


Abstract
The antidiabetic agent glyburide (glibenclamide) is frequently used for the treatment of type II diabetes and is increasingly being used for the treatment of gestational diabetes. Evidence suggests that breast cancer resistance protein/ATP-binding cassette, subfamily G, member 2 (ABCG2) expressed in the placenta protects the fetus against the accumulation of glyburide. A number of studies have investigated the significance of several single-nucleotide polymorphisms (SNPs) in the ABCG2 gene. Associations between the Q141K (C421A) SNP and ABCG2 protein expression, membrane surface translocation, efflux activity, or ATPase activity have been shown. Therefore, alterations in glyburide transport across the placenta, resulting in increased fetal glyburide exposure, may be seen in individuals carrying the C421A allele. The purpose of this study is to investigate whether the Q141K SNP causes alterations in ABCG2-mediated glyburide transport. Glyburide accumulation assays were carried out with stably transfected human embryonic kidney (HEK)-293 cells expressing wild-type ABCG2 (Arg482) and polymorphic ABCG2 (Q141K). Glyburide kinetic parameters were determined for comparison of wild-type and SNP ABCG2 activity by simultaneously fitting data for ABCG2-expressing cells (saturable transport) and empty vector-expressing cells (nonsaturable transport) by nonlinear regression analysis. The apparent Kt and Vmax values for the transfected HEK-293 cells expressing the polymorphic variant (Q141K) of ABCG2 were significantly higher than those values determined for the wild-type ABCG2-expressing cells (p < 0.05). Our results indicate that the Q141K variant of ABCG2 may have the potential to alter the placental pharmacokinetics of glyburide used in pregnancy.
Glyburide (glibenclamide) is a second-generation sulfonylurea hypoglycemic agent that is frequently used for the treatment of type 2 diabetes and is increasingly being used for the treatment of gestational diabetes. Previously, oral hypoglycemic agents were avoided in pregnancy because of fears of exposure to the fetus. However, Elliott et al. (1991) showed limited placental transfer of glyburide across the perfused placental lobule. Glyburide's limited transfer across the placenta, despite its small molecular weight and lipid solubility, was thought to be the result of its high protein binding (99.8%) and short elimination half-life. However, it was discovered, through perfusion studies using human placenta, that glyburide is actively effluxed from the fetal compartment to the maternal compartment (Kraemer et al., 2006). Subsequently, glyburide uptake studies carried out using breast cancer resistance protein (BCRP)/ATP-binding cassette, subfamily G, member 2 (ABCG2)- and multidrug resistance protein (MRP1, MRP2, MRP3)-overexpressing cell lines and human placental vesicle preparations, in the presence and absence of inhibitors, have suggested that glyburide is preferentially transported by ABCG2 (Gedeon et al., 2006, 2008). In addition, results from studies examining fetal distribution of glyburide in wild-type and Bcrp1(−/−) pregnant mice showed mean fetal glyburide concentrations in the Bcrp1(−/−) pregnant mice were two times greater than that in the wild-type pregnant mice (Zhou et al., 2008). More recently, these results were confirmed in our laboratory through human placental perfusion experiments completed with glyburide in the presence of nicardipine, an ABCG2 inhibitor, which suggests that BCRP/ABCG2 is likely responsible for the efflux of glyburide across the human placenta (Pollex et al., 2008).
ABCG2, or BCRP, is a 72-kDa plasma membrane protein belonging to the ATP-binding cassette family of transport proteins. ABCG2 is known to play a role in actively effluxing a wide spectrum of exogenous and endogenous substrates across cellular membranes. The high endogenous expression of ABCG2 in the syncytiotrophoblast of the human placenta (Maliepaard et al., 2001) has led to the postulation of a role in protecting the fetus against the accumulation of xenobiotics. Because of the potential impact of ABCG2 on drug disposition, a number of studies have investigated the significance of several single-nucleotide polymorphisms (SNPs) in the ABCG2 gene. In particular, the nonsynonymous substitution of glutamine for lysine at mRNA position 421 (C421A) has been shown in several studies to have functional significance (Imai et al., 2002; Kondo et al., 2004; Mizuarai et al., 2004; Morisaki et al., 2005; Sparreboom et al., 2005). The results of these studies vary, showing inconsistencies in the effect of the C421A (Q141K) polymorphism on ABCG2 protein activity. It has been suggested that the Q141K SNP may affect ABCG2 protein expression, membrane surface translocation, efflux activity, or ATPase activity. Of particular concern, in vivo studies have shown patients expressing the Q141K variant who were exposed to the chemotherapeutic agents topotecan and diflomotecan achieved higher plasma drug levels, suggesting a significant reduction in ABCG2-mediated excretion (Sparreboom et al., 2004). However, altered pharmacokinetic parameters of irinotecan, a known ABCG2 substrate, were not shown in a study with patients carrying the C421A allele (de Jong et al., 2004).
