


Abstract
Vascular calcification is a mortality risk factor for stage 5 chronic kidney disease patients. We investigated the role of phosphorus and vitamin D analogs in the pathogenesis of vascular calcification using in vivo, ex vivo, and in vitro models. Our results demonstrate that uremic rats receiving a hyperphosphatemia-inducing diet did not exhibit aortic calcification despite elevated levels of serum phosphorus and calcium-phosphorus (CaxP) product. The vitamin D analog 1α-hydroxyvitamin-D2 [1α(OH)D2] at 0.17 μg/kg raised serum calcium, phosphorus, CaxP product, and aortic calcification in the uremic rats, but 19-nor-1α,25(OH)2D2 (19-nor) at the same dose had no significant effect. At 0.67 μg/kg, both 1α(OH)D2 and 19-nor had similar effects on serum calcium, phosphorus, and CaxP product, but only 1α(OH)D2 induced significant aortic calcification. Only aortic rings from 1α(OH)D2-treated uremic rats exhibited a significant increase in 45Ca uptake ex vivo. When aortic rings from normal rats or a primary culture of human coronary artery smooth muscle cells were treated with phosphorus or vitamin D analogs in vitro, high phosphorus induced calcium accumulation and/or 45Ca uptake in a dose- or time-dependent manner, whereas vitamin D analogs including 1α(OH)D2 up to 100 nM had no significant effect despite the presence of a functional vitamin D receptor. However, serum from 1α(OH)D2-treated uremic rats induced 45Ca uptake into smooth muscle cells cultured in high phosphorus. These results suggest that the regulation of vascular calcification in vivo cannot be easily replicated in the ex vivo or in vitro models, and high phosphorus and some vitamin D analogs such as 1α(OH)D2 exert interactive effects on modulating vascular calcification.
Chronic kidney disease (CKD) patients experience a high mortality rate from cardiovascular (CV) diseases (Zebrack et al., 2003; Weiner et al., 2004). Vascular calcification, the deposition of calcium phosphate mineral in cardiovascular tissues, contributes to CV morbidity and mortality in CKD patients (Blacher et al., 2001; London et al., 2003; Moe et al., 2004). Two distinct forms of calcification have been reported in the vessels (Jeziorska et al., 1998; Shanahan et al., 1999). Intimal calcification is often associated with inflammation and atherosclerosis (Jeziorska et al., 1998), whereas medial calcification occurs in smooth muscle cells (Shanahan et al., 1999). Stage 5 CKD patients experience 2- to 5-fold more coronary artery calcification than age-and gender-matched individuals (Braun et al., 1996). Moreover, histologic analysis shows that coronary plaques in patients with stage 5 CKD are characterized by increased media thickness and marked calcification, suggesting that smooth muscle cell calcification plays a critical role (Schwarz et al., 2000; Moe et al., 2005).
The mechanisms involved in vascular calcification are still largely unknown. Recently it has been postulated that vascular calcification is a regulated process involving a variety of factors including hyperphosphatemia, loss of calcification inhibition, induction of bone formation in smooth muscle cells, cell death, and circulating nucleational complexes derived from bone (Raggi et al., 2002; Giachelli, 2004; Nishizawa et al., 2005; Ritz and Gross, 2005). Many proteins such as matrix Gla protein, fetuin-A, osteoprotegerin, and pyrophosphatase, have been shown to be involved in this process (Giachelli, 2004). Although it is generally recognized that elevated serum phosphorus levels contribute to vascular calcification, medial calcification is also seen in diabetics and with aging in the presence of normal serum phosphorus (Everhart et al., 1988). Therefore, the role of phosphorus on calcification requires more studies.
1,25-Dihydroxyvitamin D3 and its analogs, such as 19-nor-1α,25(OH)2D2 (paricalcitol) or 1α-hydroxyvitamin-D2, that activate vitamin D receptor (VDR) in vivo are commonly used to manage secondary hyperparathyroidism associated with CKD (Martin and Gonzalez, 2004). Recent retrospective clinical observations show that vitamin D analogs provide a survival benefit for stage 5 CKD patients with an effectiveness order of paricalcitol > calcitriol > no vitamin D analog therapy, independent of the PTH and calcium levels (Teng et al., 2003, 2005; Nakai et al., 2004); the survival benefit of vitamin D analogs is associated with a decrease in cardiovascular-related mortality (Shoji et al., 2004). More recently, results from Dialysis Outcomes and Practice Patterns Study time-dependent analyses suggest that the survival benefit is in the order of i.v. paricalcitol > i.v. calcitriol > oral vitamin D analogs = no vitamin D analogs (Young et al., 2005).
The observation that vitamin D analogs provide a survival benefit in CKD seems contradictory to the general perception that these drugs, because of their potential impact on serum phosphorus and calcium, may cause medial layer calcification in vessels (Rostand and Drueke, 1999). Information from the literature regarding how different vitamin D analogs affect vascular calcification has not been forthcoming. Furthermore, it is not known how vitamin D analogs interact with phosphorus on modulating calcification. In the present study, we used the subtotally nephrectomized uremic rat model to examine the effect of phosphorus and vitamin D analogs on aortic calcification in vivo. We also prepared aorta tissue from normal or uremic rats to study the interactive effects of phosphorus and vitamin D analogs on calcification ex vivo. To investigate further the involvement of phosphorus and vitamin D analogs in calcification, we tested the effects of phosphorus and three different vitamin D analogs on calcium accumulation and 45Ca uptake in human vascular smooth muscle cells. We went on to collect serum from uremic rats treated with vitamin D analogs and examined whether these serum samples affected smooth muscle cell calcification in vitro. Our results demonstrate that phosphorus and some vitamin D analogs are involved interactively in modulating calcification.
Materials and Methods
Materials. 1α,25-Dihydroxyvitamin D3 [1α,25(OH)2D3, calcitriol, Calcijex], 1α,25-dihydroxyvitamin D2 [1α,25(OH)2D2, the active form of doxercalciferol], 1α-hydroxyvitamin D2 [1α(OH)D2, doxercalciferol, Hectorol], and 19-nor-1α,25-dihydroxyvitamin D2 [19-nor-1α, 25(OH)2D2, 19-nor, paricalcitol, Zemplar] were from Abbott Laboratories. Other reagents were of analytical grade.
Subtotally Nephrectomized Rats. The five-sixths nephrectomized (SNX) uremic rats were obtained from Charles River Laboratories, Inc. (Wilmington, MA). The nephrectomy was performed on male Sprague-Dawley rats with a standard two-step surgical ablation procedure. Approximately 2 weeks after the surgery, the rats were put on a hyperphosphatemia-inducing diet containing 0.9% phosphorus and 0.6% calcium (Slatopolsky et al., 2003) for 4 weeks, followed by treatment with vehicle (5% ethanol + 95% propylene glycol, 0.4 ml/kg), 19-nor or 1α(OH)D2 at indicated doses i.p. three times per week for 12 (n = 4-8 per group) or 40 days (n = 4-5 per group). Twenty-four hours after the last dose, animals were anesthetized with ketamine, and blood and aortic tissue were collected.
