


Abstract
Antipsychotic medications function by blocking postsynaptic dopaminergic signaling in the central nervous system. Dopamine transmission can also be modulated presynaptically by inhibitors of vesicular monoamine transporter 2 (VMAT2), which inhibit loading of dopamine into presynaptic vesicles. Here we investigated the combination of these mechanisms in animal models of schizophrenia and weight gain (a primary side effect of antipsychotics). When dosed alone, the highly selective VMAT2 inhibitor RRR-dihydrotetrabenazine (RRR-DHTBZ, also known as [+]-α-HTBZ) elicited efficacy comparable to conventional antipsychotics in prepulse inhibition and conditioned avoidance models without eliciting weight gain. In combination experiments, synergy was observed: subthreshold doses of RRR-DHTBZ and risperidone or olanzapine produced robust efficacy, and in dose response experiments, RRR-DHTBZ increased the antipsychotic potency in the efficacy models but did not affect weight gain. The combinations did not affect plasma compound concentrations. The synergy is consistent with VMAT2 inhibition blocking the counterproductive presynaptic stimulation of dopamine by antipsychotics. These results suggest a therapeutic strategy of adding a VMAT2 inhibitor to lower the antipsychotic dose and reduce the side effect burden of the antipsychotic while maintaining and potentially enhancing its therapeutic effects.
SIGNIFICANCE STATEMENT Antipsychotics are often necessary and life-changing medications that reduce psychotic symptoms; however, these benefits come with a high side effect burden. This study shows that combining these postsynaptic dopaminergic modulators with a presynaptic dopamine modulator (vesicular monoamine transporter 2 [VMAT2] inhibitor) potentiates efficacy synergistically in animal models of schizophrenia without potentiating weight gain. Our data suggest that adding a VMAT2 inhibitor may be a viable therapeutic strategy for reducing antipsychotic side effects by lowering antipsychotic dose while maintaining therapeutic efficacy.
Introduction
Vesicular monoamine transporters (VMATs) are responsible for packaging monoamines into presynaptic vesicles that subsequently fuse with the presynaptic terminal membrane, thereby releasing those monoamines into the synaptic cleft (Erickson et al., 1996; Erickson and Varoqui, 2000; Eiden, 2020). They belong to the superfamily of solute carrier (SLC) transporters (Lin et al., 2015; Jankovic, 2016). The VMAT2 subtype (SLC18A2) is the exclusive monoamine transporter in the central nervous system (CNS) (Erickson et al., 1996). VMAT2 is the site of action of the CNS drug tetrabenazine, which binds selectively to this subtype. First described and characterized in the 1950s (Pletscher, 1957; Quinn et al., 1959), tetrabenazine attenuates behavioral deficits in a variety of animal models and displays efficacy in a number of monoamine-mediated human disorders (Jankovic, 2016). Tetrabenazine and tetrabenazine analogs are currently in clinical use for treating movement disorders, specifically tardive dyskinesia (TD) and chorea associated with Huntington’s disease. The ability of tetrabenazine to relieve these symptoms is believed to result from blockade of VMAT2 in the presynaptic terminals of striatal dopaminergic neurons, resulting in a reduction of synaptic dopamine levels (Vander Borght et al., 1995; Erickson et al., 1996; Erickson and Varoqui, 2000; Grigoriadis et al., 2017).
Tetrabenazine was originally developed as an antipsychotic, displaying efficacy for relieving the positive symptoms of schizophrenia (Lingjaerde, 1963; Malik and Balkoski, 2007). This effect can be explained by what has been known as “the dopamine hypothesis of schizophrenia” first adopted by van Rossum in 1967, which resulted from the observations that amphetamine, a dopamine releaser, exacerbated psychotic symptoms, whereas reserpine, a dopamine release inhibitor, reduced psychotic symptoms (van Rossum, 1967). Originally synthesized in the 1950s at Hoffmann-La Roche, tetrabenazine was identified as a reversible dopamine release inhibitor (Pletscher, 1957; Quinn et al., 1959). Although a few well controlled clinical trials were conducted in patients with psychosis at the time showing modest effects of tetrabenazine, the coincident development of a class of selective high affinity dopamine receptor antagonist molecules known as the phenothiazines demonstrated superior efficacy in patients with psychosis (Ashcroft et al., 1961). Today, the most commonly used antipsychotics are still those that block central D2 dopamine receptors. This mode of treatment is the primary mechanism of action of conventional antipsychotics such as haloperidol, risperidone, and olanzapine (Seeman et al., 1976; Kapur et al., 1999). The efficacy of tetrabenazine as an antipsychotic was likely a result of reduced D2 dopamine receptor signaling resulting from reductions in synaptic dopamine content after inhibition of VMAT2 activity. If this is the case, then two independent mechanisms in the relevant dopaminergic signaling pathways can be targeted for treating the positive symptoms of schizophrenia with the possibility of reducing the requirement for high doses of the individual compounds working through the two mechanisms: postsynaptic signaling (dopamine D2 receptor blockade) and reduced presynaptic dopamine packaging (VMAT2).
The aim of this study was to use well established animal models of schizophrenia and antipsychotic efficacy to determine whether targeting the presynaptic pathway (VMAT2 inhibition) concurrently with the postsynaptic mechanisms (dopamine receptor blockade) results in synergistic efficacy. In an effort to maintain unambiguous pharmacological effects for these synergy studies, we selected the single isomer RRR-dihydrotetrabenazine (RRR-DHTBZ, also known as [+]-α-HTBZ)—the most active of the four isomers generated from tetrabenazine and the only isomer generated from valbenazine—which has exquisite specificity and selectivity for VMAT2 and is devoid of any other off-target effects (Grigoriadis et al., 2017). Since the other isomers of tetrabenazine have multiple pharmacologic activities at monoamine receptors, including blockade of D2 dopamine receptors, the selective nature of the RRR-DHTBZ isomer offers the clearest interpretation of data associated with pre- and postsynaptic activity. Efficacy of VMAT2 inhibition alone in animal models of psychosis was first confirmed using RRR-DHTBZ in prepulse inhibition (PPI) and conditioned avoidance assays and compared with efficacy of antipsychotic-mediated postsynaptic dopamine inhibition. Once these independent baselines of efficacy were determined in a dose-dependent manner for each compound, the potential synergy of VMAT2 inhibition and D2-receptor blocking antipsychotics was evaluated by combining individual subthreshold doses of RRR-DHTBZ and risperidone or olanzapine in the conditioned avoidance assay. We further explored the effects of combining RRR-DHTBZ and olanzapine in an antipsychotic-induced weight gain model, a surrogate for the metabolic syndrome side effects of conventional antipsychotics, in an attempt to determine the extent to which conclusions could be drawn on the translatability of these preclinical in vivo effects.
Methods
Drug Compounds and Dosing Information
Haloperidol, olanzapine, and risperidone (all free base) were obtained from Millipore-Sigma (St. Louis, MO). RRR-DHTBZ (camphorsulfonic acid salt) and tetrabenazine hydrochloride were synthesized in-house at Neurocrine Biosciences, Inc. (San Diego, CA). For the behavioral tests, RRR-DHTBZ and tetrabenazine were dissolved in water and administered orally in a dose volume of 1 ml/kg; olanzapine, risperidone, and haloperidol were dissolved in 10% DMSO in water and administered intraperitoneally in a volume of 1 ml/kg. For the weight gain studies, RRR-DHTBZ was dissolved in water and administered orally in a volume of 3 ml/kg; olanzapine was dissolved in 1% methylcellulose and administered intraperitoneally in a volume of 3 ml/kg.
Animals
All experimental procedures were approved by the Institutional Animal Care and Use Committee of Neurocrine Biosciences, Inc. and were conducted in accordance with the Guide for the Care and Use of Laboratory Animals. Upon receipt, all animals remained housed in groups and acclimated to the colony room for at least 1 week prior to testing. During the period of acclimation, animals were examined on a regular basis, handled, and weighed to assure adequate health and suitability. Animals were maintained on a 12:12, light:dark cycle for the duration of the experiments, and all behavioral assessments were conducted during the light phase. Room temperature was maintained between 20 and 23°C with a relative humidity between 30% and 70%. Chow and water were provided ad libitum for the duration of each study. In each study, animals were randomly assigned to treatment groups that were then balanced across the testing day to account for any potential differences in circadian response.
PPI of Startle
RRR-DHTBZ was tested in the PPI of startle task, which is used as a measure of sensorimotor gating. The acoustic startle measures an unconditioned reflex response to external auditory stimulation. PPI, consisting of an inhibited startle response (reduction in amplitude) to an auditory stimulation after the presentation of a weak auditory stimulus, or prepulse, has been used as a tool for the assessment of deficiencies in sensorimotor gating, such as those seen in schizophrenia. It is well established that administration of either typical or atypical antipsychotic compounds increases the percentage of PPI typically displayed by C57BL/6J mice (Ouagazzal et al., 2001; Ralph and Caine, 2005). A pharmacological model of PPI was not used because the effectiveness of test compounds can be dependent on the mechanism of the compound used to induce PPI (Swerdlow et al., 2008).