Studies have shown considerable differences in genotype frequency, ranging from 0 to 35%, among different ethnic populations. The Han Chinese population appears to exhibit the highest frequency (34%), whereas the variant allele is very rare in sub-Sahara African populations (<1%) and occurs at a relatively low frequency in black and white populations in the United States (5 and 12%, respectively) (Imai et al., 2002; de Jong et al., 2004).
Because BCRP/ABCG2 is believed to protect the fetus against the accumulation of its substrates, it is important to determine the potential effect of the Q141K SNP on the placental transfer of drugs used in pregnancy. In particular, certain ethnic populations may be at an increased risk for fetal exposure to ABCG2 substrate drugs taken during pregnancy. Numerous studies have shown that glyburide is both safe and effective for use in pregnancy; however, its affinity for the ABCG2 protein may lead to increased fetal exposure in individuals carrying the C421A allele. Therefore, the purpose of this study is to investigate whether the Q141K SNP causes alterations in ABCG2-mediated glyburide transport.
Materials and Methods
Stably transfected human embryonic kidney (HEK)-293 cells containing full-length ABCG2 encoding wild-type (Arg482) or the SNP variant (Q141K) of ABCG2, or an empty vector to serve as a control were obtained (Dr. Robert W. Robey, National Cancer Institute, Bethesda, MD), and expression was enforced by selection in G418 (Invitrogen, Carlsbad, CA). Stable transfectants were maintained in Eagle's minimum essential medium (American Type Culture Collection, Manassas, VA) supplemented with 10% fetal bovine serum, 1% penicillin (20 U/ml), streptomycin (20 μg/ml), and G418 (2 mg/ml) as described by Morisaki et al. (2005).
Flow Cytometry.
Flow cytometry assays were performed to confirm surface expression of ABCG2 in the wild-type (Arg482) and polymorphic (Q141K) clones. Absence of surface expression of ABCG2 in the empty vector-transfected HEK-293 cells was also confirmed by flow cytometry. In brief, cells were trypsinized and incubated overnight in complete medium at 37°C in an atmosphere containing 5% CO2 on a rocker platform to enable regeneration of the BCRP protein. Cells were centrifuged at 500g for 5 min and washed three times in an isotonic phosphate-buffered saline (PBS) buffer supplemented with 0.5% bovine serum albumin (BSA). Cells were then resuspended in the same buffer. Cells were initially Fc-blocked for nonspecific binding by treatment with 1 μg of human IgG/105 cells for 15 min at room temperature (Sigma-Aldrich, Oakville, ON, Canada). The cells were then incubated with monoclonal phycoerythrin-conjugated anti-human ABCG2 antibody (R&D Systems/Cedarlane Laboratories, Burlington, ON, Canada) or phycoerythrin-conjugated mouse IgG2B negative control antibody (R&D Systems/Cedarlane Laboratories) for 30 min at 2 to 8°C. Cells were washed with PBS buffer supplemented with 0.5% BSA and resuspended for final flow cytometric analysis using a FACScan sorter with 488-nm wavelength laser excitation (BD Biosciences, Mississauga, ON, Canada). Surface expression of ABCG2 was confirmed on three independent occasions before carrying out each transport experiment.
Cellular Transport Studies.
