


Abstract
Zonampanel monohydrate (YM872; [2,3-dioxo-7-(1H-imidazol-1-yl)-6-nitro-1,2,3,4-tetrahydro-1-quinoxalinyl]acetic acid monohydrate) is a novel α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptor antagonist. The major elimination route for zonampanel has been reported to be by urine via the kidneys. The purpose of this study is to elucidate the molecular mechanism of the renal excretion of zonampanel using cells stably expressing human organic anion transporters (hOAT) 1, hOAT2, hOAT3, and hOAT4, as well as human organic cation transporters (hOCT) 1 and hOCT2. Another AMPA receptor antagonist, YM90K [6-(1H-imidazol-1-yl)-7-nitro-2,3(1H,4H)-quinoxalinedione monohydrochloride], a decarboxymethylated form of zonampanel, was also used for comparing the substrate specificity. Zonampanel inhibited the uptake of prototypical organic anion substrates, [14C]para-aminohippurate in hOAT1 and [3H]estrone sulfate in hOAT3 and hOAT4, in a competitive manner. A time- and concentration-dependent increase in [14C]zonampanel uptake was observed in cells expressing hOAT1, hOAT3, and hOAT4. The Km values of zonampanel uptake by hOAT1, hOAT3, and hOAT4 were 1.4, 7.7, and 215 μM, respectively. Considering the localization of each transporter, results suggest that zonampanel is taken up via hOAT1 and hOAT3 from the blood into proximal tubular cells and then effluxed into the lumen via hOAT4. Probenecid and cimetidine competitively inhibited [14C]zonampanel uptake by the hOATs (hOAT1, hOAT3, and hOAT4 for probenecid; hOAT3 for cimetidine). YM90K inhibited the uptake of the prototypical substrate via hOAT3 competitively, but the uptake via hOAT1 noncompetitively. These findings suggest that the prototypical organic anion substrates (para-aminohippurate and estrone sulfate), cimetidine, probenecid, and zonampanel share binding specificity in each hOAT, whereas YM90K does not in hOAT1, possibly due to it being the decarboxymethylated form.
The excessive synaptic release of glutamate and activation of postsynaptic glutamate receptors are considered to mediate ischemia-induced neuronal damage in stroke victims. In a wide variety of cerebral ischemia animal models, receptor antagonists against α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA), a glutamate receptor subtype, have been shown to be neuroprotective (Sheardown et al., 1990; Gill et al., 1992; Bullock et al., 1994; Gill 1994; Xue at al., 1994; Shimizu-Sasamata et al., 1996). However, the poor water solubility and resultant renal toxicity of the first-generation compounds may have limited their use in patients (Xue at al., 1994). Zonampanel monohydrate (YM872; [2,3-dioxo-7-(1H-imidazol-1-yl)-6-nitro-1,2,3,4-tetrahydro-1-quinoxalinyl]acetic acid monohydrate; Fig. 1) is a novel competitive AMPA receptor antagonist that is highly water soluble yet maintains selectivity and potency for AMPA receptors (Kohara et al., 1998; Takahashi et al., 1998b). Zonampanel monohydrate has been shown to reduce the volume of ischemic damage in rats (Kawasaki-Yatsugi et al., 1998, 2000; Takahashi et al., 1998a) and cats (Takahashi et al., 1998b) subjected to transient or permanent middle cerebral artery occlusion.

