


Abstract
Fabry disease (FD) is a lysosomal storage disorder caused by mutations in the gene for α-galactosidase A, inducing a progressive accumulation of globotriaosylceramide (GB3) and its metabolites in different organs and tissues. GB3 deposition does not fully explain the clinical manifestations of FD, and other pathogenetic mechanisms have been proposed, requiring the identification of new biomarkers for monitoring FD patients. Emerging evidence suggests the involvement of mitochondrial alterations in FD. Here, we propose mitochondrial-related microRNAs (miRs) as potential biomarkers of mitochondrial involvement in FD. Indeed, we demonstate that miRs regulating different aspects of mitochondrial homeostasis including expression and assembly of respiratory chain, mitogenesis, antioxidant capacity, and apoptosis are consistently dysregulated in FD patients. Our data unveil a novel noncoding RNA signature of FD patients, indicating mitochondrial-related miRs as new potential pathogenic players and biomarkers in FD.
SIGNIFICANCE STATEMENT This study demonstrates for the first time that a specific signature of circulating mitochondrial miRs (mitomiRs) is dysregulated in FD patients. MitomiRs regulating fundamental aspects of mitochondrial homeostasis and fitness, including expression and assembly of the respiratory chain, mitogenesis, antioxidant capacity, and apoptosis are significantly dysregulated in FD patients. Taken together, these new findings introduce mitomiRs as unprecedented biomarkers of FD and point at mitochondrial dysfunction as a novel potential mechanistic target for therapeutic approaches.
Introduction
Fabry disease (FD) is an X-linked inherited disorder of glycosphingolipid metabolism characterized by reduced or absent lysosomal α-galactosidase A (α-GAL A) activity due to mutations of the GLA gene (Germain, 2010), resulting in a progressive accumulation of globotriaosylceramide (GB3) and its metabolites (Germain, 2010; Miller et al., 2020). The clinical presentation of FD patients includes a poor quality of life and a reduced life expectancy due to complications in multiple organs (Morand et al., 2019). Cardiovascular disease remains the leading cause of death in these patients (Sorriento and Iaccarino, 2021). At the molecular level, GB3 deposition is generally considered one of the key pathogenetic mechanisms of this rare pathology, and the actual therapies are aimed at correcting the enzymatic deficiency, either by enzyme replacement therapy (ERT) or enzyme stabilization (e.g., using a chaperone, like Migalastat) (Ioannou et al., 2001; Pisani et al., 2012; Germain et al., 2016; Azevedo et al., 2020; Riccio et al., 2020). However, these therapeutics cannot revert FD pathology and its clinical manifestations, thereby suggesting that other pathogenetic mechanisms are involved. This aspect is further supported by the great variability of clinical phenotypes occurring among patients with the same genetic mutation (Sorriento and Iaccarino, 2021), maybe depending on different triggering mechanisms. For instance, an altered mitochondrial respiration has been suggested in fibroblasts isolated from FD patients (Das and Naim, 2009; Namdar et al., 2012; Rozenfeld and Feriozzi, 2017). An early diagnosis of FD is decisive for reducing the morbidity and mortality and activate organ-specific therapeutic interventions.
We and others have shown that microRNAs (miRs), small noncoding RNAs of ∼22 nucleotides, may serve as reliable biomarkers of various human disorders (Varzideh et al., 2022; Santulli et al., 2014; Sardu et al., 2014; Novak et al., 2015; Santulli, 2015; Santulli, 2016; Cammarata et al., 2018; Mone et al., 2022; Dama et al., 2020; Donati et al., 2021; Jankauskas et al., 2021; Kanach et al., 2021; Kansakar et al., 2022). Therefore, the miR profile could be a promising biomarker candidate for diagnosis and prognosis of FD. Alterations in over 100 circulating miRs were identified in blood samples from FD patients in treatment with ERT compared with untreated patients (Xiao et al., 2019). Increased expression of miR-29a-3p and miR-200a-3p were detected in urinary extracellular vesicles isolated from patients with FD nephropathy (Levstek et al., 2021); miR-let-7a and miR-let-7d were significantly increased in FD after therapy (Maier et al., 2021). These data point at miR profiling as a powerful diagnostic tool of FD progression and drug testing, as well as indicators of the development of specific FD phenotypes. Furthermore, deciphering a specific FD miR signature could help to identify alternative pathogenetic mechanisms and identifying specific therapeutic targets.