Transport activity was examined by measuring glyburide accumulation in stably transfected HEK-293 cells expressing wild-type ABCG2 (Arg482), polymorphic ABCG2 (Q141K), and an empty vector. In brief, HEK-293 cells were grown to 90% confluence in 12-well plates. Cells were incubated at 37°C in Eagle's minimum essential medium containing various concentrations of [3H]glyburide (0.2–100 μM; PerkinElmer Life and Analytical Sciences, Waltham, MA) for 20 min. After 20 min of incubation, cells were washed with ice-cold PBS buffer and allowed to solubilize in NaOH (1 M). Subsequently, the cells were neutralized with HCl (1 M). The intracellular concentration of [3H]glyburide was determined by adding 4 ml of scintillation fluid to vials containing 600 μl of solubilized HEK-293 cells and by determining radioactivity using a Beckman Coulter LS 6500 liquid scintillation counter (Beckman Coulter Canada, Inc., Mississauga, ON, Canada). Glyburide concentrations were extrapolated from standard curves generated from glyburide-spiked media. Glyburide accumulation was normalized to the protein concentration of lysates from each well using the Bradford assay method of protein determination and BSA standards (Bradford, 1976). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT) assays were used to examine potential glyburide toxicity under our conditions. For MTT assays, cells were prepared and treated with glyburide as they were for accumulation experiments but in 96-well plates. After treatment, cells were incubated with MTT (Sigma-Aldrich) in seeding/incubation medium. Cells were then solubilized, and the extent of MTT reduction to formazan was quantified by spectrophotometry (570 nm).
Data Analysis.
Results are reported as a mean ± S.E.M. from three independent experiments each performed in triplicate. Nonspecific binding was corrected for by subtracting [3H]glyburide accumulation determined at time 0 from all the values obtained at time 20 min. Data were visualized before fitting to confirm the appropriateness of the chosen model. Kt, Kd, and Vmax values were obtained from Scientist 3.0 software (St. Louis, MO) through simultaneously fitting data for the ABCG2-expressing cells (saturable transport) and the empty vector cells (nonsaturable transport) by nonlinear regression analysis using eqs. 1 and 2 (González-Alvarez et al., 2005):

 where S and V represent the concentration of substrate and transport rate, respectively. Vmax, Kt, and Kd represent the maximal transport rate for saturable transport, the affinity for saturable transport, and the rate constant for nonsaturable transport, respectively. Data obtained from experiments carried out on empty vector-transfected HEK-293 cells were analyzed using eq. 1 to determine the contribution of nonsaturable transport to glyburide accumulation. The goodness of fit was assessed from the experimental versus predicted values graphs, and the residuals were plotted to confirm random variation. Student's t test was used to determine the statistical significance of differences.
where S and V represent the concentration of substrate and transport rate, respectively. Vmax, Kt, and Kd represent the maximal transport rate for saturable transport, the affinity for saturable transport, and the rate constant for nonsaturable transport, respectively. Data obtained from experiments carried out on empty vector-transfected HEK-293 cells were analyzed using eq. 1 to determine the contribution of nonsaturable transport to glyburide accumulation. The goodness of fit was assessed from the experimental versus predicted values graphs, and the residuals were plotted to confirm random variation. Student's t test was used to determine the statistical significance of differences.
Assumptions of the Kinetic Model.
In using the current method to evaluate the drug efflux kinetics, several assumptions were made. Data from the HEK-293 empty vector-transfected cell line and each of the ABCG2-transfected HEK-293 cell lines (Arg482, Q141K) were used simultaneously to fit the model parameters because the assumption was made that the permeability of the empty vector-transfected cell line and each of the ABCG2-transfected HEK-293 cell lines was the same. In the present study, a HEK-293 cell line transfected with an empty vector was used to determine and correct for the degree to which glyburide moves by diffusion. The use of an inhibitor for ABCG2 would have required making the assumption that complete ABCG2 inhibition was achieved and that no differences in the ability of the inhibitor to inhibit the wild-type and polymorphic ABCG2 variant exist. The use of an empty vector-transfected HEK-293 cell line removes the need to make these assumptions and subsequently avoids the potential to underestimate the transport capacity of ABCG2. Finally, ABCG2 kinetics were determined after measurement of glyburide accumulation in the cell and were based on the extracellular concentration of glyburide exposed to the cells as described previously (Bachmeier et al., 2006). Therefore, the results represent apparent Kt values for glyburide transport.
Results
ABCG2 Surface Expression in HEK-293-Transfected Cells.
The surface expression of ABCG2 in HEK-293 cells transfected with empty vector, wild-type (Arg482), or SNP variant (Q141K) of ABCG2 was examined by flow cytometry. As shown in Fig. 1, surface expression of ABCG2 was detected at similar levels in the ABCG2-transfected HEK-293 cell lines expressing wild-type (Arg482) and the polymorphic variant (Q141K) of ABCG2 (Fig. 1). ABCG2 surface expression was not detectable in the empty vector-transfected HEK-293 cell line.