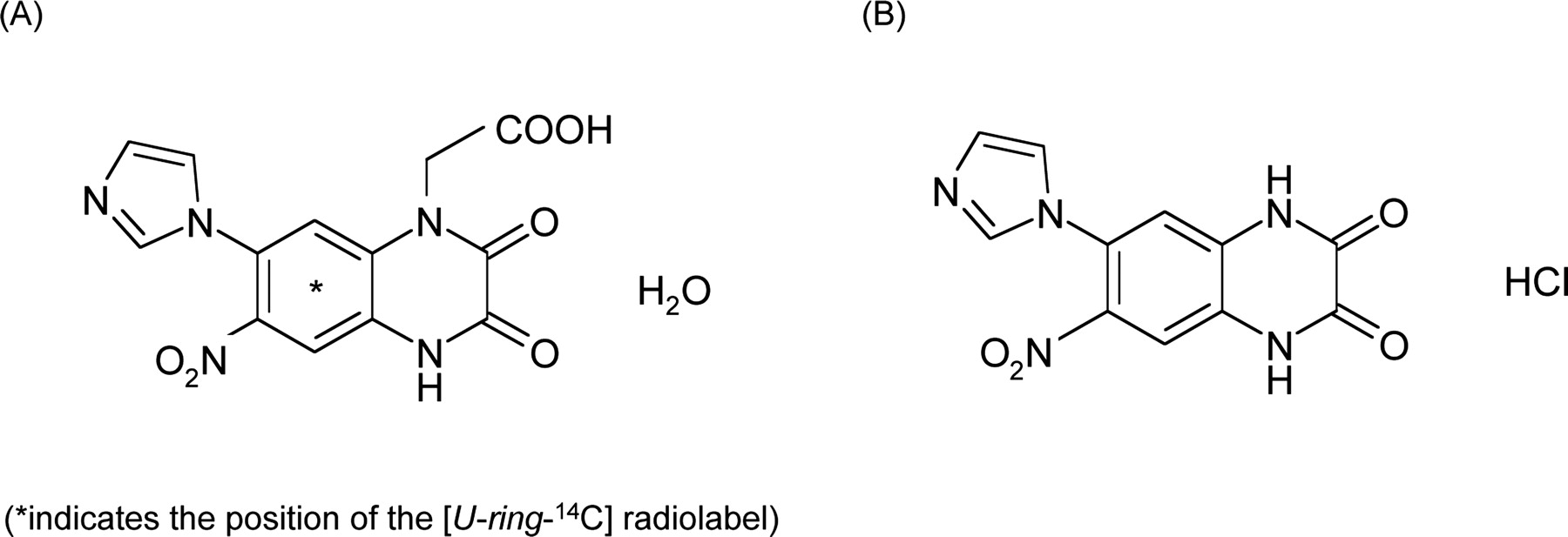
Chemical structures of zonampanel monohydrate (A) and YM90K (B).
It has been found that the major elimination route for zonampanel monohydrate in humans and animals is by renal excretion. Zonampanel monohydrate in the body was excreted into urine mainly as the unchanged form within 2 h after completion of intravenous infusion in humans, and fecal excretion was very low (unpublished observation). In addition, renal clearance of zonampanel monohydrate was much higher than the product of the fraction unbound in plasma and the glomerular filtration rate. Therefore, it is considered that renal tubular secretion plays an important role in the renal excretion of this compound.
Renal tubular secretion is accomplished through two steps of membrane transport: the uptake from blood through the basolateral membrane of the epithelial cells in the proximal tubule, and the efflux into the tubular lumen from the epithelial cells through the apical membrane of the cells. These membrane transport processes are mediated by transporters located separately in the basolateral and apical membranes. Organic anion transporters (OATs) and organic cation transporters (OCTs) are the major transporters involved in the renal tubular secretion of various endogenous metabolites, drugs, and xenobiotics (Pritchard and Miller, 1993; Inui et al., 2000; Sekine et al., 2000). In the human proximal tubule, it has been revealed that human organic anion transporter (hOAT) 1, hOAT2, hOAT3, and human organic cation transporter (hOCT) 2 are localized in the basolateral membrane of the tubular cells (Hosoyamada et al., 1999; Cha et al., 2001; Pietig et al., 2001; Enomoto et al., 2002), whereas hOAT4 is localized in the apical membrane of the cells (Babu et al., 2002a). However, the localization of hOCT1 in the human kidney remains unclear.
The purpose of this study is to elucidate the molecular mechanism of the renal excretion of zonampanel. For this purpose, we utilized cells from the second portion of the proximal tubule (S2 cells) that stably expressed the hOATs (hOAT1, hOAT2, hOAT3, and hOAT4) and the hOCTs (hOCT1 and hOCT2). In addition, we examined the differences in inhibitory effects on hOATs between zonampanel and YM90K [6-(1H-imidazol-1-yl)-7-nitro-2,3(1H,4H)-quinoxalinedione monohydrochloride; Fig. 1], a structural analog of zonampanel.
