


Abstract
Monoamine releasers such as d-methamphetamine (d-MA) can reduce cocaine use in laboratory studies and have been forwarded for the management of cocaine use disorder (CUD). However, the proven abuse liability of d-MA has limited enthusiasm for clinical use. The levorotatory isomer of MA, l-MA, appears to have lesser stimulant effects, possibly due to its preferential norepinephrine-releasing properties compared with dopamine. The present study evaluated the abuse potential of l-MA by comparing its reinforcing effects with known stimulant drugs of abuse in nonhuman primates. Adult rhesus macaques (N = 4) responded for intravenous injections of cocaine, d-MA, methcathinone (MCAT), or l-MA under a fixed-ratio (FR) schedule of reinforcement; reinforcing effectiveness was evaluated using behavioral economic demand procedures. In a separate cohort (N = 9), daily activity and food-reinforced responding were assessed during 100 days of treatment with daily dosages of l-MA (2.3 mg/kg per day, i.v.) or d-MA (0.74 mg/kg per day, i.v.) previously shown to decrease cocaine self-administration. Results show that all drugs maintained self-administration, with peak injections reaching ∼100 inj per session for cocaine, MCAT, and d-MA and ∼50 inj per session for l-MA . In demand studies, self-administration of each drug gradually decreased as FR size increased. The exponential model of demand indicated that the reinforcing effectiveness of l-MA was significantly less than the other drugs studied. Chronic l-MA treatment did not appreciably alter daily activity and only transiently suppressed food-reinforced responding. These data, coupled with previous findings that l-MA effectively reduces stimulant self-administration, suggest that l-MA, or other norepinephrine-preferring releasers, may serve as agonist medication for CUD with lesser abuse liability than common psychostimulants.
SIGNIFICANCE STATEMENT Development of pharmacotherapies for cocaine use disorder remains a formidable challenge. Agonist-based therapies show promise, but enthusiasm is tempered by the abuse liability of previously proposed medications. This study evaluated the abuse liability and chronic treatment effects of methamphetamine’s levorotatory isomer (l-MA). l-MA demonstrated lower abuse liability compared with commonly abused stimulants and produced few untoward effects. In the context of recent studies demonstrating that l-MA attenuates stimulant self-administration, these findings support l-MA’s potential as a pharmacotherapy for stimulant addiction.
Introduction
The agonist replacement approach for medications development has been successfully employed for the management of both opioid and nicotine use disorders but has not yet resulted in a Food and Drug Administration–approved medication for cocaine use disorder (CUD) (Grabowski et al., 2004; Rush and Stoops, 2012; Kohut and Bergman, 2017). This approach is predicated on the idea that illicit drug use and harm will be decreased by treatment with a drug that shares pharmacological properties with the illicit drug but with lesser abuse-related and unwanted effects (Rothman et al., 2008). Several pharmacologically similar compounds, all with prominent dopaminergic actions, have been found to effectively decrease cocaine intake in preclinical and/or clinical studies [reviewed by Czoty et al. (2016)]. In fact, a recent meta-analysis found that prescription psychostimulants, particularly amphetamines, significantly increase rates and duration of abstinence as well as cocaine-negative urines (Tardelli et al., 2020), adding additional support for the clinical benefit of utilizing agonist medications for CUD. However, identifying compounds that meet the criteria for agonist medication with reduced abuse liability has remained a formidable challenge.
We have previously suggested that the levo isomer of methamphetamine, l-methamphetamine (l-MA), which is available over the counter as the active ingredient in the nonprescription nasal decongestant Vick’s VapoInhaler may be a promising candidate medication for CUD, in part, because of its unique pharmacological profile (Kohut and Bergman, 2016, 2017; Kohut et al., 2016; Bardo et al., 2019). Both isomers—l-MA and the dextro isomer, d-methamphetamine (d-MA)—show similar time courses of distribution throughout the brain (Fowler et al., 2007) and are relatively equipotent for releasing norepinephrine (NE) (Kuczenski et al., 1995; Melega et al., 1999). However, l-MA is approximately 15- to 20-fold less potent than d-MA and other stimulant drugs of abuse for releasing dopamine (DA), which may be important in determining stimulant-like abuse-related effects (Kuczenski et al., 1995; Melega et al., 1999; Rothman et al., 2001; Kohut et al., 2017).
