


Abstract
The ability of voltage-gated potassium channel α-subunits to form heteromultimers has complicated efforts to use toxins to characterize potassium channels in native cells. Here I investigate the effects of subunit composition on toxin blocking affinity, using three members of the Shaker subfamily of potassium channel α-subunits (Kv1.1, Kv1.2 and Kv1.4), which are known to form heteromultimers in vivo,in the Xenopus oocyte expression system. These subunits were coexpressed as pairs in which one member was toxin-sensitive and the other relatively insensitive. The blocking affinity of two dendrotoxins (DTX-I and δ-DTX) and a scorpion toxin (tityustoxin-Kα) on the resulting mixed population of channels was measured to evaluate three models of toxin block. The single subunit model, in which a single toxin-sensitive subunit renders the channel toxin sensitive, best described all of the data for the two dendrotoxins and the block of tityustoxin-Kα for a mixed population of channels composed of Kv1.1 and Kv1.2 subunits. However, with tityustoxin-Kα, the data for a mixed population of Kv1.2 and Kv1.4 subunits was fit best by a model in which the toxin interacts with all four subunits for high-affinity block. The data suggest that subunit composition of potassium channels can have a large effect on toxin block and that different toxins yield strikingly diverse results with the same pair of subunits, even when they are nearly identical in blocking affinity for the toxin-sensitive subunit.
The functional characterization of potassium channels in the CNS and elsewhere will require a greater understanding of the correspondence between potassium currents defined by biophysical and pharmacological criteria and our expanding knowledge of the molecular composition of potassium channels. This task has been complicated by the now well-known ability of potassium channel subunits to coassemble with members of the same molecularly defined subfamilies into heterotetramers (Christie et al., 1990; Isacoff et al., 1990; Ruppersberg et al., 1990). Peptide toxins from a variety of animal species have been used to inhibit components of potassium current in neurons and other cells (Benoit and Dubois, 1986; Halliwell et al., 1986; Hall et al., 1994;Brew and Forsythe, 1995), and these toxins have been tested on a number of cloned α-subunits that form functional potassium channels in heterologous expression systems (Stühmer et al., 1989;Ramaswami et al., 1990; Grissmer et al., 1994;Hopkins et al., 1994a). However, the molecular identity of channels expressed in native cells has not been deduced with certainty on the basis of the latter findings because of our incomplete understanding of the subunit specificity of these toxins and of the effect of subunit composition on toxin blocking affinity.
This study addresses the effects of voltage-gated potassium channel subunit composition on toxin blocking affinity using theXenopus oocyte expression system. I have focused on three members of the Shaker subfamily of voltage-gated potassium channel α-subunits: Kv1.1, Kv1.2 and Kv1.4 (Tempel et al., 1988;Chandy et al., 1990; Ramaswami et al., 1990). These subunits are known to coassemble into heteromultimeric channels in Xenopus oocytes (Ruppersberg et al., 1990;Christie et al., 1990; Hopkins et al., 1994b) andin vivo (Wang et al., 1993; Sheng et al., 1993; Rhodes et al., 1995). I have coexpressed these subunits as pairs (Kv1.1/Kv1.2, Kv1.2/Kv1.4 and Kv1.1/Kv1.4) and have used two dendrotoxins (DTX-I and δ-DTX) and tityustoxin-Kα to evaluate three models of toxin binding to heteromultimeric potassium channels. The subunit pairs and toxins were chosen such that one member of the pair of subunits would be toxin-sensitive and the other relatively insensitive. The results demonstrate that the model that best describes the data can be both toxin- and subunit-specific. For example, the blocking affinity of tityustoxin-Kα for a mixed population of channels composed of Kv1.1 and Kv1.2 subunits could be described by a model in which a single toxin-sensitive subunit renders the channel toxin-sensitive (single subunit model). However, this model did not describe the tityustoxin-Kα data for a mixed population of Kv1.2 and Kv1.4 subunits. The single subunit model provided the closest fit to the data for a mixed population of Kv1.2 and Kv1.4 subunits using DTX-I. The blocking affinities of δ-DTX on mixed populations of Kv1.1/Kv1.2 or Kv1.1/Kv1.4 subunits were best described by the single subunit model. The data suggest that potassium channel subunit composition can have a marked effect on a toxin’s blocking affinity and that the identity of the toxin is also an important factor.
Materials and Methods
Oocyte expression.