Male C57BL/6J mice from Jackson Laboratories (Bar Harbor, ME) were used in this study. Mice were received at 7 weeks of age, and upon receipt mice were assigned unique identification numbers (tail marked) and group housed in OPTImice cages. The number of animals in each treatment group was between 9 and 11. C57BL/6J mice were pretreated with vehicle or test compound as described in Drug Compounds above and placed into holding cages prior to testing starting 30 minutes postdose. Mice were placed in the PPI chambers (Med Associates, St. Albans, VT) for a 5-minute session of white noise (70 dB) habituation. After the acclimation period, the test session was automatically initiated. The session started with a habituation block of six presentations of the startle stimulus alone followed by 10 PPI blocks of six different types of trials. Trial types were as follows: null (no stimuli); startle (120 dB); startle plus prepulse (4, 8, and 12 dB over background noise, which is 74, 78, or 82 dB); and prepulse alone (82 dB). Trial types were presented at random within each block. Each trial started with a 50-millisecond null period during which baseline movements were recorded. There was a subsequent 20-millisecond period during which prepulse stimuli were presented and responses to the prepulse measured. After a further 100 milliseconds, the startle stimuli were presented for 40 milliseconds and responses recorded for 100 milliseconds from startle onset. Responses were sampled every millisecond. The intertrial interval was variable with an average of 15 seconds (range from 10 to 20 seconds). In startle alone trials, the basic auditory startle was measured; in prepulse plus startle trials, the amount of inhibition of the normal startle was determined and expressed as a percentage of the basic startle response (from startle alone trials), excluding the startle response of the first habituation block.
Conditioned Avoidance Response
The conditioned avoidance response (CAR) assay has been shown to be a very reliable animal model for screening antipsychotic drugs (Wadenberg and Hicks, 1999). In the CAR paradigm, an animal is trained to respond to a conditioned stimulus (auditory and visual) by negative reinforcement (foot shock). Numerous studies have shown that typical and atypical antipsychotic drugs selectively suppress a trained CAR, thus making it an ideal assay to screen potential antipsychotic compounds (Wadenberg and Hicks, 1999). The experimental paradigm and behavioral scoring method used was previously described (Kalinichev et al., 2013) and is shown in Fig. 1.
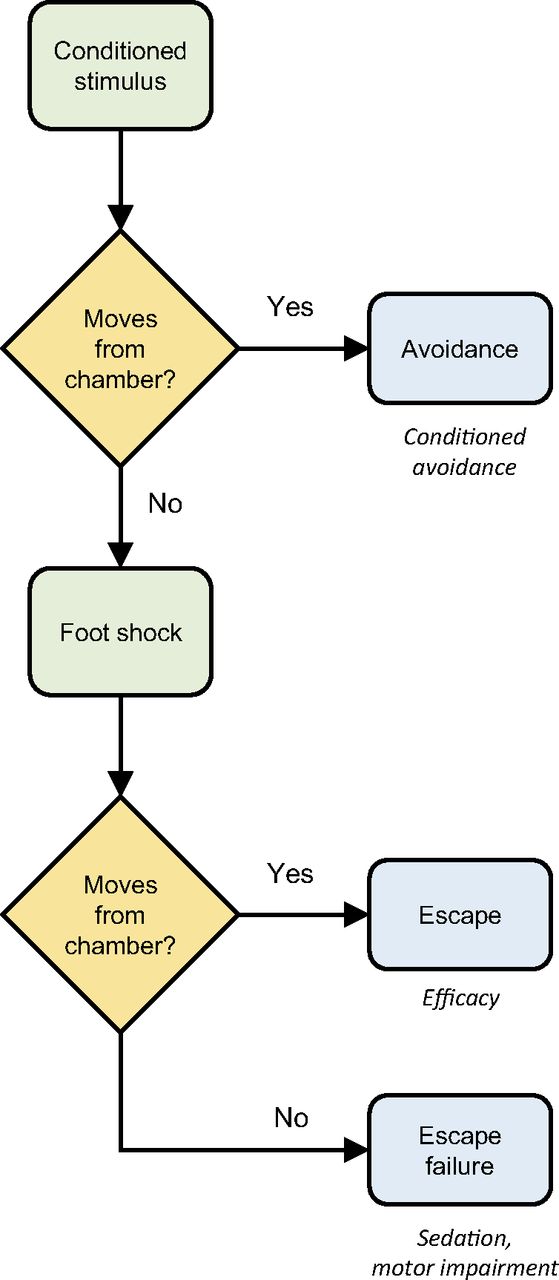
CAR model experimental paradigm and response quantification. The scoring system is that used in Kalinichev et al. (2013). Rats were trained in a two-chamber shuttle box to escape to the second chamber on presentation of the conditioned stimulus (a 10-second pulse of white noise at 80 dB) to avoid a mild foot shock. In the experiment, test compound was administered to the rat, and after the appropriate pretreatment time the conditioned stimulus was applied. The rat could either escape to the second compartment or remain in the test compartment. The former represents the conditioned avoidance response (and would be observed for an untreated rat). The latter represents inhibition of avoidance, which could in principle be due either to inhibition of the conditioned response by the test compound or inhibition of movement (typically by sedation or gross motor inhibition). These mechanisms can be discriminated in the second stage of the experiment. A mild foot shock is applied to the test chamber after presentation of the white noise pulse. If the rat moves to the second chamber, it is assumed that the inhibition of avoidance was a result of inhibition of the conditioned response. This represents efficacy of the drug for inhibiting conditioned avoidance and was scored as an escape. If the rat failed to move, this implicated failure of movement and was scored as an escape failure. Twenty of these trials were performed for each rat and the mean number of escapes and escape failures quantified. The maximum score is therefore 20.
Male Wistar rats from Envigo (Indianapolis, IN) were used in the study. Upon receipt, rats were assigned unique identification numbers and were group housed with three rats per cage in polycarbonate cages with microisolator filter tops. On each training day, experimental animals were acclimated to the test room for at least 30 minutes prior to test. Rats were placed in the CAR two-way shuttle box, and the training period of 20 trials ensued. Each test trial consisted of a conditioned stimulus (i.e., 10-second presentation of an 80 dB white noise) immediately followed by a scrambled 0.6 mA foot shock lasting up to 20 seconds. The intertrial interval ranged from 20 to 60 seconds. Rats learned to avoid foot shocks by moving from one compartment to the other when the cue was presented, and this was recorded as an avoidance response. If the animal failed to move after cue presentation, it received the foot shock, during which it could then cross compartments to escape the full shock duration. If the rat failed to leave the compartment, it endured the full 20-second foot shock and an escape failure was recorded. After 3 to 4 weeks of training in the CAR chambers, rats that passed the testing criterion of performing 16 to 20 avoidance responses while also successfully escaping the foot shock for 3 days in a row were included in the study.
CAR testing then followed the same paradigm as CAR training with the addition of test compound dosing. The number of animals in each treatment group was between 7 and 12. In some cases, data from two treatment groups were combined, giving a final group size of up to 24 animals. On the test day, trained rats were dosed with vehicle or compound as described in Drug Compounds above. When RRR-DHTBZ and antipsychotic (risperidone or olanzapine) formulations were administered in combination, they were dosed sequentially, one immediately after the other, and rats were returned to their home cage. After the 30-minute pretreatment period, the animal was placed into the two-compartment shuttle box. Animals were subsequently exposed to a series of conditioned stimuli. If the rat moved to the second compartment upon presentation of the conditioned stimulus, this was scored as an avoidance (Fig. 1). As mentioned above, an animal not leaving the chamber represents inhibition of avoidance and is indicative of compound efficacy in the model. Inhibition of avoidance can result from either a true inhibition of the conditioned behavior or from a compound-induced inability of the animal to move from the chamber; however, these possibilities were differentiated by applying a mild foot shock to the test chamber. An animal leaving the chamber subsequent to the initiation of the foot shock (i.e., an escape) displayed a true inhibition of conditioned avoidance, as the animal was physically capable of avoiding the shock (Fig. 1). Conversely, an animal not leaving the chamber upon initiation of the foot shock represents a compound-induced sedation or inhibition of movement rather than inhibition of the conditioned behavior and was scored as an escape failure (Fig. 1). Therefore, a potential antipsychotic compound will significantly decrease percentage of avoidance at a dose that does not significantly increase the number of escape failures. For inhibitors of dopaminergic signaling, escape failure likely results from sedation at lower doses and gross motor impairment at higher doses (Arnt, 1982; Wadenberg et al., 2001).
Catalepsy Bar Test
To further assess the potential for gross motor impairment, a separate cohort of animals was tested for compound-induced cataleptic effects using the bar test. Male Wistar rats from Envigo (Indianapolis, IN), age-matched to the rats used in the CAR experiments, were used in this study. On the day of testing, rats were brought to the experimental room for at least 1 hour to acclimate. Rats were dosed with vehicle or test compound as described in Drug Compounds above, and 30 minutes later cataleptic behavior was assessed. At the start of the assay, the front paws of the experimental subject were placed on a horizontal metal bar raised 6 inches above a Plexiglas platform, and time was recorded for up to 60 seconds per trial at each observational time point. The assessment ended when the animal’s front paws returned to the platform or after 60 seconds. The test was repeated in triplicate, and the average of the three trials is reported as the intensity index of catalepsy. The number of animals in each treatment group was four or five.
