


Abstract
The medium chain triglyceride (MCT) ketogenic diet is a major treatment of drug-resistant epilepsy but is problematic, particularly in adults, because of poor tolerability. Branched derivatives of octanoic acid (OA), a medium chain fat provided in the diet have been suggested as potential new treatments for drug-resistant epilepsy, but the structural basis of this functionality has not been determined. Here we investigate structural variants of branched medium chain fatty acids as new seizure-control treatments. We initially employ a series of methyl-branched OA derivatives, and using the GABAA receptor antagonist pentylenetetrazol to induce seizure-like activity in rat hippocampal slices, we show a strong, branch-point–specific activity that improves upon the related epilepsy treatment valproic acid. Using low magnesium conditions to induce glutamate excitotoxicity in rat primary hippocampal neuronal cultures for the assessment of neuroprotection, we also show a structural dependence identical to that for seizure control, suggesting a related mechanism of action for these compounds in both seizure control and neuroprotection. In contrast, the effect of these compounds on histone deacetylase (HDAC) inhibition, associated with teratogenicity, shows no correlation with therapeutic efficacy. Furthermore, small structural modifications of the starting compounds provide active compounds without HDAC inhibitory effects. Finally, using multiple in vivo seizure models, we identify potent lead candidates for the treatment of epilepsy. This study therefore identifies a novel family of fatty acids, related to the MCT ketogenic diet, that show promise as new treatments for epilepsy control and possibly other MCT ketogenic diet-responding conditions, such as Alzheimer disease.
Introduction
Epilepsy is a common and severe neurologic condition affecting 0.5%–1.0% of the population worldwide (Bell and Sander, 2001). Despite the development of a range of new antiepileptic drugs (Bialer et al., 2010), which, compared with older antiepileptic drugs, have improved tolerability, reduced side effects, and fewer drug-drug interactions (Perucca, 2002), around 30% of people with epilepsy continue to experience seizures (Bialer and White, 2010). The development of new treatments for epilepsy thus addresses a major unmet need.
As an alternative approach to drug treatment, a specialized diet has proven successful in controlling seizures in children with severe, drug-resistant epilepsy (Sills et al., 1986; Vining et al., 1998; Neal et al., 2008, 2009; Kossoff and Rho, 2009). This medium chain triglyceride (MCT) ketogenic diet, first introduced in 1971 (Huttenlocher et al., 1971), was based around a reduction in dietary carbohydrate and an increase in medium chain fatty acid intake, in the form of triglycerides containing 81% octanoic acid (OA) (caprylic acid) and 16% decanoic acid (capric acid) (Sills et al., 1986). This diet increases blood levels of OA and decanoic acid (Huttenlocher et al., 1971; Sills et al., 1986). Decanoic acid can directly inhibit seizure activity, and this may be an important mechanism for the diet’s antiseizure effect (Chang et al., 2013). In contrast, OA has no direct effect on seizures. However, specific branched derivatives of OA provide more potent seizure control than the commonly used antiepileptic drug valproic acid (VPA) (Chang et al., 2012, 2013). Although the diet remains a clear choice for the treatment of drug-resistant epilepsy in children, its use in adults is severely limited due to poor tolerability, hence development of these branched chain fatty acids may provide an alternative to the diet by overcoming its associated side effects and metabolic consequences, especially in adults.
Here we investigate structural aspects of branched medium chain fatty acids in seizure control. We initially determined the seizure control efficacy and neuroprotective activity of a structured series of methyl-branched octanoic acid (MOA) compounds. In these experiments, we used rat entorhinal cortex-hippocampus slices exposed to the GABAA receptor antagonist pentylenetetrazol (PTZ), which is widely used to generate seizures and seizure-like (paroxysmal) activity (Armand et al., 1998). We also used dissociated hippocampal neurons in culture in the presence of low magnesium to generate high-frequency epileptiform activity as an in vitro neuroprotection assay. This model results in glutamate-dependent excitotoxic cell death (Deshpande et al., 2008), which mechanistically is similar to that observed in vivo during status epilepticus (DeLorenzo et al., 2005). We also measured histone deacetylase (HDAC) inhibition; this can have teratogenic effects (Jentink et al., 2010) and has been proposed to explain the teratogenicity of the branched isomer of OA, VPA (2-propylpentanoic acid), a well established epilepsy treatment (Phiel et al., 2001; Eikel et al., 2006). Based upon favorable potency and low HDAC inhibition, we also modified structures to generate more promising compounds. Finally, we analyzed the in vivo efficacy of the most potent compounds, using distinct in vivo models to identify novel chemicals providing strongly enhanced seizure control activity compared with VPA, suggesting a potential new treatment of epilepsy and other MCT ketogenic diet-responsive conditions.
Materials and Methods
Animals
Hippocampal Slice Experiments.
Male Sprague-Dawley rats were kept under controlled environmental conditions (24–25°C; 50%–60% humidity; 12-hour light/dark cycle) with free access to food and tap water. All the experiments were performed in accordance with the 1986 guidelines for animals in scientific procedures.
Maximal Electroshock and 6 Hz Experiments.
Adult male CF No. 1 albino mice (18–25 g maximal electroshock and 26–30 g 6 Hz) and male Sprague-Dawley albino rats (100–150 g) were obtained from Charles River (Portage, MI) and were matched, where possible, for sex, age, and weight (Woolley et al., 1961; Petty and Karler, 1965). Animals were maintained on an adequate diet (Prolab RMH 3000; LabDiet, St. Louis, MO), allowed free access to food and water (Davenport and Davenport, 1948), and allowed time to adjust after transit. All mice were housed in a dedicated facility, in plastic cages with controlled humidity, exchange of air, and controlled lighting (12-hour light/dark cycle). The animals were housed, fed, and handled in a manner consistent with the recommendations in the National Council Publication, Guide for the Care and Use of Laboratory Animals. No insecticides capable of altering hepatic drug metabolism enzymes are used in the animal facility.
