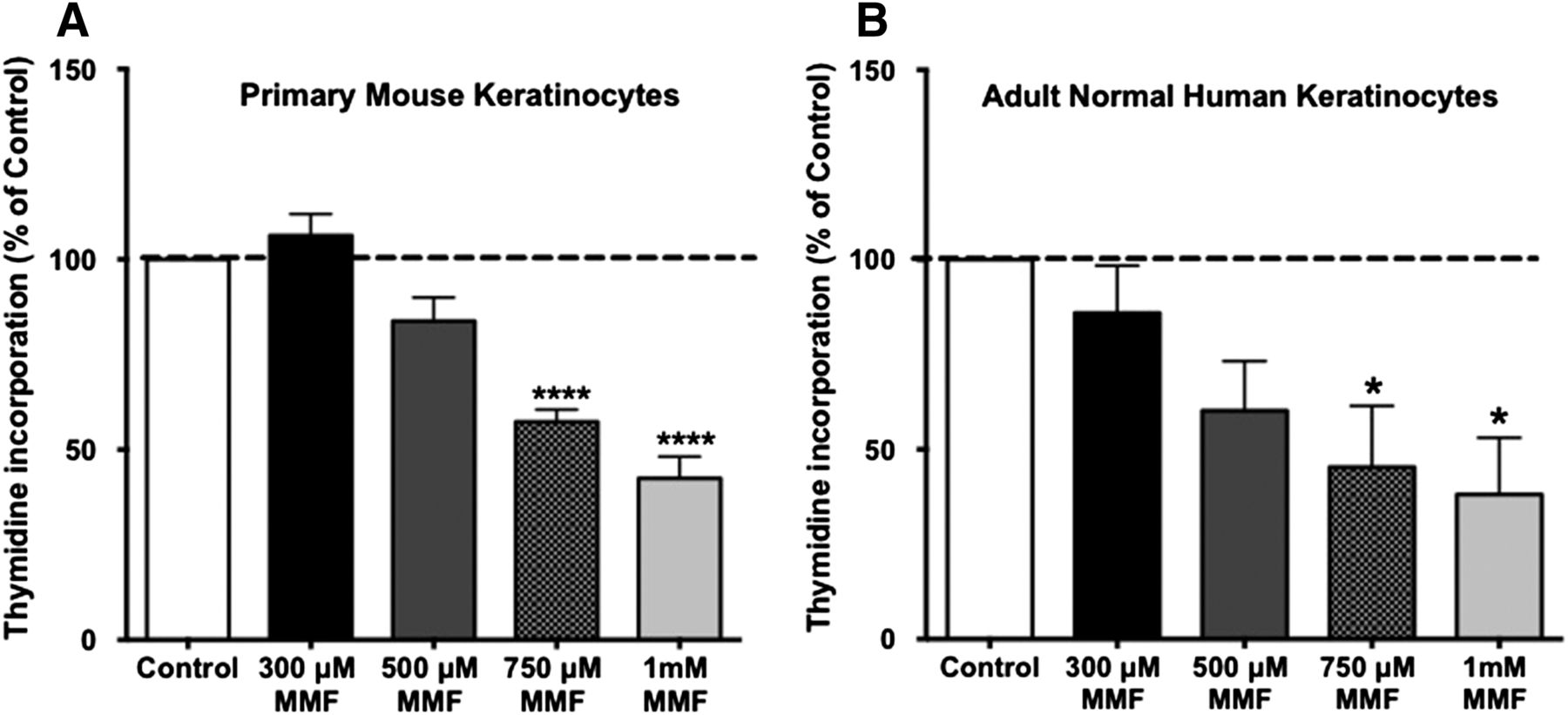
We initially wished to confirm the antiproliferative effect of MMF, which was previously demonstrated by cell counting (Thio et al., 1994), using an alternative proliferation assay. Keratinocytes were incubated for 24 hours with medium only or medium containing different concentrations of MMF (300, 500, and 750 μM, and 1 mM), and the ability of MMF to inhibit keratinocyte proliferation was examined by monitoring the incorporation of [3H]thymidine into DNA. As shown in Fig. 1, MMF significantly inhibited the proliferation of actively growing normal keratinocytes in a dose-dependent manner. Figure 1A shows the antiproliferative effect of MMF on primary mouse keratinocytes, whereas Fig. 1B illustrates a similar effect in adult normal human keratinocytes. This result suggests that MMF inhibits keratinocyte proliferation in both species, an effect that would presumably be beneficial in improving hyperproliferative psoriatic lesions in human patients.