Plasmid DNAs encoding mKv1.1 and mKv1.2 (kindly provided by Dr. Bruce Tempel at the University of Washington) were linearized with EcoRI for in vitrotranscription of capped mRNAs (mCAP kit, Stratagene) using SP6 RNA polymerase (Hopkins et al., 1994b). Plasmid DNAs encoding hKv1.2 and hKv1.4 (kindly provided by Dr. Mark Tanouye at the University of California, Berkeley), were linearized with NotI forin vitro transcription of capped, polyadenylated mRNA using T7 RNA polymerase (Ramaswami et al., 1990). Unless otherwise noted, Kv1.1 refers to mKv1.1, Kv1.2 to hKv1.2 and Kv1.4 to hKv1.4. Adult female Xenopus laevis were anesthetized with MS-222 for half an hour, followed by surgical removal of 2 to 4 ovarian lobes. Pieces of ovary were washed in CFS containing 82.5 mM NaCl, 2 mM KCl, 1 mM MgCl2 and 5 mM HEPES (pH 7.6). Single oocytes free from follicular cells and connective tissue were obtained by treatment with 1 mg ml−1 collagenase (Type A, Boehringer Mannheim, Indianapolis, IN) in CFS at room temperature (22°C) for 3 hr. Healthy oocytes at maturation stage V and VI were selected and transferred to an incubation saline solution (ISS) containing 96 mM NaCl, 2 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 0.01 mg ml−1 penicillin, 0.01 mg ml−1 streptomycin and 5 mM HEPES (pH 7.4). After 24 hr of incubation at 18°C, the oocytes were injected with 50 nl of 3 to 1000 pg nl−1 of cRNA via a microinjector (Drummond, Broomall, PA). Oocytes were maintained in ISS at 18°C and tested for current expression 2 to 6 days after injection.
Electrophysiology and data analysis.
For whole-cell current measurements, a two-electrode voltage clamp (Warner OC-725B) was used. Both the voltage-recording and the current electrode were filled with 3 M KCl and had resistances of 0.5 to 2.0 MΩ. The oocytes were placed in an 0.5-ml chamber and were perfused continuously at 5 ml min−1 with FRS consisting of 115 mM NaCl, 2.5 mM KCl, 1.8 mM CaCl2 and 10 mM HEPES (pH 7.2). Transient capacitive and linear leakage currents were subtracted on-line by the P/4 procedure (Armstrong and Bezanilla, 1974) built into the data acquisition software (PClamp, Axon Instruments, Foster City, CA). The currents were filtered at 1 kHz and sampled at 5 kHz. The oocyte membrane was clamped at −70 mV, and membrane currents were elicited by 250-msec command voltages from −70 to 50 mV. To obtain concentration-inhibition data for an inhibitor, control data were obtained from a given oocyte in FRS, and then, 5 min after perfusion of inhibitor, data were obtained again. This procedure was repeated for an ascending series of inhibitor concentrations. I have determined that, at the solution flow rate used, 5 min is an adequate time interval to obtain “steady-state” current measurements for all inhibitors used. The peak potassium current at 50 mV was measured in the presence of each inhibitor concentration and normalized to the current obtained in control FRS at the same voltage (fractional inhibition). For a given concentration-inhibition curve, each data-point represents the mean fractional inhibition value obtained from the number of oocytes given in the figure legend for each curve. For the experiments in which toxins were applied sequentially to the same oocyte, the membrane potential was held at −70 mV, and 100-msec command voltages were applied to 50 mV (0.2 Hz). I observed no cumulative inactivation or rundown with any of the subunits tested with this protocol. After a 10-pulse base-line period, a toxin was applied directly to the bath solution. After the responses displayed a steady level of block, another toxin was applied in the same manner, and the experiment was continued until a new steady level of block was achieved. For the fitting of drug-binding isotherms to concentration-inhibition data, the adequacy of the fit was statistically evaluated with the chi-square goodness-of-fit test with a significance level of P < .05. All values are expressed as mean ± S.E.M.
To determine the expression ratios for K+ channel subunit pairs in the coexpression experiments, the same concentrations of cRNA for the two subunits coinjected in the coexpression study were injected separately into two groups of oocytes from the same batch, and the average peak current was determined from at least 10 oocytes in each of the three groups (one coinjected group and two single transcript-injected groups) on the day of an experiment. The average peak current of the coinjected oocytes was always at least the sum of the average peak currents of the oocytes injected with single cRNA transcripts (data not shown). The average number of channels (and subunits) per oocyte was estimated from the expressionN = I/pi, where I is mean peak current at 50 mV, p is the probability of channel opening (p ≈ 1 at this voltage), i is the single channel current amplitude for channels formed by each subunit type (determined in separate patch-clamp experiments) and N is the number of channels. This method might tend to underestimate channels formed from hKv1.4 subunits, because they display rapidN-type inactivation in contrast to the very slow, C-type inactivation displayed by channels formed from the other subunits (Hoshi et al., 1991; Ruppersberg et al., 1991;López-Barneo et al., 1993). This potential source of error should not affect any of the conclusions if it is small and constant from experiment to experiment, because it would affect all ratio estimates similarly. I could not simply coinject fixed ratios of concentrations of cRNA, because for a given concentration of cRNA, subunit expression levels were not equal. I chose not to transcribe cDNA mixtures (Kavanaugh et al., 1992) because of the differences in translation initiation sequences for the different plasmid vectors used. These differences may cause differential translational efficiency, yielding inaccurate estimates of expression ratios based on cRNA or cDNA concentrations.
Modeling.