These pharmacological differences, although subtle, may underlie the mix of overlapping and distinct behavioral effects reported between the two isomers. In preclinical behavioral studies, treatment with either d- or l-MA similarly reduced cocaine self-administration in nonhuman primates (c.f., Negus et al., 2007; Kohut et al., 2016) and d-MA self-administration in rats (Bardo et al., 2019). Both d-MA and l-MA produce cocaine-like discriminative stimulus effects (Kohut et al., 2017), and l-MA substitutes for d-MA in drug discrimination studies (Yasar and Bergman, 1994; Bondareva et al., 2002; Desai and Bergman, 2010). Further, l-MA treatment has been shown to induce cross-sensitization to d-MA in rats (Xue et al., 2019). However, unlike d-MA, l-MA does not stimulate locomotor activity, nor does it induce locomotor sensitization with repeated dosing in rats (Nishimura et al., 2017; Siemian et al., 2017), suggesting that l-MA has lesser stimulant-like effects compared with d-MA. Taken together, these results indicate that although there are shared mechanisms, both drugs produce divergent behavioral and stimulant effects related to abuse potential.
We recently reported that l-MA is less reinforcing than d-MA in drug-naïve rats and attenuates self-administration of cocaine and d-MA in primates and rats, respectively (Kohut and Bergman, 2016; Bardo et al., 2019). The present studies were conducted to characterize the abuse-related reinforcing and stimulant-like behavioral effects of l-MA in rhesus monkeys with histories of stimulant self-administration. First, well validated drug self-administration procedures were used to compare the reinforcing effects of l-MA with standard monoaminergic drugs of abuse: d-MA, cocaine, and the substituted cathinone, methcathinone (MCAT). Behavioral economic procedures were then used to quantify the reinforcing strength of l-MA relative to the same standard stimulants. Second, because medication for the management of cocaine or other stimulant use disorders is likely to require long-term treatment in clinical populations, behavioral effects (daily activity and food-maintained responding) and the safety of chronic dosing were monitored while subjects were treated for 100 days with d- or l-MA at doses that produce comparable decreases in cocaine self-administration in rhesus monkeys (Negus et al., 2007; Kohut et al., 2016). Results indicate that l-MA is less reinforcing compared with common psychostimulants with minimal unwanted effects, strengthening its consideration as an agonist therapy for stimulant use disorders.
Materials and Methods
Subjects
Adult male and female rhesus macaques (Macacca mulatta) with histories of cocaine self-administration served as subjects. Monkeys had continuous access to water and were fed High Protein Monkey Diet (Purina Mills International, Brentwood, MO), fresh fruit, and vegetables daily in addition to 1 g banana-flavored food pellets (Formula 4TUR banana flavor, Purina Mills Test Diet, Richmond, IN) earned during experimental sessions. A 12-hour light/dark cycle was in effect, except during experimental sessions, as noted below. Subjects lived in stainless steel cages with side and front visual access to other subjects in the vivarium. Environmental enrichment was provided through access to mirrors, toys, and foraging boards; music during nonexperimental periods; and interactions with technical staff.
Animal husbandry was conducted in accordance with the guidelines provided by the Institute of Laboratory Animal Resources - National Research Council (2010) as adopted and promulgated by the US National Institutes of Health. The facility is licensed by the US Department of Agriculture, and all experimental protocols were approved by the Institutional Animal Care and Use Committee at McLean Hospital. Animal health and well being were monitored daily by trained technical staff and by the attending veterinarian.
Apparatus
Monkeys responded on custom operant panels containing three square translucent response keys (5.1 × 5.1 cm) arranged 3.5 cm apart in a horizontal row 9 cm from the top of the panel. Each key could be transilluminated with red or green stimulus lights (SuperBright LEDs; Fairchild Semiconductor, San Jose, CA). A pellet dispenser (Gerbrands model G5210, Arlington, MA) was positioned above the cage next to two syringe pumps (model 981210; Harvard Apparatus, Inc., South Natick, MA), one for each lumen of the double-lumen catheter. A plastic food cup was located near the bottom of the operant panel and provided access to response-contingent food pellets earned during experimental sessions. In self-administration experiments, two subjects sat in standard primate chairs (Primate Products, Immokalee, FL) within a custom-built sound-attenuating chamber and faced the operant panel—all other events/equipment were as described above. Experimental events were controlled by a desktop computer running Med Associates (Georgia, VT) software.
