


Abstract
Flavonoids are a class of polyphenolic compounds widely present in the diet and herbal products. The interactions of flavonoids with some major efflux transporters [e.g., P-glycoprotein, multidrug resistance-associated protein 1 (MRP1), and breast cancer resistance protein] have been reported; however, their interactions with uptake transporters are largely unknown. Organic anion-transporting polypeptide OATP1B1 is a liver-specific uptake transporter important in hepatic drug disposition. Our objective was to evaluate the effects of 20 naturally occurring flavonoids, and some of their corresponding glycosides, on the uptake of [3H]dehydroepiandrosterone sulfate (DHEAS) in OATP1B1-expressing and OATP1B1-negative HeLa cells. Many of the tested flavonoids (including biochanin A, genistein, and epigallocatechin-3-gallate) significantly inhibited [3H]DHEAS uptake in a concentration-dependent manner in OATP1B1-expressing cells, with biochanin A being one of the most potent inhibitors with an IC50 of 11.3 ± 3.22 μM. The flavonoids had negligible or small effects in OATP1B1-negative cells. Four of the eight pairs of tested flavonoids and their glycosides, namely, genistein/genistin, diosmetin/diosmin, epigallocatechin/epigallocatechin-3-gallate, and quercetin/rutin, exhibited distinct effects on [3H]DHEAS uptake. For example, genistin did not inhibit DHEAS uptake, whereas genistein did, and rutin stimulated uptake, whereas quercetin had no effect. [3H]Biochanin A uptake was similar in OATP1B1-expressing and OATP1B1-negative cells, suggesting that it is not a substrate for OATP1B1. A kinetic study revealed that biochanin A inhibited [3H]DHEAS uptake in a noncompetitive manner, with a Ki of 10.2 ± 1.89 μM. Taken together, these results indicate that flavonoids are a novel class of OATP1B1 modulators, suggesting the potential for diet-drug interactions.
Organic anion-transporting polypeptides (OATPs) are important membrane transporters expressed in various organs critical in drug disposition, such as liver, intestine, blood-brain barrier, kidney, placenta, and other organs (Tamai et al., 2000; Hagenbuch and Meier, 2004). This transporter family mediates sodium-independent uptake of a broad spectrum of amphipathic organic anions as well as steroid conjugates, organic cations, and xenobiotics (Hagenbuch and Meier, 2004; Mikkaichi et al., 2004). A number of OATPs share some overlap in substrate specificity with certain efflux transporters such as P-glycoprotein (Cvetkovic et al., 1999), multidrug resistance-associated protein 2 (MRP2) (Su et al., 2004; Spears et al., 2005), and breast cancer resistance protein (BCRP) (Nozawa et al., 2005). To date, 36 OATPs have been identified in human, rat, and mouse, and 11 of them were cloned from human tissues (Hagenbuch and Meier, 2004). Among all the human OATPs, OATP1B1 (previously known as LST-1, OATP-C, OATP-2, and SLC21A6), which has been identified by a number of groups (Abe et al., 1999; Hsiang et al., 1999; Konig et al., 2000; Tamai et al., 2000), represents one of the best characterized human OATPs. Molecular characterization revealed that OATP1B1 consists of 691 amino acids with an apparent molecular mass of 84 kDa, which is reduced to 54 kDa after deglycosylation (Konig et al., 2000). OATP1B1 is specifically expressed on the basolateral (sinusoidal) membrane of human hepatocytes and transports a broad range of compounds such as bile acids, conjugated steroids, thyroid hormones, peptides, and drugs including rifampicin and pravastatin (Abe et al., 1999; Konig et al., 2000; Hagenbuch and Meier, 2004). Due to its liver-specific expression and its capacity for transporting a large number of structurally different compounds, it has been suggested that OATP1B1 plays an important role in hepatic drug uptake and elimination (Kim, 2003). Thus, modulators of OATP1B1 may alter the pharmacokinetics of OATP1B1 substrate drugs, causing potential drug-drug interactions, exemplified by the interactions between cerivastatin and cyclosporin A (Shitara et al., 2003) and between cerivastatin and gemfibrozil (Shitara et al., 2004).
Flavonoids are a class of polyphenolic compounds widely present in fruits, vegetables, and plant-derived beverages, and in many herbal products marketed as over-the-counter dietary supplements. The average daily intake of total flavonoids from the U.S. diet was estimated to be 1 g (Kuhnau, 1976), although this may be an overestimation and actual intake may be lower (Harborne and Williams, 2000). Flavonoids have long been associated with a variety of biochemical and pharmacological properties including antioxidant, antiviral, anticarcinogenic, and anti-inflammatory activities, with no or low toxicity (Middleton et al., 2000; Havsteen, 2002). These health-promoting activities indicate that flavonoids may play a protective role in the prevention of cancer and coronary heart disease, as well as other age-related degenerative diseases (Hertog et al., 1993; Havsteen, 2002; Kohno et al., 2002). Recently, numerous studies have indicated that flavonoids could interact with several efflux transporters such as P-glycoprotein, MRP1, and BCRP, suggesting potential drug-flavonoid interactions (Conseil et al., 1998; Leslie et al., 2001; Zhang and Morris, 2003; Zhang et al., 2004). However, the interactions between flavonoids and uptake transporters, especially OATPs, have not been well characterized and remain largely unknown.
Recent in vitro and in vivo fruit juice and fexofenadine interaction studies have indicated that fruit juices and constituents preferentially inhibit OATPs rather than P-glycoprotein (Dresser et al., 2002; Kamath et al., 2005). Fruit juices at 5% of normal strength potently inhibit human OATP1A2 (OATP-A) and at least two rat Oatp transporters. Several flavonoids (quercetin, naringenin, and hesperitin) and their glycosides, at a concentration of 50 μM, significantly inhibit fexofenadine uptake mediated by rat Oatp1a5 (Oatp3) (Dresser et al., 2002), suggesting that flavonoids could interact with some OATP isoforms. However, to our knowledge, the interaction between flavonoids and human OATP1B1 has not been reported. Based on the studies with fruit juices, we hypothesized that some naturally occur-ring flavonoids may also interact with OATP1B1. In the present study, we examined the effects of 20 flavonoids on the uptake of dehydroepiandrosterone sulfate (DHEAS), a well known OATP1B1 substrate, in both OATP1B1-expressing and OATP1B1-negative cells, and investigated the potential mechanism of interaction between flavonoids and OATP1B1.