Ex Vivo VMAT2 Occupancy Measurement
Occupancy of VMAT2 by RRR-DHTBZ in rats was assessed ex vivo using homogenates of whole striatum. Male Wistar rats (satellite groups to those used in the CAR studies) were dosed with RRR-DHTBZ as described in Drug Compounds above (n = 4 or 5 per treatment group). Thirty minutes later, animals were sacrificed, and whole brains were collected and then frozen on aluminum foil on dry ice. On the day of assay, brains were partially thawed on wet ice, and whole striatum was dissected and placed in 10 ml assay buffer in a 15 ml centrifuge tube on ice. Assay buffer comprised Dulbecco’s phosphate-buffered saline (DPBS): 1.5 mM KH2PO4, 8.1 mM Na2HPO4, 2.7 mM KCl, and 138 mM NaCl (Thermo Fisher Scientific, Waltham, MA), supplemented with 10 mM MgCl2, 2 mM ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid, pH 7.4, with NaOH. Assay buffer was maintained on ice throughout the preparation procedure. Multiple samples (up to 48) were homogenized simultaneously using an Autogizer (Tomtec Inc., Hamden, CT). Tubes containing the samples were maintained in an ice bath throughout the homogenization procedure. The tissues were homogenized at 30,000 rpm for 10 seconds. The samples were then centrifuged at 45,000 × g for 20 minutes at 4°C. The supernatant was discarded, and 2 ml of fresh assay buffer was added. The pellet was dislodged by vortexing, and an additional 8 ml of buffer was added. The homogenization and centrifugation steps were then repeated once. The final pellet was suspended in 2 ml assay buffer and protein concentration determined using the Coomassie method (Thermo Fisher Scientific) using bovine serum albumin as the standard. The sample volume was then adjusted to give a protein concentration of 400 μg/ml.
Occupancy of VMAT2 in the homogenate was then determined using a radioligand binding assay; RRR-DHTBZ, which binds to VMAT2, administered in vivo will inhibit binding of a VMAT2 radioligand to the transporter. The radioligand used was [3H]-RRR-DHTBZ labeled with C3H3 at the 9 O-methyl position, custom-synthesized by Pharmeron (Irvine, CA) (formerly Quotient Bioresearch) to a specific activity of 75 Ci/mmol. The radioligand concentration used was 3 nM, and the dissociation constant (Kd) was 0.6 nM. The assay was performed as described in Grigoriadis et al. (2017). Binding was measured over 16 time points and the data analyzed to determine occupancy as described in Malany et al. (2009). The maximal occupancy obtainable in the assay was approximately 80%, determined using maximally occupying doses of RRR-DHTBZ (1 mg/kg), suggesting a pool of VMAT2 accessible in the homogenate in the binding assay but not in the intact tissue in the animal. Occupancy values were thus normalized to the maximal obtainable occupancy.
Measurement of Weight Gain in Rats Maintained on a High-Fat Diet
It has been consistently demonstrated that repeated olanzapine treatment in rats elicits gender-specific increases in body weight compared with vehicle-treated counterparts (Castellani et al., 2019), with females regularly displaying treatment-induced body weight increases not always observed in males. Therefore, although males were used in the behavioral screening models, females were used for the body weight assessment studies as they represent a more translational model of human weight gain in response to olanzapine treatment. Female Sprague Dawley rats from Charles River (Margate, Kent, UK) were individually housed in polypropylene cages with metal grid floors at a temperature of 21 ± 4°C and 55% ± 20% humidity. Animals were maintained on a reverse phase light-dark cycle (lights off for 8 hours from 1000 to 1800 hours), during which time the room was illuminated by red light. Animals had free access to a high-fat powdered diet (VRF1, Special Diet Services, Witham, Essex, UK, plus 20% lard) and tap water at all times. The diet was contained in glass feeding jars with aluminum lids (Solmedia Laboratory Suppliers, Romford, Essex, UK). Each lid had a 3–4 cm hole cut in it to allow access to the food. Animals were accustomed to these conditions for at least 2 weeks before experimentation.
Animals underwent a 3-day baseline run-in period, during which time all rats were dosed once a day with vehicle. Animals were weighed at the time of dosing. Toward the end of this baseline period, animals were weighed (to the nearest 0.1 g using an electronic top-pan balance) and allocated into eight weight-matched treatment groups by a statistician. Rats were then dosed for 14 days with vehicle, olanzapine, or RRR-DHTBZ as described in Drug Compounds above. When olanzapine and RRR-DHTBZ were administered in combination, the latter was administered as soon as possible after the former. Dosing began at approximately 0845 hours each day (referred to as “0 hours”) so that the midpoint of dosing would coincide the time of lights out. This strategy was taken to maximize the impact of olanzapine on food intake. Rats were weighed to the nearest 0.1 g every day at the 0-hour timepoint.
Plasma Pharmacokinetics
For the CAR experiments, plasma compound pharmacokinetics were assessed in age-matched satellite groups of male Wistar rats, and blood was collected via closed cardiac puncture under isoflurane anesthetic. Compounds were administered as described in Drug Compounds above, and blood was collected 30 minutes later. Whole blood was collected into K3-EDTA–coated vials and was then separated via centrifugation at 4°C at 10,000 rpm followed by transfer to individually labeled sample vials that were flash frozen in liquid nitrogen and stored at −80°C. The number of satellite animals in each treatment group was four or five. For the weight gain studies, on the day after the last weight measurement, whole blood was collected by cardiac puncture under CO2 narcosis.
RRR-DHTBZ was quantified using liquid chromatography in tandem with a mass spectrometric detector (LC-MS/MS). The plasma calibration standards were prepared by a 12-point dilution series, from 2500 to 0.5 ng/ml. The lower limit of quantification in the assay was 0.5 ng/ml. The plasma extraction procedure involved a protein precipitation extraction from 50 μl of calibration standards or study plasma samples with 175 μl acetonitrile containing 200 ng/ml internal standard (D6-RRR-DHTBZ). Samples were vortexed and centrifuged, and 75 μl of the supernatant was transferred to the sample injection plate and diluted with 225 μl water for LC-MS/MS analysis.
Statistics
PPI, CAR, Catalepsy, and Plasma Concentrations
Single-factor analysis of variance (ANOVA) was used to compare effects of different treatments in the PPI studies. In these experiments, the treatment groups compared were RRR-DHTBZ (multiple doses), tetrabenazine (multiple doses), the antipsychotic haloperidol (single dose), and the respective vehicles. Multiple comparisons were performed using Tukey’s multiple comparison post hoc test. This analysis was conducted using Prism 7.0 (GraphPad Software, Inc., La Jolla, CA) and the results shown in Supplemental Tables 1 and 2. This analysis was also performed for CAR assay data (escapes and escape failures) for the effect of multiple doses of RRR-DHTBZ and vehicle (Supplemental Tables 3 and 4).
Conditioned avoidance treatment-combination experiments (i.e., experiments incorporating antipsychotic with and without RRR-DHTBZ) were designed to enable pairwise comparisons between the presence or absence of RRR-DHTBZ treatment groups and comparisons between doses of the antipsychotic (or a single dose and vehicle in one case for risperidone, Supplemental Table 5). All the doses of antipsychotic tested alone were also treated with RRR-DHTBZ in the same experiment. This design enabled two-factor ANOVA and post hoc testing using multiple comparisons. In all cases, an individual experiment was conducted on the same cohort of animals. In many cases, the treatment groups were split across two test sessions, necessitated by the large number of treatment groups. The session details are given in the supplemental material. The ANOVA compared the effect of the different antipsychotic doses and compared the effect of the antipsychotic in the absence or presence of RRR-DHTBZ. The interaction statistic of the ANOVA tested whether the effect of RRR-DHTBZ was dependent on the antipsychotic dose. Tukey’s multiple comparison post hoc test was then run to compare all treatment groups with one another. This design enabled multiple effects to be compared in the same experiment, specifically 1) the effect of RRR-DHTBZ at each dose of antipsychotic, 2) the effect of RRR-DHTBZ in the absence of antipsychotic, and 3) the effect of the different doses of antipsychotic and vehicle in the absence of RRR-DHTBZ. The post hoc test reported significance at the P < 0.05, P < 0.01, and P < 0.001 levels, and these results are given in the supplemental material. Statistical analysis was performed using Prism 7.0. The effects tested in the conditioned avoidance test were the number of escapes and number of escape failures. If the highest number of escape failures among the treatment groups in an experiment was low (< two trials), the statistical test on escape failures was not performed.
The same two-factor ANOVA design was applied to the treatment-combination catalepsy bar test (time on bar data). The test was not performed if the highest effect among the treatment groups was small (time on bar <10 seconds). In addition, plasma antipsychotic concentrations in the satellite groups of animals dosed with compound were compared using the two-factor ANOVA design. For the RRR-DHTBZ concentration, single-factor ANOVA was performed comparing the concentration of RRR-DHTBZ across the antipsychotic treatment groups (different antipsychotic doses and antipsychotic vehicle). In all cases, the P value from this test was >0.05, so no post-test comparison between treatment groups was performed.
Outliers were assessed by calculating the standard deviation of each treatment group within an individual experiment. If a value was outside of the range of the mean ± 2 S.D., it was excluded.