Compounds for Analysis
Fatty acids used in this study were VPA (Sigma-Aldrich, St. Louis, MO), OA (Alfa Aesar, Ward Hill, MA), 2-methyl-octanoic acid (2-MOA), 3-methyl-octanoic acid (3-MOA), 6-methyl-octanoic acid (6-MOA), and 7-methyl-octanoic acid (7-MOA) (all from Ukrorgsyntez Ltd., Riga, Latvia), 4-methyl-octanoic acid (4-MOA) and 4-methyl-nonanoic acid (4-MNA) (Alfa Aesar), 4-ethyloctanoic acid (4-EOA) (Chemos GmbH, Regenstauf, Germany), and trans-4-butylcyclohexane carboxylic acid. Compounds were prepared as 1000 times stocks (1 M) in dimethylsulfoxide, except for VPA which was dissolved in distilled water. Stocks were dissolved in artificial cerebrospinal fluid (aCSF) or medium to achieve their final experimental concentrations of 1 mM. All compounds were over 95% purity.
Synthesis of 5-Methyloctanoic Acid
The 5-methyloctanoic acid synthesis procedure was adapted from Levene and Marker (1931). Magnesium turnings (0.209g, 8.55 mmol), iodine (0.002 g, catalytic amount), and anhydrous diethyl ether (2 ml) were stirred at room temperature for 10 minutes under an atmosphere of nitrogen. A solution of 1-bromo-4-methylheptane (1.5g, 7.78 mmol) in anhydrous diethyl ether (3 ml) was added dropwise over 5 minutes. The mixture was heated at reflux for 10 minutes then allowed to cool to room temperature. Dry ice (solid CO2, 1.50 g, 34.08 mmol) was added, and the mixture was allowed to warm to room temperature. Hydrochloric acid (1 M; 10 ml) was carefully added to the reaction mixture, and the crude product was extracted with ethyl acetate (3 × 20 ml). The organic extracts were combined, washed with brine (20 ml), dried (MgSO4), and evaporated under reduced pressure to give clear oil (0.884 g).
The crude product was dissolved in chloroform (20 ml) and extracted with NaOH (2M; 3 × 15 ml). The combined aqueous layers were acidified with concentrated hydrochloric acid to pH 2.0, and extracted from the aqueous layer with chloroform (3 × 20 ml). The organic extracts were combined, washed with brine (20 ml), dried (MgSO4), and evaporated under reduced pressure to give 5-MOA (0.554 g, 41%) as a colorless oil. High-resolution mass spectrometry–electrospray (m/z) found 181.1203, calculated for [C9H18NaO2]+ 181.1199) 1H-NMR (500 MHz, CDCl3) δ 10.31 (s, 1H), 2.33 (t, J 7.6 Hz, 2H), 1.73–1.54 (m, 2H), 1.48–1.38 (m, 1H), 1.37–1.20 (m, 4H), 1.20–1.05 (m, 2H), 0.92–0.84 (m, 6H). 13C-NMR (126 MHz, CDCl3) δ 180.7 (CO), 39.3 (CH2), 36.5 (CH2), 34.6 (CH2), 32.4 (CH), 22.4 (CH2), 20.2 (CH2), 19.6 (CH3), 14.3 (CH3).
In Vitro Electrophysiology
The preparation of entorhinal cortex-hippocampus slices and electrophysiologic recording in CA1 were described previously elsewhere (Armand et al., 1998; Chang and Walker, 2011). In brief, male rats (50–150 g) were decapitated after being killed by intraperitoneal injection with an overdose of pentobarbitone (500 mg/kg). The brain was removed and preserved in oxygenated ice-cold sucrose solution in mM: NaCl 87, KCl 2.5, MgCl2 7, CaCl2 0.5, NaH2PO4 1.25, NaHCO3 26.2 sucrose 75, glucose 3. Transverse slices (350 µm) were prepared with a Vibratome (1500; Intracel Ltd., Royston, UK) and were then stored in an interface chamber containing aCSF (in mM): NaCl 119, KCl 2.5, MgSO4 1.3, CaCl2 2.5, NaH2PO4 1, NaHCO3 26.2, glucose 16.6. The slices were stored for over 1 hour. During the experiment, the slices were transferred from the interface chamber into a submerged recording chamber and continuously perfused with prewarmed (about 36°C) oxygenated (95% O2, 5% CO2) aCSF.
A field potential recording was made by placing a glass microelectrode filled with aCSF solution in stratum radiatum of CA1. A bipolar stimulating electrode was positioned in the Schaffer collateral/commissural fiber pathway in stratum radiatum to confirm slice viability. PTZ (2 mM) was added to the perfusate, and [K+] was increased to 6 mM to induce epileptiform activity. PTZ-induced epileptiform discharges, population spikes consisting of positive field potentials, appeared 10–30 minutes after application of PTZ plus the increase of [K+]. Compounds were applied once the frequency and amplitude of the epileptiform discharges were stable over a period of 10 minutes. Anticonvulsant effects were evaluated by measuring the variation of frequency of the discharges every minute.
In Vitro Neuroprotection Assays
Primary Hippocampal Neuron Cell Culture.
Primary hippocampal neuron cell culture was prepared from rat pups (postnatal day 0–1). After the brains had been quickly removed and the hippocampi isolated, the hippocampi were submerged in ice-cold HEPES-buffered Hanks’ balanced salt solution (Invitrogen/Life Technologies, Carlsbad, CA). The tissue was then treated with 1% trypsin for 10 minutes at 37°C to dissociate cells. After removal of residual trypsin, the tissue was triturated and plated at a density of 2 × 105 cells/well onto a glial-supported layer that had been previously plated onto poly-l-lysine–coated cell culture coverslips (13 mm). Cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2 and 95% air, and fed once weekly with NeuroBasal A medium supplemented with B-27 (Invitrogen/Life Technologies) and Gibco GlutaMAX Supplement (Gibco/Life Technologies, Grand Island, NY).