The fraction of the total population of channels composed of a particular type of homomultimer or heteromultimer can be predicted with the binomial theorem if the subunit expression ratio is known:
The following inhibitor binding equation was used for all of the models presented:
I considered three models to describe the binding of an inhibitor to K+ channels (fig. 1): Thesingle subunit model assumes that the IC50 for an inhibitor increases linearly as the number of inhibitor-sensitive subunits decreases. A channel with one inhibitor-sensitive subunit has an IC50 value 4 times larger than a channel with four inhibitor-sensitive subunits. An inhibitor-sensitive subunit provides a “complete,” high-affinity binding site. More formally, letk and k* be the microscopic dissociation constants for inhibitor-sensitive and -insensitive subunits, respectively, that are equal to 4 × IC50 for the inhibitor-sensitive and -insensitive homomultimeric channels. Then

Three blocking affinity models. The white circles represent toxin-sensitive subunits, and the black circles represent toxin-insensitive subunits. See “Materials and Methods” for more details.
The energy additivity model assumes that all four subunits contribute to the binding site for an inhibitor; it is based on the model for external tetraethylammonium (TEA) binding to TEA-sensitive Kv1.1 (Heginbotham and MacKinnon, 1992; Kavanaugh et al.,1992). As the number of inhibitor-sensitive subunits is decreased, IC50 increases exponentially because of an effect on the binding free energy of the inhibitor with the channel:
The dominant negative model embodies the idea that all channels that have at least one inhibitor-insensitive subunit will be as insensitive to the inhibitor as a channel with four inhibitor-insensitive subunits. It is based on a model to describe charybdotoxin blocking affinity on some smooth muscle K+channels, which were believed to be heteromultimers composed of Kv1.2 and Kv1.5 subunits (Russell et al., 1995).
DTX-I and δ-DTX were obtained from Alomone Labs (Jerusalem, Israel). Tityustoxin-Kα was obtained from Dr. Mordecai Blaustein (University of Maryland). TEA was obtained from Sigma (St. Louis, MO). The toxins and TEA were diluted in FRS to obtain the final concentrations.
Results
TEA blocking affinity described by energy additivity model.
TEA blocking affinity for heteromultimeric K+ channels composed of TEA-sensitive Kv1.1 and mutant TEA-insensitive Kv1.1 subunits is well described by the energy additivity model (Heginbotham and MacKinnon, 1992; Kavanaugh et al., 1992). This is also true for heteromultimeric channels composed of Kv1.1 and relatively TEA-insensitive Kv1.2 subunits (Hopkins et al., 1994b). I attempted to reproduce this result as a check on the methods used here. Figure 2A depicts an experiment in which TEA-sensitive Kv1.1 was coinjected with TEA-insensitive Kv1.2 with a fractional expression of 0.6 for Kv1.1. The concentration-inhibition curve for the resultant mixed population of channels was well described by the energy additivity model, but not by the single subunit model or the dominant negative model. Figure 2B shows that as the fraction of Kv1.1 subunits was varied in three separate experiments, the IC50 values for the mixed population of channels was well fit by the line predicted for the energy additivity model. The single subunit and dominant negative models did not provide adequate fits to the data. The dominant negative model was found to provide very poor fits to any of the data sets; it will not be further depicted.

TEA blocking affinity data well fit by the energy additivity model. A) Concentration-inhibition curves using TEA on Kv1.1 (N = 5 oocytes), Kv1.2 (N = 8) and a mixed population of channels where the measured fractional expressions of Kv1.1 and Kv1.2 were 0.6 and 0.4, respectively (N = 3). The IC50 for the mixed population of channels was 5 mM. The predicted IC50 values were 0.6 mM for the single subunit model under these conditions, 3 mM for the energy additivity model and 52 mM for the dominant negative model. B) The IC50 values for TEA as a function of the fractional expression of TEA-sensitive Kv1.1 subunits in three mixing experiments. The predicted IC50 values for the three models as a function of the fractional expression of Kv1.1 are also shown as line segments. The lines were obtained by calculating the predicted IC50 values on the basis of each of the three models for each experimental expression ratio and drawing line segments through the predicted IC50 values. The energy additivity model provided the best fit at all measured expression ratios.
δ-DTX, DTX-I and tityustoxin-Kα block of homomultimeric K+ channels.
δ-DTX is a dendrotoxin isolated from the venom of Dendroaspis angusticeps that blocks some voltage-gated K+ channels (Benishin et al.,1988; Hall et al., 1994). Whereas Kv1.1 was very sensitive to δ-DTX, Kv1.2 was much less sensitive, and Kv1.4 was completely insensitive to this toxin (table 1; figs.3 and 4).