Several miRs are specifically associated with the mitochondrial phenotype in human diseases (Zhang et al., 2021). Some of them, also known as mitochondrial miRs (mitomiRs), are produced by the mitochondrial genome to regulate the mRNA transcription in loco. Other mitomiRs can target nuclear-encoded mRNAs localized on the mitochondrial surface or are necessary for mitochondrial homeostasis (Song et al., 2019). The mechanisms underlying mitomiR biogenesis and action sites are still poorly understood, however, they are a fascinating class of miRs that could allow tracking of mitochondrial status and health in pathologic conditions. Indeed, differentially expressed mitomiRs have been observed in heart failure (Pinti et al., 2017; Wang et al., 2017) where mitochondrial dysfunction is evident (Santulli et al., 2015; Sties et al., 2018; Del Campo et al., 2021; Guitart-Mampel et al., 2021; Morciano et al., 2021; Yang et al., 2021; Schwemmlein et al., 2022). Despite their potential power in mirroring systemic mitochondrial homeostasis, the functional contribution of mitomiRs in FD has never been investigated. Here, we evaluated in FD patients a cluster of mitomiRs and miRs with a recognized role in regulating mitochondrial fitness. Our data provide new insights about mitomiRs as biomarkers and therapeutic targets, adding a new mosaic tile in the delineation of the complex FD pathophysiology, potentially implicating mitochondrial dysfunction.
Materials and Methods
Study Population
FD patients with a confirmed FD diagnosis and healthy controls were recruited from the FD Clinic of Federico II University Hospital from March 2021 to February 2022. FD patients met the following inclusion criteria: 1) Adult ≥18 years of age, 2) confirmed diagnosis of classic FD, and 3) written informed consent. The diagnostic algorithm was based on the measurement of α-GAL A activity and on the genetic confirmatory testing (Vardarli et al., 2020). The study was officially approved by the Institutional Ethical Committee (protocol n. 181/19).
Blood Collection and Circulating miRs Determination
Peripheral blood was collected in EDTA-tubes and plasma was obtained by centrifugation as previously reported (Gambardella et al., 2020a). We extracted microRNAs using the miRVana miRNA isolation kit (Thermo Fisher Scientific; Waltham, MA) according to the manufacturer’s protocol. The quality of miR was determined using Agilent Small RNA Kit (Matarese et al., 2020; Mone et al., 2021). A custom panel of mitomiRs was quantified as we previously described (Gambardella et al., 2021).
Isolation of Peripheral Blood Mononuclear Cells and Western Blot Analysis
Peripheral blood mononuclear cells (PBMCs) were isolated as previously described (Santulli et al., 2011; Gambardella et al., 2020a; Gambardella et al., 2022). Briefly, PBMC rings were obtained by blood stratification (diluted 1:1 in PBS) on ficoll gradient using HISTOPAQUE-1077 (Merck; Germany). PBMC rings were lysed and the protein concentration was determined spectrophotometrically. Western blot analysis was conducted as previously reported (Gambardella et al., 2020a). Briefly, proteins were separated by 4%–12% SDS-PAGE gel and transferred to an Immobilon-P nitrocellulose filter (Merck); levels of electron transport chain (ETC) subunits were determined by using antibodies for total OXPHOS (abcam), and purified proteins (ETC complexes) as positive control. Actin levels used as loading control were visualized by a specific primary antibody (Santa Cruz Biotechnology, Dallas, TX). Secondary peroxidase-conjugated antibodies (ImmunoReagents; Raleigh, NC) were used to visualize antigen-antibody complexes on nitrocellulose by chemiluminescence. A standard chemiluminescence reaction kit (Thermo Fisher Scientific) was used for autoradiography on film.