Materials and Methods
Materials. [14C]Zonampanel monohydrate (specific radioactivity: 1.23 GBq/mmol) was synthesized at Amersham Biosciences UK, Ltd. (Little Chal-font, Buckinghamshire, UK) (Fig. 1). Zonampanel monohydrate and YM90K were synthesized by the Chemical Technology Laboratories of Yamanouchi Pharmaceutical Co., Ltd. [14C]para-Aminohippurate ([14C]PAH) (2 GBq/mmol), [3H]prostaglandin F2α ([3H]PGF2α) (6808 GBq/mmol), and [3H]estrone sulfate ([3H]ES (1961 GBq/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [14C]Tetraethylammonium ([14C]TEA) (2 GBq/mmol) was obtained from American Radiolabeled Chemicals (St. Louis, MO). PAH, PGF2α, tris salt, and ES sodium salt were purchased from Sigma-Aldrich (St. Louis, MO). TEA bromide was obtained from Wako Pure Chemicals Industries, Ltd. (Osaka, Japan). Other materials used included fetal bovine serum, trypsin, and geneticin from Invitrogen (Carlsbad, CA), recombinant epidermal growth factor from Wakunaga (Hiroshima, Japan), insulin from Shimizu (Shizuoka, Japan), RITC 80-7 culture medium from Iwaki Co. (Tokyo, Japan), and TfX-50 from Promega (Madison, WI). All other chemicals and reagents were of analytical grade and purchased from commercial sources.
Preparation of [14C]Zonampanel, Zonampanel, and YM90K Solutions. Stock solutions of [14C]zonampanel monohydrate (3 mM; 3.68 MBq/ml), zonampanel monohydrate (2 mM), and YM90K (600 μM) were dissolved in Dulbecco's modified phosphate-buffered saline (D-PBS, Sigma-Aldrich).
Cell Culture and Establishment of S2 hOATs and S2 hOCTs. S2 hOATs and S2 hOCTs were obtained by the method described by Takeda et al. (2002a,b). S2 cells transfected with pcDNA 3.1 (Invitrogen), a mammalian expression vector lacking an insert, and pSV2neo, a neomycin-resistant gene, were designated as S2 pcDNA 3.1 (mock) and used as the control. These cells were used for 10 to 40 passages. Clonal cells were isolated using a cloning cylinder and screened by determining the saturable uptake of the optimal substrate for each transporter, i.e., [14C]PAH for hOAT1 (Hosoyamada et al., 1999), [3H]PGF2α for hOAT2 (Enomoto et al., 2002), [3H]ES for hOAT3 (Cha et al., 2001) and hOAT4 (Cha et al., 2000), and [3H]TEA for hOCT1 and hOCT2 (Okuda et al., 1996; Zhang et al., 1998).
Inhibition Study. First, we studied the inhibitory effects of zonampanel and YM90K at a high concentration (1 mM zonampanel, and 300 μM YM90K, because of poor solubility) on the uptake of prototypical substrates for hOATs and hOCTs. Subsequently, the IC50 values of zonampanel or YM90K, upon the uptake of prototypical substrates, were estimated for hOATs in which more than 50% inhibition by 1 mM zonampanel or 300 μM YM90K had been observed. For hOATs in which an IC50 value of less than 300 μM was observed, the Ki values were also determined. In these inhibition studies, the cells were incubated at 37°C in 0.5 ml of D-PBS containing either [14C]PAH (2 min; for hOAT1), [3H]PGF2α (0.5 min; for hOAT2), [3H]ES (2 min; for hOAT3 and hOAT4), or [3H]TEA (15 min for hOCT1 and 5 min for hOCT2) in the absence or presence of zonampanel or YM90K. As for the experiments using zonampanel at 1 mM and YM90K at 300 μM, and those for the determination of IC50 values, concentrations of prototypical substrates were set as follows: [14C]PAH, 5 μM; [3H]PGF2α, 50 nM; [3H]ES, 50 nM; [3H]TEA, 5 μM. In addition, to evaluate the inhibitory effects of probenecid and cimetidine on the uptake of zonampanel and the prototypical substrates, the cells (S2 hOAT1, S2 hOAT3, and S2 hOAT4 for probenecid; S2 hOAT3 for cimetidine) were incubated at 37°C for 2 min (for uptake of the prototypical substrates) and 5 min (for zonampanel uptake) in D-PBS containing various concentrations of [14C]zonampanel or prototypical substrates, in the absence or presence of either probenecid or cimetidine. Because of the high CV (coefficient of variation) of low radioactivity counts in the mock, the values for uptake in the mock were not subtracted from those in S2 hOATs or S2 hOCTs. For the kinetic analysis of the inhibitory effects, Lineweaver-Burk plot analyses were performed. When the inhibition was competitive, the Ki values were calculated based on the equation, Ki = concentration of inhibitor/[(Km of substrate with inhibitor/Km of substrate without inhibitor) - 1]. When the inhibition was noncompetitive, Ki = concentration of inhibitor/[(Vmax of substrate without inhibitor/Vmax of substrate with inhibitor) - 1].