IC50 values for the four inhibitors used on Kv1.1, Kv1.2 and Kv1.4 homomultimers in Xenopus oocytes

δ-DTX blocking affinity for Kv1.1/Kv1.2 subunits well fit by the single subunit model. A) Whole-cell potassium currents from Xenopus oocytes recorded at a holding potential of −70 mV using test pulses to 50 mV. Top: Potassium currents measured in control FRS and in the presence of 1 nM δ-DTX for Kv1.1 subunit homomultimers. Center: Same for Kv1.2 homomultimers. Bottom: Same for a mixed population of Kv1.1 and Kv1.2 subunits expressed in a 1:1 ratio (fractional expression of Kv1.1 was 0.5). B) Concentration-inhibition curves using δ-DTX on Kv1.1 (N = 11), Kv1.2 (N = 6) and a mixed population of channels where the measured fractional expression of Kv1.1 was 0.5 (N = 9). The IC50 value for the mixed population of channels was 100 pM. The predicted IC50 values were 89 pM for the single subunit model under these conditions and 3 nM for the energy additivity model. C) The IC50 values for δ-DTX as a function of the fractional expression of δ-DTX-sensitive Kv1.1 subunits in three mixing experiments. The predicted IC50 values for the single subunit and energy additivity models as a function of the fractional expression of Kv1.1 are also shown as line segments. The single subunit model provided the best fit at all measured expression ratios.

δ-DTX blocking affinity for Kv1.1/Kv1.4 subunits well fit by the single subunit model. Concentration-inhibition curves using δ-DTX on Kv1.1 (N = 11), Kv1.4 (N = 3) and a mixed population of channels where the measured fractional expression of Kv1.1 was 0.24 (N = 11). The IC50 for the mixed population of channels was 634 pM. The predicted IC50 values were 1 nM for the single subunit model under these conditions and 404 nM for the energy additivity model.
DTX-I is a dendrotoxin isolated from the venom of Dendroaspis polylepis that was first shown to augment ACh release at neuromuscular synapses and has since been shown to block some K+ channels (Strydom, 1972; Harvey and Karlsson, 1982). Both Kv1.1 and Kv1.2 are sensitive to DTX-I, Kv1.2 being somewhat more sensitive to the toxin (table 1; fig. 7). As with δ-DTX, Kv1.4 was insensitive to DTX-I (table 1; fig. 7).
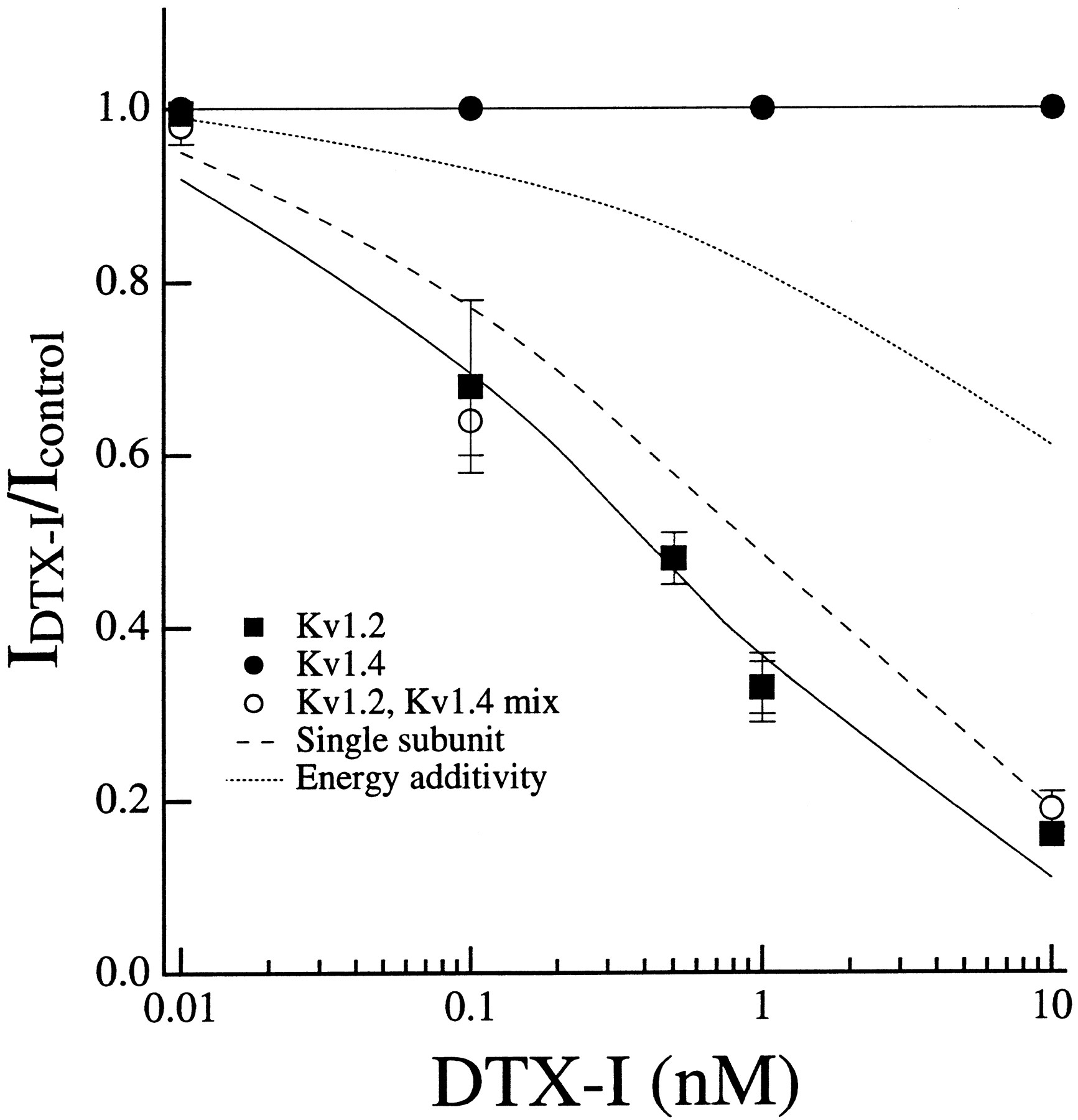
DTX-I blocking affinity for Kv1.2/Kv1.4 subunits well fit by the single subunit model. Concentration-inhibition curves using DTX-I on Kv1.2 (N = 13), Kv1.4 (N = 4) and a mixed population of channels where the measured fractional expression of toxin-sensitive Kv1.2 was 0.55 (N = 6). The IC50 for the mixed population of channels was indistinguishable from the IC50 of DTX-I for Kv1.2 homomultimers (410 pM). The predicted IC50 values were 914 pM for the single subunit model under these conditions and 24 nM for the energy additivity model.