Statistical Analysis
All experiments were carried out at least in triplicate by blinded investigators; we performed ANOVA with Bonferroni post hoc test or unpaired t test, as appropriate, where applicable. A significance level of P < 0.05 was assumed for all statistical evaluations. Statistics were computed with GraphPad Prism (v. 9.0) software (San Diego, CA).
Results
Alterations of Several mitomiRs in FD Patients
After having excluded 11 patients with late-onset nonclassic variants of FD, we enrolled 63 D and 14 healthy controls. Table 1 summarizes the main clinical characteristics of our population. Circulating miRs were assessed on plasma from FD patients compared with age-matched healthy controls. Specifically, the analysis was focused on a custom panel of key mitomiRs. Overall, a considerable dysregulation of mitomiRs was detected, with FD patients showing a pronounced clusterization (Fig. 1). Indeed, for most of the analyzed mitomiRs, we observed a significant difference in the relative expression between the two groups.
Characteristics of FD and control populations
Data are expressed as mean ± S.D. or percent.

Clusterization analysis of circulating mitomiRs. Heat map showing the profile of mitomiRs’ relative abundance between healthy controls and patients with FD.
The mitomiRs affected in the FD population regulate several aspects of mitochondrial health and biology.
In FD patients, we found the upregulation of miR-181c, miR-338, miR-130, and miR-210, which inhibit mitochondrial ETC expression and assembly (Fig. 2A). Indeed, the expression of ETC complexes was reduced in PBMCs isolated from FD patients (Fig. 2B).

Analysis of miRs associated with mitochondrial health and function. (A) Significant dysregulation of circulating miRs related to the expression and assembly of mitochondrial respiratory chain in patients with Fabry disease (FD). Median (dashed line) and quartiles (dotted lines) are indicated in the violin plots. (B) Western blot analysis of mitochondrial ETC in PBMCs revealed a significant downregulation of I-II-IV and ATP synthase complexes in FD; actin was used as loading control; data are mean ± S.E. (C) Purified proteins provided by the supplier, used as molecular weight referrals. *P < 0.05, **P < 0.005, ***P < 0.0005, FD versus control. CTRL +, positive control.
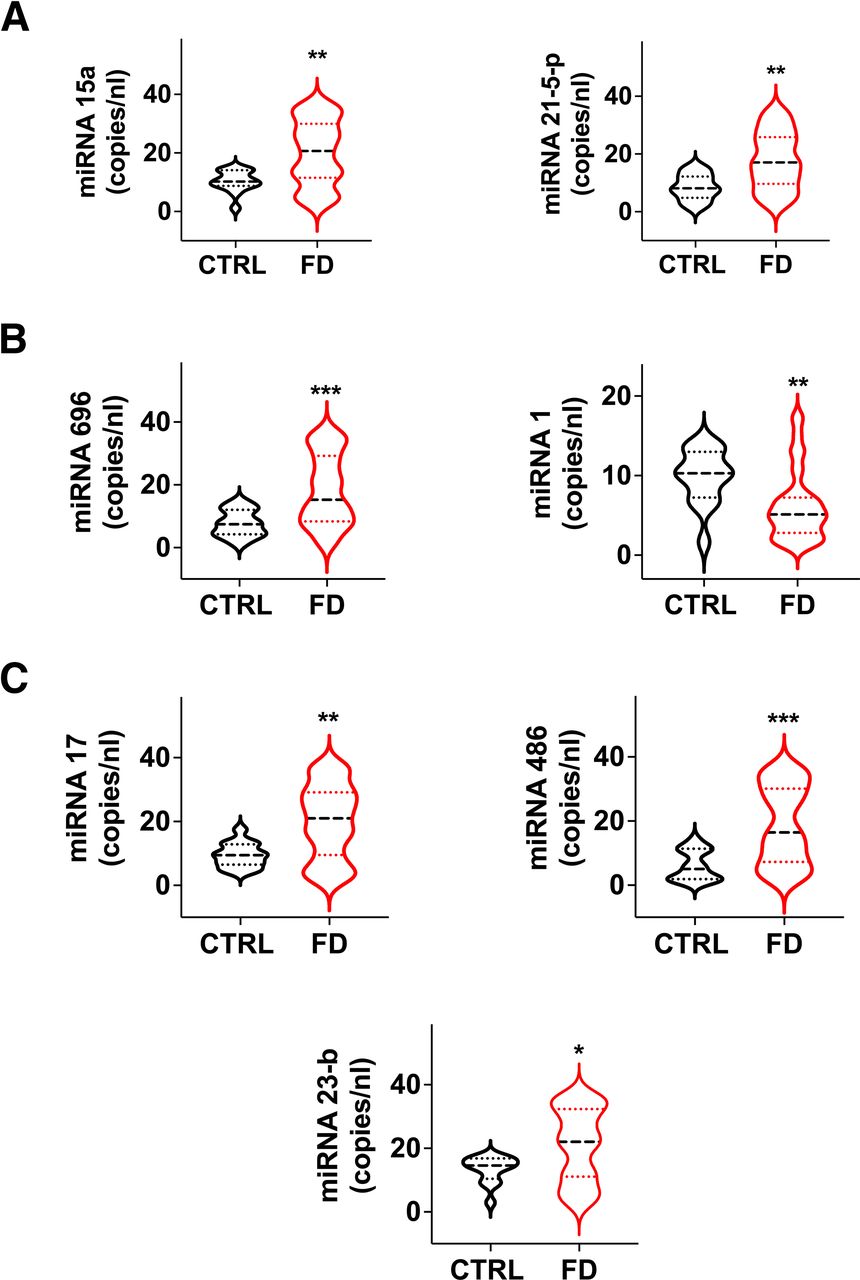