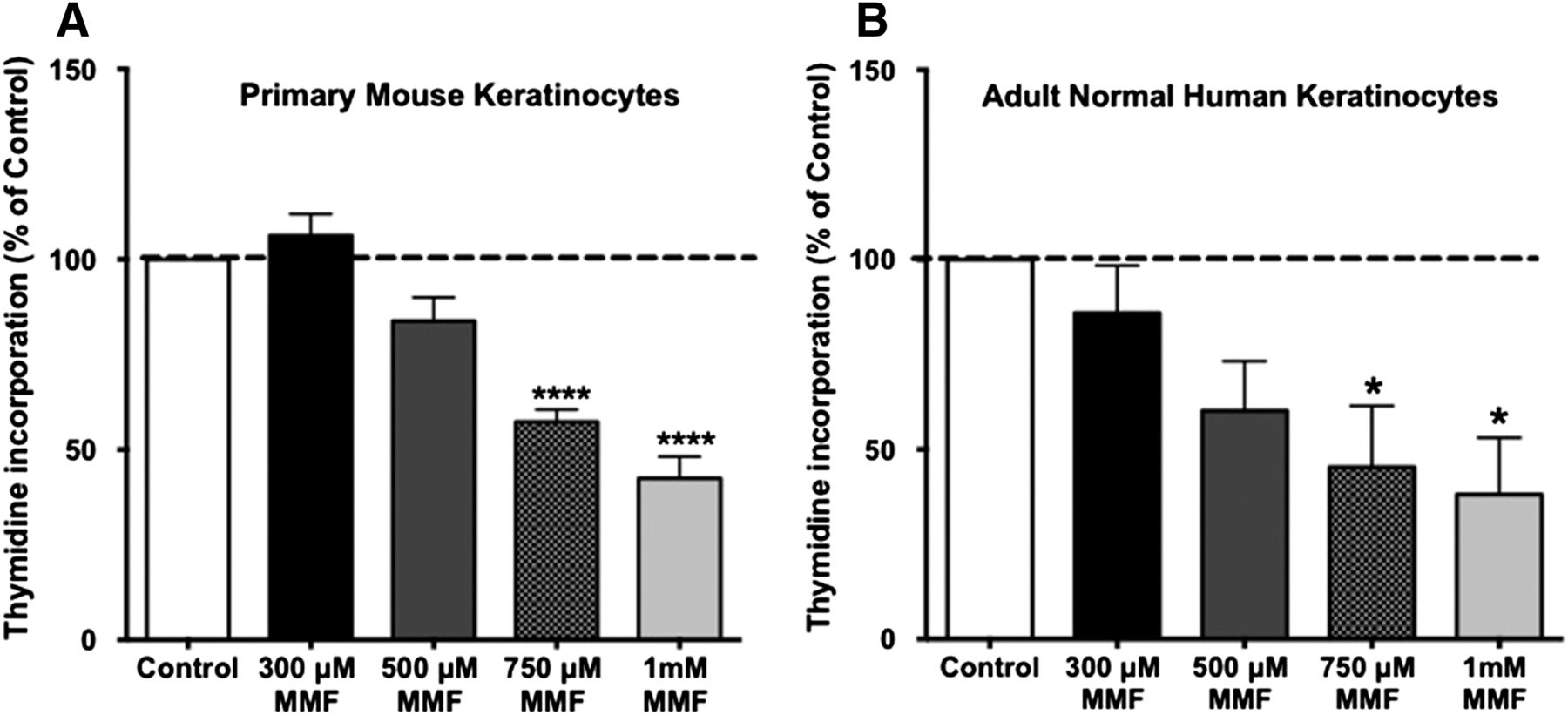
MMF exerted an antiproliferative effect on keratinocytes. Near-confluent primary mouse keratinocytes (A) or normal human epidermal keratinocytes (B) were treated with the indicated concentrations of MMF (or no MMF for the control) in K-SFM or complete KBM-Gold, respectively, for 24 hours. [3H]Thymidine incorporation was measured as in Griner et al. (1999). Values, expressed as means ± S.E.M., represent the percentage over the control of five separate experiments performed in duplicate. *P ≤ 0.05; ****P ≤ 0.0005 versus the control. For primary mouse keratinocytes, the mean (± S.E.M.) value of the control was 14,043 ± 2042 cpm/well. For adult normal human keratinocytes, the mean control value was 170,478 ± 7378 cpm/well.