Fourteen days after cell culture preparation, cultures were used for experimentation. The maintenance medium was replaced with HEPES-buffered aCSF with or without MgCl2 (2 mM), containing 125 mM NaCl, 2.5 mM KCl, 25 mM HEPES, 2 mM CaCl2, 30 mM glucose, pH 7.4, and osmolarity adjusted to 310 mOsm with sucrose. Low magnesium treatment with/without OA derivate was performed with aCSF without added MgCl2, whereas sham controls were treated with aCSF containing MgCl2.
Lactate Dehydrogenase Activity Assay.
Four hours after cells were exposed to a low magnesium environment in the absence or presence of compounds, neuronal cell injury was quantitatively assessed by the measurement of lactate dehydrogenase (LDH) released into the extracellular fluid from damaged cells using a commercial kit (TOX7; Sigma-Aldrich). Release of LDH into the extracellular fluid is an indication of compromised membrane integrity (Koh and Choi, 1987), thus providing a means to monitor cell toxicity without damaging the cells in culture.
The assay was conducted according to the manufacturer’s protocol. In brief, duplicate samples of media (50 µl) were transferred to wells of a clear 96-well flat-bottom microtiter plate (Corning Costar, Corning, NY), and 20 µl of LDH mixture containing equal amounts of LDH cofactor, substrate, and dye solutions were added (all supplied in the TOX7 kit). The plates were incubated in the dark for 30 minutes at room temperature. LDH release was assayed by absorbance change (490 nm) using a microplate reader, with results normalized to sham control. Data are presented as mean percentage of cell death ± S.E.M. (compared with sham control).
Viability Staining.
Four hours after cells were exposed to the low magnesium environment in the absence or presence of compounds, cell death was visualized by staining with 10 µM propidium iodide (PI) (excitation at 488 nm; emission at 550 nm) (Sigma-Aldrich) and 10 µM Hoechst 33342 (excitation at 530 nm; emission at 620 nm) (Sigma-Aldrich). PI is a cell-impermeant fluorescent biomarker that will fluoresce upon binding nuclear chromatin; thus, fluorescence provides an indication of loss of cell membrane integrity as a marker for cell death. Hoechst 33342 is a cell-permeant nuclei stain that provides a total cell count. After treatment, cells were stained for 30 minutes at room temperature, washed to remove unbound dye, and cell death and total cell count were visualized and quantified using ImageJ (http://imagej.nih.gov/ij/).
HDAC Activity Assay.
HDAC activity was analyzed by using an in vitro commercial assay kit, the HDAC fluorescent activity assay/drug discovery kit HDAC activity (Biomol, Plymouth Meeting, PA). The assay was conducted at room temperature according to the manufacturer’s protocol, using Trichostatin A (a HDAC inhibitor; at 1 μM concentration) as a positive control. Data were derived from at least triplicate experiments, with each experiment comprising at least duplicate measurements (n = 6) normalized to controls (without VPA or fatty acids).
In Vivo Seizure Model
The 6 Hz “Psychomotor” Seizure Test.
Adult male CF1 mice (18–25 g) were pretreated with each compound at 100–150 mg/kg i.p. After pretreatment, each mouse was given a drop of 0.5% tetracaine hydrochloride to each eye, followed by low-frequency (6 Hz) stimulation (32 mA) for 3 seconds delivered through corneal electrodes. Animals were manually restrained during stimulation and released immediately after the stimulation and observed for the presence or absence of seizure activity at five time points (0.25, 0.5, 1, 2, and 4 hours). Typically, the 6 Hz stimulation results in a seizure characterized by a minimal clonic phase that is followed by stereotyped, automatistic behaviors, including twitching of the vibrissae, and Straub tail. Animals not displaying such behaviors were considered protected.
Maximal Electroshock Test.
The maximal electroshock (MES) test is a model for generalized tonic-clonic seizures (Loscher, 2011) that provides an indication of a compound’s ability to prevent seizure spread when all neuronal circuits in the brain are maximally active. Animals are pretreated with compounds (100–125 mg/kg) by intraperitoneal injection. Seizure activity was induced by delivery of 60 Hz of alternating current (50 mA in mice, 150 in rats) for 0.2 seconds through corneal electrodes that had been primed with an electrolyte solution containing an anesthetic agent (0.5% tetracaine HCl). An animal is considered “protected” from MES-induced seizures upon abolition of the hind limb tonic extensor component of the seizure (Swinyard et al., 1995; White et al., 1995, 2007).
Subcutaneous Metrazol Seizure Threshold Test.
Subcutaneous injection of pentylenetetrazol produces clonic seizures in animals. The subcutaneous metrazol seizure threshold test (scMET) test allows detection of test compound efficacy in raising the seizure threshold, thus demonstrating protection from clonic seizure induction. Animals were pretreated with each compound (100 mg/kg) in a similar manner to the MES test. Seizure activity was induced by administration of metrazol (CD97: 85 mg/kg mice; 70 mg/kg rats) into skin in the midline of the neck. Animals were observed for the following 30 minutes for the presence or absence of a seizure. Animals were considered “protected” if they did not exhibit an episode of clonic spasms of the fore and/or hind limbs, jaws, or vibrissae (3–5 seconds).
Corneal Kindled Mouse Model.