Measurement of Blood Chemistry. Serum total calcium, blood-ionized calcium, serum phosphorus, creatinine, and BUN concentrations were measured using an Abbott Aeroset (Abbott Park, IL). Serum PTH was measured using a rat intact PTH enzyme-linked immunosorbent assay kit obtained from ALPCO/Immutopics, Inc. (Windham, NH).
Tissue Preparation and von Kossa Staining. Aorta was collected into liquid nitrogen for quick freezing. Aorta was postfixed in 10% formalin, processed overnight, and followed by embedding into paraffin. Sections were stained by the von Kossa method and counterstained with hematoxylin.
Measurement of Calcium and Phosphorus Contents in Aorta from Five-Sixths Nephrectomized Rats. Aorta tissues were dried at 90°C for 4 h and weighed. Samples were incinerated to ash at 900°C for 8 h using an electric muffle furnace (type F62700; Barnstead International, Dubuque, IA) and then were dissolved in 6 N HCl with lanthanum chloride (0.1%). The solutions were diluted with distilled water, and calcium and phosphorus concentrations were measured using an autoanalyzer (Aeroset; Abbott).
Measurement of 45Ca Uptake into Aorta Isolated from Five-Sixths Nephrectomized Rats. Aorta from uremic rats treated with drugs was denuded, cut open into 2- to 3-mm ring strips, and placed in DMEM (with 0.9 mM phosphorus) containing 0.2 μCi/ml 45Ca for 3 days at 37°C in a 5% CO2 atmosphere. Afterward, the aortic strips were washed three times in PBS. The tissues were dried in an oven, weighed, and then dissolved in a solution containing 6% H2O2 and 56% HClO4 (final concentrations). Radioactivity was measured by liquid scintillation.
Measurement of 45Ca Uptake into Aorta Isolated from Normal Rats. Aorta was cut open and denuded and then cut into 2- to 3-mm ring strips and placed in DMEM containing different concentrations of test agents for 6 days at 37°C in a 5% CO2 atmosphere. Media containing the appropriate test agents were changed every 3 days. Then, 0.2 μCi/ml 45Ca was added, and the incubation continued for another 3 days. Afterward, the tissues were processed as described above, and radioactivity was measured by liquid scintillation.
Cell Culture. Primary culture of human coronary artery smooth muscle cells (Cambrex, Walkersville, MD) were grown in smooth muscle growth medium-2 containing 5% fetal bovine serum at 37°C in a humidified 5% CO2-95% air atmosphere. Cells were grown to >80% confluence and used within five passages. Primary culture of human coronary artery endothelial cells (Cambrex) were grown to confluence in EGM-2 Bullet Kit medium containing 2% fetal bovine serum (Cambrex) at 37°C and used within four passages.
Measurement of 45Ca Uptake into Cultured Smooth Muscle or Endothelial Cells. Smooth muscle cells were cultured in DMEM containing different concentrations of test agents for 5 days at 37°C in a 5% CO2 atmosphere. Media containing the appropriate test agents were changed every 3 days. Then, 0.2 μCi/ml 45Ca was added, and the incubation was continued for another day. Cells were then washed with PBS and solubilized in 0.1 N NaOH. Radioactivity was measured by liquid scintillation. In the coculture studies, endothelial and smooth muscle cells were initially cultured separately with endothelial cells in inserts. Cells were then put together in DMEM containing test agents, and 45Ca uptake was determined as described above.
Measurement of Calcium Content in Cultured Smooth Muscle Cells. Smooth muscle cells were cultured in DMEM containing different concentrations of test agents for different periods of time (as indicated) at 37°C in a 5% CO2 atmosphere. Media containing the appropriate test agents were changed every 3 days. Cells were then washed with PBS two times and extracted with 0.6 N HCl for 24 h at room temperature, and the calcium content in the extract was determined using the Calcium CPC LiquiColor test kit (StanBio, Boerne, TX). The remaining cells were washed with PBS and solubilized in 0.1 N NaOH containing 0.1% SDS. The protein content was determined using the BCA protein assay kit (Pierce Chemical, Rockford, IL).
Real-Time Reverse Transcription Polymerase Chain Reaction. Real-time reverse transcription (RT)-polymerase chain reaction (PCR) was performed with an iCycler (Bio-Rad, Hercules, CA). Each sample has a final volume of 25 μl containing 100 ng of cDNA, 0.4 μM concentrations each of the forward and reverse PCR primers, and 0.1 μM TaqMan probe for the gene of interest (Applied Biosystems, Foster City, CA). Temperature conditions consisted of a step of 5 min at 95°C, followed by 40 cycles of 60°C for 1 min and 95°C for 15 s. Data were collected during each extension phase of the PCR reaction and analyzed with the software package (Bio-Rad). Threshold cycles were determined for each gene.
SDS-Polyacrylamide Gel Electrophoresis and Western Blot Analysis. Cells (1 × 106 cells per sample) pretreated with or without test agents were solubilized in 100 μl of SDS-polyacrylamide gel electrophoresis sample buffer (Invitrogen, Carlsbad, CA), and the protein content in each sample was determined by the Pierce Chemical BCA protein assay. Samples were resolved by SDS-polyacrylamide gel electrophoresis using a 4 to 12% NuPAGE gel (Invitrogen), and proteins were electrophoretically transferred to polyvinylidene difluoride membrane for Western blotting. The membrane was blocked for 1 h at 25°C with 5% nonfat dry milk in PBS-T and then incubated with a rabbit anti-VDR (Active Motif Inc., Carlsbad, CA) polyclonal antibody (500-fold dilution) in PBS-T overnight at 4°C. The membrane was washed with PBS-T and incubated with a horseradish peroxidase-labeled anti-rabbit antibody for 1 h at 25°C. The membrane was then incubated with detection reagent (SuperSignal WestPico; Pierce Chemical). Specific bands were visualized by exposing the paper to Kodak BioMax films. Band intensity was quantified by Quantity One (Bio-Rad).
Data Analysis. One-way ANOVA Dunnett's test with 95% confidence intervals of difference was used to determine a p value among treatment groups. Statistical comparisons between two treatment groups were performed by unpaired t test with 95% confidence intervals of difference.
Results
In the first study, uremic rats with hyperphosphatemia were treated with 0.17 μg/kg 1α(OH)D2 [the precursor of 1α,25(OH)2D2, which is activated by conversion to 1α,25(OH)2D2 in the liver] and 19-nor [an analog of 1α,25(OH)2D2] at three times per week i.p. for 40 days. As shown in Fig. 1, the serum creatinine and BUN levels were significantly elevated in the SNX rats. Both 19-nor and 1α(OH)D2 at 0.17 μg/kg had no significant effect on creatinine or BUN (versus own group, Day 0). Figure 2 shows that 19-nor and 1α(OH)D2 effectively suppressed serum PTH throughout the treatment period. Serum calcium trended lower in the SNX (uremic)-vehicle group on Day 41, whereas serum phosphorus tended to be higher. As shown in Fig. 3, there was no significant difference in the aortic calcium and phosphorus contents among sham, SNX-vehicle, and 19-nor-treated groups, whereas 1α(OH)D2 treatment resulted in a significant elevation in the aortic calcium (36-fold) and phosphorus (36.2-fold versus vehicle) content. Figure 4 shows representative sections of aorta stained by von Kossa staining. No staining was detected in either vehicle or 19-nor-treated groups. In contrast, extensive staining was observed in the medial layer of aorta in the 1α(OH)D2-treated group, indicating the presence of calcification. These results suggest that 1α(OH)D2, but not 19-nor, exhibited significant effects on raising serum calcium, phosphorus, and the CaxP product, which is consistent with a previous report by Slatopolsky et al. (2002). Furthermore, only 1α(OH)D2 induced aortic calcification.