Surgery
Silicone catheters (i.d. 0.028 inches, o.d. 0.088 inches; Saint Gobain, Beaverton, MI) were surgically implanted in an internal or external jugular or femoral vein under aseptic conditions. Monkeys were initially administered atropine (0.05 mg/kg, i.m.) and then sedated with ketamine (5–10 mg/kg, i.m.). After intubation, anesthesia was maintained with isoflurane (1%–2% mixed with oxygen) throughout the procedure. After surgery, monkeys were given a single dose of penicillin G at 20,000 units/kg, i.m., and cephalexin 20 mg/kg, oral, twice daily for 5 days. Analgesic doses of buprenorphine (0.018 mg/kg, i.m.) and Metacam (meloxicam; 0.1 mg/kg, s.c. or oral) were administered twice daily for 3 days.
The intravenous catheter exited at the midscapular region. Subjects wore custom-fitted nylon jackets to protect the catheter. In tethered experiments, jackets were connected to a flexible stainless steel cable and fluid swivel (Lomir Biomedical, Malone, NY) that permitted monkeys to move freely within the home cage. Catheter patency was evaluated periodically by intravenous administration of methohexital sodium (3 to 4 mg/kg).
Drug Self-Administration
Two male and two female subjects were trained to respond for food pellets and drug injections per previously published methods (Kohut et al., 2013, 2016). Each daily 110-minute behavioral session consisted of two response components separated by a 5-minute time-out (TO) period. During the first component, a red light illuminating the center response key signaled the availability of 1 g banana-flavored food pellets under a fixed-ratio 10 (FR10):TO 10-second schedule of reinforcement for 10 minutes. During the second component, green lights illuminating the center response key signaled the availability of intravenous injections of drug or saline for 100 minutes under an FR10:TO 30-second schedule. This component was immediately preceded by illumination of a yellow light for 10 seconds and the noncontingent delivery of a single injection of saline or drug (i.e., cocaine, d-MA, MCAT, or l-MA) at the dose that was subsequently available. During the TO after each reinforcer delivery (food or drug), the center response key was illuminated with yellow lights. During the intercomponent TO period, all lights were off, and responding had no scheduled consequences.
During training sessions, saline or a unit dose of cocaine was typically available under a double alternation schedule that varied irregularly between subjects. Once responding was stable, dose-response functions for cocaine (0.001–0.1 mg/kg per inj), d-MA (0.001–0.1 mg/kg per inj), MCAT (0.001–0.1 mg/kg per inj), and l-MA (0.01–0.32 mg/kg per inj) were determined in a mixed order by substituting the various doses of each drug for cocaine. In general, each drug dose was studied for 2 to 3 days, and responding was considered stable if the number of injections over that period was within 15% of the 2- to 3-day mean before switching to another dose. Complete dose-effect functions for a single drug were obtained before moving to the next one. Subjects could earn an unlimited number of drug injections during each daily session, and sessions were monitored by technical staff.
Behavioral Economic Demand Curves
The relative reinforcing strengths of cocaine, d-MA, MCAT, and l-MA were determined using behavioral economic demand procedures (Hursh, 1993; Hursh and Silberberg, 2008). These studies were identical to dose-effect determinations, except that, after 3 days of stable self-administration under the FR10:TO30-second schedule of reinforcement, the response requirement for drug delivery was increased across successive sessions (i.e., 10, 32, 56, 100, 320, 560, 1000, 3200) until subjects failed to earn a single injection at a given ratio. Two doses of each drug were studied: the dose that maintained peak levels of responding during dose-effect determinations and a one-half log dose higher than the peak (+1/2-log peak; i.e., first dose on the descending limb). The doses used were as follows: cocaine = 0.01 and 0.032 mg/kg per inj, d-MA = 0.0032 and 0.01 mg/kg per inj, MCAT = 0.01 and 0.032 mg/kg per inj, and l-MA = 0.1 and 0.32 mg/kg per inj. Drug availability remained unlimited during this procedure.
Chronic Treatment
The effects of 100-day chronic treatment with saline, d-MA (0.032 mg/kg per hour; 0.74 mg/kg per day), or l-MA (0.1 mg/kg per hour; 2.3 mg/kg per day) on food-maintained responding, daily activity, and complete blood count (CBC)/chemistry profile were evaluated. These doses of d- and l-MA were chosen because previous studies have shown that 7–10 days of continuous treatment produces similar decreases in ongoing cocaine self-administration (by approximately 50%) in rhesus monkeys with minimal effects on food-maintained responding (Negus et al., 2007; Kohut et al., 2016). Nine subjects (two males and one female per condition) with histories of cocaine self-administration were studied; age and body weight were matched across conditions prior to the initiation of these studies. Each subject was drug-free for >3 months prior to the onset of chronic dosing procedures. Saline, d-MA, or l-MA was administered intravenously every 20 minutes for 23 hours each day (1100–0700 hours), as in our previous reports (Kohut et al., 2016).