Tityustoxin-Kα is a peptide toxin isolated from the Brazilian scorpion Tityus serrulatus. It blocks voltage-gated potassium currents in some preparations (Blaustein et al.,1991; Eccles et al., 1994; Rogowski et al., 1994) and can block Kv1.2 with high affinity (Werkman et al.,1993). I have also found that tityustoxin-Kα can inhibit Kv1.2 (IC50 = 105 pM for mKv1.2; IC50 = 550 pM for hKv1.2) and that Kv1.1 and Kv1.4 are insensitive to tityustoxin-Kα (table 1; figs. 5A and6).

Tityustoxin-Kα blocking affinity for Kv1.1/Kv1.2 subunits well fit by the single subunit model. A) Concentration-inhibition curves using tityustoxin-Kα on Kv1.1 (N = 4), Kv1.2 (N = 5) and a mixed population of channels where the measured fractional expression of toxin-sensitive Kv1.2 was 0.22 (N = 10). The IC50 for the mixed population of channels was 1.2 nM. The predicted IC50 values were 10 nM for the single subunit model under these conditions and 233 nM for the energy additivity model. B) The IC50 values for tityustoxin-Kα as a function of the fractional expression of tityustoxin-Kα-sensitive Kv1.2 subunits in three mixing experiments. The predicted IC50 values for the single subunit and energy additivity models as a function of the fractional expression of Kv1.2 are also shown as line segments. The single subunit model provided the best fit at all measured expression ratios.

Tityustoxin-Kα blocking affinity for Kv1.2/Kv1.4 subunits well fit by the energy additivity model. Concentration-inhibition curves using tityustoxin-Kα on Kv1.2 (N = 5), Kv1.4 (N = 3) and a mixed population of channels where the measured fractional expression of toxin-sensitive Kv1.2 was 0.71 (N = 5). The IC50 for the mixed population of channels was 7 nM. The predicted IC50 values were 845 pM for the single subunit model under these conditions and 7 nM for the energy additivity model.
δ-DTX blocking affinity for Kv1.1/Kv1.2 and Kv1.1/Kv1.4 heteromultimers described by single subunit model.
When Kv1.1 and Kv1.2 were coexpressed in a 1:1 ratio (Kv1.1 fractional expression 0.5), the resulting channels had high sensitivity to δ-DTX (fig. 3A, and B). The data from the mixed population of channels were well described by the single subunit model (fig. 3B). When the fraction of Kv1.1 subunits was varied in three separate experiments, the IC50 values for the mixed population of channels were well fit by the line predicted for the single subunit model. The energy additivity model provided a poor fit to the data (fig. 3C).
I obtained similar results when Kv1.1 was coexpressed with Kv1.4 (fig.4). In this experiment, δ-DTX-insensitive Kv1.4 subunits were overexpressed relative to Kv1.1 subunits (Kv1.1 fractional expression 0.24). The mixed population of channels displayed an IC50of 634 pM, and the IC50 values predicted by the single subunit and energy additivity models for this expression ratio were 1 and 404 nM, respectively.
Tityustoxin-Kα blocking affinity for Kv1.2/Kv1.1 heteromultimers described by single subunit model.
When tityustoxin-Kα-sensitive Kv1.2 and tityustoxin-Kα-insensitive Kv1.1 were coexpressed (Kv1.2 fractional expression 0.22), the resulting channels were very sensitive to tityustoxin-Kα (fig. 5A). The single subunit model provided the best fit to the data. In three experiments using different expression ratios, the IC50 values for the mixed population of channels were well fit by the line predicted for the single subunit model and were not well fit by the line predicted for the energy additivity model (fig. 5B).
Tityustoxin-Kα blocking affinity for Kv1.2/Kv1.4 heteromultimers not described by single subunit model.