Serum creatinine and BUN levels in SNX uremic rats consuming the hyperphosphatemia-inducing diet. Rats were dosed with vehicle, 1α(OH)D2, or 19-nor at 0.17 μg/kg i.p. three times per week for 40 days. Handling of animals was as described under Materials and Methods. On days 0, 13, and 41 after the drug treatment, blood samples were collected for the measurement of serum creatinine (A) and BUN (B). Mean ± S.E. was calculated for each group (n = 4-5). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. *, p < 0.05; **, p < 0.01 versus sham day 0; #, p < 0.05 versus own group day 0.
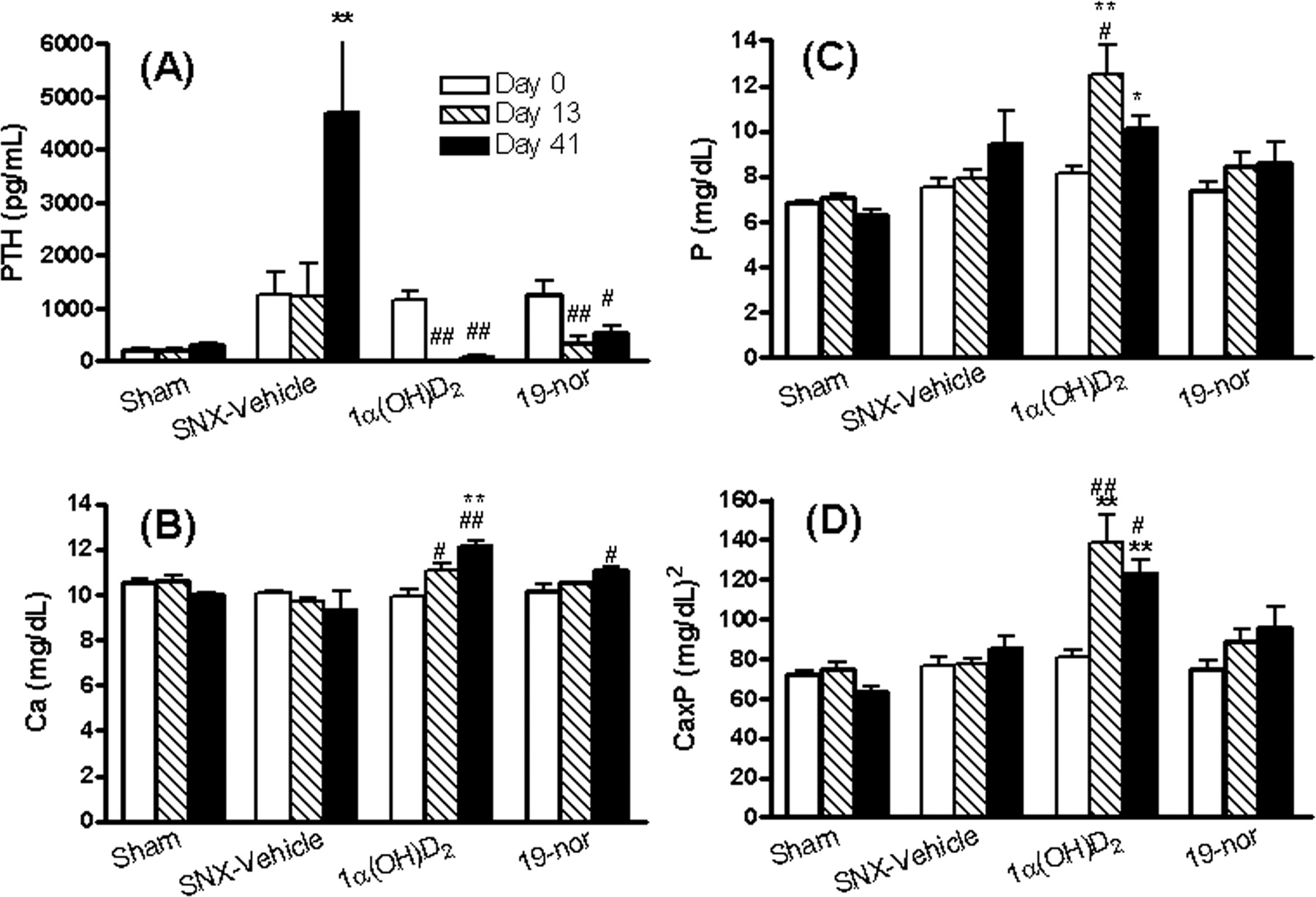
The serum calcium, phosphorus, CaxP product, and PTH levels in uremic rats. Rats were dosed with vehicle, 1α(OH)D2, or 19-nor at 0.17 μg/kg i.p. three times per week for 40 days as in Fig. 1. On days 0, 13, and 41 after the drug treatment, blood samples were collected for the measurement of serum PTH (A), calcium (B), phosphorus (C), and CaxP product (D). Mean ± S.E. was calculated for each group (n = 4-5). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. *, p < 0.05; **, p < 0.01 versus sham day 0; #, p < 0.05; ##, p < 0.05 versus own group day 0.

The calcium and phosphorus contents in the aorta of uremic rats. Rats were dosed with vehicle, 1α(OH)D2, or 19-nor at 0.17 μg/kg i.p. three times per week for 40 days as in Fig. 1. Twenty-four hours after the last dose, aorta was collected and processed for the determination of calcium (A) and phosphorus (B) as described under Materials and Methods. Mean ± S.E. was calculated for each group (n = 4-5). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. **, p < 0.01 versus sham.
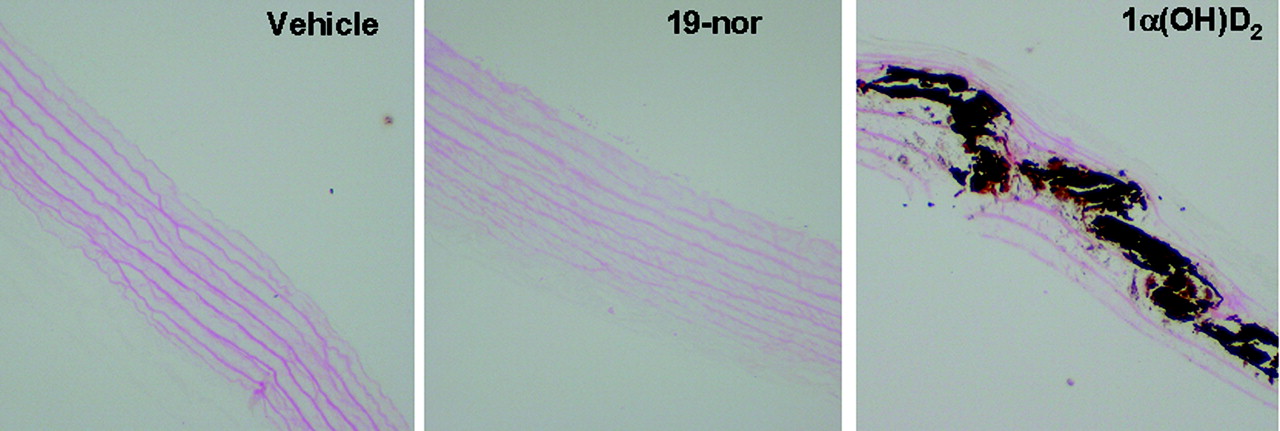
von Kossa staining in aorta. Uremic rats were dosed with vehicle, 1α(OH)D2, or 19-nor at 0.17 μg/kg i.p. three times per week, for 40 days as in Fig. 1. Twenty-four hours after the last dose, aorta was collected and processed for von Kossa staining.