Surface expression of ABCG2 in transfected HEK-293 cells. Cells were incubated for 30 min with either phycoerythrin-labeled negative control antibody or the phycoerythrin-labeled anti-ABCG2 antibody 5D3 (red, negative control; green, Arg482; blue, Q141K).
[3H]Glyburide Transport.
After initial time course experiments, glyburide accumulation was found to be linear, reaching a plateau by 30 min in transfected HEK-293 cells. Inhibition of ABCG2/BCRP by fumitremorgin C (FTC) (10 μM) resulted in marked increases in intracellular accumulation of [3H]glyburide in HEK-293 cells expressing the wild-type (Arg482) ABCG2 or SNP variant (Q141K) of ABCG2 after 60 min of exposure to 20 μM glyburide (Fig. 2A). Similar findings were seen after 20 min of exposure to 20 μM glyburide. [3H]Glyburide accumulation experiments carried out for 20 min at various concentrations of glyburide (0.2–100 μM) show inhibitable transport in both Arg482- and Q141K-transfected HEK-293 cells exposed to up to 20 μM glyburide (Fig. 2B). At concentrations of 100 μM glyburide, the accumulation of glyburide in wild-type (Arg482) transfected cells was similar in the presence or absence of FTC and was similar to accumulation in cells transfected with empty vector, suggesting saturation of active transport (data not shown). Data obtained suggest that full FTC-mediated inhibition of BCRP/ABCG2 is seen in the wild-type transfected cells at higher drug concentrations than in the SNP-transfected cells (Q141K) (Fig. 2B).

A, glyburide accumulation in the presence and absence of FTC (10 μM) in HEK-293 cells transfected with wild-type ABCG2 (Arg482), ABCG2 polymorphic variant (Q141K), and control (empty vector) after 60 min of exposure to 20 μM glyburide. Data represent mean ± S.E.M. from an experiment performed in triplicate (n = 3); *, p < 0.05, **, p < 0.001. B, accumulation of glyburide in HEK-293 cells transfected with wild-type ABCG2 (Arg482), ABCG2 polymorphic variant (Q141K), and empty vector control after 20 min of exposure to various concentrations of glyburide in the presence and absence of FTC (10 μM). Data are mean ± S.E.M. from an experiment performed in triplicate (n = 3).
ABCG2 Kinetic Studies.
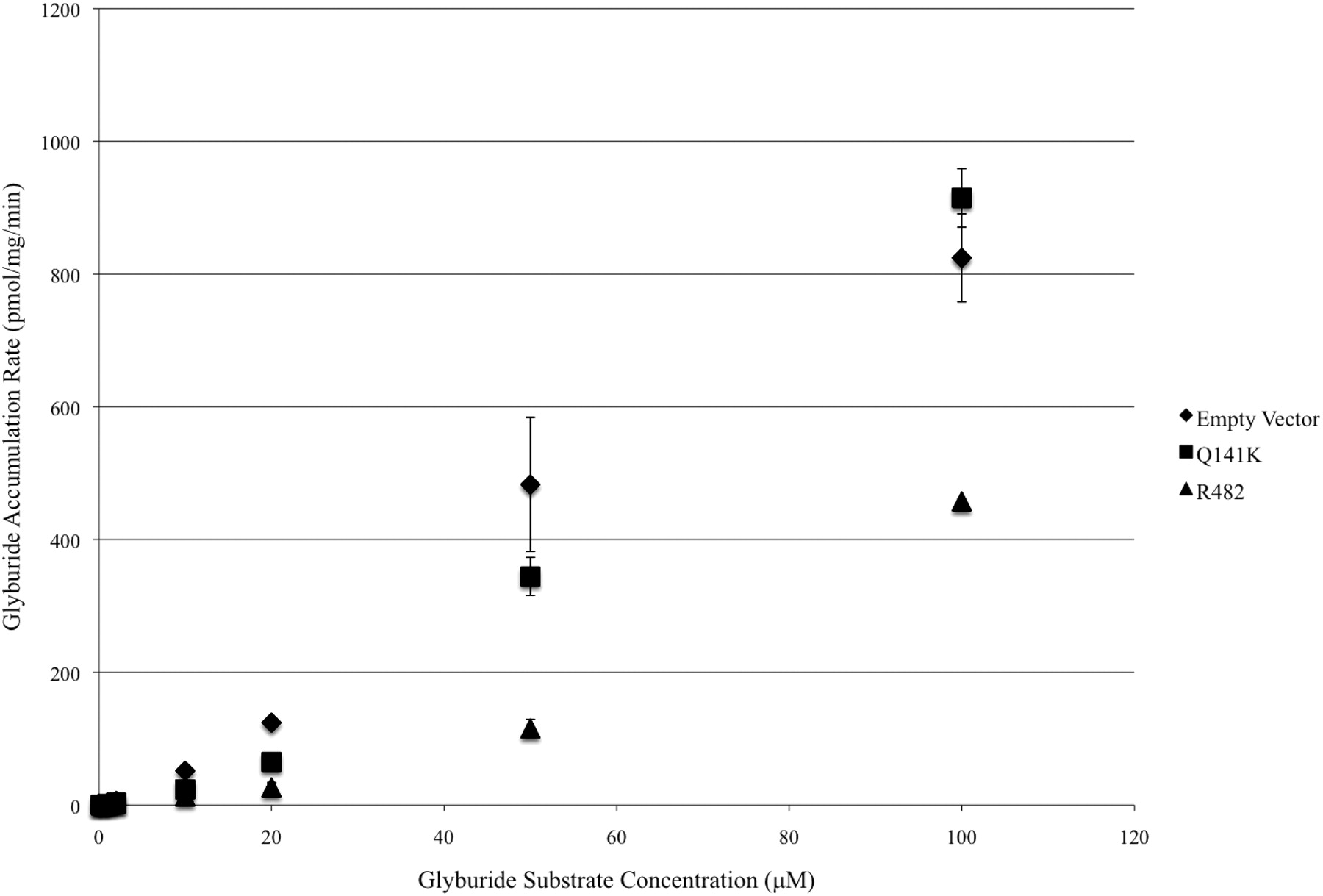