Induction of Keratinocyte Differentiation by MMF.
Differentiation is another important factor required for epidermal homeostasis, and psoriasis is characterized by abnormal keratinocyte differentiation (Nestle et al., 2009). We next investigated whether the antiproliferative effect of MMF is accompanied by a prodifferentiative action. We tested for the effect of MMF on two keratinocyte differentiation markers that represent two different stages of the keratinocyte differentiation process: K10 protein levels (an early marker of keratinocyte differentiation) and TGase enzyme activity (a late marker of keratinocyte differentiation) (Eckert et al., 2005; Helwa et al., 2013). To investigate the ability of MMF to stimulate early differentiation, near-confluent cultures of primary mouse keratinocytes were treated with different concentrations of MMF for 24 hours; then cell lysates were prepared and Western blot analysis was used to detect the levels of K10 in MMF-treated and untreated cells. Our data indicate that a 24-hour treatment with MMF significantly increased K10 protein expression, with a maximum effect at a 500 µM concentration (Fig. 2). The activity of TGase, measured as the incorporation of the radiolabeled substrate [3H]putrescine into casein, was then monitored to investigate the capacity of MMF to induce late differentiation in keratinocytes. Near-confluent cultures of primary mouse keratinocytes and adult normal human keratinocytes were treated with different concentrations of MMF for 24 hours, and TGase enzyme activity assay was monitored as described in Materials and Methods. MMF induced a significant increase in TGase activity in both mouse and human keratinocytes (Fig. 3) with a maximum elevation at the 1 mM concentration. These data suggest that at lower concentrations, MMF triggers early differentiation of keratinocytes and at higher concentrations, MMF induces keratinocytes to terminally differentiate. Thus, MMF has antiproliferative and prodifferentiative effects on keratinocytes, stimulating both early and late markers of keratinocyte differentiation, a result that suggests that this drug may be able to correct the imbalance between proliferation and differentiation in psoriatic keratinocytes.

MMF at a lower concentration induced the protein expression of K10, the early marker of differentiation. Near-confluent primary mouse keratinocytes were treated with 0, 300, 500, and 750 µM, and 1 mM MMF for 24 hours. Cells were then harvested and proteins were analyzed by Western blotting. Cytokeratin 10 levels were determined using rabbit anti-K10 antibody (Covance) and the LI-COR Odyssey system. (A) A representative blot is shown. (B) The quantification and statistical analysis of the results of four separate experiments are shown, with values normalized to actin levels and shown as the means ± S.E.M. **P ≤ 0.005 versus the control.