Although 19-nor has been shown to suppress PTH in uremic rats at doses that did not affect serum calcium or phosphorus (Slatopolsky et al., 1995), 19-nor could be hypercalcemic at very high doses (Takahashi et al., 1997). To investigate whether the difference in aortic calcification was due to the differential effect of 19-nor and 1α(OH)D2 on serum calcium, phosphorus, and CaxP product, we tested a substantially higher dose (0.67 μg/kg) of 19-nor and 1α(OH)D2 in the hyperphosphatemic uremic rats using the same dosing paradigm (three times per week) and a 12-day treatment period. The studies were not carried out to 40 days because of the fact that animals did not survive well after prolonged treatment with vitamin D analogs at this high dose, especially in the 1α(OH)D2-treated group. The blood chemistry results are summarized in Table 1. Serum creatinine and BUN levels were significantly elevated in SNX-vehicle, which was not affected by either 19-nor or 1α(OH)D2 (versus SNX-vehicle). Significant decreases in serum calcium and an elevation in serum phosphorus were observed in the SNX-vehicle group (versus sham). The serum CaxP product trended upward without achieving statistical significance. As expected, 19-nor and 1α(OH)D2 at 0.67 μg/kg produced similar elevations in serum calcium, phosphorus, and CaxP product (versus sham). Both compounds effectively suppressed PTH. Figure 5 shows that the calcium and phosphorus contents in aorta were not different for sham, SNX-vehicle, and the 19-nor-treated groups. In contrast, 1α(OH)D2 produced a significant increase in the aortic calcium (10.4-fold) and phosphorus (15.2-fold versus SNX-vehicle) contents. As a control, we have also treated uremic rats consuming a normal diet (0.9% phosphorus and 1.13% calcium) with 19-nor and 1α(OH)D2 at 0.34 μg/kg for 12 days. No aortic calcification was observed in either SNX-vehicle or drug-treated animals (data not shown). These results suggest that an increase in serum phosphorus and calcium do not necessarily produce aortic calcification in uremic rats, and 19-nor and 1α(OH)D2 exert differential effects on aortic calcification independent of serum phosphorus and calcium levels.
Blood chemistry results in five-sixths nephrectomized uremic rats Five-sixths nephrectomized uremic rats consuming a hyperphosphatemia-inducing diet were treated with vehicle, 1α (OH)D2, or 19-nor at 0.67 μg/kg i.p. three times per week for 12 days. Data are means ± S.E. (n = 4 in sham, 6 in SNX-vehicle, 8 in the 1α(OH)D2-treated group, and 6 in the 19-nor-treated group). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons.

Calcium and phosphorus contents in the aorta of uremic rats dosed with 1α(OH)D2 or 19-nor at 0.67 μg/kg. Rats were dosed with vehicle or drugs i.p. three times per week for 12 days as in Table 1. Twenty-four hours after the last dose, aorta was collected and processed for the determination of calcium (A) and phosphorus (B) as described under Materials and Methods. Mean ± S.E. was calculated for each group (n = 4-8 as in Table 1). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. **, p < 0.01 versus sham.