Materials and Methods
Materials. [3H]DHEAS (60 Ci/mmol, >97% purity) was purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [3H]Biochanin A (14 Ci/mmol, >97% purity) was purchased from Moravek Biochemicals (Brea, CA). Flavonoids and their glycosides were purchased from Sigma-Aldrich (St. Louis, MO) and Indofine Chemical Company (Hillsborough, NJ). Silymarin is composed of silibinin (major component), silydianin, and silychristin, and the molar concentration was calculated based on the molecular weight of silibinin. All other reagents were of either high-performance liquid chromatography or analytical grade and, unless otherwise stated, were purchased from Sigma-Aldrich. Dulbecco's modified Eagle's medium, fetal bovine serum, penicillin, and streptomycin were purchased from Invitrogen (Carlsbad, CA).
Cell Culture. HeLa cells stably transfected with human OATP1B1 were cultured under similar conditions as described previously. Under these conditions, the expression of OATP1B1 in HeLa cells has been shown to be induced significantly and can be readily detected by Western blot analysis (Wang et al., 2003). Briefly, HeLa cells were cultured in 75-cm2 flasks with Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum at 37°C in a humidified 5% CO2/95% air atmosphere. The culture medium also contained 100 units/ml penicillin and 100 μg/ml streptomycin. For uptake and inhibition experiments, the cells were seeded at a density of 5 × 105 cells per 35-mm (diameter) Petri dish. For induction of OATP1B1, cells were cultured in medium containing 100 μM ZnSO4. After a 24-h incubation, an additional 50 μM ZnSO4 was added to this medium for the final 24 h before use.
Uptake Studies. Cells were washed three times with 2 ml of uptake buffer (135 mM NaCl, 1.2 mM MgCl2, 0.81 mM MgSO4, 27.8 mM glucose, 2.5 mM CaCl2, and 25 mM HEPES, pH 7.2). They were then preincubated with 1 ml of uptake buffer at 37°C for 10 min. At the end of the preincubation period, the uptake of [3H]DHEAS (0.5 μM) was performed by incubating cells with 1 ml of uptake buffer containing [3H]DHEAS together with 50 μM flavonoids or the vehicle (0.3% dimethyl sulfoxide at 37°C for 5 min). Rifampicin (100 μM) was used as a positive control. The uptake reaction was stopped by aspiration, followed by washing with 3 ml of ice-cold buffer (137 mM NaCl, and 14 mM Tris base, pH 7.2) three times. The same procedures were used for the [3H]biochanin A (1 μM) uptake experiments. Cells were then solubilized using a solution of 1 ml of 0.3 N NaOH and 1% SDS. Aliquots were used to determine the radioactivity by liquid scintillation counting (1900 CA, Tri-Carb liquid scintillation analyzer; PerkinElmer Life and Analytical Sciences). Results were normalized by protein content determined by the bicinchoninic acid protein assay. The uptake of [3H]DHEAS was expressed as percentage of the control (uptake in OATP1B1-negative cells in the presence of 0.3% dimethyl sulfoxide).
Concentration-Dependent Inhibition Studies. Concentration-dependent studies were performed following the same procedures as described above, except that the effects of flavonoids were tested at varying concentrations (0.3–50 μM). Concentration-dependent effects were determined for three flavonoids [biochanin A, genistein, and epigallocatechin-3-gallate (EGCG)]. Specific uptake was obtained by subtracting [3H]DHEAS uptake into OATP1B1-negative cells from the uptake into OATP1B1-expressing cells. The specific uptake of [3H]DHEAS was expressed as percentage of the control (in the presence of 0.3% DMSO). The IC50 value, the concentration of flavonoid required to inhibit 50% of specific [3H]DHEAS uptake, was obtained by fitting data with the following equation using WinNonlin Professional 2.1 (Pharsight, Mountain View, CA): F = 100 × (1 – Imax · Cγ)/(ICγ50 + Cγ), where C is the concentration of flavonoid, F is the percentage of the specific uptake of [3H]DHEAS, Imax is the maximum percentage of inhibition, and γ is the Hill coefficient. For each flavonoid, the IC50 value (expressed as mean ± S.D.) was determined from three separate experiments and each experiment had triplicate measurements.
Inhibition of DHEAS Uptake by Biochanin A. To determine the inhibition kinetics, [3H]DHEAS (0.5, 1, and 5 μM) uptake was determined at 5 min both in the absence and presence of varying concentrations of biochanin A. The specific uptake was obtained by subtracting [3H]DHEAS uptake into OATP1B1-negative cells from the uptake into OATP1B1-expressing cells. Kinetic parameters were obtained using nonlinear regression fitting (WinNonlin) according to the following equations: v = [Vmax/(1 + I/Ki) × C]/(Km + C) (for noncompetitive kinetics), and v = (Vmax × C)/[Km/(1 + I/Ki) + C] (for competitive kinetics), where v is the specific uptake velocity of the substrate (pmol/mg protein/min), Vmax is the maximum specific uptake rate (pmol/mg protein/min), C is the substrate concentration (μM), Km is the Michaelis-Menten constant (μM), I is the inhibitor concentration, and Ki is the biochanin A inhibition constant. The kinetic model selection was based on visual examination and the goodness of fit (CV% and Akaike's information criterion). Kinetic parameters (expressed as mean ± S.D.) were determined from three separate triplicate experiments.
Statistical Analysis. Data were analyzed for statistically significant differences using ANOVA followed by a Dunnett's post hoc test or by a Student's t test. p values <0.05 were considered statistically significant.