Higher concentrations of MMF increased TGase activity, a late marker of keratinocyte differentiation. Near-confluent cultures of primary mouse keratinocytes (A) and normal human keratinocytes (B) were treated with the indicated concentrations of MMF for 24 hours and TGase activity was assayed. For each experiment, the TGase activity in treated keratinocytes was normalized to protein content and then to the TGase activity of control (untreated) cells. Data are expressed as the percentage of control and represent the means ± S.E.M. of at least three separate experiments in duplicate. *P ≤ 0.05 versus the control. For primary mouse keratinocytes, the mean value of the control was 3075 ± 1171 cpm/µg protein. For adult normal human keratinocytes, the mean control value was 3523 ± 183 cpm/µg protein. In addition, a positive control (1 mM calcium-containing medium) performed with the same cells yielded an approximate 444 ± 48% increase in TGase activity in mouse keratinocytes and 195 ± 34% in human keratinocytes.
MMF Has an Inhibitory Effect on Cytokine Production by Keratinocytes.
Keratinocytes play an active role in the production of inflammatory mediators (cytokines and chemokines) in the psoriatic cytokine network (Steinhoff et al., 2001; Gröne, 2002; Nestle et al., 2009). Accordingly, we hypothesized that MMF exerts a unique anti-inflammatory action by inhibiting major inflammatory mediators produced not only by immune cells (Litjens et al., 2004b) but also by keratinocytes. We opted to examine the effect of MMF on the expression of TNFα, IL-1α, and IL-6 in keratinocytes treated with TPA, which is known to activate keratinocytes and induce inflammatory mediator release (Kim et al., 2014). Near-confluent primary mouse keratinocytes were treated with different concentrations of MMF for 24 hours. At 22 hours, cells were spiked with 100 nM TPA (for 2 hours), and mRNA was isolated. qRT-PCR results showed that MMF significantly inhibited the TPA-induced expression of TNFα (Fig. 4A), IL-6 (Fig. 4B), and IL-1α (Fig. 4C), thus revealing a specific anti-inflammatory effect of this drug on keratinocytes. Moreover, we confirmed the qRT-PCR results with TNFα using an ELISA assay to monitor the secretion of TNFα from both primary mouse and normal human keratinocytes. Near-confluent primary mouse keratinocytes were treated as above and the supernatants were collected for assay by ELISA. The secretion of TNFα by MMF-pretreated cells activated with TPA was significantly inhibited, with levels returned almost to basal levels. For normal human keratinocytes, near-confluent cells were treated with different concentrations of MMF for 16 hours and cells were then cotreated with 100 nM TPA for an additional 8 hours, prior to ELISA assay of the supernatants. Our data showed that MMF also inhibited TNFα production in TPA-activated human keratinocytes compared with keratinocytes treated with TPA only. The inhibition in the case of human cells was so effective that TNFα levels in the supernatants of MMF-pretreated cells were below the limit of detection (2 pg/ml) of the ELISA kit (Fig. 5).

MMF downregulated the mRNA expression of inflammatory cytokines in keratinocytes treated with TPA. Near-confluent cultures of primary mouse keratinocytes were treated with 0, 300, 500, or 750 µM, or 1 mM MMF for 22 hours. Cells were then cotreated with or without 100 nM TPA for 2 hours (for a total of 24 hours of exposure to MMF). RNA was isolated and transcribed to cDNA, and qRT-PCR was performed to determine the mRNA levels of TNFα (n = 6), IL-6 (n = 3), and IL-1α (n = 3). Values represent the means ± S.E.M. of 3–6 separate experiments performed in duplicate. **P ≤ 0.005; ***P ≤ 0.0005 versus TPA alone; $P ≤ 0.05; $$P ≤ 0.005 versus control (untreated) cells.

MMF inhibited the production of TNFα by keratinocytes treated with TPA. Near-confluent cultures of primary mouse keratinocytes and normal epidermal keratinocytes were treated with 0, 300, 500, or 750 µM, or 1 mM MMF for 22 and 16 hours, respectively. Cells were then cotreated with or without 100 nM TPA for an additional 2 hours and 8 hours, respectively (for a total of 24 hours of exposure to MMF). TNFα ELISA assays were performed and values represent the means ± S.E.M. of three separate experiments performed in duplicate. **P ≤ 0.005; ***P ≤ 0.0005 versus TPA alone; $P ≤ 0.05 versus control (untreated) cells. Please note that initially we treated both primary mouse keratinocytes and adult normal human keratinocytes with TPA for the same length of time (2 hours); however, the TNFα levels were undetectable in the control untreated human keratinocytes as well as in the MMF-treated human cells. Therefore, we increased the time of incubation to 8 hours in an unsuccessful attempt to bring the TNFα levels in the control and/or MMF-treated samples into the detectable range.