Uptake Experiments for Zonampanel. Time-dependence for the uptake of [14C]zonampanel was examined at 10 μM. Concentration dependence for the uptake of [14C]zonampanel was also examined for the transporters in which significant time-dependent uptake was observed. The Km and Vmax values were estimated using Eadie-Hofstee plots. Because of the high CV of low radioactivity counts in the mock, the values for uptake in the mock were not subtracted from those in each of the S2 hOATs. Uptake experiments were performed as described previously (Kimura et al., 2002; Takeda et al., 2002b). Briefly, S2 hOATs, S2 hOCTs, and S2 pcDNA 3.1 (mock) were seeded in 24-well tissue culture plates (Corning, Costar 3524) at a cell density of 1 × 105 cells/well. After being cultured for 2 days, the cells were washed three times with D-PBS (containing 137 mM NaCl, 3 mM KCl, 8 mM NaHPO4, 1 mM KH2PO4, 1 mM CaCl2, and 0.5 mM MgCl2; pH 7.4) and then preincubated in the same solution in a water bath set at 37°C for 10 min. Subsequently, the solution was removed, and the cells were incubated with 0.5 ml of [14C]zonampanel at 37°C in D-PBS, at various concentrations, for the period as indicated in each experiment. After removing the [14C]zonampanel solution, the uptake was stopped by the addition of ice-cold D-PBS, and the cells were washed three times with fresh portions of the same ice-cold D-PBS. The cells in each well were lysed with 0.5 ml of 0.1 N sodium hydroxide and mixed with 2.5 ml of Aquasol-2 (PerkinElmer Life and Analytical Sciences), and the radioactivity was measured using a β-scintillation counter (LSC-6100; Aloka, Tokyo, Japan).
Statistical Analysis. Data are expressed as the mean ± S.E. Statistical differences in the uptake of prototypical substrate for each hOAT and hOCT with or without 1 mM zonampanel or 300 μM YM90K were determined using the Student's unpaired t test. Differences were considered significant at P < 0.05.
Results