The class of miRs regulating mitochondrial-dependent metabolic pathways was also affected. Levels of miR-15a and miR-21-5p, regulating the expression of Uncoupling Protein 2 (UCP-2) and lipid content and peroxidation (Li et al., 2012; Nasci et al., 2019), were significantly augmented in FD patients compared to healthy controls (Fig. 3A).

Analysis of circulating miRs associated with mitochondrial health and function. Significant dysregulation in FD patients of miRs regulating mitochondrial metabolism (A), mitochondrial biogenesis and renewal (B), and mitochondrial oxidative damage and apoptosis (C). Medians (dashed line) and quartiles (dotted lines) are indicated in the violin plots. *P < 0.05, **P < 0.005, ***P < 0.0005.
Mitochondrial biogenesis and renewal are essential for a proper mitochondrial function, and are therefore strictly regulated both at the transcriptional and post-transcriptional levels (Jornayvaz and Shulman, 2010); miR-696 regulates mitochondrial biogenesis by targeting peroxisome proliferator-activated receptor gamma coactivator-1α (PGC-1α) (Aoi et al., 2010), while miR-1 has been reported to enhance the transcription of mitochondrial genomes (Zhang et al., 2014). Interestingly, miR-696 was upregulated while miR-1 was downregulated in the FD population (Fig. 3B).
Additionally, the antioxidant capacity and the mitochondrial proapoptotic pathway can be modulated by miRs: miR-17 and miR-23b are able to suppress critical primary mitochondrial antioxidant enzymes (Schmidt, 1990; Xu et al., 2010); miR-486 may regulate apoptosis via the p53-BCL-2–mediated mitochondrial apoptotic pathway (Sun et al., 2017). In our analysis, miR-17, miR-23b, and miR-486 were consistently upregulated in FD patients (Fig. 3C), further supporting our view of mitochondrial stress and/or damage in FD.
Effect of Treatment on mitomiRs Levels
We then dichotomized FD population in naïve and treated patients to test the effect of treatment on the levels of those mitomiRs that were significantly different between FD and controls (miR-181c, miR-338, miR-130a, miR-210, miR-15a, miR-21-5-p, miR-696, miR-1, miR-17, miR-486, miR-23b). Treatment influenced miR-1. In fact, this mitomiR is downregulated in untreated FD patients, and is restored by treatment to the levels observed in control subjects (Fig. 4; Table 2). No effect of treatment was observed for any of the other explored mitomiRs, which remained statistically different from control subjects (Table 2).