Results
Effects of Flavonoids on [3H]DHEAS Uptake. To determine whether flavonoids modulate OATP1B1 activity, uptake studies were performed with DHEAS (a known OATP1B1 substrate) in OATP1B1-expressing (induced by Zn2+) and OATP1B1-negative (noninduced) HeLa cells in the presence or absence of flavonoids (50 μM) (Fig. 1). The expression of OATP1B1 under the same culture conditions has been characterized previously (Wang et al., 2003). The time course study of [3H]DHEAS uptake indicated that the uptake of DHEAS was linear within a 5-min time period (data not shown). Therefore, we chose 5 min as an appropriate time for [3H]DHEAS uptake studies. As shown in Table 1, in the absence of flavonoids, [3H]DHEAS uptake in OATP1B1-expressing cells was significantly higher than that in OATP1B1-negative control cells (452 ± 105% versus 100 ± 7.93% of the control, p < 0.001). Rifampicin (100 μM) (a known OATP1B1 inhibitor) significantly decreased [3H]DHEAS uptake in OATP1B1-expressing cells (p < 0.01, percentage decrease in mean value, –56.8%) with no significant effects on [3H]DHEAS uptake in OATP1B1-negative cells (p > 0.05, percentage decrease in mean value, –2.88%). All the tested flavonoids except for benzoflavone, chrysin, galangin, and kaempferol produced a significant decrease in [3H]DHEAS uptake in OATP-expressing cells with no or small effects on [3H]DHEAS uptake in OATP1B1-negative cells. The flavonoids biochanin A, fisetin, silibinin, and silymarin produced the greatest effects, resulting in significant decreases in [3H]DHEAS uptake (p < 0.01, percentage decrease in mean values: –68.6%, –56.5%, –66.5%, and –68.9%, respectively), comparable to or even greater than that caused by rifampicin (–56.8%) in OATP1B1-expressing cells. In OATP1B1-negative cells, the flavonoids apigenin, chrysin, fisetin, galangin, luteolin, and silibinin, at a concentration of 50 μM, produced statistically significant increases in [3H]DHEAS uptake (p < 0.01 or p < 0.05, percentage increase in mean values: +92.8%, +17.5%, +37.9%, +17.8%, +17.8%, and + 13.9%, respectively).
Effects of flavonoids on the uptake of [3H]DHEAS in both OATP1B1-expressing and OATP1B1-negative cells
The uptake of [3H]DHEAS (0.5 μM) in both OATP1B1-expressing and OATP1B1-negative cells in the absence or presence of flavonoids (50 μM) was performed as described under Materials and Methods. Rifampicin (100 μM) was used as a positive control. The uptake of [3H]DHEAS was expressed as the percentage of control (uptake in OATP1B1-negative cells in the presence of 0.3% DMSO). The data are expressed as mean ± S.D., n ≥ 6. * p < 0.05, ** p < 0.01 compared with the control of either OATP1B1-expressing or OATP1B1-negative cells. The values in parentheses represent the percentage of decrease or increase in means relative to the control value for either OATP1B1-expressing or OATP1B1-negative cells.
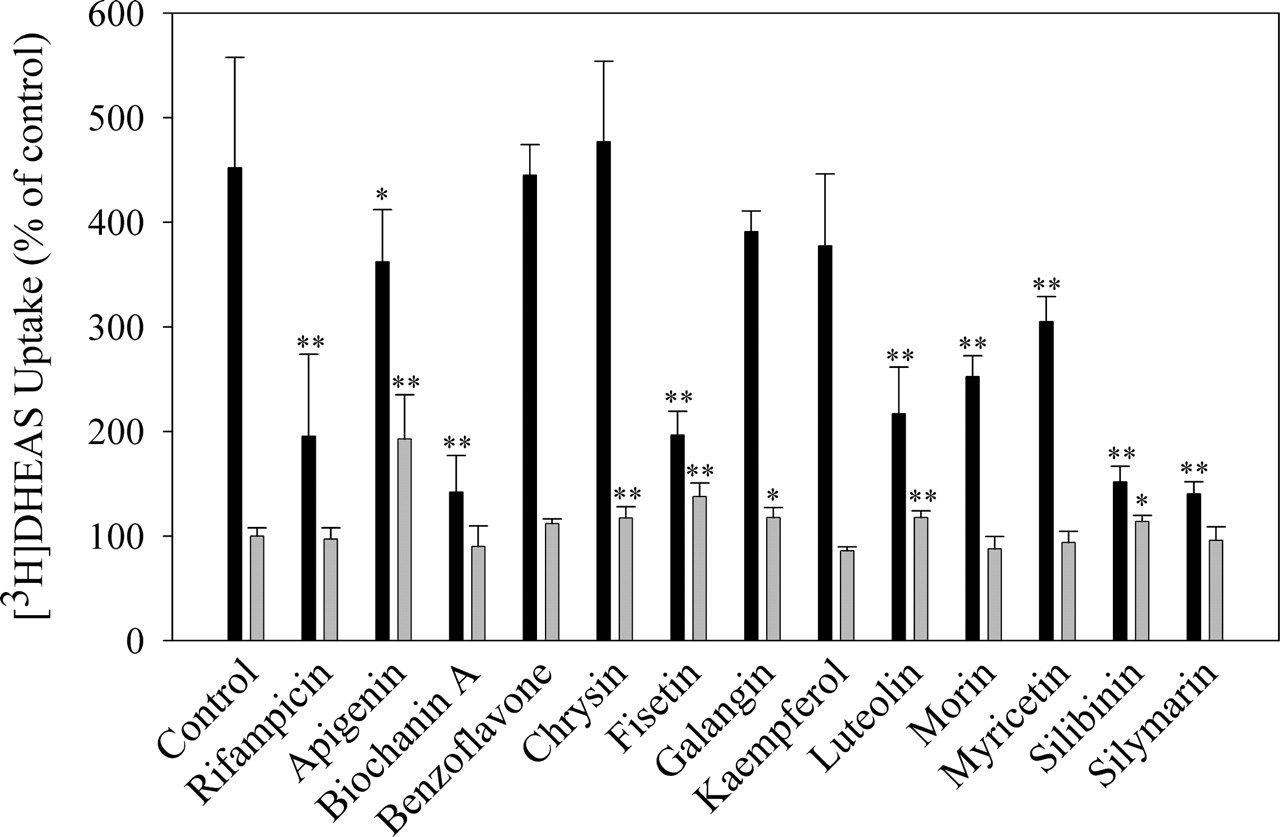
Effects of flavonoids on the uptake of [3H]DHEAS in both OATP1B1-expressing (solid bar) and OATP1B1-negative HeLa cells (shadowed bar). The uptake of [3H]DHEAS (0.5 μM) in both OATP1B1-expressing and OATP1B1-negative cells in the absence or presence of flavonoids (50 μM) was performed as described under Materials and Methods. Rifampicin (100 μM) was used as a positive control. The uptake of [3H]DHEAS is expressed as the percentage of control (uptake in OATP1B1-negative cells in the presence of 0.3% DMSO). The data are expressed as mean ± S.D., n ≥ 6. *, p < 0.05; **, p < 0.01 compared with the control of either OATP1B1-expressing or OATP1B1-negative cells. In the absence of flavonoids, the quantitative level of [3H]DHEAS uptake in OATP1B1-expressing and OATP1B1-negative HeLa cells was 15.0 ± 9.10 and 3.08 ± 1.22 pmol/mg protein, respectively (mean ± S.D., p < 0.001).