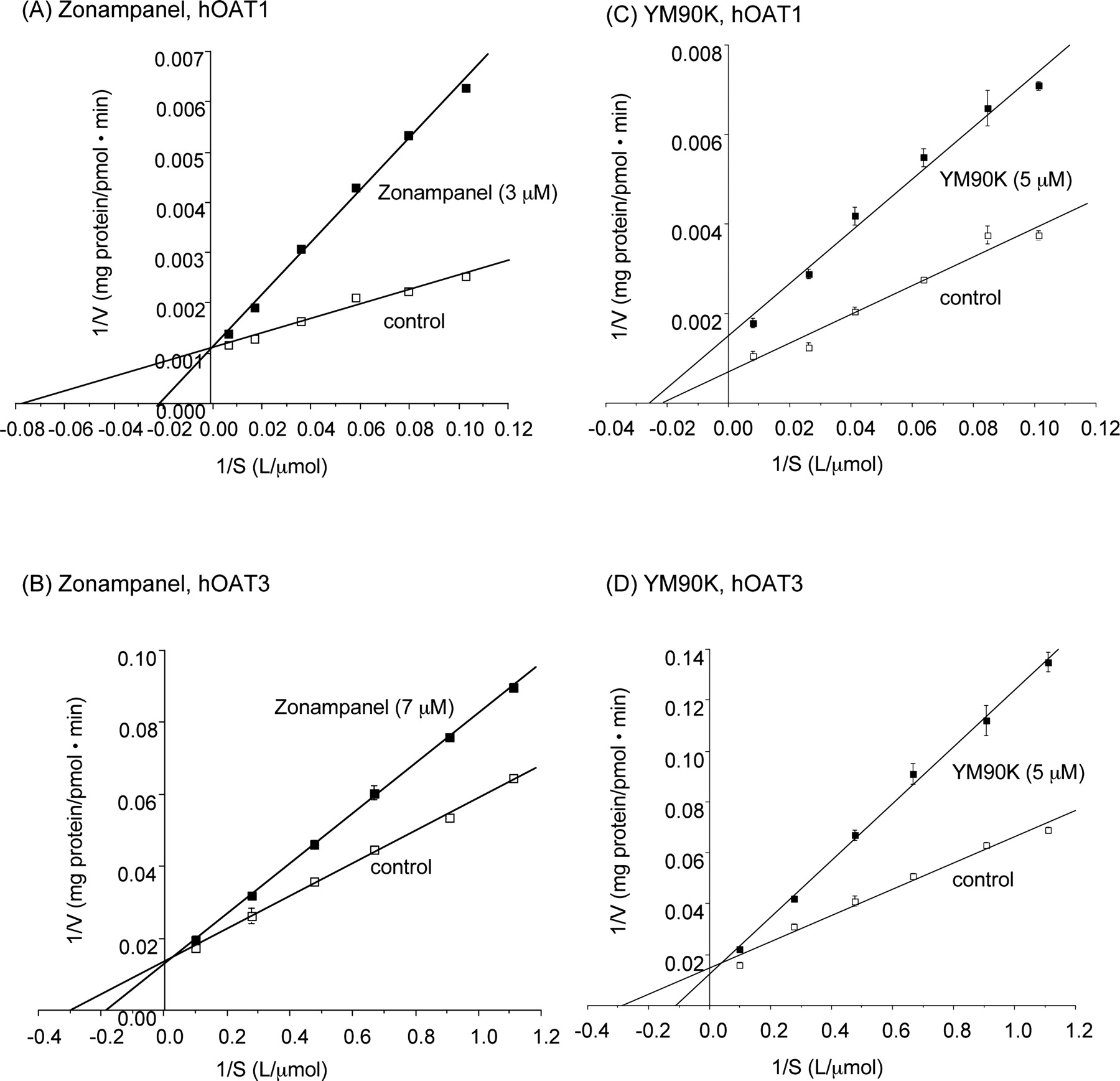
Inhibitory Effects of Zonampanel and YM90K on hOAT- and hOCT-Mediated Organic Ion Uptake. As shown in Tables 1 and 2, zonampanel and YM90K at a high concentration (1 mM zonampanel and 300 μM YM90K) exhibited no inhibitory effect on the uptake of the prototypical substrate by hOCT1 and hOCT2. In contrast, zonampanel (1 mM) significantly inhibited the uptake of prototypical substrates for hOAT1, hOAT2, hOAT3, and hOAT4 (Table 1). Similarly, YM90K (300 μM) also inhibited these hOATs significantly (Table 2). The IC50 or Ki values for zonampanel upon uptake of the prototypical substrates were determined for hOAT1, hOAT3, and hOAT4 in which more than 50% inhibition by 1 mM zonampanel was observed. Zonampanel inhibited the hOAT4-mediated uptake by less than 50% at 300 μM, the maximum concentration tested, indicating that the IC50 value ranged from 300 to 1000 μM. Similarly, the IC50 and Ki values for YM90K upon uptake of the prototypical substrates were determined for hOAT1 and hOAT3 in which more than 50% inhibition by 300 μM YM90K was observed. As shown in Fig. 2, zonampanel inhibited hOAT1- and hOAT3-mediated uptake competitively. YM90K inhibited hOAT3-mediated uptake competitively and hOAT1-mediated uptake noncompetitively.
Inhibitory effect of zonampanel on organic anion uptake by hOATs and on organic cation uptake by hOCTs
Inhibitory effect of YM90K on organic anion uptake by hOATs and on organic cation uptake by hOCTs

Inhibitory effect of zonampanel (A and B) and YM90K (C and D) on hOATs-mediated prototypical substrate uptake. S2 hOAT1 (A and C), S2 hOAT3 (B and D), and the mock were incubated at 37°C for 2 min in D-PBS containing various concentrations of prototypical substrates (PAH for hOAT1, ES for hOAT3 and hOAT4) in the presence or absence of zonampanel or YM90K. Lineweaver-Burk plot analyses were performed from one typical experiment. Each value represents the mean ± S.E. of three determinations.