Operant sessions to measure food-maintained responding consisted of four 1-hour periods of banana-flavored food pellet access each day: 1100, 1500, 1900, and 0600 hours the following morning. During response components, a red light illuminating the center response key signaled the availability of 1 g banana-flavored food pellets under a fixed ratio 2, variable ratio 16 second-order schedule of reinforcement. A 10-second TO period followed delivery of each food pellet, during which stimulus lights were turned off. This schedule of reinforcement was used for consistency with our previous studies of chronic treatment with d- or l-methamphetamine on cocaine and food-maintained responding in rhesus monkeys (Negus et al., 2007; Kohut et al., 2016). If 25 food pellets were delivered before the end of the 1-hour session, then all stimulus lights were turned off, and responding had no scheduled consequences for the remainder of that session.
Daily activity was measured using a commercially available three-way accelerometer (FitBit Zip; FitBit, San Francisco, CA) attached to the back of each subject’s nylon jacket. The accelerometer used has been shown to produce reliable activity data in human studies (Imboden et al., 2018) and was used here to measure total daily activity counts, as we have previously reported from macaque subjects (Jacobs et al., 2017). Data were downloaded at least twice weekly through Bluetooth technology.
CBC/chemistry profiles were determined in each subject prior to (baseline) and at approximately 30, 60, 90, and 100 days of chronic treatment to monitor blood electrolytes, cholesterol, triglycerides, and liver and kidney function. Blood samples were collected under ketamine anesthesia during monthly health checks via a temporary catheter placed in a saphenous or tibial vein. To collect samples, subjects were anesthetized with 10 mg/kg ketamine, and the site was thoroughly cleaned with a chlorhexidine solution and wiped with alcohol. Samples were analyzed using a standard CBC/chemistry panel by a commercial laboratory (IDEXX laboratories, Westbrook, ME).
Drugs
Cocaine HCl and racemic methcathinone HCl were obtained from the National Institute on Drug Abuse Drug Supply program. d-Methamphetamine HCl was purchased from Sigma-Aldrich (St. Louis, MO) and l-methamphetamine HCl was provided by B. Blough. All drug solutions were dissolved in sterile saline and filtered using a 0.22-micron syringe-driven filter (Millipore Corporation, Billerica, MA). All drug doses were calculated using the salt weights.
Data Analysis
Data were collected according to a preset plan for assessment of drug self-administration and chronic treatment. The number of responses and reinforcement deliveries were averaged for the last 2 days for each dose of drug or FR value and used for further analysis. The primary dependent variables for self-administration were total injections earned per session, and, for behavioral economic analysis, demand intensity and elasticity derived from demand curves (see below).
One-way ANOVA for repeated measures with Dunnett’s post hoc tests were used to compare the number of injections of various doses of each drug to identify those doses that were self-administered at levels higher than those observed for saline.
The demand curve analysis employed the exponential model of demand (Hursh and Silberberg, 2008; Hursh, 2014). Non-normalized consumption (Q; injections earned) and price (P; FR value) for each drug were fit to the exponential model of demand equation: log Q = log Q0 + k(e−αP− 1); k denotes the range of the exponential model and was experimentally derived and shared across all conditions, α represents a free parameter to quantify the elasticity of each drug from the demand curve, and Q0 represents a free parameter to quantify the maximum intensity of demand. Significant differences for each drug were determined when 95% confidence intervals did not overlap between drugs for a given parameter, in accordance with previous studies (Kohut and Bergman, 2016).
Activity data were downloaded from FitBit servers using custom software and organized as total activity counts per day. Mean daily activity (total activity counts/24 hours) was determined over a 7- to 10-day period under baseline conditions for each subject prior to chronic treatment, and daily activity was normalized to their own baseline. The primary dependent variable in food-maintained responding experiments was the total number of food pellets earned per day. Activity counts and food-maintained responding data were binned in 5-day epochs prior to statistical analysis. Two-way repeated measures ANOVA with factors of treatment and day were followed by Dunnett’s post hoc tests, when appropriate, to determine whether food-maintained responding, activity, or CBC/chemistry values during treatment with d- or l-MA differed from the saline control group at any time point during chronic treatment. All figures and statistical analyses were carried out using GraphPad Prism 6.0 for Mac (GraphPad Software Inc.) and used a critical P < 0.05.