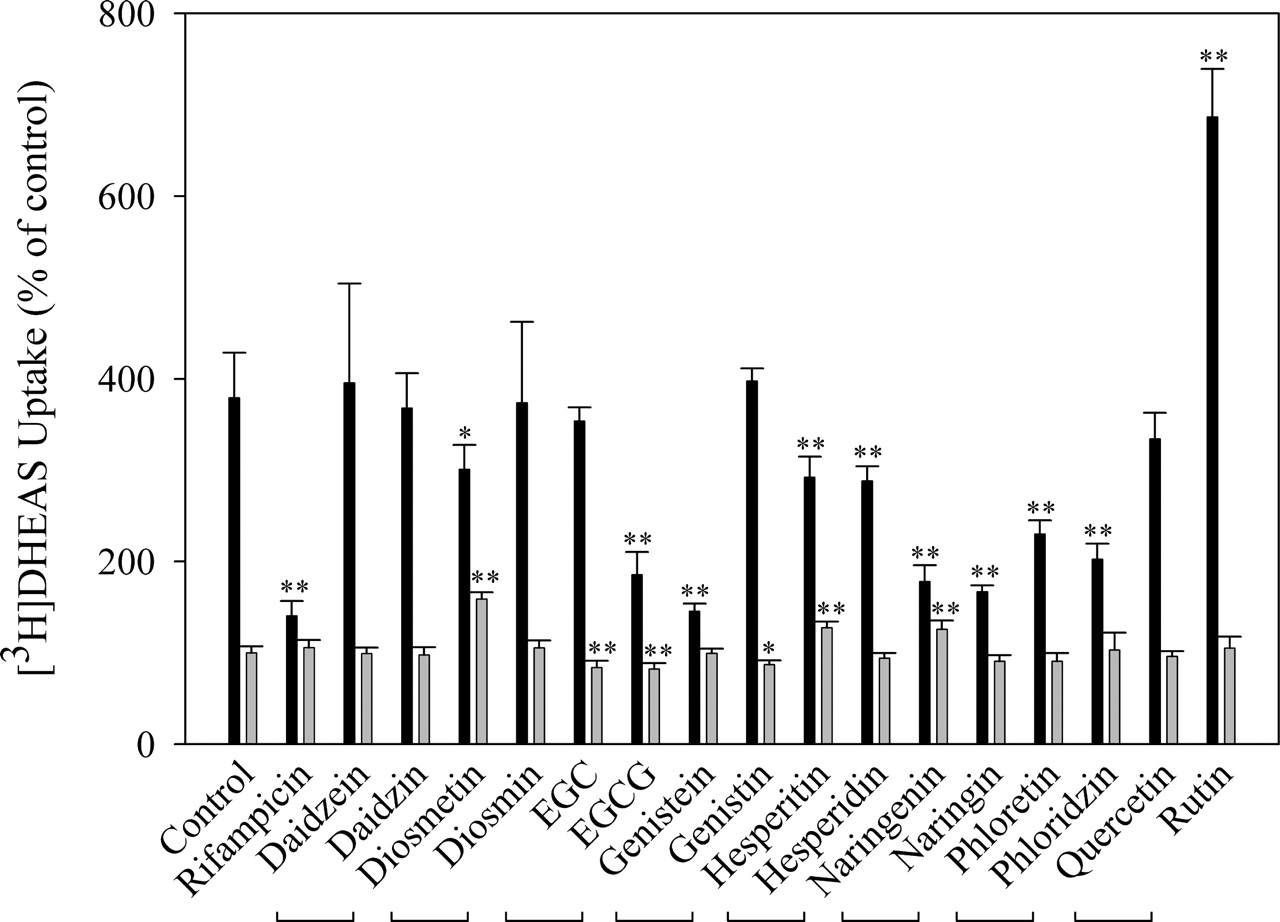
Effects of Flavonoids and Their Glycosides on [3H]DHEAS Uptake. Many flavonoids exist in both aglycone and glycone forms. Different biochemical and pharmacological activities between these two forms have been reported for a number of flavonoids (Kwon et al., 2004; Lin et al., 2005). To compare the effects of flavonoids and their glycosides on [3H]DHEAS uptake, eight pairs of flavonoids and corresponding glycosides were characterized in both OATP1B1-expressing and OATP1B1-negative cells (Fig. 2). As shown in Table 2, in OATP1B1-expressing cells, both daidzein and its glycoside daidzin had no effects on [3H]DHEAS uptake (p > 0.05, percentage change in mean values: +4.34% and –2.94%, respectively). Three pairs of flavonoids and their corresponding glycosides (hesperitin/hesperidin, naringenin/naringin, and phloretin/phloridzin) all significantly decreased [3H]DHEAS uptake (p < 0.01). On the other hand, genistein and diosmetin significantly inhibited [3H]DHEAS uptake (p < 0.01 or p < 0.05, percentage decrease in mean values: –61.6% and –20.6%, respectively) with no significant effects found for their glycosides genistin and diosmin (p > 0.05, percentage change in mean values: +4.88% and –1.36%, respectively). In contrast, epigallocatechin (EGC) and quercetin had no significant effects (p > 0.05, percentage change in mean values: –6.70% and –11.8%, respectively) on [3H]DHEAS uptake, whereas their glycosides EGCG and rutin significantly decreased and increased [3H]DHEAS uptake, respectively (p < 0.01, percentage change in mean values: –51.0% and +81.1%, respectively). In OATP1B1-negative control cells, most flavonoids and their glycosides had negligible effects on [3H]DHEAS uptake. However, diosmetin, hesperitin, and naringenin, at a concentration of 50 μM, produced small but statistically significant increases in [3H]DHEAS uptake (p < 0.01, percentage increase in mean values: +59.1%, +27.5%, and + 26.0%, respectively). EGC, EGCG, and genistin resulted in small but statistically significant decreases in [3H]DHEAS uptake (p < 0.01 or p < 0.05, percentage decrease in mean values: –16.1%, –17.6%, and –12.8%, respectively).