Zonampanel Uptake Mediated by hOATs and hOCTs. In the time course for [14C]zonampanel uptake in S2 cells stably expressing hOATs and hOCTs, S2 hOAT1, S2 hOAT3, and S2 hOAT4 exhibited a significantly higher [14C]zonampanel uptake than did the mock (Fig. 3). No significant uptake was observed in S2 hOAT2, S2 hOCT1, or S2 hOCT2 (data not shown). Figure 4 illustrates the saturable concentration-dependent uptake of [14C]zonampanel in S2 hOAT1, S2 hOAT3, and S2 hOAT4 together with the Eadie-Hofstee plots. The estimated Km values and the resultant Vmax/Km values of [14C]zonampanel uptake are shown in Table 3.

Time course of zonampanel uptake by hOATs. S2 hOAT1 (A), S2 hOAT3 (B), S2 hOAT4 (C), and the mock were incubated in D-PBS containing 10 μM [14C]zonampanel at 37°C up to 30 min. Each value represents the mean ± S.E. of three determinations from one typical experiment.

Concentration dependence of zonampanel uptake by hOATs. S2 hOAT1 (A), S2 hOAT3 (B), S2 hOAT4 (C), and the mock were incubated in D-PBS containing various concentrations of [14C]zonampanel at 37°C for 5 min. The Eadie-Hofstee plots were analyzed. Each value represents the mean ± S.E. of three determinations from one typical experiment.
Kinetic parameters for zonampanel uptake by hOATs
S2 hOAT1, S2 hOAT3, S2 hOAT4, and the mock were incubated in D-PBS containing various concentrations of [14C]zonampanel at 37°C for 5 min.
Inhibitory Effects of Probenecid and Cimetidine on hOAT-Mediated Zonampanel and Organic Ion Uptake. Probenecid competitively inhibited the uptake of [14C]zonampanel uptake as well as prototypical organic anion uptake via hOAT1, hOAT3, and hOAT4 (Fig. 5; Table 4). Cimetidine competitively inhibited the uptake of prototypical organic anion and [14C]zonampanel via hOAT3 (Fig. 5; Table 4).

Inhibitory effect of probenecid (A, B, and C) and cimetidine (D) on hOAT-mediated zonampanel uptake. S2 hOAT1 (A), S2 hOAT3 (B and D), S2 hOAT4 (C), and the mock were incubated in D-PBS containing various concentrations of [14C]zonampanel in the presence or absence of probenecid or cimetidine at 37°C for 5 min. Lineweaver-Burk plot analyses were performed from one typical experiment. Each value represents the mean ± S.E. of three determinations.
Inhibitory effect of probenecid and cimetidine on prototypical substrate and zonampanel uptake by hOATs
S2 hOAT1, S2 hOAT3, S2 hOAT4, and the mock were incubated at 37°C for 2 min (for the prototypical substrate uptake) and 5 min (for zonampanel uptake) in D-PBS containing various concentrations of prototypical substrates and [14C]zonampanel in the presence or absence of probenecid or cimetidine. Prototypical substrates were PAH for hOAT1, and ES for hOAT3 and hOAT4. Lineweaver-Burk plot analyses were performed for the determination of Ki values.
Discussion
As described previously, renal tubular secretion is considered to play an important role in the renal excretion of zonampanel. This compound possesses an anionic moiety (carboxyl group) and a cationic moiety (imidazole ring) in its molecule (Fig. 1), classifying it as a zwitterionic compound. Numerous zwitterionic compounds are known to interact with both renal organic anion and cation transport systems (Ullrich et al., 1993). These organic anion and cation transporters are known to be the major transporters involved in the renal tubular secretion of numerous compounds. Zonampanel is considered to exist mostly as an anion under physiological conditions (pH ca 7.4). However, it has been reported that the interaction with organic anion and cation transporters is independent of the degree of ionization of the substrates (Ullrich and Rumrich, 1992). Thus, it was expected that zonampanel would be taken up via organic cation transporters as well as organic anion transporters. However, in this study, zonampanel was transported via hOAT1, hOAT3, and hOAT4, but not via organic cation transporters.