Weight Gain Statistics
Statistical reports for the weight gain study are provided in the supplemental material (Supplemental Tables 12 and 13). The statistical methods assume that data are normally distributed with equal variance in the groups. Body weight and body weight gain were analyzed by two-way analysis of covariance with treatment and cohort as factors and body weight on day 1 as the covariate. Multiple comparisons were conducted as follows: In the experiment RS1046 (Fig. 8), multiple comparisons were by Williams’ test to compare RRR-DHTBZ to vehicle, separate Dunnett’s tests to compare olanzapine and the combination to vehicle, Williams’ test to compare the combination to RRR-DHTBZ 0.15 mg/kg, and the multiple t test to compare the combination to the same dose of olanzapine. In the experiment RS1262, multiple comparisons were by separate Williams’ tests to compare olanzapine and RRR-DHTBZ to vehicle, Dunnett’s test to compare the combination to vehicle, Williams’ test to compare the combination to olanzapine 0.3 mg/kg, and the multiple t test to compare the combination to the same dose of RRR-DHTBZ. The rationale for applying the Williams’ and Dunnett’s tests was as follows: Williams’ test was used to compare increasing doses of a compound to vehicle control assuming that the dose-response relationship is monotonic (i.e., increasing the dose causes increased response). This assumption is reasonable for RRR-DHTBZ but not for olanzapine, which appears to have a bell-shaped dose-response relationship (e.g., 3 mg/kg does not have as big of an effect as 1.2 mg/kg) (Fig. 8). Williams’ test was also appropriate to compare RRR-DHTBZ + olanzapine to RRR-DHTBZ 0.15 mg/kg because, again, it is reasonable to assume a monotonic dose-response relationship. Dunnett’s test was used to compare several groups to a single group, making no assumptions about the dose-response relationship. It was also used to compare the combination to vehicle because a monotonic dose-response relationship would not be expected if olanzapine reversed the effect of RRR-DHTBZ.
Results
Efficacy of the Selective VMAT2 Inhibitor RRR-DHTBZ in Animal Models of Schizophrenia
Tetrabenazine has previously demonstrated efficacy in animal models of schizophrenia (Heise and Boff, 1960; Levison and Freedman, 1967; Bocknik et al., 1968; Kuribara and Tadokoro, 1983; Rigdon and Weatherspoon, 1992). These effects were originally presumed to be mediated solely by inhibition of VMAT2 activity. However, some of the metabolites of tetrabenazine also inhibit other receptors that modulate antipsychotic activity, albeit at lower potency. For example, the R,S,S isomer of DHTBZ binds the D2 receptor with a Ki of 53 nM and 5HT7 receptor with a Ki of 6 nM (Grigoriadis et al., 2017). Here we confirmed that VMAT2 inhibition can elicit antipsychotic-like efficacy using the highly selective tetrabenazine metabolite RRR-DHTBZ (VMAT2 Ki of 2 nM, >1000-fold selective in a broad-panel selectivity screen, including monoamine receptors) (Grigoriadis et al., 2017). The compound was tested in two models: PPI of startle, a model of sensorimotor gating (Geyer et al., 2001); and CAR, a model of reinforcement learning (Wadenberg and Hicks, 1999; Wadenberg, 2010). PPI in rodent models has a direct translatability to humans in that deficits in PPI are observed in schizophrenic individuals, and drugs that are effective in this model are effective antipsychotics in humans (Geyer et al., 2001; Kumari and Sharma, 2002; Swerdlow et al., 2008). The CAR model also has excellent predictive validity. All clinically effective antipsychotics, but not most other classes of drugs, are effective in suppressing CAR at clinically relevant doses (Arnt, 1982; Olsen et al., 2008; Wadenberg, 2010). In addition, pathways mediating CAR behavior have been invoked in the efficacy of antipsychotics for attenuating the positive symptoms of schizophrenia, notably the mesolimbic dopamine pathway (Wadenberg et al., 1990).
Prepulse Inhibition
RRR-DHTBZ was tested in a genetic PPI model. The inbred C57BL/6J mouse strain displays a reduced level of PPI of acoustic startle compared with the parental C57BL line (Ralph et al., 2001). This reduction can be reversed by antipsychotics (i.e., the compounds increase PPI in C57BL/6J mice) (Ouagazzal et al., 2001; Ralph and Caine, 2005). In the genetic PPI model, RRR-DHTBZ increased PPI in C57BL/6J mice in a dose-dependent manner at stimulus intensities of 74, 78, and 82 dB, with a statistically significant effect observed at 10 mg/kg (Fig. 2 for 74 dB; Supplemental Tables 1 and 2 for 78 and 82 dB). Likewise, tetrabenazine elicited a dose-dependent increase of PPI, with a statistically significant effect observed at 30 mg/kg (Fig. 2; Supplemental Table 1). The increase of PPI observed at the highest doses of RRR-DHTBZ and tetrabenazine was not significantly different from the effect of haloperidol alone, used as a positive control (Fig. 2; Supplemental Tables 1 and 2). These results indicate that selective inhibition of VMAT2 elicits efficacy in the PPI model equivalent to the antipsychotic haloperidol.
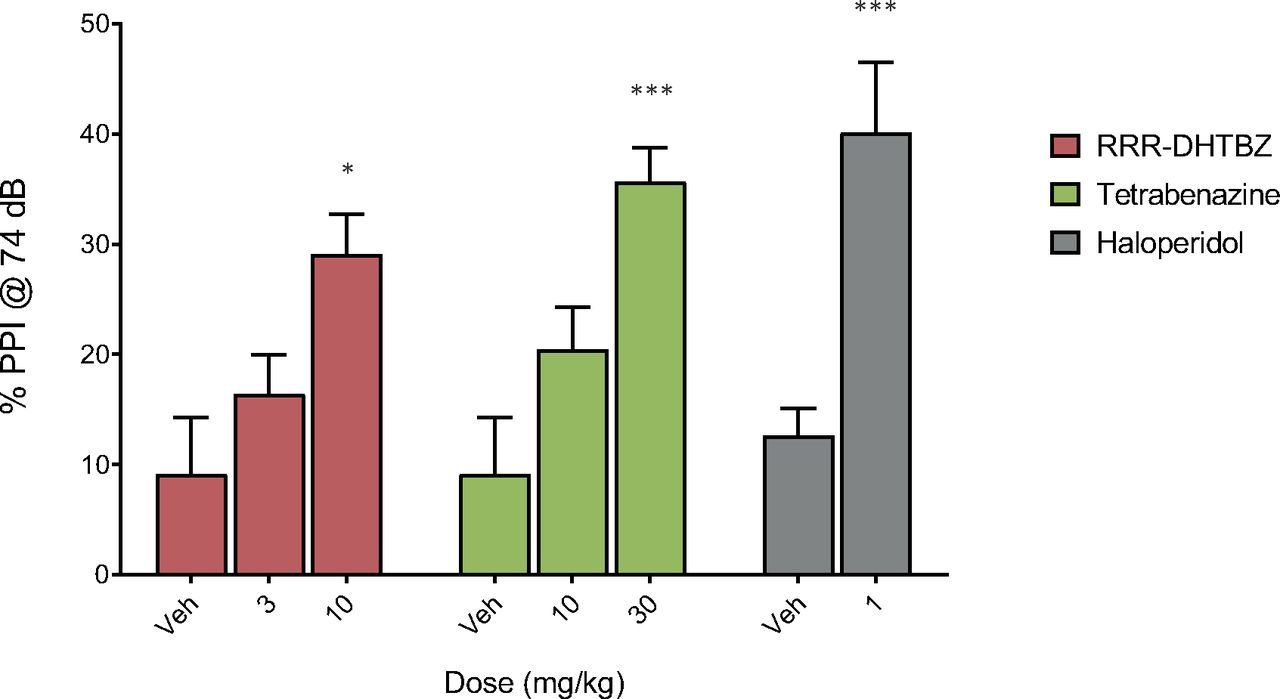
VMAT2 inhibition reverses PPI deficit in C57BL/6J mice. This inbred mouse strain displays a deficit in PPI of acoustic startle. The compounds tested were the selective VMAT2 inhibitor RRR-DHTBZ (also known as [+]-α-HTBZ), tetrabenazine, and haloperidol (the positive control) administered 30 minutes prior to measurement of PPI. PPI was measured at a stimulus intensity of 74 dB (shown in figure) and 78 and 82 dB (data in Supplemental Table 1). The experiments were performed twice with similar results (Supplemental Tables 1 and 2) except the experiment for tetrabenazine, which was performed once (Supplemental Table 1). Data in the figure are from Supplemental Table 1. The number of animals in each treatment group was 10 or 11. Statistical significance of PPI between treatment groups was tested by single-factor ANOVA followed by Tukey’s multiple comparison post hoc test. The asterisks denote significant difference between test-compound treatment and the corresponding vehicle. *P < 0.05, ***P < 0.001.
Conditioned Avoidance
The effect of RRR-DHTBZ on conditioned avoidance was tested in rats. The experimental paradigm and behavioral scoring methods are shown in Fig. 1. CAR scoring was used as previously described (Kalinichev et al., 2013). On the test day, rats previously trained in a two-compartment shuttle box were exposed to the conditioned stimulus (a 10-second pulse of white noise at 80 dB). If the rat moved to the second compartment upon presentation of the conditioned stimulus, this was scored as an avoidance (Fig. 1). An animal not leaving the chamber represents inhibition of avoidance. Inhibition of avoidance can result from inhibition of the conditioned behavior (antipsychotic efficacy) or impairment of the ability of the animal to move from the chamber (sedation/motor impairment). These possibilities were differentiated by applying a mild foot shock to the test chamber (Fig. 1). An animal leaving the chamber subsequent to initiation of the foot shock represents inhibition of conditioned avoidance, the behavior of interest. This event represents efficacy of the test compound in the model and was scored as an escape (Fig. 1; Kalinichev et al., 2013). The animal not leaving the chamber represents an inhibition of movement rather than inhibition of the conditioned behavior. This result was scored as an escape failure (Fig. 1). For inhibitors of dopaminergic signaling, escape failure likely results from sedation at lower doses and gross motor impairment at higher doses (Arnt, 1982; Wadenberg et al., 2001). To further assess the potential for gross motor impairment, a separate cohort of animals was tested by measuring catalepsy using the bar test.