Effects of flavonoids and their glycosides on the uptake of [3H]DHEAS in both OATP1B1-expressing and OATP1B1-negative cells
The uptake of [3H]DHEAS (0.5 μM) in both OATP1B1-expressing and OATP1B1-negative cells in the absence or presence of flavonoids (50 μM) was performed as described under Materials and Methods. Rifampicin (100 μM) was used as a positive control. The uptake of [3H]DHEAS was expressed as the percentage of control (uptake in OATP1B1-negative cells in the presence of 0.3% DMSO). The data are expressed as mean ± S.D., n ≥ 6. * P < 0.05; ** P < 0.01 compared with the control of either OATP1B1-expressing or OATP1B1-negative cells. The values in parentheses represent the percentage of decrease or increase in means relative to the control value for either OATP1B1-expressing or OATP1B1-negative cells.
Concentration-Dependent Effects of Flavonoids on [3H]DHEAS Uptake. The concentration-dependent effects of flavonoids on [3H]DHEAS uptake were investigated for three flavonoids, biochanin A, genistein, and EGCG, which demonstrated high OATP1B1 inhibition activity when tested at a concentration of 50 μM. As shown in Fig. 3, the inhibition of [3H]DHEAS uptake by biochanin A, genistein, and EGCG were flavonoid concentration-dependent. At 10 μM, all the tested flavonoids had significant effects on [3H]DHEAS uptake (p < 0.001). In addition, 3 μM biochanin A significantly decreased [3H]DHEAS uptake (Fig. 3; 85.8 ± 4.14% of the control, p < 0.001). The IC50 values for biochanin A, genistein, and EGCG for inhibiting [3H]DHEAS uptake were 11.3 ± 3.22, 14.9 ± 3.76, and 14.1 ± 1.44 μM, respectively.

Effects of flavonoids and their glycosides on the uptake of [3H]DHEAS in both OATP1B1-expressing (solid bar) and OATP1B1-negative HeLa cells (shadowed bar). The uptake of [3H]DHEAS (0.5 μM) in both OATP1B1-expressing and OATP1B1-negative cells in the absence or presence of flavonoids (50 μM) was performed as described under Materials and Methods. The two flavonoids with a connecting line represent a flavonoid and its corresponding glycoside. Rifampicin (100 μM) was used as a positive control. The uptake of [3H]DHEAS is expressed as the percentage of control (uptake in OATP1B1-negative cells in the presence of 0.3% DMSO). The data are expressed as mean ± S.D., n ≥ 6. *, p < 0.05, **, p < 0.01 compared with the control of either OATP1B1-expressing or OATP1B1-negative cells.
Uptake of [3H]Biochanin A by OATP1B1. To further characterize the OATP1B1-modulating activities of the flavonoids, biochanin A was used as a model flavonoid and the uptake of [3H]biochanin A was studied in both OATP1B1-expressing and OATP1B1-negative cells. As shown in Fig. 4, [3H]biochanin A uptake was similar in OATP1B1-expressing and OATP1B1-negative HeLa cells in the absence of 100 μM rifampicin (1.27 ± 0.23 and 1.20 ± 0.22 nmol/mg protein, respectively, p > 0.05), as well as in the presence of 100 μM rifampicin (1.30 ± 0.23 and 1.23 ± 0.21 nmol/mg protein, respectively, p > 0.05).