Results
Drug Self-Administration
Initially, control values were established for the self-administration of intravenous saline and several doses of intravenous cocaine. When saline was available for self-administration, subjects generally responded for <20 injections per session. When cocaine was available for self-administration, the cocaine dose-effect curve described an inverted U-shaped function in all monkeys: the number of injections per session increased in a dose-dependent manner [FDose(5,15) = 8.41; P < 0.001], with 0.01 mg/kg cocaine producing peak levels of responding at the group level (107.4 ± 18.6 inj per session), which decreased as the unit dose increased further. Responding for unit doses of 0.0032–0.032 mg/kg per inj was significantly greater than saline vehicle (all P < 0.05; Fig. 1).

Self-administration of cocaine (circles), MCAT (squares), d-MA (downward triangles), and l-MA (upward triangles). Abscissa, drug dose in milligrams per kilogram per injection (log scale). Ordinate, total drug injections for a given dose earned per session. All data points represent mean ± S.E.M. of four subjects. Filled symbols indicate significant (P < 0.05) differences from saline self-administration.
When d-MA [FDose(5,15) = 5.88; P < 0.001], MCAT [FDose(5,15) = 15.7; P < 0.0001], and l-MA [FDose(4,12) = 4.86; P = 0.02] were substituted for cocaine, self-administration of each drug followed a similar inverted U-shaped dose-effect function, but with varying maximal levels of responding (see Fig. 1). Peak levels of self-administration for d-MA and MCAT were similar to cocaine, with subjects earning, on average, 93.8 (± 8.5) and 100.3 (± 10.7) inj per session at 0.003 mg/kg d-MA and 0.01 mg/kg MCAT, respectively. Doses of 0.0032 and 0.01 mg/kg per inj d-MA and 0.0032–0.032 mg/kg per inj MCAT elicited responding greater than saline (all P < 0.05). The peak number of injections of l-MA earned during self-administration sessions (50.0 ± 6.6 inj at 0.1 mg/kg) was approximately 50% of that earned when cocaine, d-MA, or MCAT were available, and only intake of the peak dose of l-MA was significantly different from saline self-administration (P = 0.01).
Figure 2 shows representative cumulative records for self-administration of cocaine, d-MA, MCAT, and l-MA during dose-effect determinations at the peak and +1/2-log peak dose of each drug. The within-session pattern of responding for cocaine and MCAT at both doses was characterized by relatively consistent responding throughout the session with injections earned at regular intervals. This pattern of responding was also evident during the first half of d-MA sessions but with longer “break” periods during the second half of sessions in which bouts of injections were more spaced, likely reflecting the accumulation of drug due to d-MA’s longer duration of effects and half-life relative to cocaine (Newton et al., 2005). In contrast to cocaine, MCAT, and d-MA, the within-session pattern of responding for l-MA was almost exclusively restricted to the first 60 minutes of the session and more spaced injections (i.e., 10–30 minutes) at the higher unit dose. This pattern was consistent in all subjects studied—i.e., >80% of all injections earned of 0.1 mg/kg l-MA were earned in the first half of the session.