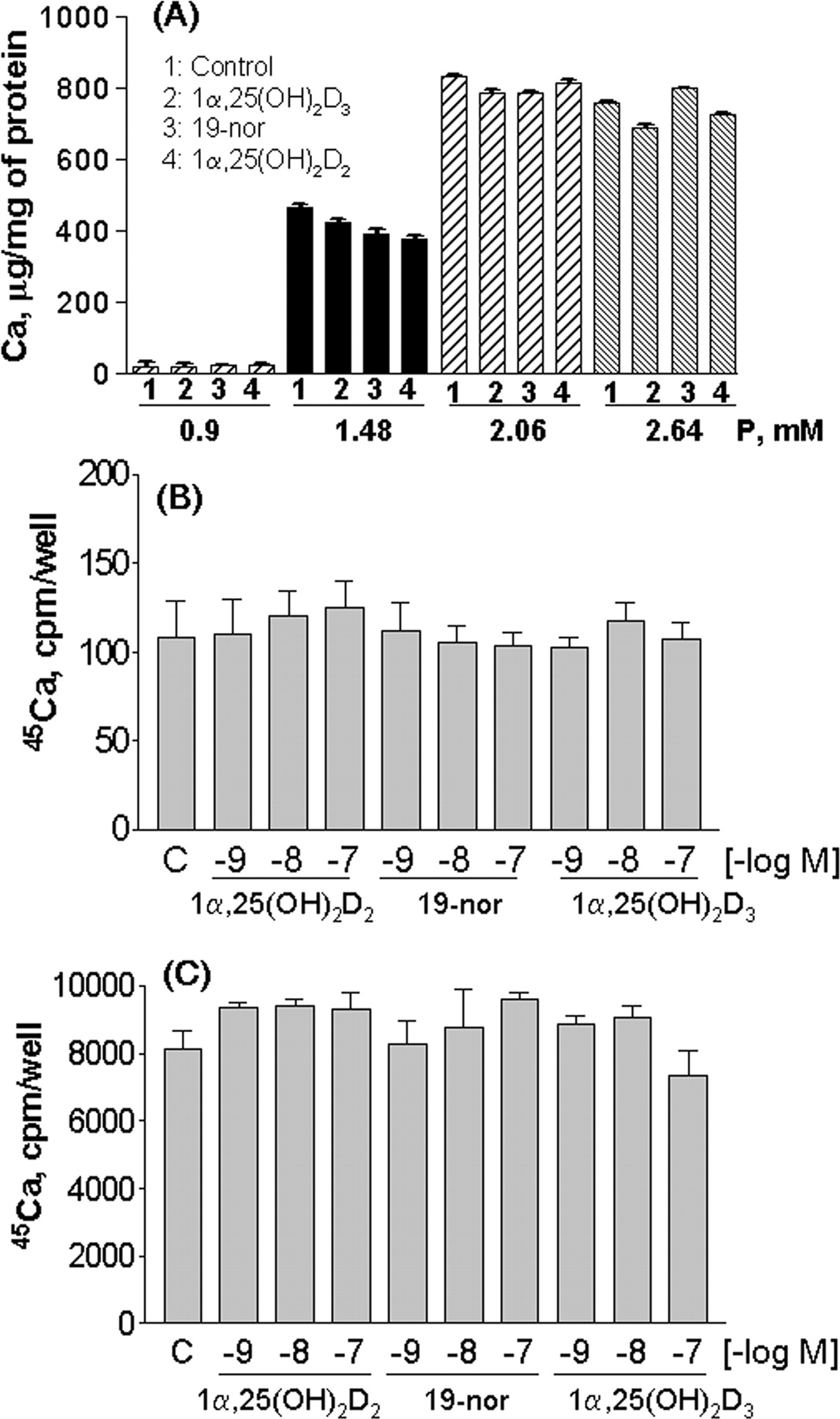
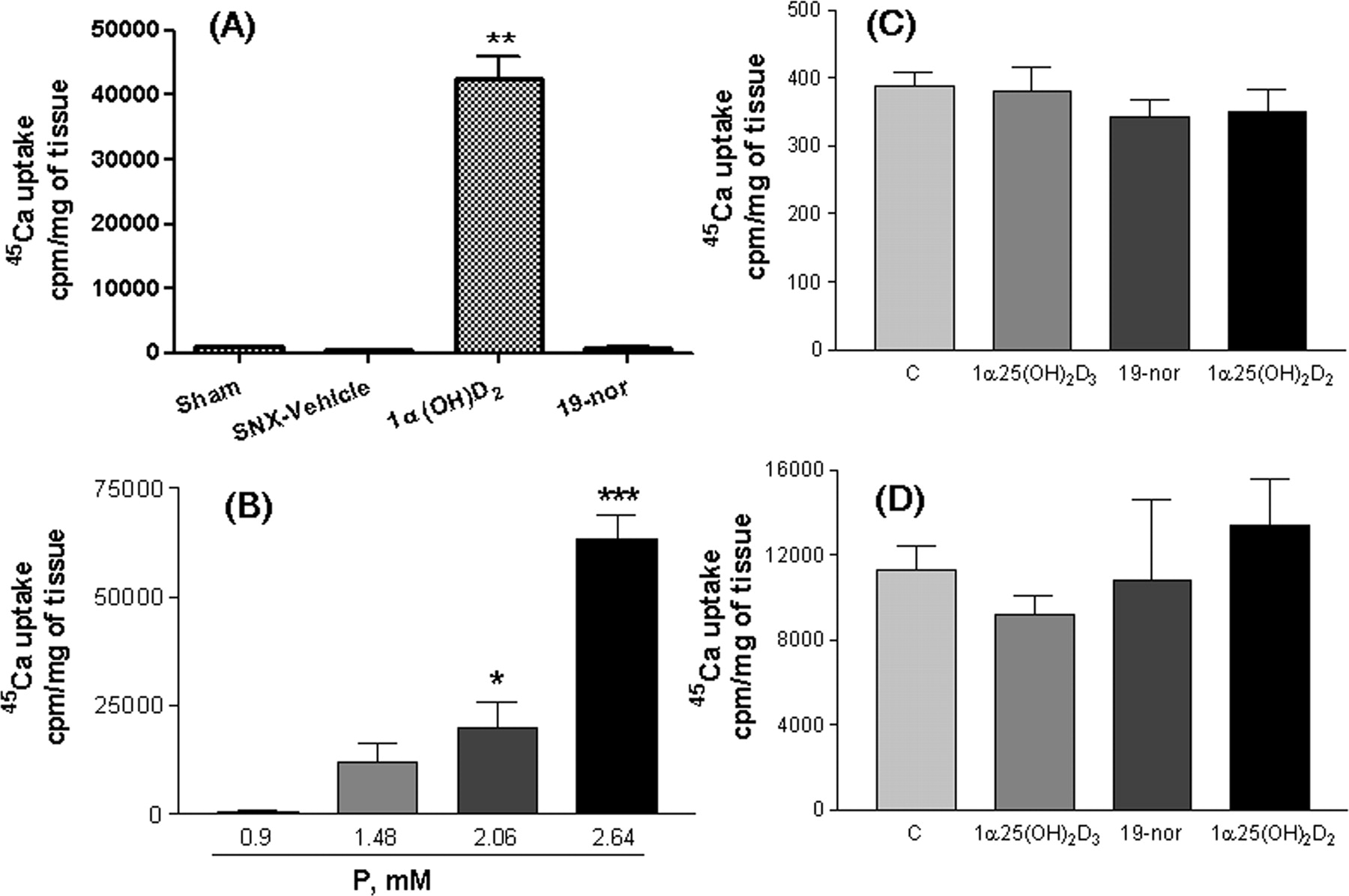
To further investigate the different effects of phosphorus and vitamin D analogs on aortic calcification, we examined 45Ca uptake into aortic ring strips prepared from the hyperphosphatemic uremic rats treated with vehicle, 19-nor, or 1α(OH)D2 at 0.67 μg/kg (i.p. for 12 days, three times per week). In these studies, the aortic tissue was denuded of the endothelial layer to minimize potential variability among samples. Consistent with the in vivo observations shown above, a significant increase in 45Ca uptake (40-fold versus sham) was observed in aorta prepared from the 1α(OH)D2-treated uremic rats, whereas minimal 45Ca uptake occurred in aorta prepared from the sham, SNX-vehicle, or 19-nor-treated uremic rats (Fig. 6A). In contrast, when aorta was prepared from normal rats and then treated with phosphorus and/or vitamin D analogs ex vivo, the results were strikingly different. Figure 6B shows that an increase in phosphorus in the culture medium significantly increased 45Ca uptake into aortic rings from normal rats in a dose-dependent manner. However, 1α,25(OH)2D, 19-nor, or 1α,25(OH)2D3 had no effect on 45Ca uptake in the presence of either 0.9 mM (Fig. 6C) or 2.06 mM phosphorus (Fig. 6D).
Taken together, the above results suggest that phosphorus and vitamin D analogs exert contradictory effects on aortic calcification in vivo versus ex vivo. Although high phosphorus does not necessarily cause aortic calcification in the in vivo situation, it profoundly induces calcification in aorta from normal rats ex vivo. As a comparison, vitamin D analogs have no effect on calcification ex vivo but display differential effects on aortic calcification in the hyperphosphatemic uremic rat.

Uptake of 45Ca into denuded aortic rings. A, aortic rings were prepared from uremic rats dosed with vehicle, 1α(OH)D2, or 19-nor at 0.67 μg/kg i.p. three times per week, for 12 days as in Table 1. 45Ca uptake was determined as described under Materials and Methods. Mean ± S.D. was calculated for each group (n = 4-8 animals/group as in Table 1). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. **, p < 0.01 versus sham. For B to D, aortic rings were prepared from normal rats and processed, and 45Ca uptake was determined as described under Materials and Methods. B, aortic rings from normal rats (n = 4) were cultured in DMEM containing different concentrations of phosphorus plus 3 U/ml alkaline phosphatase. C, aortic rings from normal rats (n = 6) were cultured in DMEM containing 0.9 mM phosphorus with or without 100 nM 1α,25(OH)2D3, 19-nor, or 1α,25(OH)2D2. D, aortic rings from normal rats (n = 3) were cultured in DMEM containing 2.06 mM phosphorus and 3 U/ml alkaline phosphatase with or without 100 nM 1α,25(OH)2D3, 19-nor, or 1α,25(OH)2D2. Mean ± S.D. was calculated for each group. One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. *, p < 0.05; ***, p < 0.001 versus 0.9 mM phosphorus (B) or control (C and D).