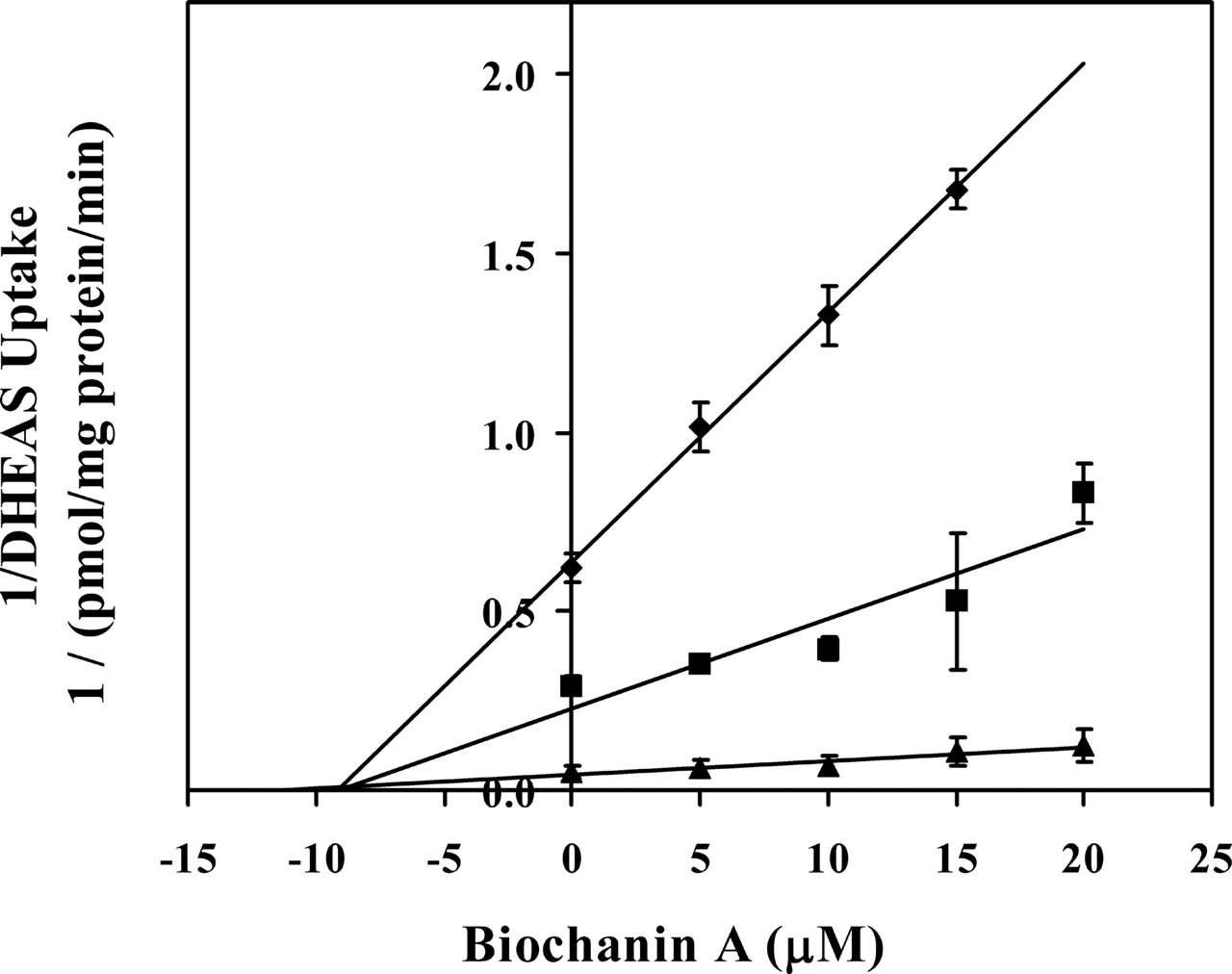
Inhibition of OATP1B1-Mediated Uptake of [3H]DHEAS by Biochanin A. To gain insight into the mechanism underlying the inhibition of OATP1B1-mediated uptake of DHEAS by biochanin A, [3H]DHEAS uptake kinetics in the absence and presence of biochanin A was determined by Dixon plot analysis as shown in Fig. 5. Based on visual examination and computer fitting results (data not shown), the kinetic study revealed that biochanin A inhibited OATP1B1-mediated [3H]DHEAS uptake in a noncompetitive manner with a Ki value of 10.2 ± 1.89 μM, and a Vmax value of 154 ± 7.61 pmol/mg protein.
Discussion
OATPs are key membrane transporters that mediate the sodium-independent uptake of a wide range of endogenous and exogenous compounds, including drugs in clinical use (Hagenbuch and Meier, 2004). The importance of this transporter family in drug disposition has been increasingly recognized. The coexpression of OATPs with some efflux transporters in multiple tissues and their partially overlapping substrate specificity suggest that OATPs may play an important role together with efflux transporters in overall drug absorption and disposition (Kim, 2003). OATP1B1, a well studied human OATP, is one of the major sodium-independent bile salt transporters localized on the basolateral membrane of human hepatocytes (Trauner and Boyer, 2003). OATP1B1 transports a large number of structurally divergent compounds and has been implicated as an important determinant in hepatic drug uptake and elimination (Kim, 2003). Thus, analogous to the cases of efflux transporters, modulation of OATP1B1 may alter the pharmacokinetics of OATP1B1 substrate drugs, causing potential drug-drug interactions.
Flavonoids are a class of polyphenolic compounds widely present in diet and herbal products with exceptional safety records (Havsteen, 2002). In addition to their many anticancer properties, flavonoids have been found to interact with several efflux transporters (Conseil et al., 1998; Leslie et al., 2001; Zhang et al., 2004). Recent studies have indicated that several flavonoids present in fruit juices significantly inhibit fexofenadine uptake mediated by rat Oatp1a5 (Oatp3) at a concentration of 50 μM (Dresser et al., 2002). Therefore, it is likely that some flavonoids may also be modulators of OATP1B1. The elucidation of the effects of flavonoids on OATP1B1-mediated transport may help to better predict potential flavonoid-drug pharmacokinetic interactions, which is an important issue considering the widespread consumption of large amounts of flavonoids in food or herbal products.
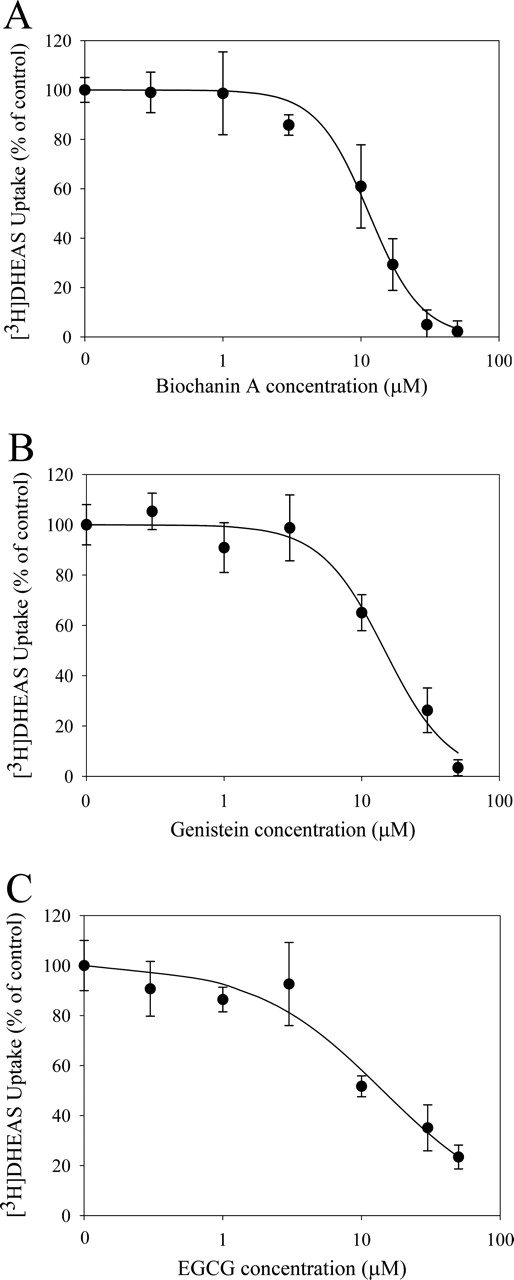
Concentration-dependent effects of flavonoids on [3H]DHEAS uptake in HeLa cells. Concentration-dependent effects were determined for three flavonoids [biochanin A (A), genistein (B), and EGCG (C)] in the presence of varying concentrations (0.3–50 μM) of flavonoids or vehicle (0.3% DMSO). Specific uptake was obtained by subtracting DHEAS uptake into OATP1B1-negative HeLa cells from the uptake into OATP1B1-expressing HeLa cells. The specific uptake of [3H]DHEAS is expressed as percentage of the control (in the presence of 0.3% DMSO). Each data point represents the mean ± S.D., determined from three separate triplicate experiments.