RRR-DHTBZ produced a dose-dependent suppression of conditioned avoidance, manifested as an increase in the number of escapes (Fig. 3; Supplemental Tables 3 and 4; Table 1). The lowest dose eliciting a statistically significant effect was 0.3 mg/kg. At this dose, there was no detectable catalepsy (Supplemental Table 4) and there were no escape failures (Supplemental Tables 3 and 4). The next highest dose in the experiments, 0.7 mg/kg (Supplemental Table 3) or 0.6 mg/kg (Supplemental Table 4), elicited a greater number of escapes, indicating an increased suppression of conditioned avoidance. At these doses, a significant number of escape failures were recorded (Supplemental Tables 3 and 4; Table 1), consistent with mild sedation contributing to the overall suppression of avoidance. No significant catalepsy was detected at these doses (Supplemental Table 4). At a higher dose (1.2 mg/kg), catalepsy was observed (Supplemental Table 4; Table 1), indicating gross motor impairment, which is anticipated for a high dose of an agent that suppresses dopaminergic transmission. A high dose of risperidone (1 mg/kg) also resulted in catalepsy (Supplemental Table 7; Table 1), which is typically observed in rats treated with high doses of antipsychotics (Wadenberg et al., 2001). These findings imply that specific inhibition of VMAT2 can suppress conditioned avoidance responding to the same extent as known antipsychotics.

VMAT2 inhibition suppresses conditioned avoidance responding in rats. The compounds tested were the selective VMAT2 inhibitor RRR-DHTBZ, tetrabenazine, and the conventional antipsychotics risperidone and olanzapine. Inhibition of conditioned avoidance was quantified as the number of escapes, as described in Fig. 1. Data in the figure are from Experiment 1 (Supplemental Table 3) except the data for olanzapine, which are from Experiment 3 (Supplemental Table 8). The experiments were performed multiple times with similar results (twice for RRR-DHTBZ, Supplemental Tables 3 and 4; three times for risperidone, Supplemental Tables 3, 6, and 7; twice for olanzapine, Supplemental Tables 8 and 11) except for tetrabenazine, which was performed once (Supplemental Table 3). The number of animals in each group was between 9 and 24. Statistical significance of the number of escapes between treatment groups was tested by single-factor ANOVA followed by Tukey’s multiple comparison post hoc test. The asterisks denote significant difference between test-compound treatment and the corresponding vehicle. *P < 0.05, ***P < 0.001.
Minimum effective plasma concentration and dose for dopaminergic behaviors and weight gain in rats
The minimum plasma concentration and dose required to elicit statistically significant activity was determined (see supplementary tables and footnotes below). CAR escapes, escape failures, and catalepsy were measured in male Wistar rats 30 minutes after a single dose of the compounds. Weight gain was measured in female Sprague Dawley rats and the plasma concentration measured 30 minutes after the final dose of a once-daily dosing regimen for 15 days. Values indicate plasma concentration in ng/ml, with dose (mg/kg) in parentheses.
The efficacy of RRR-DHTBZ in the CAR model was compared with tetrabenazine and with conventional antipsychotics. Tetrabenazine significantly increased the number of escapes at 3 mg/kg (Fig. 3; Supplemental Table 3), consistent with previous studies (Heise and Boff, 1960; Bocknik et al., 1968; Kuribara and Tadokoro, 1983). The magnitude of the effect was not significantly different from that of 0.7 mg/kg RRR-DHTBZ (Supplemental Table 3A). The plasma concentration of RRR-DHTBZ resulting from administration of 3 mg/kg tetrabenazine (34 ng/ml) was similar to that resulting from administration of 0.7 mg/kg RRR-DHTBZ (52 ng/ml, Supplemental Table 3). These findings are consistent with the hypothesis that the effect of tetrabenazine in the CAR model results primarily from inhibition of VMAT2 activity. The conventional antipsychotics risperidone and olanzapine increased the number of escapes at a historically standard dose of 0.3 mg/kg for risperidone (Fig. 3; Supplemental Tables 3, 6, and 7; Table 1) and 3 mg/kg for olanzapine (Fig. 3; Supplemental Tables 8 and 11; Table 1) (Wadenberg et al., 2001). The magnitude of the effect was similar to that for 0.7 mg/kg RRR-DHTBZ (Fig. 3). (The effects of 0.3 mg/kg risperidone and 0.7 mg/kg RRR-DHTBZ were compared directly in one experiment and were not found to be significantly different [Supplemental Table 3A]). These findings indicate that inhibition of VMAT2 activity can be as effective as a conventional antipsychotic in inhibiting conditioned avoidance.
Relationship between RRR-DHTBZ Plasma Concentration, Inhibition of Conditioned Avoidance, and VMAT2 Occupancy
The pharmacokinetic/pharmacodynamic relationships of conventional antipsychotics for inhibiting conditioned avoidance have been studied in detail. The effective plasma concentration for inhibiting conditioned avoidance closely matches the therapeutic plasma concentration for treating schizophrenia (Olsen et al., 2008). D2 receptor occupancy of >50% is typically required for both therapeutic efficacy (Farde et al., 1988; Kapur et al., 1999) and for half-maximal effect in CAR (Wadenberg et al., 2001; Olsen et al., 2008). Here we investigated these relationships for VMAT2 inhibition using RRR-DHTBZ.
The plasma concentration-effect relationship of RRR-DHTBZ for suppressing CAR is shown in Fig. 4. Significant suppression of CAR (manifested as an increase of escapes) was observed at a plasma RRR-DHTBZ concentration of 19 ng/ml (Fig. 4; Table 1). The maximal effect for suppressing CAR was observed in range of 100 ng/ml (Fig. 4). These plasma concentrations are in the same range as the therapeutic RRR-DHTBZ plasma concentration in humans for treating TD (20–48 ng/ml) resulting from administration of valbenazine (Skor et al., 2017). VMAT2 occupancy was assessed in a dose-dependent manner by ex vivo binding in the striatum in a satellite group of rats. Significant suppression of CAR was observed at 87% VMAT2 occupancy and above. The dose dependence of the behavioral efficacy demonstrated that there was no significant effect on conditioned avoidance below 59% occupancy (Fig. 4).

Concentration-response for CAR suppression and VMAT2 occupancy by RRR-DHTBZ. The pink bar is the range of VMAT2 occupancy resulting in statistically significant suppression of CAR (87%–100%). CAR suppression was quantified as the number of escapes, as described in Fig. 1 (n = 9 or 10 animals in each treatment group). VMAT2 occupancy was measured in the striatum using an ex vivo occupancy assay (n = 4 or 5). Plasma concentration and VMAT2 occupancy were measured in satellite groups of animals (n = 4 or 5). Each point on the graph is a treatment group from an individual experiment, and the data points from all the experiments are shown on the graph (see Supplemental Tables 3 and 4 for data values). CAR data are included for RRR-DHTBZ doses that did not cause catalepsy.
Window between Dopaminergic Behaviors
Dopaminergic inhibitors manifest numerous behaviors in animal models, and the CAR assay represents one preclinical model displaying high sensitivity to dopamine receptor antagonists (Arnt, 1982). In the CAR assay, antipsychotic compounds elicit an inhibition of a previously trained conditioned avoidance response, a hallmark behavioral signature of both typical and atypical antipsychotic treatment in rats (Fig. 1). It is well documented, however, that antipsychotic compounds can also elicit on-target sedative effects or cataleptic behavior in preclinical models assessing antipsychotic-like activity. Therefore, a meaningful inhibition of the conditioned avoidance response in the CAR assay must be accompanied by the ability of the animal to move and escape the subsequent anticipated foot shock as reflected in a higher number of escape behaviors. If the animals show an inhibited avoidance response but fail to escape the foot shock or if the compound itself elicits cataleptic behavior resulting from too robust of a decrease in striatal dopamine, interpretation of the CAR response is confounded at that dose and resultant exposure. We compared the window between these behaviors for RRR-DHTBZ and for the conventional antipsychotics risperidone and olanzapine. Fig. 5 shows the behavior versus plasma concentration for the three compounds, and Table 1 includes the minimal effective plasma concentrations for each behavioral endpoint.

Window between dopaminergic behaviors in the conditioned avoidance model. Dopaminergic inhibition produces multiple behaviors in the CAR test (Fig. 1). At low doses, inhibition of conditioned avoidance predominates (manifested as an increase in the number of escapes). At higher doses, sedation can occur, evident as an increase in escape failures. Further increases in dose results in gross motor impairment (catalepsy), measured using the bar test. The plasma concentration versus effect for these behaviors was assessed for RRR-DHTBZ (A), risperidone (B), and olanzapine (C). Data for RRR-DHTBZ are from Supplemental Tables 3 and 4, data for risperidone from Supplemental Tables 5–7, and data for olanzapine from Supplemental Tables 8–11, with group sizes of 7–24 animals for escapes and escape failures and 4–5 animals for catalepsy.