This model involves the use of corneal kindled mouse model for prediction of efficacy in human partial seizures (Rowley and White, 2010). Adult male CF1 mice (18–25 g) mice were stimulated through corneal electrodes (3 mA, 60 Hz, 3 seconds) after administration of 0.5% tetracaine hydrochloride to each eye. This procedure was performed twice daily for an average of 12 days. Animals were considered kindled when they displayed five consecutive stage 5 seizures according to the Racine scale (Racine, 1972): stage 1, facial automatisms; stage 2: head nodding and more serve facial and mouth movements (jaw opening); stage 3: forelimb clonus with a lordotic posture; stage 4: bilateral forelimb clonus continuing along with rearing; stage 5: some rats fall to one side first and then show evidence of forelimb clonus. After 5 to 7 days of stimulation-free period, once the mice were fully kindled, the test compounds were administrated (100 mg/kg i.p.). Mice in each group were then tested at various time points (0.25, 0.5, 1, 2, 4 hours) after drug dosing. Mice displaying a seizure score <3 are considered protected.
Statistical Analyses
For all data, the results provided are presented as mean ± S.E.M. Statistical comparisons were performed by using the one-way analysis of variance followed by Tukey for post hoc analysis using GraphPad (GraphPad Software, San Diego, CA).
Results
In Vitro Seizure Control Activity of Methyl-Branched Octanoic Acid Derivatives in the PTZ/Hippocampal Model Is Structurally Specific.
The discovery of a direct role for branched derivatives of OA, rather than the inactive unbranched compound, in multiple in vitro seizure models and an in vivo model of status epilepticus (Chang et al., 2012, 2013) suggests that improved activity may be found by examining the range of branching structures possible for these fatty acids. We thus initially examined the seizure control activity of a series of branched OA derivatives containing a methyl group side chain (Fig. 1A) at a single concentration (1 mM) as an early indication of comparative activity. For these experiments, we induced seizure-like activity in rat hippocampal slices by the application of PTZ and monitored the frequency of burst activity in the CA1 region. Consistent with our previous study (Chang et al., 2013), OA has no effect on the frequency of burst discharges 20–40 minutes after application of PTZ (98.4% ± 7.2% of baseline, n = 4) (Fig. 1B).

MOA derivatives show strong structure-specific potency in an in vitro model of epileptiform activity control. (A) The structure of the straight chain, eight carbon, OA, and derivatives branched at the second carbon to the seventh carbon. (B) Summary of the change in the frequency of PTZ-induced burst discharges in area CA1 recorded from the stratum pyramidal by application of OA and derivatives (1 mM). The frequency of epileptiform activity is plotted against time. (C) Comparison of the mean frequency of PTZ-induced burst discharges, averaged from 20–40 minutes after compound addition (data shown as mean ± S.E.M.). ***P < 0.001, statistically significant difference at compared with OA. Data are provided for all compounds tested at 1 mM from at least four to five independent repeats.
In contrast, methyl-group substitution on the third to the seventh carbon of the octanoic backbone resulted in compounds that significantly reduced discharge frequency, and the position of the branching point related to activity (Fig. 1B). Inhibition of seizure-like activity increased through substitution of the methyl group on the second carbon (2-MOA: 78.1% ± 3.6% of baseline, n = 5), to the third (3-MOA: 62.3% ± 2.6% of baseline, n = 5) and fourth carbon (4-MOA: 48.6% ± 2.7% of baseline, n = 5). Inhibition of seizure-like activity was greatest with the methyl branch chain located at the fifth position (5-MOA; 8.2% ± 5.4% baseline, n = 4). Inhibition of seizure-like activity then decreased compared with that of 5-MOA when the methyl branch chain was substituted on the sixth (6-MOA: 25.4% ± 7.9% of baseline, n = 5) and then further reduced on the seventh position (7-MOA: 39.2% ± 4.0% of baseline) (Fig. 1B).
These potent compounds show enhanced seizure-activity control compared with the established epilepsy treatment VPA, application of which results in a small but significant reduction in discharge frequency in this model (77.1% ± 2.0% of baseline) (Chang et al., 2013). These results suggest a strong structure-relative effect of branched OA derivatives directly on controlling seizures.