Effect of treatment on circulating miR-1 in Fabry disease (FD). Evaluation of mitomiRs in untreated and treated FD patients. Untreated FD patients show a reduction in circulating miR-1; treatment restores miR-1 levels in FD patients to the levels observed in control subjects. Data are mean ± S.E. *P < 0.05 versus controls; **P <0.05 versus untreated FD patients.
Effects of treatment on circulating mitomiRs in FD
Data are expressed as mean ± SE Data were analyzed by ANOVA with Bonferroni post hoc test.
Discussion
Our data reveals for the first time a dysregulation of mitomiRs in FD. In particular, miRs regulating different aspects of mitochondrial biology including oxidative phosphorylation capacity and energetic metabolism, mitochondrial biogenesis, mitochondrial oxidative damage, and apoptosis, were consistently affected. The upregulation of miRs targeting ETC complexes, thus inhibiting their expression and/or assembly, was particularly interesting. Indeed, miR-181c has been reported to directly bind the 3' untranslated region (3′UTR) of mitochondrial respiratory chain complex 1, determining its downregulation (Das et al., 2012). Similarly, miR-338 induces the downregulation of mitochondrial transport chain complex IV, and its overexpression decreases mitochondrial activity (Aschrafi et al., 2008). A potential interaction between miR-130 and ETC complex III was described (Kren et al., 2009) as well. Additionally, the protein involved in the complex IV assembly, Cytochrome C Oxidase Assembly Factor Heme A:Farnesyltransferase (COX10), is the target of miR-210 (Colleoni et al., 2013).
Consistently, we observed the downregulation of ETC complexes in cells derived from FD patients compared with controls. According with the reduced mitochondrial respiration recorded in fibroblasts from FD patients (Lucke et al., 2004), our data indicate that the oxidative phosphorylation capacity could be affected in FD cells and indicate mitomiR dysregulation as a possible underlying mechanism.
Fundamental miRs regulating mitochondrial biogenesis were also affected in FD patients. Indeed, miR-696 can target and downregulate PGC-1α, the master regulator of mitochondrial biogenesis (Aoi et al., 2010), whereas miR-1 is able to modulate the expression of transcripts encoded by the mitochondrial genome (Zhang et al., 2014). Specifically, miR-1 has been reported to enter the mitochondria, where it can stimulate, rather than repress, the translation of specific mitochondrial transcripts, playing a protective role for mitochondria. Consistently, in our analysis, miR-696 was upregulated in FD patients while miR-1 was downregulated, supporting the hypothesis that both mitochondrial and nuclear factors orchestrating the synthesis of mitochondrial components could be compromised in FD patients.
Altogether, our data strongly suggest the implication of mitochondrial dysfunction in FD and shed light on mitomiRs as potential functional players in contributing to aberrant mitochondrial homeostasis. MitomiR dysregulation could represent a new signature of FD underlying the alterations of mitochondrial homeostasis in FD (Fig. 5).

MitomiR dysregulation as signature of FD. MitomiRs are significantly dysregulated in FD. The altered miRs are involved in fundamental mechanisms required for the maintenance of mitochondrial homeostasis, including ETC expression and assembly, mitochondrial metabolism, mitogenesis, mitochondrial oxidative damage and apoptosis. This emphatic pattern could represent a new FD footprint. Cyt-C, Cytochrome C; ETC, electron transport chain; FD, Fabry disease.
Consistent with our results, alterations of mitochondrial function and regulation are common in lysosome storage disorders (de la Mata et al., 2016) but they have not been fully investigated in FD. Moreover, our findings are in agreement with a recent report indicating that miR-184 levels are altered in FD patients (Salamon et al., 2021). Indeed, miR-184 is known to target another gene that is crucial for mitochondrial function, namely Slc25a22 (Morita et al., 2013), a mitochondrial carrier that transports glutamate (Goubert et al., 2017) and asymmetric dimethyl L-arginine (Gambardella et al., 2020b). Our results call for future investigations in this direction, unveiling the essential role of mitochondria as a new frontier in FD research.