Discussion
DMF is considered the essential ingredient of the antipsoriatic drug Fumaderm. However, DMF does not seem to be the bioactive ingredient because it is not detected in patients’ blood for a sufficient length of time to be exerting an action on its own (Litjens et al., 2004a). Furthermore, according to previous clinical and experimental studies, most of the DMF is hydrolyzed to MMF, which, in turn, is thought to act as an antipsoriatic agent (Rostami Yazdi and Mrowietz, 2008; Rostami-Yazdi et al., 2009). Therefore, in our study, we focused on the bioactive ingredient of the drug MMF, rather than the main ingredient, an apparent prodrug with low bioavailability, using MMF concentrations that have recently been examined in other cell types (Bozard et al., 2012; Ananth et al., 2013; Promsote et al., 2014).
We show for the first time that MMF has direct prodifferentiative and anti-inflammatory effects on keratinocytes. Furthermore, using a reliable proliferation assay (incorporation of [3H]thymidine into DNA), we confirmed previous published results concerning the antiproliferative effects of MMF (Thio et al., 1994). Our results show that MMF significantly inhibited proliferation with a greater than 50% reduction in DNA synthesis at the highest concentration (1 mM; Fig. 1). This effect was observed in both primary mouse as well as normal human keratinocytes. However, to date, no studies have investigated the effect of MMF on keratinocyte differentiation. To investigate the effect of MMF on keratinocyte differentiation, we selected two differentiation markers that are relevant to the abnormality in psoriatic skin and that reflect two different stages of the differentiation process. K10 is one of the first proteins expressed by keratinocytes and reflects the fundamental switch from a proliferative basal phenotype to the postmitotic phenotype; this switch is abnormal in psoriasis, in which diseased keratinocytes maintain their proliferative capacity into suprabasal layers (Candi et al., 2005). K10 can be regarded as the “keratinization marker of keratinocytes” (Moll et al., 2008), and it has been reported that K10 is greatly reduced in psoriatic skin (Iizuka et al., 2004). We observed that MMF significantly induced K10 protein expression at a concentration of 500 µM (Fig. 2). The biphasic effect of MMF on K10, with concentrations greater than 500 µM inducing less of an increase in K10 levels, is likely because higher concentrations of MMF and with the long treatment interval (24 hours) drive keratinocytes to late differentiation. At later stages of differentiation, a decline in K10 expression is expected, accompanied by an increase in TGase activity, a late marker of differentiation, as shown in Fig. 3.
TGase is another important protein that facilitates terminal keratinocyte differentiation and the formation of the cornified envelope (stratum corneum) (Eckert et al., 2005). Thus, K10 and TGase delineate the effect of MMF not only on initial keratinocyte differentiation but also on the terminal stage involving stratum corneum formation and hence barrier integrity. Furthermore, psoriatic lesions have previously been shown to exhibit aberrant epidermal TGase distribution (Candi et al., 2002), and inhibition of TGase in mouse skin leads to hyperproliferation and parakeratosis (Harrison et al., 2007). MMF significantly increased TGase enzyme activity at a 1 mM concentration by almost 2-fold (200%) in primary mouse keratinocytes and about 1.5-fold (150%) in adult normal human keratinocytes.