When tityustoxin-Kα-sensitive Kv1.2 and tityustoxin-Kα-insensitive Kv1.4 were coexpressed (Kv1.2 fractional expression 0.71), the sensitivity of the resulting channels to tityustoxin-Kα was much lower than predicted for the single subunit model and was well fit by the energy additivity model (fig. 6). The single subunit model predicted that the sensitivity of the mixed population of channels to tityustoxin-Kα should be nearly identical to that for Kv1.2 homomultimers in this experiment, because I estimated that 71% of the expressed subunits were toxin-sensitive. The single subunit model predicted an IC50 of about 800 pM for the mixed channel data, whereas the energy additivity model predicted an IC50 of about 7 nM. The IC50 for the mixed subunit data was 7 nM.
DTX-I blocking affinity for Kv1.2/Kv1.4 heteromultimers described by single subunit model.
To determine whether the results with tityustoxin-Kα on Kv1.2/Kv1.4 heteromultimers was a general property of toxin binding to this subunit pair, I performed a similar mixing experiment but instead used DTX-I. This toxin was chosen because its blocking affinity for Kv1.2 homomultimers was nearly identical to that of tityustoxin-Kα (table 1) and, like tityustoxin-Kα, it displayed low blocking affinity for Kv1.4 (fig. 7). In contrast to the results with tityustoxin-Kα, I found that the blocking affinity for DTX-I on Kv1.2/Kv1.4 heteromultimers was more closely described by the single subunit model than by the energy additivity model (fig. 7).
Sequential application of tityustoxin-Kα and DTX-I to oocytes coexpressing Kv1.2/Kv1.4 heteromultimers.
To obtain further evidence that the blocking affinity of tityustoxin-Kα and DTX-I for a mixed population of Kv1.2 and Kv1.4 subunits cannot be described by the same blocking affinity models, I performed experiments in which each toxin was applied sequentially at the same concentration to the same oocyte. Two such experiments are shown in figure8A and B. Regardless of the order in which the toxins were applied, DTX-I displayed significantly greater potency against the mixed population of channels than did tityustoxin-Kα, in spite of their nearly identical potency against Kv1.2 homomultimers (table 1). Results from 16 oocytes are shown in figure 8C. DTX-I (1 nM) blocked about 30% of the potassium current on average, whereas 1 nM tityustoxin-Kα blocked about 10% of the current, and this difference was statistically significant. Figure 8D shows the toxin-binding isotherms for DTX-I and tityustoxin-Kα on Kv1.2 and Kv1.4 and the predicted toxin-binding isotherms for the single subunit and energy additivity models for the measured fractional expression (Kv1.2: 0.19) in this batch of oocytes. These results predict that a toxin whose blocking affinity is described by the single subunit model under these conditions should block about 25% of the current, whereas for the energy additivity model, the toxin should block about 5% of the current. These results strongly suggest that for a mixture of Kv1.2 and Kv1.4 potassium channel subunits coexpressed in oocytes, the blocking affinity of DTX-I can be described by the single subunit model but that of tityustoxin-Kα cannot.

DTX-I and tityustoxin-Kα blocking affinity for Kv1.2/Kv1.4 subunits described by different models. A) Experiment in which DTX-I and tityustoxin-Kα (1 nM each) were bath-applied sequentially to the same oocyte in which cRNA encoding Kv1.2 and Kv1.4 had been coinjected. Test pulses to 50 mV were applied at 5-sec intervals from a holding potential of −70 mV. B) Same as panel A except that tityustoxin-Kα was applied first, followed by DTX-I. C) Summary results from 16 oocytes from the same batch in which 8 oocytes were perfused first with DTX-I and then with tityustoxin-Kα, and 8 oocytes were perfused first with tityustoxin-Kα and then with DTX-I (each at 1 nM). The measured fractional expression of toxin-sensitive Kv1.2 was 0.19. The magnitude of the decrease in the fractional response amplitude in the presence of the two toxins, relative to the base-line amplitude, is indicated by the stipled (tityustoxin-Kα) and gray (DTX-I) regions of the bars. The difference in the extent of block between DTX-I and tityustoxin-Kα was significant (P < .05), regardless of the order of toxin application. D) Model results for data presented in panel C. Concentration-inhibition curves using DTX-I and tityustoxin-Kα on Kv1.2 (N = 13 and 5, respectively), Kv1.4 (N = 4 and 3, respectively) and the predicted isotherms for the single subunit and energy additivity models, assuming a fractional expression of 0.19 for toxin-sensitive Kv1.2 (∼4:1). The IC50 values of DTX-I and tityustoxin-Kα for Kv1.2 homomultimers were 410 pM and 550 pM, respectively. The predicted model isotherms for each toxin are so similar that they were “lumped” into one curve for simplicity. Using a concentration of 1 nM, about 25% of the steady-state current should be blocked under the single subunit model and about 5% of the current should be blocked under the energy additivity model. This is very close to what was observed in the experiments summarized in panel C for DTX-I and tityustoxin-Kα, respectively.