The diagnosis of FD is usually obtained by detecting GB3 in urine and plasma and is confirmed by genetic analysis. However, GB3 levels and genetic information often are not reliable prognostic indicators, probably because other GB3-independent mechanisms could contribute to organ damage and dysfunction in FD. Several alternative strategies have been proposed in recent years to monitor FD patients (Simonetta et al., 2020). Although they require further investigation, mitomiRs could represent a new class of biomarkers in FD. Our data show that most mitomiRs don’t seem to be sensitive to currently available FD therapies, suggesting that their dysregulation is probably an intimate mechanism of FD, probably independent from GB3 accumulation (Weidemann et al., 2013; Braun et al., 2019), which is reduced by treatment. In this view, their levels could be extremely useful to monitor the onset of GB3-independent damage, highlighting once again the urgent need for new therapeutic targets, beyond current therapies. Intriguingly, miR-1 levels were instead significantly different between untreated and treated patients. In particular, the treatment seems to restore miR-1 levels in FD patients, indicating this miR as a valuable candidate marker of FD, useful also to monitor the response to therapies. Our research has several limitations. The effect of treatment, for instance, was tested in different patients, as our study had not been designed to compare pre- and post- effects. It is likely that our observations are limited to the Caucasian population, since no other ethnicities were investigated. Also, the relative abundance of mitomiRs should be confirmed in a larger population of FD patients and controls.
In conclusion, our results support the involvement of mitochondria in FD. In this view, mitomiRs could have a pathogenic role to induce the mitochondrial alterations described in FD, and therefore could also represent a potential therapeutic target. Furthermore, miR-1 could be a useful biomarker to monitor the response to treatment in FD.
Acknowledgments
The authors kindly thank the trainees of the Sport Medicine School and of Urgent Medicine School: Dr. Rosita Mottola, Dr. Luca Allocca, Dr. Amos Cocola, Dr. Giada Annarumma, Dr. Pasquale Perrella, Dr. Vincenza Notarangelo, and Dr. Marco Rumolo. The authors thank Dr. Stanislovas S. Jankauskas, Dr. Urna Kansakar, and Dr. Xujun Wang for insightful discussion and technical assistance.
Authorship Contributions
Participated in research design: Gambardella, Santulli, and Iaccarino.
Conducted experiments: Gambardella, Fiordelisi, Sorriento, Cerasuolo, Buonaiuto, Avvisato, Pisani, Varzideh, Riccio.
Contributed new reagents or analytic tools: Gambardella, Fiordelisi, Sorriento, Cerasuolo, Buonaiuto, Avvisato, Pisani, Varzideh, Riccio.
Performed data analysis: Gambardella, Fiordelisi, Santulli, and Iaccarino.
Wrote or contributed to the writing of the manuscript: Gambardella, Sorriento, Pisani, Santulli, Iaccarino.
Footnotes
- Received April 2, 2022.
- Accepted May 25, 2022.
The Iaccarino’s Laboratory is supported in part by PRIN [Grant 2017HTKLRF] and Campania Bioscience [Grant PON03PE_00060_8] (to G.I.), and 54 2020 FRA (to D.S.). The Santulli’s Laboratory is supported in part by the National Institutes of Health National Heart, Lung, and Blood Institute [Grant R01-HL159062], [Grant R01-HL146691], and [Grant T32-HL144456]; National Institute of Diabetes and Digestive and Kidney Diseases [Grant R01-DK123259] and [Grant R01-DK033823] (to G.S.), by the Diabetes Action Research and Education Foundation (to G.S.), and by the Monique Weill-Caulier and Irma T. Hirschl Trusts (to G.S.). F.V. is supported by a postdoctoral fellowship of the American Heart Association [AHA-21POST836407].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
↵1 J.G. and A.F. contributed equally to this work as first authors.
↵2 G.S. and G.I. contributed equally to this work as senior authors.
Abbreviations
- ERT
- enzyme replacement therapy
- ETC
- electron transport chain
- FD
- Fabry disease
- α-GAL A
- α-galactosidase A
- GB3
- globotriaosylceramide
- miR
- microRNA (miRNA)
- mitomiR
- mitochondrial microRNA
- PBMCs
- peripheral blood mononuclear cells
- PGC-1α
- peroxisome proliferator-activated receptor gamma coactivator-1α
- UCP-2
- uncoupling protein-2
- Copyright © 2022 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}