We also investigated the effect of MMF on the third major hallmark of psoriasis: inflammation. TNFα, IL-6, and IL-1α are key cytokines in the psoriatic cytokine network and can be secreted by immune cells as well as keratinocytes, thus amplifying their proinflammatory potential. These cytokines are also capable of activating keratinocytes (Nestle et al., 2009). Awareness of the role of TNFα in the psoriatic cytokine network has led investigators to the development of antipsoriatic biologic drugs that target TNFα, such as Etanercept (Griffiths, 2010). Moreover, the serum levels of TNFα and IL-6, another proinflammatory cytokine, are significantly higher in psoriasis patients compared with healthy controls (Arican et al., 2005). IL-1α is a known regulator of keratinocyte proliferation and differentiation, as well as immune function (Taniguchi et al., 2014). Indeed, IL-1α overexpression in the basal epidermal layer (in an IL-1α transgenic mouse model) produces a psoriasis-like phenotype with keratinocyte hyperproliferation and immune cell infiltration (Groves et al., 1995; Gudjonsson et al., 2007). Furthermore, with the extensive cross-talk between keratinocytes and immune cells in psoriasis, it is unclear which is the major source of these cytokines, and the role of activated keratinocytes in cytokine production cannot be excluded. In fact, keratinocytes are thought to be the primary producers of IL-1 in the skin (Brotas et al., 2012). To stimulate cytokine production by keratinocytes, we used TPA, which is a known inducer of the keratinocyte inflammatory response (Kim et al., 2014). However, one caveat of our study is the use of normal keratinocytes, since exposure of psoriatic keratinocytes to an inflammatory milieu may alter their responses to various agents including possibly MMF. Nevertheless, with the technical difficulty of obtaining sufficient psoriatic keratinocytes to perform all of the reported experiments, the absence of an available psoriatic keratinocyte cell line, and the clinical success of Fumaderm in psoriatic patients, our results can be regarded as a proof of concept regarding the effects of MMF in keratinocytes. Future studies are required to validate these data in psoriatic keratinocytes. Nevertheless, our results show that MMF was a potent inhibitor of inflammatory cytokine expression by TPA-activated keratinocytes. As mentioned earlier, in an inflammatory response, keratinocytes are a major source of a spectrum of different cytokines including IL-6 and TNFα (Gröne, 2002; Nestle et al., 2009). By stimulating keratinocytes with TPA, we mimicked the initiation phase of the psoriatic process (Helwa et al., 2013), and MMF was capable of efficiently inhibiting this stimulation. This conclusion was reached based on qRT-PCR and ELISA findings. In the case of human keratinocytes, MMF treatment reduced the TPA-induced TNFα secretion to undetectable levels, reflecting the effective anti-inflammatory action of MMF on keratinocytes. Thus, we can conclude that MMF inhibits proliferation, induces differentiation, and decreases the expression/production of cytokines by keratinocytes and hence can presumably improve psoriatic lesions by downregulating the psoriatic cytokine network and restoring keratinocyte homeostasis (Fig. 6).

The role of keratinocytes in the psoriatic cytokine network and the effect of MMF in ameliorating the disease process. This figure illustrates the interactions between keratinocytes and immune cells, which mediate the initiation, progression, and maintenance of the inflammatory process in psoriasis. A stimulus, such as trauma or an infection, can initiate an inflammatory response in genetically susceptible individuals. Keratinocytes are capable of initiating an immune response through the production of various proinflammatory cytokines, which feed back on the keratinocytes, inducing further activation and production of cytokines. Activated keratinocytes exhibit hyperproliferation and abnormal differentiation. MMF is capable of counteracting these effects to restore the balance between proliferation and differentiation as well as exerting an anti-inflammatory effect, all of which would improve psoriatic symptoms.
Most available antipsoriatic drugs target immune cells and/or mediators, and most researchers and dermatologists believe that targeting these cells and molecules is the ultimate goal for an antipsoriatic agent. However, considering the essential role of keratinocytes in the psoriatic cytokine network, targeting keratinocytes as well will amplify the drug’s effect and may not only be effective but also safe, because there will be less compromise of the patient’s immune system. Indeed, hyperproliferation, absent or abnormal differentiation, and inflammation (inflammatory cytokine production and immune cell infiltration) are the three major hallmarks of psoriasis. A drug with a pleiotropic mode of action that can target all three of these aspects should be considered the ultimate therapeutic strategy to treat psoriasis. Considering the already-proven clinical efficacy of Fumaderm (Mrowietz et al., 1998), studies to determine the effects and mechanism of action of this drug certainly seem warranted.