Although hOAT1 and hOAT3 have overlapping substrate selectivity, some differences in characteristics have been reported concerning its localization and substrate specificity. hOAT1 exists in the second segment (S2) of the proximal tubule only (Hosoyamada et al., 1999), whereas hOAT3 exists in the first, second, and third segments (S1, S2, and S3) (Cha et al., 2001). hOAT1 mediates sodium-independent uptake of PAH (Km = 9.3 μM) (Hosoyamada et al., 1999) but shows little or no transport of PGE2, methotrexate, or taurocholate, as was found to be the case in oocytes (Cha et al., 2001). In addition, hOAT1, but not hOAT3, exhibits transport properties as an exchanger (Hosoyamada et al., 1999; Cha et al., 2001). hOAT3 mediates the transport of ES (Km = 3.1 μM), methotrexate (Km = 10.9 μM), cimetidine (Km = 57.4 μM), and PAH (Km = 87.2 μM) in a sodium-independent manner. hOAT3 also mediates the transport of dehydroepiandrosterone sulfate, ochratoxin A, PGE2, estradiol glucuronide, taurocholate, glutarate, cAMP, salicylate, and urate (Cha et al., 2001). hOAT4 has been shown to mediate the high affinity transport of dehydroepiandrosterone sulfate (Km = 0.63 μM) and ES (Km = 1.01 μM) in a sodium-independent manner (Cha et al., 2000). hOAT4 also transports various anionic drugs including ochratoxin A (Km = 22.9 μM), PAH, and methotrexate (Babu et al., 2002a,b). However, this transporter exhibits relatively narrow substrate recognition compared with hOAT1 and hOAT3 (Cha et al., 2000; Babu et al., 2002a). This study revealed that zonampanel is transported with high affinity by hOAT1 and hOAT3, with the Km values of 1.4 and 7.7 μM, respectively (Table 3). The Km value of zonampanel for hOAT3 was comparable with those of ES and methotrexate (7.7 versus 3.1-10.9 μM), whereas that of zonampanel for hOAT1 was lower than that of PAH (1.4 versus 9.3 μM).
Zonampanel at 1 mM significantly inhibited the uptake of prototypical substrates via hOATs. The extent of inhibition (percentage of control uptake) was strongest for hOAT1 (1.0%) followed by hOAT3 (14.2%) and hOAT4 (28.7%), the weakest being hOAT2 (58.8%). On the contrary, zonampanel showed no inhibitory effect on hOCT1 and hOCT2. These results indicate that zonampanel has a high affinity for hOAT1, hOAT3, and hOAT4, and correspond well with the theory that zonampanel is transported via these hOATs.
The Km values of zonampanel for hOAT1, hOAT3, and hOAT4 were comparable with the Ki values (as for hOAT4, IC50) of zonampanel for the uptake of prototypical organic anion substrates via the corresponding hOAT [1.4 versus 1.4 μM, 7.7 versus 11.8 μM, and 215 versus 300-1000 μM, respectively (Tables 1 and 3)]. The inhibition pattern by zonampanel was competitive for at least hOAT1 and hOAT3. Probenecid and cimetidine inhibited the uptake of prototypical substrates and zonampanel via hOATs (hOAT1, hOAT3, and hOAT4 for probenecid; hOAT3 for cimetidine) in a competitive manner. In addition, their Ki values for the uptake of prototypical substrates via hOATs were similar to those for the uptake of [14C]zonampanel: 5.9 versus 5.1 μM (hOAT1), 4.6 versus 2.6 μM (hOAT3), and 67.7 versus 57.8 μM (hOAT4) for probenecid, and 43.2 versus 42.9 μM (hOAT3) for cimetidine (Table 4). These findings suggest that the prototypical organic anion substrates (PAH and ES) used in this study as well as zonampanel bind to the same site in each hOAT, and that transport of the prototypical substrates or zonampanel is inhibited by the binding of probenecid or cimetidine at the respective sites. On the other hand, YM90K, a compound structurally analogous to zonampanel, inhibited the uptake of prototypical substrate via hOAT3 competitively, but inhibited the uptake via hOAT1 noncompetitively. These results suggest that YM90K binds to a different site in hOAT1 from that on the prototypical substrate. The structural difference between YM872 and YM90K is a carboxylmethyl group (Fig. 1). This carboxylmethyl group may therefore be involved in the binding specificity for substrates in hOAT1.