Uptake of [3H]biochanin A in both OATP1B1-expressing (solid bar) and OATP1B1-negative HeLa cells (open bar). The uptake of [3H]biochanin A (1 μM) in both OATP1B1-expressing and OATP1B1-negative cells in the absence or presence of rifampicin (100 μM) was performed as described under Materials and Methods. The uptake of [3H]biochanin A is given as mean ± S.D., n = 6.

Dixon plot analysis of inhibition of OATP1B1-mediated uptake of [3H]DHEAS by biochanin A in HeLa cells. The uptake of [3H]DHEAS at the fixed concentrations of 0.5 μM (♦), 1 μM (▪), and 5 μM (▴) was measured both in the absence and presence of the indicated inhibitor concentration. Data represent the reciprocal of the uptake rate difference between OATP1B1-expressing cells and OATP-negative cells. Each data point represents the mean ± S.D., determined from three separate triplicate experiments.
In the present study, we examined the effects of 20 flavonoids, representing all the chemical subclasses of flavonoids, on OATP1B1-mediated DHEAS transport. DHEAS, one of the major steroids in the systemic circulation, is a known substrate for several OATPs (Kim, 2003) as well as MRPs (Zelcer et al., 2003). For OATP1B1, a 3.2- to 7.3-fold ratio of DHEAS uptake in OATP1B1-expressing cells over control cells was observed in different experimental systems (Konig et al., 2000; Kullak-Ublick et al., 2001). The high uptake ratio of DHEAS indicates that it is a suitable substrate for the study of OATP1B1-mediated transport. In this study, we used an OATP1B1-expressing HeLa cell line, in which the expression of OATP1B1 can be substantially induced by Zn2+ (Wang et al., 2003). Although very low levels of some endogenous transporters, such as P-glycoprotein and MRP1/2 (Sakaeda et al., 2002), are present in parent HeLa cells and may possibly contribute to the DHEAS transport, we observed a 3- to 4-fold net difference in DHEAS uptake between OATP1B1-expressing and OATP1B1-negative cells in the absence of flavonoids. This net difference in DHEAS uptake is most likely due to OATP1B1 since substantial OATP1B1 expression is induced in the presence of Zn2+ and may represent the major difference between OATP1B1-expressing and OATP1B1-negative cells. In OATP1B1-expressing cells, the flavonoids biochanin A, fisetin, silibinin, and silymarin (50 μM) produced significant decreases in [3H]DHEAS uptake, which were comparable to or even greater than that caused by 100 μM rifampicin, indicating that biochanin A, fisetin, silibinin, and silymarin strongly inhibited the OATP1B1-mediated uptake of [3H]DHEAS. The flavonoids apigenin, luteolin, morin, and myricetin produced smaller, but statistically significant decreases in [3H]DHEAS uptake in OATP1B1-expressing cells with no or small effects in OATP1B1-negative cells, suggesting that these flavonoids are also OATP1B1 inhibitors. Some flavonoids (apigenin, chrysin, fisetin, galangin, luteolin, and silibinin), at a concentration of 50 μM, produced statistically significant increases in [3H]DHEAS uptake in OATP1B1-negative cells. The exact reason(s) for this is currently unknown but might possibly be due to alterations in cell membrane permeability by these flavonoids at high concentrations and/or the interaction of these flavonoids with an endogenous transporter(s) expressed in HeLa cells that mediates [3H]DHEAS transport. A very low level of endogenous MRP1, which was detected by real-time polymerase chain reaction in HeLa cells (Sakaeda et al., 2002), may be responsible for the small changes in [3H]DHEAS uptake observed, since some flavonoids may also interact with MRP1 (Nguyen et al., 2003) and DHEAS seems to be transported by MRP1 as well (Zelcer et al., 2003).