After transport studies carried out at increasing concentrations (0.2–100 μM) of [3H]glyburide in ABCG2-transfected HEK-293 cells expressing wild-type (Arg482) ABCG2 (Fig. 3), the apparent Kt and Vmax values were 13.07 ± 1.21 μM and 101.23 ± 34.46 pmol/min · mg protein, respectively. In the transfected HEK-293 cells expressing the polymorphic variant (Q141K) of ABCG2, the apparent Kt and Vmax values were 80.37 ± 22.96 μM and 384.79 ± 69.71 pmol/min · mg protein, respectively. Both the apparent Kt and Vmax values for the transfected HEK-293 cells expressing the polymorphic variant (Q141K) of ABCG2 were found to be significantly higher than those values determined for the wild-type ABCG2-expressing cells (p < 0.05).

Rate of glyburide transport in wild-type (Arg482), SNP (Q141K), and empty vector-transfected HEK-293 cells after exposure to various concentrations of glyburide (0.2–100 μM) for 20 min. Data represent the mean ± S.E.M. from a single experiment performed in triplicate (n = 3).
Discussion
Reports reveal that glyburide is often used as first-line therapy for gestational diabetes treatment (Gabbe et al., 2004). In general, the use of glyburide for the treatment of diabetes in pregnancy has been increasing because of the large number of reports of use in pregnancy failing to show increased risk for negative fetal outcomes (Feig et al., 2007). Much of the reassurance on the safety of the use of glyburide in pregnancy arose from several studies showing not only limited transfer across the placenta compared with older sulfonylurea hypoglycemic agents but also the existence of a protective efflux mechanism mediated by BCRP (Elliott et al., 1994; Kraemer et al., 2006; Gedeon et al., 2008; Pollex et al., 2008).
The recent identification of the nonsynonymous SNP, Q141K (Honjo et al., 2002; Imai et al., 2002), in the coding region of ABCG2 and the potential functional consequences on drug disposition has led to an increasingly important area of research. To elucidate the potential effects of the Q141K SNP on BCRP-mediated glyburide transport, we examined and compared the accumulation of glyburide in HEK-293 cells stably transfected with full-length ABCG2 encoding wild-type or the SNP variant (Q141K) of ABCG2. Transport studies showed BCRP-mediated glyburide transport in both wild-type and SNP ABCG2-transfected cells. Treatment of the cells with 10 μM FTC, a potent inhibitor of BCRP, resulted in significant increases in the intracellular accumulation of [3H]glyburide (Fig. 3), confirming BCRP-mediated transport.