To further understand these observations, we tested phosphorus and vitamin D analogs in primary cultures of human coronary artery smooth muscle cells (SMCs). As shown in Fig. 7A, increasing the phosphorus concentration from 0.9 to 2.64 mM induced a dose-dependent increase in the cellular calcium content in SMCs. Figure 7B shows that the effect of phosphorus on calcium content in the cells was time-dependent. Consistently, Fig. 7C shows that an increase in the phosphorus concentration from 0.9 to 3.22 mM dose-dependently increased 45Ca uptake into these cells, reaching a plateau at 2.06 mM phosphorus. In contrast to the effects of phosphorus, as shown in Fig. 8A, 1α,25(OH)2D2, 19-nor, or 1α,25(OH)2D3 at 100 nM had no effect on the cellular calcium content when cells were cultured in medium containing phosphorus ranging from 0.9 to 2.64 mM. In addition, 1α,25(OH)2D2, 19-nor, or 1α,25(OH)2D3 at 1 to 100 nM had no effect on 45Ca uptake at either 0.9 (Fig. 8B) or 2.06 mM phosphorus (Fig. 8C).

Effect of phosphorus on calcium content and 45Ca uptake in cultured human coronary artery smooth muscle cells. A, human coronary artery smooth muscle cells were cultured in medium containing different concentrations of phosphorus plus 3 U/ml alkaline phosphatase for 9 days. B, cells were cultured in medium containing 0.9 or 2.06 mM phosphorus plus 3 U/ml alkaline phosphatase for different periods of time. C, cells were cultured in medium containing different concentrations of phosphorus plus 3 U/ml alkaline phosphatase for a total of 6 days. For A and B, calcium content was determined. For C, 45Ca uptake was determined as described under Materials and Methods. Mean ± S.D. was calculated for each group (n = 6). For A and C, one-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. ***, p < 0.001 versus 0.9 mM phosphorus. For B, statistical comparisons were performed by unpaired t test. ***, p < 0.001 versus 0.9 mM phosphorus on corresponding days.
Effect of phosphorus and vitamin D analogs on calcium content and 45Ca uptake in cultured human coronary artery smooth muscle cells. A, cells were cultured in medium containing phosphorus ranging from 0.9 to 2.64 mM plus 3 U/ml alkaline phosphatase with control, 100 nM 1α,25(OH)2D3, 19-nor, or 1α,25(OH)2D2 for 6 days. Calcium content was determined as described under Materials and Methods. B, cells were cultured in medium containing 0.9 mM phosphorus with or without different concentrations of 1α,25(OH)2D3, 19-nor, or 1α,25(OH)2D2 for 6 days. C, cells were cultured in medium containing 2.06 mM phosphorus plus 3 U/ml alkaline phosphatase with or without different concentrations of 1α,25(OH)2D3, 19-nor, or 1α,25(OH)2D2 for 6 days. In B and C, uptake of 45Ca was determined as described under Materials and Methods. Mean ± S.D. was calculated for each group (n = 4). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. None of the values was statistically significant versus control (no drug).
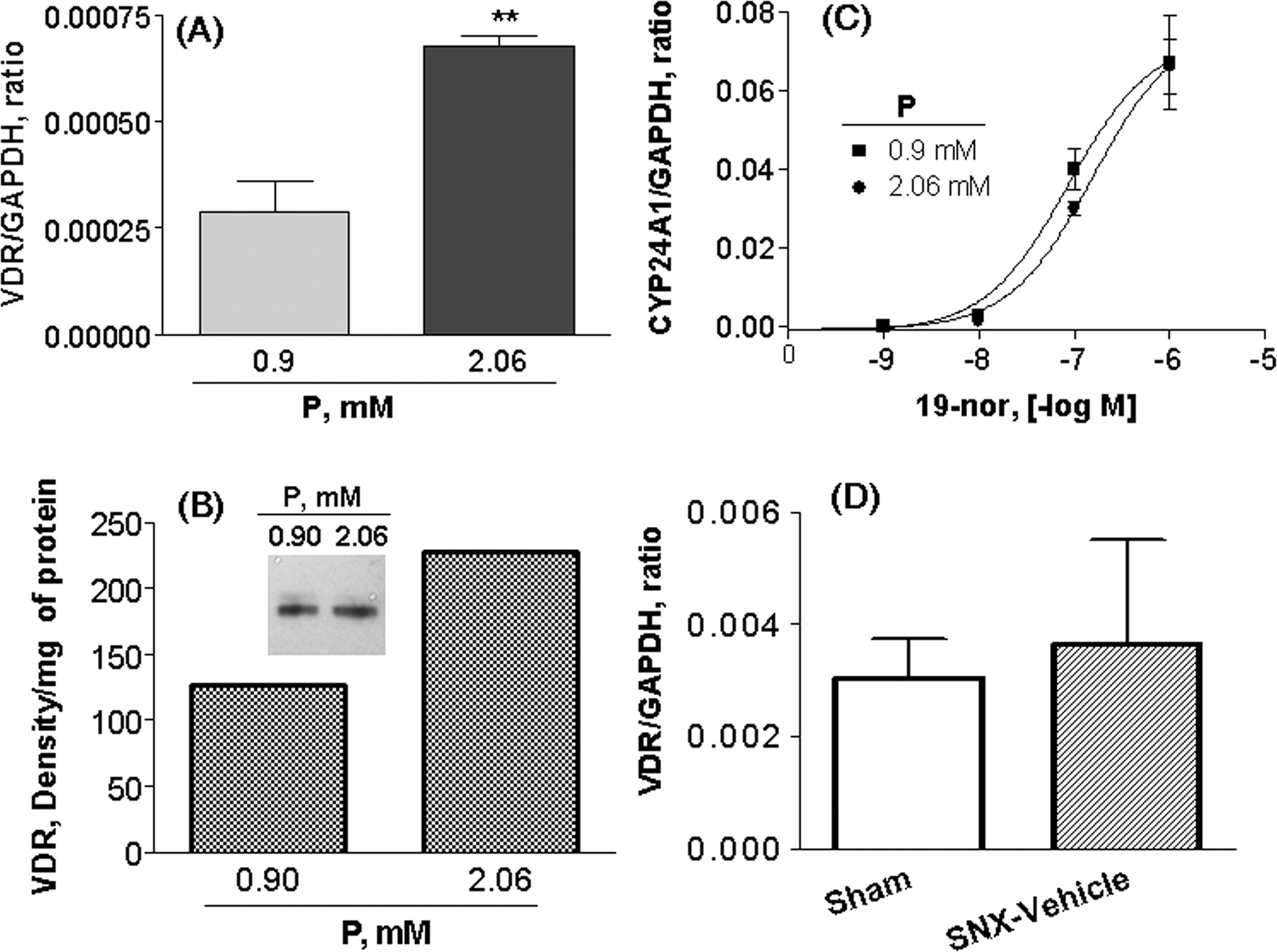
To investigate whether the lack of effect of vitamin D analogs on calcification in vitro was due to the absence of VDR in smooth muscle cells and in aorta tissues, VDR expression was examined. In Fig. 9A, the expression of VDR mRNA was detected in SMCs cultured at 0.9 or 2.06 mM phosphorus; the expression of VDR mRNA was higher at 2.06 mM phosphorus. Consistently, Fig. 9B shows that, analyzed by Western blotting, the expression of VDR at 2.06 mM phosphorus was approximately 1.8-fold of that at 0.9 mM phosphorus. As expected, 19-nor profoundly stimulated the expression of CYP24A1 mRNA in a dose-dependent manner in SMCs at 0.9 or 2.06 mM phosphorus with EC50 at 87 and 157 nM, respectively (Fig. 9C). Figure 9D shows that VDR mRNA was detected in aorta prepared from sham or SNX-vehicle rats. These results confirm that VDR was expressed under either the normal or high phosphorus condition, suggesting that the lack of effects of vitamin D analogs on calcification in vitro is not due to the absence of VDR.