To compare the effects of flavonoids and their glycosides on [3H]DHEAS uptake, eight pairs of flavonoids and their corresponding glycosides were characterized in both OATP1B1-expressing and OATP1B1-negative cells. In OATP1B1-expressing cells, three pairs of flavonoids and glycosides (hesperitin/hesperidin, naringenin/naringin, and phloretin/phloridzin) all significantly decreased [3H]DHEAS uptake (p < 0.01) when compared with the uptake value in the absence of flavonoids. Fruit juices and their major flavonoid components hesperitin and naringenin, and their glycosides, have been shown to significantly inhibit fexofenadine uptake mediated by rat Oatp1a5 (Oatp3) (Dresser et al., 2002). The potent inhibitory effects of these flavonoids on OATP1B1-mediated uptake of [3H]DHEAS indicate that fruit juices may also represent potent inhibitors of OATP1B1, suggesting the potential for hepatic drug interactions. Interestingly, both daidzein and its glycoside daidzin had no effect on [3H]DHEAS uptake. Daidzein is an isoflavone with a chemical structure similar to that of biochanin A and genistein, except that no hydroxyl group is present at the 5-position of the A-ring in the daidzein structure. Considering the potent inhibitory effects of biochanin A and genistein on [3H]DHEAS uptake, it is reasonable to speculate that attachment of a hydroxyl group at the 5-position may markedly attenuate or totally abolish OATP1B1-inhibitory activity of the flavonoids. On the other hand, genistein and diosmetin significantly inhibited [3H]DHEAS uptake with no significant effects found for their glycosides genistin and diosmin. In contrast, EGC and quercetin had no significant effects on [3H]DHEAS uptake, whereas their glycosides EGCG and rutin significantly decreased and increased [3H]DHEAS uptake, respectively. These results indicate that attachment of a sugar moiety may differently modulate OATP1B1 activity, suggesting that glycosidation of flavonoid aglycones should be considered as an important modulator of the biological activities of flavonoids. Interestingly, rutin is the only flavonoid identified in this study that can stimulate OATP1B1 activity. Although the reason for such increase is not clear at present, increased substrate uptake has been reported for some other OATP isoforms, such as rat Oatp1a4 (Oatp2) (Sugiyama et al., 2002) and human OATP2B1 (OATP-B) (Tamai et al., 2001). Future studies will be essential to clarify the underlying mechanisms.
It should be noted that, in OATP1B1-negative control cells, most flavonoids and their glycosides had negligible effects on [3H]DHEAS uptake with the exception of diosmetin, hesperitin, and naringenin, which, at a concentration of 50 μM, produced small but statistically significant increases in [3H]DHEAS uptake. Interestingly, EGC, EGCG, and genistin resulted in small but statistically significant decreases in [3H]DHEAS uptake. The exact reason(s) for these changes is currently unknown but may possibly be due to inhibition of a low endogenous OATP1B1 expression in OATP1B1-negative HeLa cells, resulting in lower uptake, although OATP1B1 expression in these cells was not detectable by Western blot analysis (Wang et al., 2003). Additionally, inhibition of an efflux transporter such as MRP1 may result in higher uptake values.
To investigate the concentration-dependent effects of flavonoids on OATP1B1-mediated transport, the effects of three flavonoids with strong OATP1B1-inhibitory activities when tested at a concentration of 50 μM, namely, biochanin A, genistein, and EGCG, were evaluated. These three flavonoids demonstrated concentration-dependent inhibition of OATP1B1. Biochanin A seems to be one of the most potent OATP1B1 inhibitors with an IC50 value of 11.3 ± 3.22 μM. Significant inhibition of OATP1B1 by biochanin A could be produced at concentrations as low as 3 μM. Our laboratory, as well as others, previously investigated flavonoids as inhibitors of P-glycoprotein-, MRP-, and BCRP-mediated drug resistance in cancer cell lines and demonstrated that some flavonoids, such as biochanin A and genistein, can also inhibit P-glycoprotein, MRP1, and BCRP (Castro and Altenberg, 1997; Nguyen et al., 2003; Zhang and Morris, 2003; Zhang et al., 2004), indicating that these flavonoids may potentially affect in vivo drug disposition by inhibiting different transporter systems. Some other flavonoids, however, preferentially inhibit one major transporter over the others. For example, chrysin has been shown to be a potent BCRP inhibitor (Zhang et al., 2004), whereas it has no OATP1B1-inhibitory activity, as shown in the present study. In contrast, EGCG is not a BCRP inhibitor (Zhang et al., 2004), whereas it can effectively inhibit OATP1B1 activity. Future studies regarding the structure-activity relationships for the flavonoid-mediated inhibition of OATP1B1, together with the existing information on some major efflux transporters, may help us better understand and design inhibitors with preferred potency and specificity.
To understand the mechanism of OATP1B1 inhibition by flavonoids, biochanin A was selected as a model flavonoid and the interaction between biochanin A and OATP1B1 was further characterized. The uptake studies indicated that [3H]biochanin A uptake was similar in OATP1B1-expressing and OATP1B1-negative HeLa cells in the absence of 100 μM rifampicin, as well as in the presence of 100 μM rifampicin, suggesting that biochanin A is not likely a substrate of OATP1B1. This is expected since biochanin A itself is a neutral compound and most substrates of OATP1B1 identified so far are organic anions (Hagenbuch and Meier, 2004). The kinetics study revealed that biochanin A inhibited OATP1B1-mediated [3H]DHEAS uptake in a noncompetitive manner with a Ki value of 10.2 ± 1.89 μM, and a Vmax value of 154 ± 7.61 pmol/mg protein. This is in general agreement with some other OATP1B1 inhibitors, such as cyclosporin A and indinavir (Tirona et al., 2003), which have been shown to inhibit OATP1B1-mediated estradiol glucuronide uptake in a noncompetitive manner as well (Campbell et al., 2004).
In conclusion, many naturally occurring flavonoids can modulate OATP1B1-mediated uptake of [3H]DHEAS, indicating that they are a novel class of OATP1B1 modulators. From studies with biochanin A, investigating the possible mechanism(s) of this interaction, we have found that biochanin A is not a substrate for OATP1B1 and it represents a noncompetitive inhibitor. From the drug interaction point of view, considering the high consumption of flavonoids in the diet and in herbal products, hepatic uptake interactions of these flavonoids with drugs that are OATP1B1 substrates may result following coadministration.
Acknowledgments
We thank Dr. Shuzhong Zhang for critical review and helpful suggestions.
Footnotes
-
Financial support for this study was provided by grants from the Susan G. Komen Breast Cancer and Cancer Research and Prevention Foundations.
-
Article, publication date, and citation information can be found at http://dmd.aspetjournals.org.
-
doi:10.1124/dmd.105.005926.
-
ABBREVIATIONS: OATP, organic anion-transporting polypeptide; BCRP, breast cancer resistance protein; DHEAS, dehydroepiandrosterone sulfate; MRP, multidrug resistance-associated protein; EGCG, epigallocatechin-3-gallate; EGC, epigallocatechin; DMSO, dimethyl sulfoxide.
- Received June 6, 2005.
- Accepted August 4, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}