Zonampanel monohydrate administered intravenously to humans is excreted into urine mainly in the unchanged form. Fecal excretion is very low, suggesting that biliary excretion is a minor elimination route. This may be due to differences in the expression levels of hOATs and hOCTs in the human liver and kidney. hOAT1 mRNA and hOAT3 mRNA are expressed strongly in the human kidney (Hosoyamada et al., 1999; Cha et al., 2001). Expression of hOAT2 mRNA in human tissues is unknown; however, rat OAT2 mRNA is detected strongly in the liver and weakly in the kidney (Sekine et al., 1998b). hOAT4 mRNA is abundantly expressed in the human kidney and placenta (Cha et al., 2000). Northern blot analysis indicates that hOCT1 mRNA is expressed primarily in the human liver, whereas reverse transcriptase polymerase chain reaction analysis shows that hOCT1 mRNA is also expressed, in lower abundance, in the human kidney and intestine, as well as in other tissues (Gorboulev et al., 1997; Zhang et al., 1997). hOCT2 mRNA is expressed mainly in the human kidney (Gorboulev et al., 1997). The fact that zonampanel monohydrate was not subjected to biliary excretion in humans is consistent with the fact that zonampanel is not recognized by hOAT2 and hOCT1, which are localized in the human liver. Efficient urinary excretion of zonampanel, a good substrate for hOAT1, hOAT3, and hOAT4, is probably due to the expression of these hOATs in the human kidney. In contrast, tetracycline, which is suggested to be transported by hOAT2 as well as hOAT1, hOAT3, and hOAT4 (Babu et al., 2002b), is known to be eliminated not only via glomerular filtration and tubular excretion, but also via the biliary route (Cambers, 2001).
In the human proximal tubule, it has been found that hOAT1 and hOAT3 are localized in the basolateral membrane of the tubular cells (Hosoyamada et al., 1999; Cha et al., 2001), whereas hOAT4 is in the apical membrane (Babu et al., 2002a). This localization of the hOATs suggests that the renal tubular secretion of zonampanel progresses via uptake from the blood into proximal tubular cells by hOAT1 and hOAT3 located in the basolateral membrane, and via the subsequent efflux from the cells into the lumen by hOAT4 located on the apical side. However, the possibility of efflux from renal epithelial cells into the blood via hOAT1 and hOAT3 localized in the basolateral membrane and/or reabsorption from the lumen into renal epithelial cells via hOAT4 (localized on the apical side) cannot be ruled out. Rat OAT1 localized in the basolateral membrane of proximal tubular cells has been shown to act as a homoexchanger, PAH/PAH (Takeda et al., 1999), as well as a heteroexchanger (dicarboxylate/PAH exchange) (Sekine et al., 1998a), and may mediate the bidirectional transport of cephaloridine (Takeda et al., 1999). It has been reported that hOAT4 mediates the reabsorption as well as the efflux of some compounds such as ochratoxin A and tetracycline (Babu et al., 2002a,b). In this study, uptake of zonampanel by the hOATs decreased slowly after reaching its maximum (Fig. 3), suggesting that zonampanel could be effluxed from renal epithelial cells via each hOAT after it is taken up. However, taking into consideration the rapid and almost complete renal excretion of zonampanel in vivo, we suspect that the contribution to the efflux from the cells to the blood and reabsoption from the lumen back to the cells is minor.
In conclusion, these results suggest that the efficient urinary excretion of zonampanel is due to the renal tubular secretion mediated by the organic anion transporters hOAT1, hOAT3, and hOAT4. Zonampanel, cimetidine, probenecid, and the prototypical organic anion substrates used in this study share binding specificity for each hOAT. However, YM90K does not share binding specificity with hOAT1, which suggests that the carboxymethyl group provides the binding specificity for the substrate in hOAT1. In addition, studying S2 cells expressing each hOAT and hOCT serve to further elucidate the molecular mechanism of renal transport for various compounds.
Footnotes
-
ABBREVIATIONS: AMPA, α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid; OAT, organic anion transporter; OCT, organic cation transporter; hOAT, human OAT; hOCT, human OCT; PAH, para-aminohippurate; ES, estrone sulfate; D-PBS, Dulbecco's modified phosphate-buffered saline; TEA, tetraethylammonium; PG, prostaglandin, S1, S2, and S3, the first, second, and third segment of the proximal tubule; YM872, zonampanel monohydrate ([2,3-dioxo-7-(1H-imidazol-1-yl)-6-nitro-1,2,3,4-tetrahydro-1-quinoxalinyl]acetic acid monohydrate); YM90K, 6-(1H-imidazol-1-yl)-7-nitro-2,3(1H,4H)-quinoxalinedione monohydrochloride.
- Received February 17, 2004.
- Accepted June 29, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}