One key difference between the in vivo and in vitro models is the presence or absence of the endothelial cells. To further investigate whether endothelial cells play a role, we tested phosphorus and vitamin D analogs in the coculture of human coronary artery smooth muscle and endothelial cells. Figure 10A shows that no significant 45Ca uptake was observed in endothelial cells. As a comparison, Fig. 10B shows that increasing the medium phosphorus concentration from 0.9 to 2.06 mM induced a 15-fold increase in 45Ca uptake into smooth muscle cells. However, 1α,25(OH)2D2 or 19-nor at 100 nM exhibited no significant effect compared with control (no drug). These results suggest that the contradictory effects of phosphorus and vitamin D analogs on calcification in vivo versus in vitro are not likely due to the presence or absence of endothelial cells.

VDR expression and effect of 19-nor on CYP24A1 mRNA. A to C, SMCs were cultured in medium containing 0.9 or 2.06 mM phosphorus plus 3 U/ml alkaline phosphatase for 5 days. Real-time RT-PCR and Western blotting were performed as described under Materials and Methods. A, the expression level of VDR mRNA was normalized to the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA level. Values shown are means ± S.D. (n = 4). Statistical comparisons were performed by unpaired t test. **, p < 0.01. B, The band intensity from Western blotting was normalized to the protein level in each sample. Results shown are representative of two independent experiments. C, SMCs were treated with increasing concentrations of 19-nor at 0.9 or 2.06 mM phosphorus, and RNA was isolated for real-time RT-PCR. The expression level of CYP24A1 mRNA was normalized to the GAPDH mRNA level. Values shown are means ± S.D. (n = 4). D, aorta tissues were isolated from sham or SNX (uremic) rats, and RNA was isolated for real-time RT-PCR. Values shown are means ± S.D. (n = 6 per group).
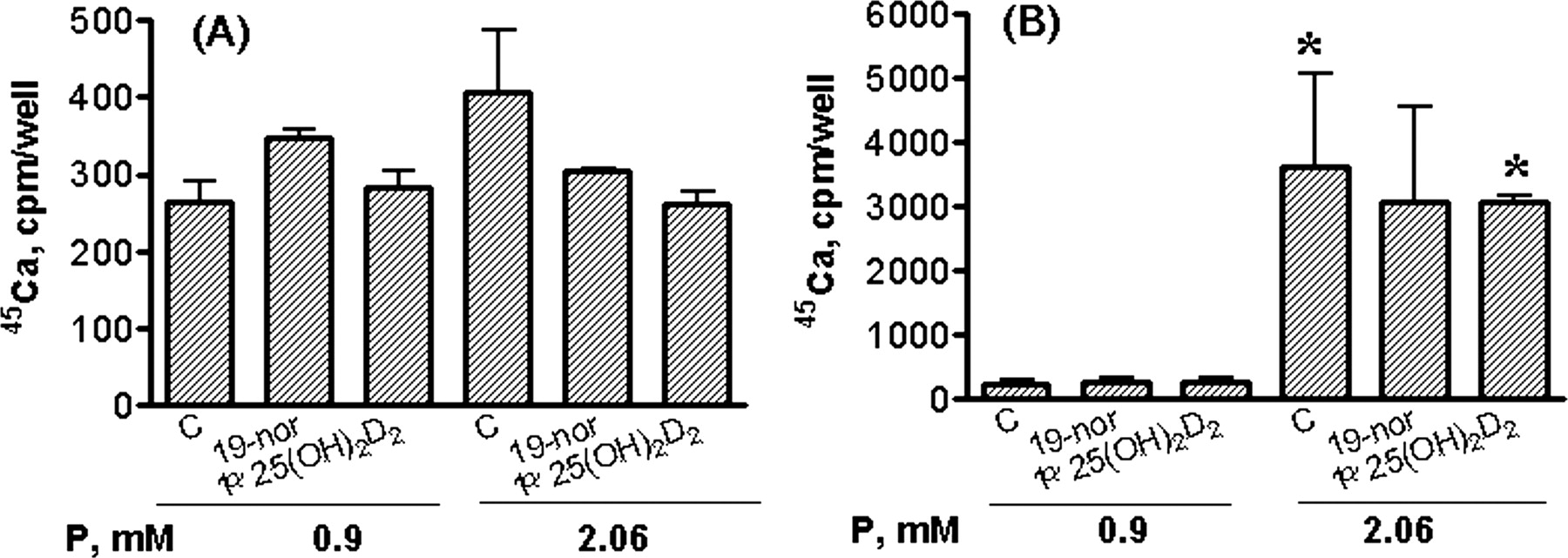
We then determined 45Ca uptake into smooth muscle cells treated with serum prepared from hyperphosphatemic uremic rats. Figure 11 shows that serum from 1α(OH)D2-treated uremic rats induced a significant increase in 45Ca uptake into smooth muscle cells cultured at 2.06 mM phosphorus [2.7- and 3.0-fold versus sham for 0.17 and 0.33 μg/kg 1α(OH)D2, respectively]. As a comparison, serum prepared from uremic rats treated with 19-nor resulted in a 40% decrease in 45Ca uptake into smooth muscle cells. Interestingly, none of the serum samples exhibited any significant effect on 45Ca uptake when cells were cultured at 0.9 mM phosphorus. In addition, 2.06 mM phosphorus in this study only induced calcium uptake by ∼5-fold (versus. 0.9 mM phosphorus). These results suggest that 1) the uremic milieu from the 1α(OH)D2-treated animals induced an increase in calcium uptake, and 2) the effect was only evident under a high-phosphorus environment.
Discussion
Vascular calcification has become an important area of research during the past decade largely because it is often associated with cardiovascular morbidity and mortality, especially for CKD patients (Blacher et al., 2001; London et al., 2003; Moe et al., 2004). However, so far the available data regarding how factors such as vitamin D analogs and phosphorus are involved in the calcification process remain controversial. Moreover, the perception that vitamin D analog usage may be associated with a higher risk for vascular calcification contradicts recent retrospective clinical observations that vitamin D analogs provide a survival benefit for dialysis patients. The goal of the present study was to investigate the interactive effects of vitamin D analogs and phosphorus on calcification. Although contradictory results were obtained from in vivo and in vitro models, it is perhaps not surprising because vascular calcification is a complex process involving a delicate balance between various inhibitory and inducing factors.