A major question remains concerning the mechanism of action of MMF, and studies are in progress in our laboratory to investigate the possible signaling pathways that underlie the effects of MMF. Among these possibilities is the G protein–coupled receptor GPR109A (HCA2). Tang et al. (2008) reported that MMF is an agonist of GPR109A. However, little is known about the physiologic functions of GPR109A in the skin and the possible mechanisms activated by its binding of MMF. In addition, in vitro and animal studies, as well as clinical data, suggest that MMF upregulates the antioxidative transcription factor Nrf2 (Onderdijk et al., 2014; Promsote et al., 2014). Our group is currently attempting to address these questions.
To our knowledge, this is the first report to show a direct prodifferentiative, anti-inflammatory effect of MMF on keratinocytes. Moreover, we also confirmed the antiproliferative effect of MMF on keratinocytes. These effects seem to account, at least in part, for its action as an antipsoriatic agent. These findings, in addition to previously published data, suggest that MMF is exerting inhibitory functions on the three major cell types involved in the pathogenesis of psoriasis (keratinocytes, immune cells, and endothelial cells) (Nibbering et al., 1993; García-Caballero et al., 2011). Together with the clinical data obtained for the antipsoriatic drug Fumaderm (Mrowietz et al., 1998; Rostami Yazdi and Mrowietz, 2008), used in Germany since 1994, these results suggest that MMF may be an ideal therapy for treating moderate to severe psoriasis. Our findings may help to pique interest in the antipsoriatic actions of MMF, which may encourage physicians in other countries (including the United States) to investigate the use of the drug for the treatment of psoriasis. These data also suggest that MMF may be effective for treating other hyperproliferative skin disorders. In addition, a drug containing DMF as its major ingredient, Tecfidera or BG-12 (Ruggieri et al., 2014) is currently available on the US market for the treatment of multiple sclerosis (Nicholas et al., 2012; Salmen and Gold, 2014). Thus, our results suggest the possibility of repurposing this already US Food and Drug Administration–approved drug for the benefit of psoriatic patients.
Acknowledgments
The authors thank Purnima Merai for excellent technical assistance in preparation of the mouse epidermal keratinocytes.
Authorship Contributions
Participated in research design: Helwa, Patel, Karempelis, Kaddour-Djebbar, Choudhary, Bollag.
Conducted experiments: Helwa, Patel, Karempelis, Kaddour-Djebbar, Choudhary.
Performed data analysis: Helwa, Bollag.
Wrote or contributed to the writing of the manuscript: Helwa, Kaddour-Djebbar, Choudhary, Bollag.
Footnotes
- Received July 30, 2014.
- Accepted October 17, 2014.
This research was supported by the National Institutes of Health National Institute of Arthritis and Musculoskeletal and Skin Diseases [Grant R01-AR45212]; and the US Department of Veterans Affairs [Merit Award CX000590]. W.B.B. is supported by a Department of Veterans Affairs Research Career Scientist Award. The contents of this article do not represent the views of the Department of Veterans Affairs or the US Government.
Abbreviations
- DMF
- dimethylfumarate
- ELISA
- enzyme-linked immunosorbent assay
- FAE
- fumaric acid ester
- IL
- interleukin
- K1
- keratin 1
- K10
- keratin 10
- K-SFM
- keratinocyte serum-free medium
- MMF
- monomethylfumarate
- qRT-PCR
- quantitative real-time polymerase chain reaction
- TGase
- transglutaminase
- TNFα
- tumor necrosis factor α
- TPA
- 12-O-tetradecanoylphorbol-13-acetate
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}