Discussion
I have characterized the blocking affinity of three peptide toxins on three cloned potassium channel subunits either expressed alone as homomultimers or coexpressed as a mixed population of channels (homomultimers and heteromultimers) in Xenopus oocytes. For two subunit combinations (Kv1.1/Kv1.2 and Kv1.1/Kv1.4) δ-DTX blocking affinity for the mixed population of channels was well described by the single subunit model. The blocking affinity of DTX-I for Kv1.2/Kv1.4 subunits was also described by this model. In contrast, the results with tityustoxin-Kα were dependent on subunit composition. For Kv1.1/Kv1.2 subunits, the data with tityustoxin-Kα were consistent with the single subunit model, whereas for Kv1.2/Kv1.4 subunits, the single subunit model did not fit the data.
The methodology and its assumptions.
Other studies addressing similar questions have used tandem cDNA constructs of potassium channel subunits to constrain the stoichiometry of the channels (Isacoffet al., 1990; Liman et al., 1992; Heginbotham and MacKinnon, 1992; Kavanaugh et al., 1992). Although this approach has yielded important information, it is difficult to know that the stoichiometry has been constrained in the manner intended (McCormack et al., 1992) and that heteromultimeric channels constructed in this manner are functionally equivalent to channels with identical stoichiometry that coassemble. I have chosen the mixing approach of unaltered subunits because the expressed channels more closely approximate their “native” state. The mixing approach complements these other methods.
The methods used here assume that subunit mixing in oocytes can be described by the binomial theorem and that subunit position in the tetramer can be ignored, which reduces the number of types of heteromultimers to model from 14 to 3. There is evidence both for (Naranjo and Miller, 1996) and against (Taglialatela et al.,1994) the first assumption. If like subunits are more likely to coassemble than unlike ones, then there would be more homomultimers and 3:1 heteromultimers, and fewer 2:2 heteromultimers, than would be predicted by chance. It might be expected that this type of preferential assembly would yield biphasic concentration-inhibition curves. This was not observed. The converse case, that nonidentical subunits preferentially coassemble, would result in enrichment in heteromultimers at the expense of homomultimers. This case would predict that when toxin-insensitive subunits were in greater abundance than toxin-sensitive ones, the toxin sensitivity of the channels would be greater than predicted, because there would be fewer toxin-insensitive homomultimers. I observed this effect (compare fig.5A), although not consistently. Overall, there were no consistent features in the data that suggested significant deviations from random coassembly of subunits. For the second assumption, I have insufficient information to model all possible heteromultimers with these or any other models. However, there is evidence that this assumption is not unreasonable (Naranjo and Miller, 1996).
To check the modeling methods, I used TEA binding to channels composed of TEA-sensitive Kv1.1 and TEA-insensitive Kv1.2 subunits, which was described by the energy additivity model in a previous study (Hopkinset al., 1994b) and when Kv1.1 was coexpressed with TEA-insensitive mutant Kv1.1 subunits in two other studies (Heginbotham and MacKinnon, 1992; Kavanaugh et al., 1992). The data were well fit by the energy additivity model only (fig. 2, A and B). This suggests that any systematic errors or violations of the modeling assumptions were not large enough to yield misleading results.
Additionally, I obtained a Hill coefficient of 0.95 for the TEA data for Kv1.1 (table 1), which has been shown to display a Hill coefficient of about 1 for the rat homolog of this subunit in a previous study (Christie et al., 1990). This suggests that Hill coefficients less than 1 obtained for the three toxins were not due to the specific methods used to obtain the concentration-inhibition data, because they were the same for both TEA and the toxins.
δ-DTX and DTX-I blocking affinities described by the single subunit model.
In mixing experiments with Kv1.1 and Kv1.2 subunits, the blocking affinity of δ-DTX was best described by the single subunit model for several expression ratios of the two subunits (fig. 3). This suggests that heteromultimeric channels composed of these subunits would display blocking affinities very similar to those of Kv1.1 homomultimers. Similar results were obtained when Kv1.1 and Kv1.4 were coexpressed (fig. 4).
DTX-I was found to display blocking affinity described by the single subunit model when Kv1.2 and Kv1.4 subunits were coexpressed (fig. 7). I have previously demonstrated similar results with DTX-I when Kv1.1 and Kv1.3 were coexpressed (Hopkins et al., 1994a). When Kv1.2 and the DTX-I-insensitive mutant Kv1.1 were coexpressed in oocytes, the DTX-I data for the resulting channels were well fit by the single subunit model (table 2; Hopkinset al., 1994b). Therefore, DTX-I blocking affinity for heteromultimeric channels in which one type of subunit was toxin-sensitive and the other was not were always best described by the single subunit model.
Best inhibitor blocking model for three coexpressed subunit pairs inXenopus oocytes
That DTX-I and δ-DTX display blocking affinity consistent with the single subunit model raises the question of whether this is a general property of all dendrotoxins. Using tandem tetramers with wild-type and toxin-insensitive mutant subunits, Tytgat et al. (1995)found that the blocking affinity of α-DTX was best described by the energy additivity model. Their data suggest that the data presented here may not be representative of all dendrotoxins.
The model describing blocking affinity of tityustoxin-Kα is subunit-specific.