In Vitro Neuroprotective Activity of Methyl-Branched Octanoic Acid Derivatives in the Low Magnesium Model Is Structurally Specific.
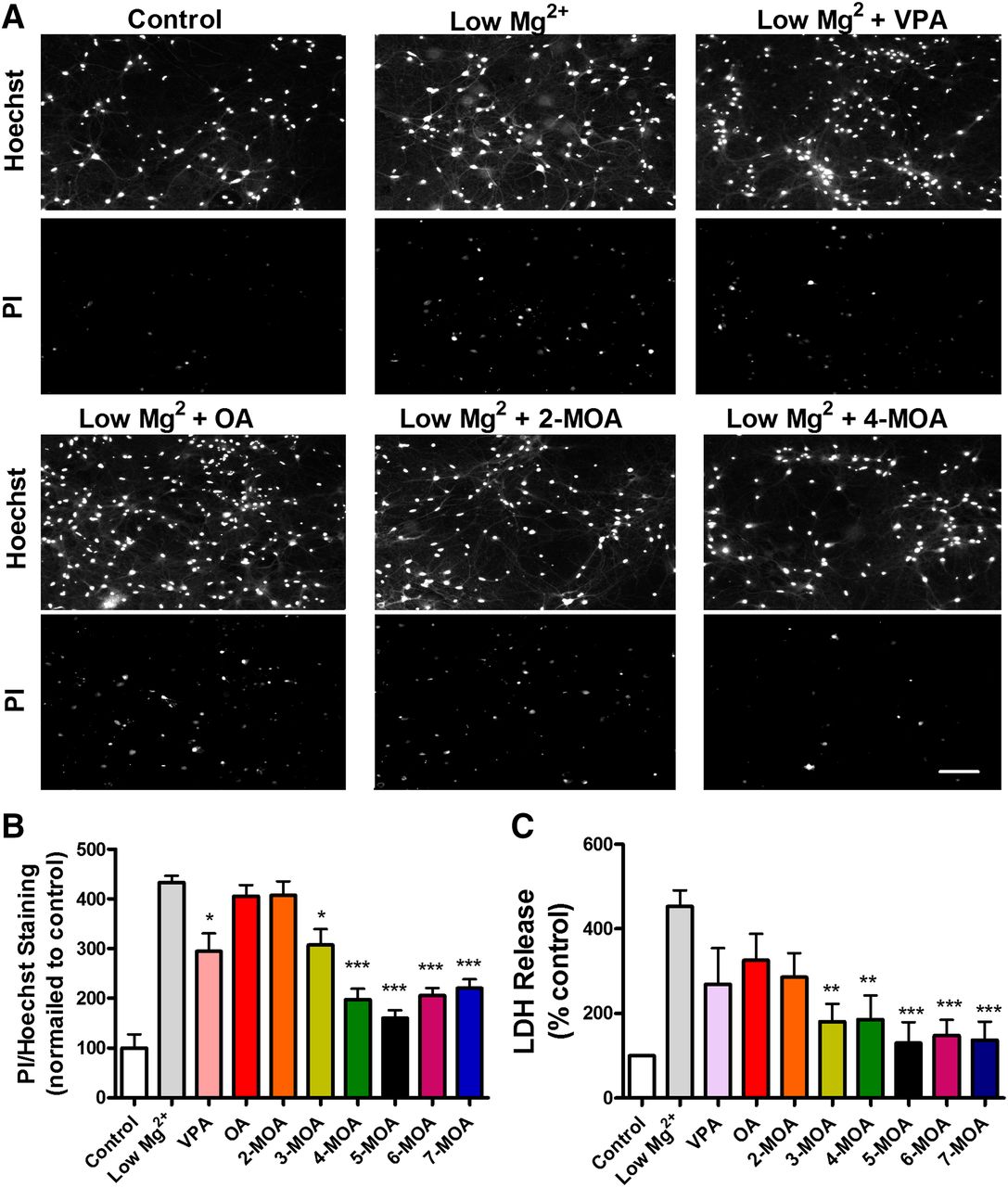
The development of new antiepileptic treatments has recently included the analysis of neuroprotective properties of these compounds, whereby novel treatments can reduce neuronal cell damage or death resulting from seizure activity. To investigate this effect in the series of branched-chain OA compounds, we used a model of glutamate receptor–dependent excitotoxic cell death, which is mechanistically similar to that observed in vivo during status epilepticus (DeLorenzo et al., 2005). This consisted of exposure of hippocampal neurons in culture to low magnesium (Deshpande et al., 2008). Cell death in this model occurs in a time-dependent manner, increasing from 30% to 100% with the increase of low magnesium exposure duration from 2 to 12 hours (Deshpande et al., 2008). We thus induced epileptiform activity by exposure of neurons in culture to low magnesium conditions for 4 hours and monitored cell death by recording uptake of the PI dye as a marker for dead cells (Fig. 2, A and B) and the release of LDH into the media as a biomarker for cellular cytotoxicity and cytolysis (Fig. 2C).

Branched methyl-octanoic acid derivatives show strong structure-specific potency in an in vitro model of neuroprotection. (A) Primary rat hippocampal neurons, maintained under low Mg2+ conditions for 4 hours to trigger status epilepticus-induced cell damage, were treated with OA and derivatives, shown for control (without low Mg2+ treatment), after low Mg2+ treatment, and after treatment in the presence of OA, 2-MOA, 4-MOA, and 5-MOA. Cells were costained with the nuclear dye Hoechst 33342 to visualize the total neuronal population, and the exclusion dye PI to visualize neuronal death. Scale bar = 20 µm. (B) Quantification of change in ratio of PI staining to total cell number after treatment with the series of branched methyl-octanoic acid derivatives. (C) Comparison of the LDH release 4 hours after induction of status epilepticus in low Mg2+ model in hippocampal neurons in culture. All data shown as mean ± S.E.M. *P < 0.05; **P < 0.01; ***P < 0.001, all statistically significant difference compared with low Mg2+ alone group. Data are provided for all compounds tested at 1 mM from at least five independent experiments.
In these experiments, exposure to low magnesium conditions significantly increased neuronal cell death, measured both by PI uptake (433% ± 30% of control) and LDH release (453% ± 38% of control). Application of OA provided no significant protection against neuronal injury in this model (PI uptake: 406% ± 45% of control, LDH release: 326% ± 62% of control). Similarly, addition of a methyl group to the second carbon, 2-MOA, did not provide significant protection measured either by PI uptake or by LDH release (407% ± 45% of control; 286% ± 56% of control, respectively) (Fig. 2, A–C). In contrast, 3-MOA to 7-MOA all resulted in a significant attenuation of PI uptake and LDH release, with 4-MOA reducing PI uptake and LDH release to 197% ± 11% and 186% ± 57% of control, respectively (Fig. 2, A–C). In comparison, VPA had an intermediate neuroprotective effect (PI uptake: 295% ± 12% of control; LDH release: 269% ± 32% of control).
Together, these results indicate that the branching pattern of MOA derivatives strongly regulates neuroprotective activity, similar to the structure-function relationship obtained from the in vitro PTZ-induced seizure model. These data also suggest that compounds containing branch points on the fourth to seventh carbon of the OA backbone have greater neuroprotective potential than VPA.
Physiochemical Properties of Methyl-Branched Octanoic Acid Derivatives Do Not Correlate to Efficacy.
To examine a potential physiochemical basis of seizure control activity for these compounds, lipophilicity (LogP) and acidity (pKa) values were estimated for each compound. Data were derived using literature sources and predictive software. No gross trends were observed for either physical parameter over the OA series (Supplemental Fig. 1). This suggests that it was unlikely that seizure control activity and neuroprotective efficacy of these medium chain fatty acids were due to purely physiochemical characteristics, but rather to biologic function.