Regarding BCRP transport kinetics, the values obtained for apparent affinity (Kt) of glyburide for BCRP suggest that glyburide has greater affinity for the wild-type ABCG2 protein compared with the affinity determined for the Q141K ABCG2 variant. Because glyburide accumulation in the HEK-293 cells was determined, BCRP kinetics were indirectly assessed, as described previously, based on the knowledge that accumulation is a function of both uptake/diffusion and efflux processes (Bachmeier et al., 2006). The apparent Kt values determined following this method of analysis likely overestimate the actual Kt values because these estimates were based on extracellular glyburide concentrations and do not necessarily reflect the amount of glyburide available for BCRP-mediated efflux. However, because the transfected cell lines are not expected to have differences in the expression and activity of uptake processes, it is unlikely that the cell lines differ in terms of their uptake capacity. The primary difference between the wild-type and SNP variant-transfected cells stems from BCRP protein activity. Therefore, differences in glyburide transport parameters, as shown by these results, are likely reflective of altered BCRP-mediated efflux activity. Metabolism of glyburide in this system is also thought to be minimal and similar in both wild-type and Q141K ABCG2-transfected cells; however, intracellular binding and glyburide metabolism could also confound the estimation of Kt values.
It has been previously reported that the Q141K variant is associated with increased sensitivity to chemotherapeutic agents as a result of a decrease in protein expression in transfected cells (Imai et al., 2002). Data obtained in our study did not show reduced surface expression of Q141K ABCG2. Thus, the impaired ability of the ABCG2 protein to efflux glyburide seen in cells transfected with Q141K ABCG2 was likely the result of SNP-induced alterations in the function of the protein itself. Similar to our findings, Morisaki et al. (2005) and Zamber et al. (2003) did not observe a correlation between decreased expression of ABCG2 and the Q141K SNP. However, Morisaki et al. (2005) noted that a larger proportion of the surface-localized Q141K ABCG2 protein is intracellular, implying less efficient post-translational processing in the SNP variant and thus providing a potential mechanism for reduced efficiency.
In summary, data from our accumulation studies indicate that the HEK-293 cell line expressing the Q141K polymorphic variant of the ABCG2/BCRP gene exhibits a reduced affinity for BCRP-mediated glyburide efflux. Therefore, the Q141K variant of ABCG2 may have the potential to alter the placental pharmacokinetics of glyburide and other clinically used BCRP substrate drugs in pregnancy. Further studies using a BCRP efflux model are required to directly determine the kinetic parameters for BCRP-mediated glyburide transport and the potential clinical relevance. However, the results of this study provide sufficient reason to further investigate the impact of SNPs on ABCG2-mediated glyburide transport because the potential exists for increased fetal exposure to glyburide in pregnancy among patients carrying the Q141K polymorphic variant allele of ABCG2. The results of this study also provide an important potential source of variability in fetal exposure and outcome after maternal exposure to glyburide in pregnancy. As we become progressively more aware of the transporters that influence the transplacental transport of drugs used in pregnancy, the study of transporter polymorphisms and the potential consequences of alterations in function will become increasingly important.
Acknowledgments.
We thank Dr. Robert Robey and colleagues at the National Cancer Institute, National Institutes of Health (Bethesda, MD) for providing HEK-293 stably transfected cells and the BCRP inhibitor, FTC.
Footnotes
This work was supported by the Canadian Institutes of Health Research [Grant 1797].
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
doi:10.1124/dmd.109.030791.
-
ABBREVIATIONS:
- BCRP
- breast cancer resistance protein
- ABCG2
- ATP-binding cassette, subfamily G, member 2
- MRP
- multidrug resistance protein
- SNP
- single-nucleotide polymorphism
- HEK
- human embryonic kidney
- PBS
- phosphate-buffered saline
- BSA
- bovine serum albumin
- MTT
- 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
- FTC
- fumitremorgin C.
- Received October 14, 2009.
- Accepted February 16, 2010.
- Copyright © 2010 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}