The blocking affinity of tityustoxin-Kα for Kv1.1/Kv1.2 channels was well described by the single subunit model, whereas for Kv1.2/Kv1.4 channels, the data were well fit by the energy additivity model (figs. 6 and 8). However, I favor the idea that some other model is more appropriate to describe these data for the following reasons. The energy additivity model describes the binding of an inhibitor that interacts with four subunits simultaneously. That tityustoxin-Kα binding to Kv1.2 can be described by the single subunit model when coexpressed with Kv1.1 suggests that a single Kv1.2 subunit provides an adequate high-affinity binding site and this is incompatible with the energy additivity model when Kv1.2 is coexpressed with another subunit (Kv1.4). Another model that could describe these data would invoke electrostatic repulsion between Kv1.4 subunits and the toxin. Thus Kv1.4 subunits would not be a neutral influence on the binding of tityustoxin-Kα to the heteromultimeric channels that were also composed of toxin-sensitive Kv1.2 subunits but would instead contribute electrostatic repulsion. The net result would be decreased blocking affinity of the toxin for the channel. Tityustoxin-Kα is a basic peptide (Rogowski et al., 1994), and Kv1.4 has a basic residue (lysine) at a position in the pore-forming region that is occupied by a valine in Kv1.2. If tityustoxin-Kα binds in the pore-forming region, then this site is an obvious candidate for an electrostatic repulsive interaction between the toxin and Kv1.4 subunits. Support for this hypothesis comes from Gross and MacKinnon (1996), who used lysine-scanning mutagenesis of the agitoxin binding site of Shaker potassium channels. Many of these lysine point mutations decreased the affinity of agitoxin for the channel even when it was present in only one subunit of the tetramer. The quantitative effects of these mutations were much closer to what would be predicted by the energy additivity than to what the single subunit model would predict (compare table 1; figs. 2 and 3 of Gross and MacKinnon). This is interesting because another nonlysine point mutation in the pore-forming region of Shaker (D431N), which renders it insensitive to agitoxin, yields data that are well described by the single subunit model when this mutant subunit is mixed with wild-type subunits inXenopus oocytes (MacKinnon, 1991). Another study has shown that the binding free energy of a charged ligand to a protein is influenced predictably by the number of positive or negative charges on the protein that are remote from the binding site (Gao et al., 1996). Thus electrostatic interactions between a peptide toxin with a net positive charge and positive residue(s) on a channel subunit could decrease the net binding free energy of the toxin to a channel with toxin-sensitive subunits. That the charge distribution on the toxin is also important is suggested by the toxin specificity (tityustoxin-Kα but not DTX-I) of this effect.
Implications for the use of toxins to characterize potassium channels in native cells.
Peptide toxins are used to block potassium currents in neurons and other cells, and in some cases a degree of selectivity for components of current has been achieved (Benoit and Dubois, 1986; Halliwell et al., 1986). However, the molecular identity of the inhibited channels cannot be inferred from data from cloned channel subunits. First, the subunits that are constituents of channels under consideration may not have been cloned and characterized. Second, few if any of the toxins are highly selective for one subunit (Hopkins et al., 1996). Third, the data presented here suggest that the subunit composition of the channel can dramatically affect toxin blocking affinity, even for a toxin whose blocking affinity can be described by the single subunit model under some conditions. Finally, other factors peculiar to the cells under study may affect toxin binding independent of the intrinsic affinity of the toxin for the channels.
How, then, might toxins be helpful in characterizing potassium channels in native cells? Evidence suggests that some toxins are selective by class of channels, such as voltage-gated vs.large-conductance calcium-activated potassium channels (Galvez et al., 1990; Garcia-Calvo et al., 1993). In the context of molecular and functional characterization of potassium channelsin situ, a toxin would be most useful when used in conjunction with other toxins and experimental techniques. Toxins will continue to be useful in discerning the roles of potassium current components that shape spike firing patterns and other facets of cellular physiologic activity (Brew and Forsythe, 1995).
Acknowledgments
I thank Drs. Richard Aldrich, George Miljanich and Stephen M. Smith for comments on the manuscript, Dr. Bruce Tempel for mKv1.1 and mKv1.2, Dr. Mark Tanouye for hKv1.2 and hKv1.4 and Beth Sampson for technical assistance.
Footnotes
-
Send reprint requests to: William F. Hopkins, Neurex Corporation, 3760 Haven Ave., Menlo Park, CA 94025.
- Abbreviations:
- α-DTX
- α-dendrotoxin
- CFS
- calcium-free solution
- δ-DTX
- δ-dendrotoxin
- DTX-I
- dendrotoxin-I
- FRS
- frog Ringer solution
- IC50
- compound concentration inhibiting 50% of potassium current
- ISS
- incubation saline solution
- tityustoxin-Kα
- scorpion toxin
- Received November 5, 1997.
- Accepted February 23, 1998.
- The American Society for Pharmacology and Experimental Therapeutics
References
JPET articles become freely available 12 months after publication, and remain freely available for 5 years.Non-open access articles that fall outside this five year window are available only to institutional subscribers and current ASPET members, or through the article purchase feature at the bottom of the page.
|




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}