Histone Deacetylase Inhibitory Activity of Methyl-Branched Octanoic Acid Derivatives Is Structurally Specific.
To investigate potential HDAC inhibitory potential of this series of branched OA derivatives, compounds were tested using an in vitro HDAC inhibition assay. In this assay, human HeLa cell nuclear extract was used as the source of HDAC activity. VPA showed a relatively strong inhibition of HDAC activity (IC50: 2.1 ± 0.1 mM; Fig. 3, A and B). In this assay, OA showed strong inhibitory activity (IC50: 6.4 ± 0.3 mM), but the addition of a methyl group to the second carbon to give 2-MOA caused a significant reduction in HDAC inhibitory activity (IC50, 13.6 ± 2.8 mM). Addition of a methyl group to the third carbon to give 3-MOA resulted in a similar small decrease in HDAC inhibition compared with OA (IC50, 8.7 ± 0.9 mM). Branched derivatives based upon the fourth to seventh carbon exhibited a highly significant reduction in HDAC inhibitory potency compared with OA (IC50, 4-MOA: 30.5 ± 1.4 mM; 5-MOA: 20.8 ± 0.8 mM; 6-MOA; 20.2 ± 2.8 mM; 7-MOA: 17.9 ± 1.2 mM) (Fig. 3, A and B). These data suggest that all branched chain OA compounds examined here are less likely than VPA to be teratogenic, and compounds branched on the fourth, fifth, sixth, and seventh carbon are unlikely to have any significant HDAC inhibitory effect at concentrations likely to be produced in vivo.

Branched methyl-octanoic acid derivatives show structure-specific potency in an in vitro human HDAC enzyme activity assay. (A) Quantification of HDAC inhibition assay employing human nuclear enzyme extracts (from HeLa cells) as the source of HDAC activity shown as fitted dose-response curves (compared with VPA). (B) Comparison of the IC50 of HDAC inhibition with different treatments, showing mean ± S.E.M. for three independent measurements at six concentrations (0.1–10 mM). **P < 0.01, statistically significant difference compared with VPA; ***P < 0.001, statistically significant difference compared with VPA.
Related Derivatives Show No HDAC Inhibition and Strong Activity in the PTZ/Hippocampal Model.
Our previous studies indicated that compounds containing longer branch chains and extended backbones also have strong antiseizure activity (Chang et al., 2012, 2013). To expand the chemical space examined in this study, we tested three compounds related to the OA series (Fig. 4A) using both the HDAC inhibitory assays (Fig. 4B) and the PTZ-induced hippocampal slice seizure model experiments (Fig. 4C) at the same concentration used for the previous compounds (1 mM).

Specific modified methyl-octanoic acid derivatives show no activity in an in vitro human HDAC enzyme activity assay and potency in a seizure control in vitro model of epileptiform activity. (A) The structure of various fatty acids with branch chain on the fourth carbon, 4-EOA, 4-MNA, and related 4-BCCA. (B) Quantification of HDAC inhibition assay employing human nuclear enzyme extracts (from HeLa cells) as the source of HDAC activity shown as fitted dose-response curves. (C) Summary of the change in the frequency of PTZ-induced burst discharges in area CA1 recorded from the stratum pyramidal by application of 4-EOA (n = 8), 4-MNA (n = 6), and 4-BCCA (n = 5). **P < 0.01, statistically significant difference compared with VPA; ***P < 0.001, statistically significant difference compared with VPA.
The addition of a 4-ethyl group to OA to produce 4-EOA rather than 4-MOA led to no HDAC inhibition but enhanced epileptiform activity control in the PTZ-induced hippocampal slice model (10.5% ± 4.0% of baseline, n = 8; Fig. 4, A–C) over the related methyl-branched compound. Elongation of the backbone length from the same starting compound, to form 4-MNA, also led to no HDAC inhibition and enhanced epileptiform activity control (2.0% ± 2.9% of baseline, n = 6; Fig. 4, A–C) over the parent compound.
Finally, analysis of a cyclic derivative, where a butyl group is attached to a cyclohexane carboxylic acid (4-butylcyclohexane carboxylic acid; 4-BCCA) again provided no HDAC inhibition with very strong epileptiform activity control (1.7% ± 1.5% of baseline, n = 5; Fig. 4, A–C). These compounds also did not show significant changes to physiochemical properties (Supplemental Fig. 1). The enhanced activity of these compounds over VPA suggests further possible lead candidates for better epilepsy treatments.
In Vivo Seizure Control Activity of Derivatives Suggests Structurally Specific Lead Candidates.
The advantage of in vitro models is that efficacy does not depend on the pharmacokinetics and blood-brain barrier penetration of the compounds. However, these in vitro models of epileptiform activity may not necessarily predict in vivo efficacy both because of pharmacokinetics and also because these reduced models have only a small portion of the brain circuitry intact. In the development of new antiepileptics, in vivo models are therefore used both to better predict efficacy and to provide evidence for physiochemical properties, such as in vivo stability and blood-brain barrier penetrance.
We examined the efficacy of the most active compounds in multiple in vivo epilepsy models in collaboration with the Anticonvulsant Screening Program at the US National Institutes of Health National Institute of Neurological Disorders and Stroke. In these experiments, multiple doses were used to estimate ED50 levels and give comparative potencies compared with VPA. We first employed a low frequency (6 Hz), long duration (3 seconds) corneal stimulation model because numerous second-generation antiepileptic drugs show poor seizure control in this assay (Loscher, 2011). In this model, compounds are preadministered to mice via intraperitoneal injection, and mice are challenged with sufficient current to elicit a psychomotor response (32 mA for 3 seconds) (Toman et al., 1952). The model causes forelimb clonus and then the automatistic behavior characteristic of limbic epilepsy in humans.