The potency of RRR-DHTBZ for inhibiting conditioned avoidance (increasing escapes) was higher than that for eliciting escape failures, which was in turn higher than that for causing catalepsy (Fig. 5), with minimally effective plasma concentrations for the three behaviors of 19, 52, and 400 ng/ml, respectively. This magnitude of the potency difference between the activities was similar to that for risperidone (Fig. 5; Table 1; minimally effective plasma concentrations of 45, 280, and 360 ng/ml for CAR inhibition, escape failures, and catalepsy, respectively). This finding suggests that presynaptic dopaminergic inhibition with RRR-DHTBZ elicits a similar window between dopaminergic behaviors as postsynaptic inhibition with risperidone. For olanzapine, there was minimal apparent difference of the concentration-response relationship between CAR inhibition and escape failures (Fig. 5; Table 1). Catalepsy was not detected at the doses used (3 mg/kg and below) but has been detected at higher doses in other studies (Wadenberg et al., 2001).
Effect of Combining a VMAT2 Inhibitor with a Conventional Antipsychotic on Suppression of Conditioned Avoidance
Synergistic Interaction Revealed by Subthreshold Dose Combination
The results above show that VMAT2 provides an additional site of action and mechanism to the D2 dopamine receptor for efficacy in animal models of schizophrenia. We next examined the effect of combining a VMAT2 inhibitor and a conventional D2-blocking antipsychotic using the CAR model. First, we tested for synergy by examining the effects of combining subthreshold doses of RRR-DHTBZ and risperidone or olanzapine. The threshold doses of these three compounds were established by assessing each compound in a dose-dependent manner. The lowest doses tested producing statistically significant suppression of CAR were 0.3 mg/kg, 0.3 mg/kg, and 3 mg/kg for RRR-DHTBZ, risperidone, and olanzapine, respectively (Supplemental Tables 3, 4, 6, 7, 8, and 11; Table 1).
RRR-DHTBZ at half of the threshold dose (i.e., 0.15 mg/kg) was combined with one-third of the threshold dose (i.e., 0.1 mg/kg) of risperidone. When administered alone, the individual treatments did not significantly suppress CAR (Fig. 6A; Supplemental Tables 5 and 6). However, when combined, a profound and significant suppression of CAR was observed, manifested as an increase in the number of escapes (Fig. 6A; Supplemental Tables 5 and 6). Remarkably, the magnitude of the effect produced by the combination was similar to the effect of the threshold dose of the compounds administered individually: 6.5–7.1 escapes for the combination of 0.15 mg/kg RRR-DHTBZ and 0.1 mg/kg risperidone (Supplemental Tables 5 and 6), 4.8–11.4 escapes for 0.3 mg/kg RRR-DHTBZ (Supplemental Tables 3 and 4), and 7.2–8.0 escapes for 0.3 mg/kg risperidone (Supplemental Tables 6 and 7). The synergistic effect was not due to a drug-drug interaction potentially affecting the plasma concentration of the compounds. RRR-DHTBZ did not significantly affect the plasma concentration of risperidone and vice versa in these experiments (Supplemental Tables 5B, 5C, 6D, and 6E; see legend to Fig. 6 for example). Furthermore, the synergistic effect did not result in the induction of catalepsy at the doses tested (Supplemental Table 6).

Synergy between RRR-DHTBZ and antipsychotics for suppressing CAR at subthreshold doses; risperidone (A) and olanzapine (B). Data in the figure are from the experiment in Supplemental Table 6 for risperidone and 9 for olanzapine. Statistical analysis was performed using two-factor ANOVA followed by Tukey’s multiple comparison post hoc test (Supplemental Table 6A for risperidone and 9A for olanzapine). The compounds dosed alone did not significantly affect the number of escapes compared with vehicle (P > 0.05). The effect of the combination was significantly different from vehicle (asterisks in figure) and from the compounds administered alone (P < 0.001). Escape failure and catalepsy were minimal in all treatment groups. RRR-DHTBZ did not significantly affect the plasma concentration of the antipsychotic or vice versa (Supplemental Table 6, D and E for risperidone and 9, C and D for olanzapine). The experiments were performed twice with similar results (Supplemental Tables 5 and 6 for risperidone and 9 and 10 for olanzapine), with group sizes of between 7 and 24 animals.
Olanzapine at one-fifth of its threshold dose (i.e., 0.6 mg/kg) was tested in combination with RRR-DHTBZ at half of the threshold dose (i.e., 0.15 mg/kg). When administered alone, neither treatment significantly affected CAR (Fig. 6B; Supplemental Tables 9 and 10). When administered together, significant suppression of CAR was again observed, evident as an increase in the number of escapes (Fig. 6B; Supplemental Tables 9 and 10). Similar to risperidone, the magnitude of the effect of the combination (8.6–10.3 escapes, Supplemental Tables 9 and 10) was similar to that of the threshold dose of the compounds administered individually (4.8–11.4 escapes for 0.3 mg/kg RRR-DHTBZ, Supplemental Tables 3 and 4; 8.6–12.3 escapes for 3 mg/kg olanzapine, Supplemental Tables 8 and 11). Once more, the synergistic effect was not due to a nonspecific drug-drug interaction. RRR-DHTBZ did not significantly affect the plasma concentration of olanzapine and vice versa in these experiments (Supplemental Tables 9C, 9D, 10C, and 10D; see legend to Fig. 6 for example). The synergistic effect did not result in the induction of catalepsy at the doses tested (Supplemental Table 10). It is interesting to note that the synergy observed was independent of the antipsychotic used and that both studies reached the maximal efficacy observed with the high dose antipsychotics alone (compare Figs. 5 and 6).
RRR-DHTBZ Increases the Potency of Risperidone and Olanzapine in the CAR Model
The effects of a subthreshold dose of RRR-DHTBZ on the potency of the conventional antipsychotics were tested by combining RRR-DHTBZ with a range of doses of risperidone or olanzapine. Conditioned avoidance suppression (number of escapes) was plotted versus the plasma concentration of the compound (Fig. 7, A and D). RRR-DHTBZ at its subthreshold dose of 0.15 mg/kg increased the potency of both risperidone and olanzapine, evidenced as a clear leftward shift of the response-versus-concentration curve for both antipsychotics (Fig. 7, A and D; Table 1). This finding confirms the synergistic interaction between the two classes of dopaminergic modulator. The effect of the combination on the other dopaminergic behaviors, escape failures, and catalepsy was also tested (Fig. 7, B and E for escape failures; Fig. 7, C and F for catalepsy). RRR-DHTBZ at 0.15 mg/kg did not appear to affect the windows between the behaviors for risperidone or olanzapine; RRR-DHTBZ potentiated escape failures and catalepsy to a similar extent, as it potentiated suppression of conditioned avoidance (Fig. 7; see Table 1 for minimum effective plasma concentrations and doses). This finding was expected since escape failures and catalepsy are both primarily mediated by dopaminergic inhibition for the compounds tested.

CAR suppression, escape failures, and catalepsy versus plasma concentration for combination of conventional antipsychotics with RRR-DHTBZ. The subthreshold dose of RRR-DHTBZ (0.15 mg/kg) was combined with a range of doses of risperidone (A–C) or olanzapine (D–F) and the data plotted versus plasma concentration of the antipsychotic. Suppression of CAR indicates antipsychotic efficacy (A and D), escape failures indicate sedation (B and E), and catalepsy indicates motor impairment (C and F). CAR suppression (number of escapes) and escape failures were scored as described in Fig. 1, and catalepsy was assessed using the bar test. The plasma concentration for risperidone represents the sum of the concentrations of risperidone and its fully active metabolite 9-OH-risperidone. Each point on the graph is a treatment group from an individual experiment, and the data points from all of the experiments are shown on the graph (see Supplemental Tables 5–7 for risperidone data and 8–11 for olanzapine data), with group sizes of 7–24 animals for escapes and escape failures and 4–5 animals for catalepsy.
Weight Gain
The results above indicate that VMAT2 inhibition enhances the efficacy of the antipsychotics in behavioral models predictive of therapeutic efficacy for relieving the positive symptoms of schizophrenia. One of the primary side effects of antipsychotic drugs, however, is weight gain and associated metabolic syndrome abnormalities. We therefore assessed whether VMAT2 inhibition affected weight gain, alone or in combination with an antipsychotic, in an antipsychotic-induced weight gain model. Olanzapine was selected as the test compound in this model since it has been demonstrated to cause a particularly high degree of weight gain in humans (Lieberman et al., 2005; Leucht et al., 2013). Although there are mixed reports regarding gender differences in olanzapine-induced weight gain in humans, preclinical studies have consistently shown that female rats exhibit a greater olanzapine-induced weight gain than do conspecific males when compared with vehicle-treated counterparts (Castellani et al., 2019). Female rats were therefore used in the weight gain experiments, as they better model the adverse weight observed in humans following olanzapine treatment than do male rats.
In this female rat model, olanzapine yielded a bell-shaped dose response. The lowest dose tested, 0.3 mg/kg, produced a small but not significant 39% increase in body weight (Fig. 9; Supplemental Table 13). The next-highest doses, 0.6 and 1.2 mg/kg, produced statistically significant 75% and 74% increases in weight, respectively (Fig. 8; Supplemental Table 12; Table 1). Increasing the dose to 3 mg/kg reduced the effect back to the same level observed with 0.3 mg/kg (Figs. 8 and 9; Supplemental Tables 12 and 13). These results are in agreement with previous studies (Heal et al., 2012).

Body weight gain in female rats fed a high-fat diet treated with olanzapine and RRR-DHTBZ alone and in combination: maximally effective olanzapine doses. In this experiment (RS1046), RRR-DHTBZ was combined with maximally effective doses of olanzapine for increasing weight gain (0.6 and 1.2 mg/kg). Body weight gain was determined over 14 days with data values calculated as body weight on day 14 minus body weight on day 1. The asterisks above the bars denote significant difference (P < 0.05) between the treatment group and vehicle. The numbers below the x-axis are P values of statistical comparison between the indicated treatment groups. Data and statistical analysis are in Supplemental Tables 12 and 12A, respectively. The group size was 10 animals.