First, we found that an increase in phosphorus and CaxP product in vivo does not necessarily result in aortic calcification, but high phosphorus consistently induces calcification in vitro. This observation suggests that high phosphorus may be an inducing factor for calcification, but a counter-regulatory mechanism exists in vivo to balance out the calcification-promoting effects of hyperphosphatemia. Previously, it had been suggested that some factors in serum may act as potent inhibitors of smooth muscle cell calcification (Shanahan, 2005). Thus, one possible explanation for the aortic calcification observed in 1α(OH)D2-treated hyperphosphatemic uremic rats is that the endogenous protective mechanism crucial for suppression of high phosphorus-induced vascular calcification is disturbed. Exactly what mechanisms are altered by 1α(OH)D2 treatment will require more studies.
Effect of vitamin D analogs and phosphorus on 45Ca uptake to cells in the coculture of human coronary artery endothelial and smooth muscle cells. Endothelial (A) and smooth muscle (B) cells were initially cultured separately with endothelial cells in inserts. Cells were then put together in medium containing phosphorus at 0.9 or 2.06 mM plus 3 U/ml alkaline phosphatase with control, 19-nor, or 1α,25(OH)2D2 (100 nM) for 6 days. Uptake of 45Ca was determined as described under Materials and Methods. Mean ± S.D. was calculated for each group (n = 3). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. *, p < 0.05 versus control (no drug) at 0.9 mM phosphorus.

Effect of serum from uremic rats on 45Ca uptake into human coronary artery smooth muscle cells. Cells in 0.5 ml of medium containing phosphorus at 0.9 or 2.06 mM plus 3 U/ml alkaline phosphatase were treated with 10% serum from sham (n = 5) or hyperphosphatemic uremic rats that were dosed with vehicle (SNX-vehicle, n = 3), 19-nor at 0.33 μg/kg (n = 3), or 1α(OH)D2 at 0.17 (n = 4) or 0.33 μg/kg (n = 3) i.p. three times per week, for 40 days. Serum from each treatment group was pooled. Uptake of 45Ca was determined as described under Materials and Methods. Mean ± S.D. was calculated for each group (n = 3). One-way ANOVA Dunnett's test with 95% confidence intervals of difference was performed for statistical comparisons. **, p < 0.01 versus sham at 2.06 mM phosphorus.
Second, no aortic calcification was observed in uremic rats consuming a normal diet and treated with 1α(OH)D2. In addition, serum from uremic rats treated with 1α(OH)D2 induced calcium uptake into cultured smooth muscle cells only at 2.06 mM but not at 0.9 mM phosphorus. Taken together, these observations suggest that 1) vascular calcification may not occur in uremic rats in the absence of permissive conditions such as hyperphosphatemia, and 2) some vitamin D analogs such as 1α(OH)D2 interact with high phosphorus to undermine the endogenous protective mechanism against vascular calcification.
Third, results from von Kossa staining of aorta sections prepared from the 1α(OH)D2-treated uremic rats suggest that calcification occurred in the aortic medial layer (Fig. 4). Consistent with this observation, denuded aortic rings prepared from 1α(OH)D2-treated uremic rats exhibited a significant increase in 45Ca uptake ex vivo. The results suggest that calcification under uremic conditions predominantly occurs in smooth muscle cells. In addition, endothelial cells do not seem to be involved. However, these data do not explain why 1α,25(OH)2D2 and 1α,25(OH)2D3 failed to induce calcification in smooth muscle cells in vitro. Viewed collectively, these observations demonstrate that 1) the complex regulation of vascular calcification in vivo cannot be easily replicated in the ex vivo or in vitro models, and 2) it is important to use a relevant disease state model such as the hyperphosphatemic uremic rat for vascular calcification studies.
Finally, we think it is necessary to compare and discuss findings from the present study versus reports in the literature. Previously, Hirata et al. (2003) showed that 1α,25(OH)2D3 induced vascular calcification in five-sixths nephrectomized uremic rats, whereas 1,25-dihydroxy-22-oxacalcitriol, an analog of 1α,25(OH)2D3, did not show any effect. They also showed that 1,25-dihydroxy-22-oxacalcitriol at a high dose (6.25 μg/kg) raised serum calcium, phosphorus, and CaxP levels to the same level as that for 1α,25(OH)2D3 at 0.125 μg/kg, but only 1α,25(OH)2D3 induced aortic calcification. Our observations confirm that different vitamin D analogs exert differential effects on vascular calcification independent of the serum calcium, phosphorus, and CaxP levels. To the best of our knowledge, the differential effect of 1α(OH)D2 and 19-nor on vascular calcification has not been reported previously. Although our observation that high phosphorus induces an increase in calcium content and 45Ca uptake into smooth muscle cells is consistent with previous reports by others (Wada et al., 1999; Steitz et al., 2001), the lack of effect of vitamin D analogs on inducing calcium uptake into cultured smooth muscle cells is in conflict with that reported by Jono et al. (1998). It is worth mentioning that, although Jono et al. (1998) showed calcification induced by calcitriol in cultured bovine aortic smooth muscle cells, a recent report by Wolisi and Moe (2005) indicated that neither 1α,25(OH)2D3 nor 19-nor showed any effect on calcification in bovine vascular smooth muscle cells. At present, these disparate results from studying cultured vascular cells cannot be readily reconciled, although it is possible that different preparations of smooth muscle cells may differ in their functional response to vitamin D analogs.
In summary, we demonstrate that high phosphorus stimulates 45Ca uptake and induces an accumulation of calcium in smooth muscle cells in vitro and ex vivo, but does not induce aortic calcification in vivo. In contrast, vitamin D analogs exhibit no effects on calcification in vitro and ex vivo, but display differential effects on aortic calcification in vivo. Furthermore, serum from uremic rats treated with 1α(OH)D2-induced calcium uptake into cultured smooth muscle cells only under high phosphorus conditions. These results suggest that the complex regulation of vascular calcification in vivo cannot be easily replicated in ex vivo or in vitro models, and some vitamin D analogs such as 1α(OH)D2 may induce aortic calcification in uremic rats when a permissive condition such as hyperphosphatemia is present.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.106.101261.
-
ABBREVIATIONS: CDK, chronic kidney disease; CV, cardiovascular; VDR, vitamin D receptor; PTH, parathyroid hormone; 1α,25(OH)2D3, 1,25-dihydroxyvitamin D3; 1α,25(OH)2D2, 1α,25-dihydroxyvitamin D2; 1α(OH)D2, 1α-hydroxyvitamin D2; 19-nor, 19-nor-1α,25-dihydroxyvitamin D2; SNX, five-sixths nephrectomized; BUN, blood urea nitrogen; DMEM, Dulbecco's modified Eagle's medium; PBS, phosphate-buffered saline; RT, reverse transcription; PCR, polymerase chain reaction; PBS-T, phosphate-buffered saline-Tween 80; ANOVA, analysis of variance; CaxP, calcium-phosphorus product; SMC, human coronary artery smooth muscle cell.
- Received January 11, 2006.
- Accepted April 6, 2006.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}