Our initial analysis of methyl-branched derivatives with the highest activity and lowest risk of HDAC inhibition, 4-MOA and 5-MOA, and the related 4-MNA showed limited seizure control (Table 1). However, analysis of 4-EOA and 4-BCCA showed an ED50 of 110 and 81 mg/kg, respectively, indicating that both are more potent than VPA, which has an ED50 of 263 mg/kg in this model (Barton et al., 2001).
In vivo seizure control data for active compounds
Summary of in vivo seizure control in multiple models.
Compounds showing potential efficacy in the 6 Hz model were then tested in three further in vivo seizure models. The MES seizure model is one of the primary preclinical models used in epilepsy drug development (Barton et al., 2001). Using this model, 4-EOA and 4-BCCA both had EC50 values of 100 mg/kg (Table 1). The scMET model, thought to determine the seizure threshold for clonic seizures, showed 4-EOA and 4-BCCA had EC50 values of 100 and ∼150 mg/kg, respectively, whereas VPA had an equivalent effect at 191 mg/kg (Rowley and White, 2010). Finally, in the corneal kindled mouse model, which provides an in vivo model for temporal lobe epilepsy (White, 2003), 4-EOA and 4-BCCA had EC50 values of 71 and 44 mg/kg, respectively, compared with VPA which showed equivalent activity at 174 mg/kg. These data strongly suggest enhanced activity of 4-EOA and 4-BCCA over VPA in a range of distinct seizure models.
Discussion
The MCT ketogenic diet is a widely used and effective treatment of drug-resistant epilepsy in children, but it is rarely employed for adult treatment due to poor tolerability. Our recent discovery of the antiseizure efficacy of branched derivatives of OA, the major constituent of the diet, may provide lead candidates for pharmacologic treatment of epilepsy based around this family of compounds. This led us to investigate these structures in a more comprehensive manner.
In this study, we initially used a single-dose PTZ model to identify differences in activity of a structured series of MOA derivatives to provide a first indication of branching effects. PTZ is widely used as an epileptogenic agent. In the hippocampal slice model used here, PTZ triggers spontaneous interictal-like repetitive burst discharges (Hori and Katsuda, 1974; Rostampour et al., 2002), which are similar to the seizure activity generated by increasing extracellular K+ concentrations in the cerebral cortex in vivo (Futamachi et al., 1974; Moody et al., 1974; Heinemann et al., 1977). The induction of PTZ discharges in the in vitro preparation offers the advantage of a simplified model for studying the pharmacology of antiepileptic drugs (Piredda et al., 1985).
We found a strong structure-related change in the control of PTZ-induced seizure-like activity in a rat hippocampal slice model, with highly significant inhibition of seizure activity by compounds branching at carbons 3–7. Our previous studies showed that VPA, the currently used branched chain fatty acid with a branch point on the second carbon, has a much smaller effect in this model (seizure activity is reduced to 77.1% ± 2.0% of baseline at 1 mM) (Chang et al., 2013). These results suggest that alternative branch points on medium chain fatty acids may provide enhanced potency over VPA in seizure control.
It is interesting to note that a recent study found that orally dosed OA increased the threshold of seizure induction in a range of seizure models (Wlaz et al., 2012). However, our data indicate no direct role of OA in seizure control, suggesting that metabolites and derivatives of OA may provide this therapeutic effect.
Neuroprotective activity may also prove useful in reducing the development of epilepsy as well as cognitive and neurologic decline after a prolonged seizure or brain insult (Dam, 1980; De Lanerolle et al., 1989; Barton et al., 2001). Similar to the seizure control data, we found a strong structural dependence of medium chain fatty acid–mediated neuroprotection in a low magnesium model. Compounds branched from the fourth to seventh carbon show greatest activity in protection against seizure-related cell death (Koh and Choi, 1987). This protective activity is greater than that shown by VPA, suggesting enhanced therapeutic efficacy for these compounds. A link between activity in the in vitro PTZ and neuroprotective models may relate to the suppression of seizure activity or decreased ion fluxes as opposed to a primary neuroprotective effect. Further studies will be necessary to determine whether these active compounds function through a common mechanism for both seizure control and neuroprotection.
Direct inhibition of HDAC by VPA (Göttlicher et al., 2001; Gurvich et al., 2004) is thought to give rise to teratogenicity (Koren et al., 2006; Jentink et al., 2010), causing a variety of major and minor malformations, including neural tube defects, cleft lip and palate, cardiovascular abnormalities, genitourinary defects, developmental delay, endocrinological disorders, limb defects, and autism (Alsdorf and Wyszynski, 2005). This biochemical activity provides a crucial side-effect profile to consider in the development of carboxylic acid–based antiepileptic drugs. Structural requirements for this activity for VPA congeners suggest a critical role for branching of the parent compound on the second carbon of the fatty acid backbone (Bialer and Yagen, 2007; Bialer et al., 2010).
Our data indicate that all compounds in our OA series show reduced HDAC inhibition, with branching at the fourth carbon providing the least activity, with around a 15-fold increase in IC50 values. This suggests that these compounds, especially 4-MOA (IC50 30.5 ± 1.4 mM), are unlikely to significantly inhibit HDAC activity in vivo at concentrations found in plasma for VPA (0.4–0.7 mM (Diagnostic and Statistical Manual IV; American Psychiatric Association, 2000) or OA (306 μM; Haidukewych et al., 1982). These data also support the enhanced safety of these novel branched fatty acids in seizure control, showing a reduced potential for HDAC inhibitory activity associated with teratogenicity.