The effect of VMAT2 inhibition alone was tested using RRR-DHTBZ. The dose range used was based on the results of the CAR experiments: 0.15 mg/kg, the subthreshold dose for suppressing CAR; 0.3 mg/kg, the threshold dose; and 0.5 mg/kg, representing a higher efficacious dose (Fig. 3). RRR-DHTBZ did not significantly affect body weight (Figs. 8 and 9) except at 0.5 mg/kg in Experiment 2 (Fig. 9). The plasma concentration versus weight gain relationship is shown in Fig. 10. RRR-DHTBZ did not increase body weight over a plasma concentration range effective for suppressing CAR (Fig. 10) or the therapeutic range for treating TD (20–48 ng/ml) (Skor et al., 2017). At the highest plasma concentration tested, 210 ng/ml, RRR-DHTBZ significantly increased weight gain (Fig. 10; Table 1). This plasma concentration is 12-fold higher than the minimally effective concentration for suppressing CAR (19 ng/ml, Fig. 6; Table 1). In combination treatment groups, RRR-DHTBZ was administered at the subthreshold dose previously shown to synergize with olanzapine and suppress the CAR response (0.15 mg/kg). This treatment did not significantly affect weight gain elicited by olanzapine. This was observed for the submaximally efficacious dose of 0.3 mg/kg olanzapine (Fig. 9) and the maximally efficacious doses of 0.6 and 1.2 mg/kg (Fig. 8). The weight gain data were plotted as a function of olanzapine concentration in Fig. 11. The data for olanzapine in combination with RRR-DHTBZ overlie those for olanzapine alone (Fig. 11; Table 1). One potential explanation for the lack of synergy by RRR-DHTBZ in weight gain versus the synergistic action in CAR is lower plasma concentration of the compounds in the weight gain model; however, this was not observed. At the synergizing plasma concentration of olanzapine in the CAR study, there was no synergy in weight gain (Fig. 11; concentration in CAR study indicated by arrow was 42 ng/ml, the average of the 0.6 mg/kg values in Supplemental Tables 9 and 10). At the synergizing dose of RRR-DHTBZ (0.15 mg/kg), the concentration of RRR-DHTBZ was 24 ng/ml in the CAR study and 60 ng/ml in the weight gain study (average of the RRR-DHTBZ/olanzapine combination values in Supplemental Tables 12 and 13). To test this possibility further, a higher dose of RRR-DHTBZ was examined; 0.3 mg/kg RRR-DHTBZ (plasma concentration of 110 ng/ml) also did not significantly alter weight gain elicited by 0.3 mg/kg olanzapine (Fig. 9).

Body weight gain in female rats fed a high-fat diet treated with olanzapine and RRR-DHTBZ alone and in combination: submaximally effective olanzapine dose. In this experiment (RS1262), RRR-DHTBZ was combined with a submaximally effective dose of olanzapine for increasing weight gain (0.3 mg/kg). Body weight gain was determined over 14 days with data values calculated as body weight on day 14 minus body weight on day 1. The asterisks above the bars denote significant difference (P < 0.05) between the treatment group and vehicle. The numbers below the x-axis are P values of statistical comparison between the indicated treatment groups. Data and statistical analysis are in Supplemental Tables 13 and 13A, respectively. The group size was 10 animals.

Body weight gain and CAR suppression versus RRR-DHTBZ concentration. Body weight gain was measured over 14 days in female rats fed a high-fat diet. CAR data are from Fig. 4. Weight gain data are in Supplemental Tables 12 and 13. The dashed red line is body weight gain of vehicle-treated rats. The asterisk denotes significant difference (P < 0.05) between the treatment group and vehicle for weight gain. (See Supplemental Tables 12A and 13A for statistical analysis.) The group size was 9–10 animals for CAR and 10 animals for weight gain.

Olanzapine-induced elevation of weight gain in the absence and presence of RRR-DHTBZ, plotted against olanzapine plasma concentration. The dose of RRR-DHTBZ was 0.15 mg/kg. Body weight gain was measured over 14 days in female rats fed a high-fat diet. The arrow indicates synergizing plasma concentration of olanzapine in the CAR study (42 ng/ml, average of the 0.6 mg/kg values in Supplemental Tables 9 and 10). Data values are given in Supplemental Tables 12 and 13. The group size was 8–10 animals for CAR and 10 animals for weight gain.
Discussion
VMAT2 inhibitors such as tetrabenazine were first developed in the early 1960s as monoamine-depleting agents for the treatment of the positive symptoms of schizophrenia (Lingjaerde, 1963). Tetrabenazine was ultimately superseded by more specific pharmacological agents, the first-generation phenothiazine antipsychotics such as chlorpromazine and later the butyrophenones such as haloperidol that directly, specifically, and potently target the postsynaptic D2 dopamine receptor (Seeman et al., 1976). Subsequently, VMAT2 inhibitors have found utility in the treatment of involuntary hyperkinetic movement disorders (Huntington’s chorea and TD, the approved drugs being tetrabenazine, deutetrabenazine, and valbenazine) (Yero and Rey, 2008; Touma and Scarff, 2018). Given the existence of approved medications targeting pre- and postsynaptic dopamine mechanisms, particularly valbenazine with its improved pharmacokinetics allowing once-daily dosing, it is now possible to clinically evaluate the combination of these mechanisms on dopaminergic morbidities such as schizophrenia. The goal of this study was to test the interaction between VMAT2 inhibition and classic antipsychotics in preclinical models of the positive symptoms of schizophrenia.
We first determined the efficacy of VMAT2 inhibition alone and, for the first time, compared it directly with that of conventional antipsychotics, using the highly selective VMAT2 inhibitor RRR-DHTBZ (also known as [+]-α-HTBZ). Previous studies have used tetrabenazine, which metabolizes into four independent and pharmacologically active metabolites that block receptors that have previously been hypothesized to play a role in treating psychosis (e.g., the D2 dopamine receptor and 5HT7 receptor) (Meltzer, 1999). RRR-DHTBZ displayed dose-dependent efficacy in both antipsychotic models (PPI and CAR) equivalent in magnitude to either typical or atypical antipsychotics (Figs. 2 and 3), confirming that inhibition of VMAT2 and by extension decreasing presynaptic dopamine release can elicit efficacy in antipsychotic models. The effective plasma concentration of RRR-DHTBZ in CAR, an animal model with translational validity regarding effective concentration for traditional antipsychotics, was similar to that of RRR-DHTBZ in humans, resulting from valbenazine dosing to treat TD (52 ng/ml at 0.7 mg/kg [Supplemental Table 3] versus 48 ng/ml at 80 mg/kg in humans) (Skor et al., 2017). The maximal detected efficacy of RRR-DHTBZ was similar to that of the antipsychotics haloperidol (in PPI, Fig. 2), risperidone, or olanzapine (in CAR, Fig. 3). This finding indicates that presynaptic modulation resulting from VMAT2 inhibition can be as effective as established antipsychotics in the behavioral models. Like the antipsychotics, VMAT2 inhibition elicited nonspecific motor effects at higher doses as anticipated for a dopaminergic inhibitor (Arnt, 1982; Wadenberg et al., 2001). At the higher doses, RRR-DHTBZ elicited escape failures (indicative of sedation, ≥0.6 mg/kg) and at the highest dose tested produced catalepsy (motor inhibition, 1.2 mg/kg) (Supplemental Tables 3 and 4). The magnitude of these effects and the window between efficacy and motor disturbance were similar to that of risperidone and olanzapine in the CAR model (Fig. 5; Supplemental Tables 5–10; Table 1). Collectively, if these findings translate to humans, selective VMAT2 inhibition as a stand-alone therapy might be as effective as conventional antipsychotics for treating the positive symptoms of schizophrenia.
We next investigated the effect of combining VMAT2 inhibition with antipsychotics on efficacy in the CAR model. A robust analytical design was implemented to enable the interaction to be rigorously quantified (Fig. 1; Kalinichev et al., 2013). RRR-DHTBZ was observed to synergize with the antipsychotics risperidone or olanzapine in suppressing conditioned avoidance behavior (Figs. 6 and 7). This was first shown using the classic synergy test (i.e., combination of subthreshold doses of the two agents). In this paradigm, the combination of nonsignificant, subthreshold doses resulted in efficacy equivalent to that of maximally effective doses of the agents administered alone (Fig. 6; Supplemental Tables 5, 6, 9, and 11). This was not due to a drug-drug interaction; the combination did not significantly affect the exposure of either agent. This provided confidence that the synergy that was observed was pharmacological or mechanism-based for the dopamine system. In the next experiments, the effect of RRR-DHTBZ on the dose-response relationship of the antipsychotics for suppressing CAR was tested (Fig. 7). More precisely, the concentration-response relationship was evaluated for both risperidone and olanzapine in the presence of subthreshold doses of the VMAT2 inhibitor. RRR-DHTBZ at a subthreshold dose produced a leftward shift of the concentration-response curve of both antipsychotics tested, olanzapine and risperidone (Fig. 7). This finding confirms that RRR-DHTBZ increased the behavioral potency of the antipsychotics in the CAR model. If these effects can be recapitulated in humans, this synergistic effect would allow a novel therapeutic approach in which low-dose administration of a VMAT2 inhibitor could in turn lower the dose of antipsychotic necessary to potentially produce efficacy in reducing positive symptoms. Reciprocally, the synergy enables a low dose of VMAT2 inhibitor to be used. This could potentially mitigate the side effects of VMAT2 inhibition noted in the early studies of tetrabenazine’s antipsychotic efficacy in humans, including somnolence and potentially depression (Lingjaerde, 1963).