To provide lead candidates for seizure control treatment, we next focused on identifying new structures with low HDAC inhibitory activity (based on 4-MOA) to develop compounds showing activity equivalent or greater than that shown for 5-MOA. Addition of a longer branch chain (ethyl-branch derivatives), lengthening the backbone (nonanoic acid), or incorporation of a ring structure all provided equivalent (4-EOA) or enhanced (4-MNA, 4-BBCA) seizure control in the PTZ-induced hippocampal model compared with 5-MOA and did not show HDAC inhibitory activity. These compounds thus provide exciting new leads for further screening.
We next translated our in vitro results to multiple in vivo models (using a range of doses to determine potency) because these models are more relevant to human seizures and epilepsy. This is particularly important because the efficacy of compounds in vivo is not necessarily predicted by in vitro models because of the complex in vivo metabolic kinetics. Furthermore, compounds tested in vitro may not reach an effective brain concentration due to low blood-brain barrier permeability. We first employed the 6 Hz electric shock model (Barton et al., 2001), in which VPA, levetiracetam, phenytoin, lamotrigine, lacosamide, carisbamate, and retigabine have shown efficacy (Bialer et al., 2010). The methyl-branched derivatives in this model show low potency in this model, which may mirror that shown for other established epilepsy treatments such as topiramate (Rowley and White, 2010), or may be due to physiologic restrictions, such as half-life or blood-brain barrier permeability; further studies will be necessary to address these questions.
However, the enhanced potency of 4-EOA and 4-BCCA over VPA in this model provides an encouraging step forward in developing new treatments in this chemical space. Because these compounds also show significantly decreased IC50 values compared with VPA in the widely used MES and scMET screening models and, importantly, in the corneal kindling model of temporal lobe epilepsy (Lothman and Williamson, 1994), these compounds have significant potential as antiseizure drugs. This contrasts with orally administered straight-chain OA (Wlaz et al., 2012), which increased the PTZ dose necessary to induce myoclonic twitch and clonic convulsions without an effect on tonic convulsions and increased the current intensity needed to induce psychomotor seizures in the 6 Hz model, but had no effect in the MES model. Thus, our initial in vitro epilepsy model suggested novel branching structures that show efficacy in one (PTZ-induced hippocampal slice model), but only two compounds (4-EOA and 4-BCCA) showed enhanced potency over VPA in multiple in vivo models, thus providing potential new structures for further analysis.
The discovery of this new chemical space for compounds with antiseizure activity, comprising branched and cyclic derivatives of a medium chain fatty acid in the MCT ketogenic diet, has implications for other therapeutic areas. The MCT diet also provides a positive effect in decreasing brain excitability in young animals (De Almeida Rabello et al., 2008), as well as playing a neuroprotective role in traumatic brain injury and stroke (Gasior et al., 2006). Finally, the diet also has potential for other neurologic disorders characterized by neuronal cell death (Stafstrom and Rho, 2012), such as Alzheimer disease (Reger et al., 2004; Henderson et al., 2009) and Parkinson disease (VanItallie et al., 2005). Future research into the use of these compounds for these conditions may provide important advances in therapy development.
In summary, a series of novel MOA derivatives were evaluated for seizure control initially using an in vitro model of seizure-like activity. Unlike the widely used short chain fatty acids, VPA, and related compounds branched on the second carbon, the potency of OA derivatives was enhanced with branching from the middle of the carboxylic acid backbone. A similar structure-activity relationship was also seen for neuroprotection against neuronal death induced by low magnesium levels. Small changes in structure to these active compounds to lengthen side chain and inclusion of a ring structure in the backbone further enhanced activity. A range of in vivo seizure control assays showed improved potency over VPA for these compounds. Given the remarkable value of the MCT diet in the treatment of epilepsies (Huttenlocher et al., 1971; Sills et al., 1986; Brandt et al., 2003; Neal et al., 2008, 2009), and its shortcomings in terms of adverse effects, there is interest in the search for derivatives of medium chain fatty acids found in the MCT diet with improved pharmacokinetic or safety profiles. We demonstrate that structurally specific medium chain fatty acids have potent antiseizure and neuroprotective properties, with clear clinical implications in providing a new chemical space for the design of more potent and safer treatments for epilepsy.
Acknowledgments
The authors gratefully acknowledge the National Institutes of Health National Institute of Neurological Disorders and Stroke Anticonvulsant Screening Program (ASP) for kindly performing the pharmacologic studies via the ASP’s contract site at the University of Utah.
Authorship Contributions
Participated in research design: Chang, Zuckermann, Walker, Williams.
Conducted experiments: Chang, Zuckermann, Williams, Cano-Jaimez.
Contributed new reagents or analytic tools: Close, Spencer.
Performed data analysis: Chang, Zuckermann, McEvoy.
Wrote or contributed to the writing of the manuscript: Chang, Zuckermann, Spencer, McEvoy, Walker, Williams.
Footnotes
- Received August 1, 2014.
- Accepted October 16, 2014.
This study was supported by the National Council for Replacement Refinement and Reduction [G0900775] (to R.S.B.W. and M.C.W.); and SMSdrug.net (to J.S.). R.S.B.W. is funded (in other projects) by the Doctor Hadwen Trust and did not participate in experiments involving animals, or cells or tissues from animals or from human embryos.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- aCSF
- artificial cerebrospinal fluid
- BCCA
- butylcyclohexane carboxylic acid
- EOA
- ethyl-octanoic acid
- HDAC
- histone deacetylase
- LDH
- lactose dehydrogenase
- MCT
- medium chain triglyceride
- MES
- maximum electric shock
- MOA
- methyl-branched octanoic acid
- MNA
- methyl-nonanoic acid
- OA
- octanoic acid
- PI
- propidium iodide
- PTZ
- pentylenetetrazol
- scMET
- subcutaneous metrazol seizure threshold test
- VPA
- valproic acid
- Copyright © 2014 by The American Society for Pharmacology and Experimental Therapeutics
References
JPET articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|





{kind=link}
{kind=link}
{kind=link}
{kind=link}