A plausible mechanism for the synergistic effect is presented in Fig. 12. This mechanism is based on the known mechanics of striatal dopamine synaptic neurotransmission. In particular, it is based on the presynaptic autoreceptor function of the D2 receptor (Ford, 2014). D2 receptors in the postsynaptic membrane mediate dopaminergic signaling in the postsynaptic neuron. These receptors are the therapeutic site of action of antipsychotics (Kapur and Mamo, 2003). However, the D2 receptor is also expressed on the presynaptic terminal where it plays an autoreceptor role in a negative feedback loop that prevents excessive dopaminergic transmission (Kehr et al., 1972; Ford, 2014). In this mechanism, dopamine released into the synapse diffuses to the presynaptic terminal where it activates presynaptic D2 receptors. This results in diminished synthesis, release, and uptake of dopamine. Activation of the autoreceptor decreases dopamine synthesis by decreasing tyrosine hydroxylase activity (Kehr et al., 1972; Wolf and Roth, 1990). A problem with antipsychotics is that they interfere with this feedback mechanism since they block the autoreceptor with equal affinity (Carlsson et al., 1977; Kapur and Mamo, 2003). This means that antipsychotics stimulate dopamine presynaptically, counteracting their inhibitory postsynaptic efficacy. The antipsychotic binds to the presynaptic D2 receptor, blocking the negative feedback loop, resulting in elevated dopamine synthesis (Carlsson et al., 1977) and extracellular concentration (Arbilla and Langer, 1981; Huang et al., 2020). A VMAT2 inhibitor would in principle blunt the presynaptic action of the antipsychotic. We propose that the elevated dopamine in the presynaptic terminal resulting from autoreceptor blockade can be prevented from activating postsynaptic receptors by inhibiting its uptake into presynaptic vesicles via inhibition of VMAT2. (Uncontrolled cytoplasmic buildup of dopamine, resulting from increased synthesis and blocked vesicular packaging, is likely blunted by a second known feedback mechanism since dopamine is a competitive inhibitor of tyrosine hydroxylase, the rate-limiting enzyme in dopamine synthesis [Daubner et al., 2011]). As a result, the elevated synaptic dopamine is reduced. This reduces competition for binding of the antipsychotic to postsynaptic receptors, which in turn increases the efficacy of the antipsychotic. This correction of the self-inhibitory action of the antipsychotic explains the synergistic, as opposed to simply additive, action of VMAT2 inhibition on antipsychotic efficacy. This hypothesis is consistent with recent results quantifying bulk dopamine concentrations in the CNS by microdialysis (Huang et al., 2020). Risperidone and olanzapine both elevated dopamine, presumably as a result of presynaptic D2 receptor blockade, and coadministration of RRR-DHTBZ reduced this elevation, presumably by inhibiting presynaptic packaging of dopamine. This hypothesis could be explored in future studies at higher spatial and temporal resolution using recently reported biosensors of synaptic dopamine concentration (Patriarchi et al., 2018).

Proposed mechanism of synergy between VMAT2 inhibition and D2 dopamine receptor blockade in antipsychotic efficacy. The D2 dopamine receptor is located postsynaptically where it mediates antipsychotic efficacy (Kapur and Mamo, 2003) and presynaptically where it mediates negative feedback mechanisms by functioning as an autoreceptor (Ford, 2014). (A) Untreated dopamine synapse. Dopamine released into the synapse activates postsynaptic receptors to modulate postsynaptic signaling (1). Dopamine also activates presynaptic receptors (2), which results in diminished dopamine synthesis by a reduction of tyrosine hydroxylase activity (3). This negative feedback loop maintains control of dopaminergic neurotransmission. (B) Antipsychotic-treated synapse. The antipsychotic blocks postsynaptic D2 receptors (4). However, blockade of the autoreceptor (5) ultimately reduces the amount of block achieved, limiting efficacy. This results from elevated dopamine, a consequence of blocking negative feedback by inhibiting the action of dopamine at the autoreceptor. Specifically, antipsychotic treatment increases tyrosine hydroxylase activity (6) (Carlsson et al., 1977), increasing dopamine synthesis and presynaptic dopamine. This dopamine is subsequently released into the synapse, where it competes with antipsychotic for binding to postsynaptic receptors (7). (C) VMAT2 inhibitor-treated synapse. This mechanism inhibits postsynaptic transmission indirectly by reducing synaptic dopamine release by blocking dopamine packaging into presynaptic vesicles (8). (D) Combination of both treatments. The VMAT2 inhibitor relieves the presynaptic dopaminergic effect of the antipsychotic by blocking the packaging of the extra dopamine into presynaptic vesicles (9), attenuating the elevated synaptic dopamine resulting from antipsychotic binding to the autoreceptor (10). This reduces competition for binding of the antipsychotic to postsynaptic receptors, increasing occupancy (11). This in turn decreases dopamine signaling and increases the efficacy of the antipsychotic. Runaway elevation of cytoplasmic dopamine is likely prevented by feedback inhibition of synthesis; dopamine is a competitive inhibitor of tyrosine hydroxylase (12) (Daubner et al., 2011).
Clearly, reducing antipsychotic dose with the synergistic approach will only be beneficial if there is no concomitant potentiation of side effects. Weight gain is associated with the use of all conventional antipsychotics, as indicated on their labels. Consequently, we evaluated the effect of VMAT2 inhibition in an antipsychotic-induced weight gain model, employing olanzapine, the antipsychotic with the highest weight gain and metabolic syndrome liability (Lieberman et al., 2005; Leucht et al., 2013). When dosed alone, RRR-DHTBZ did not significantly affect body weight at doses and exposures effective in the rodent CAR model or in humans (Fig. 10). An increase was observed at the highest exposure (210 ng/ml), but this was 10-fold higher than the effective concentration for suppressing CAR. In the combination experiment, RRR-DHTBZ did not change the effect of olanzapine, either at the efficacious doses (0.6 and 1.2 mg/kg, Fig. 8) or subefficacious dose (0.3 mg/kg, Fig. 9) of olanzapine. RRR-DHTBZ did not change the plasma concentration-effect relationship of olanzapine in the weight gain model (Fig. 11), indicating that it did not affect the potency of olanzapine to elicit weight gain in contrast to its ability to enhance the efficacy of olanzapine in the CAR model. These findings indicate that the synergizing effect of VMAT2 inhibition in the CAR model is not manifested in the weight gain model at the doses tested. One explanation for the lack of synergy on weight gain is the hypothesis that weight gain is mediated by targets other than the D2 dopamine receptor alone. Implicated targets include the M3 muscarinic acetylcholine receptor, 5HT2 serotonin receptor subtypes, and the H1 histamine receptor (Roerig et al., 2011).
In summary, these findings demonstrate that VMAT2 inhibition is effective in animal models of schizophrenia and potentiates the behavioral potency of antipsychotics in these models without potentiating weight gain. If translatable to humans, these effects could provide new treatment modalities for schizophrenia with a reduced side effect burden while maintaining and potentially enhancing therapeutic efficacy.
Acknowledgments
The authors are grateful to Dr. Taleen Hanania of PsychoGenics, Inc. for guidance with experimental design. Editorial assistance was provided by Jennifer Kaiser from Prescott Medical Communications Group (Chicago, IL) with support from Neurocrine Biosciences, Inc.
Authorship Contributions
Participated in research design: Hoare, Kudwa, Luo, Grigoriadis.
Conducted experiments: Hoare, Kudwa, Luo.
Performed data analysis: Hoare, Kudwa, Luo.
Wrote or contributed to the writing of the manuscript: Hoare, Kudwa, Luo, Grigoriadis.
Footnotes
- Received October 18, 2021.
- Accepted February 10, 2022.
This work was funded by Neurocrine Biosciences, Inc.
The studies included in this report were supported by Neurocrine Biosciences, Inc. (San Diego, CA). All authors are current or former full-time employees of Neurocrine and may hold stock or other interest in the company.
Primary laboratory of origin: Neurocrine Biosciences, Inc. (San Diego, CA).
Some findings of these studies were previously presented as follows: Grigoriadis D, Hoare S, Kudwa A, and Boikess S (2017) Can presynaptic and postsynaptic monoaminergic mechanisms work synergistically? Current findings and future directions. 56th Annual Meeting of the American College of Neuropsychopharmacology; 2017 Dec 3–7; Palm Springs, CA. American College of Neuropsychopharmacology, Brentwood, TN.
Grigoriadis D, Hoare S, Kudwa A, and Boikess S (2017) VMAT2 inhibition – a novel mechanism for maintained efficacy at lowered antipsychotic doses. 72nd Annual Meeting of the Society of Biological Psychiatry; 2017 May 18–20; San Diego, CA. Society of Biological Psychiatry, Brentwood, TN.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- CAR
- conditioned avoidance response
- CNS
- central nervous system
- PPI
- prepulse inhibition
- RRR-DHTBZ
- RRR-dihydrotetrabenazine
- TD
- tardive dyskinesia
- VMAT
- vesicular monoamine transporter
- Copyright © 2022 by The Author(s)
This is an open access article distributed under the CC BY-NC Attribution 4.0 International license.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}