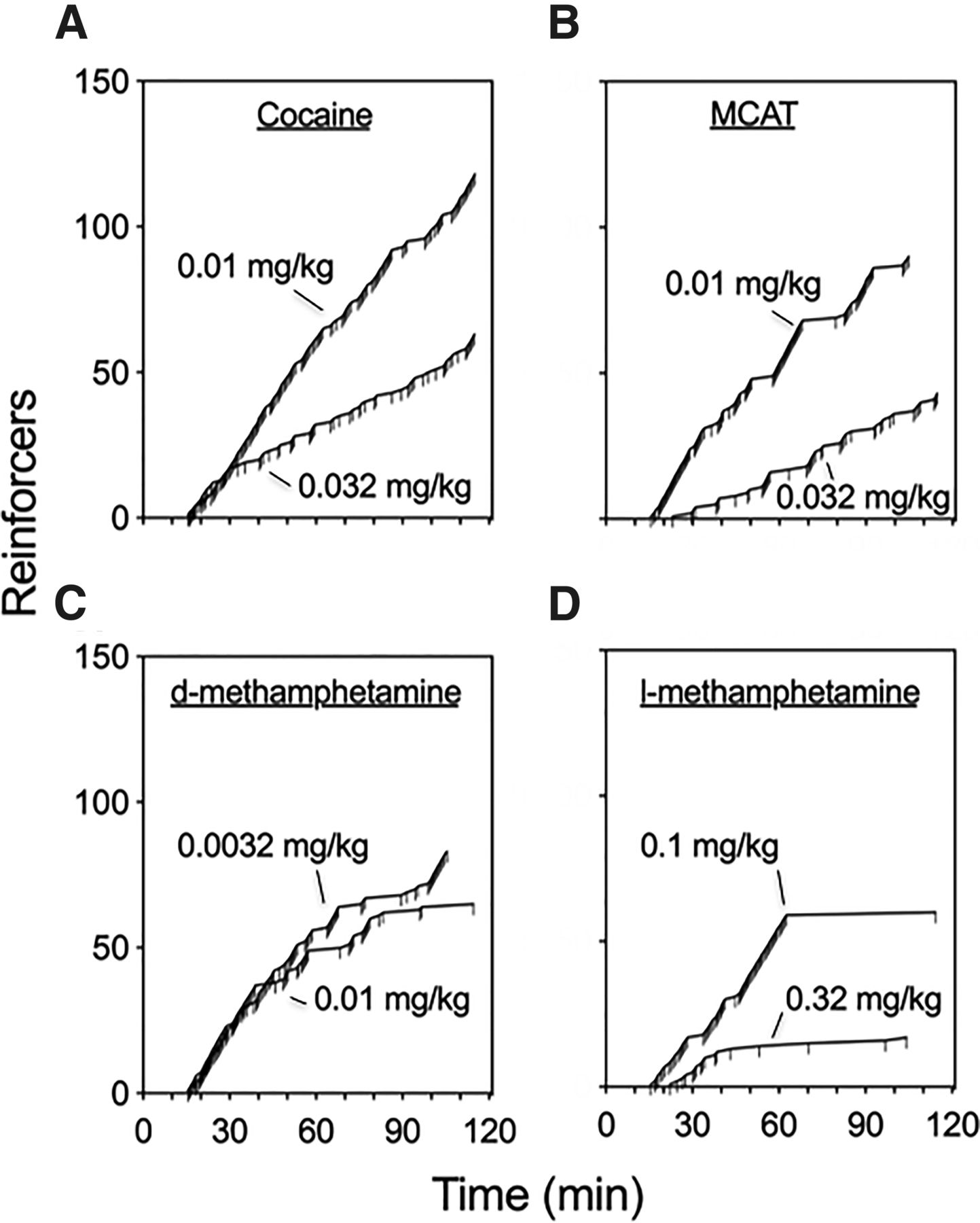
Representative cumulative records for administration of peak and +1/2-log peak doses of cocaine, MCAT, d-MA, or l-MA. Abscissa, total time of session in minutes. Ordinate, number drug reinforcers earned. Ticks represent reinforcer delivery. Most stimulants (A–C) engendered consistent and robust self-administration patterns, except for l-MA (D), which was lesser and more erratically administered.
Behavioral Economic Demand Curves
Demand curves in Fig. 3, A and B show self-administration of 0.01 and 0.032 mg/kg cocaine and MCAT, 0.0032 and 0.01 mg/kg d-MA, and 0.1 and 0.32 mg/kg l-MA as a function of increasing response requirements (i.e., price). During demand procedures, responding for each drug decreased as the response requirement for drug delivery increased. The exponential model of demand provided a good fit for all data sets (R2 range: 0.95–0.99). Overlapping confidence intervals were observed for intensity (Q0) across all drugs at the peak dose, but the 1/2-log increase in dose resulted in a lower Q0 for l-MA compared with MCAT. Comparison of demand elasticity (α) for the peak and +1/2-log peak dose of each drug indicated that l-MA, at both doses, was more elastic than that of cocaine, d-MA, and MCAT, as confidence intervals did not overlap. Cocaine, d-MA, and MCAT did not differ from each other, and no change in elasticity was observed with dose (Fig. 3, C and D).

Demand curves and parameter estimates for the peak dose (white symbols) and +1/2-log peak doses (filled symbols) of cocaine (Coc), MCAT, d-MA, and l-MA observed in self-administration. Doses are milligrams per kilogram per injection. (A and B) Each data point represents the mean number of injections earned as a function of price for n = 4 subjects. Abscissa, FR requirement for an injection (log scale). Ordinate, total drug injections earned for a given session. Lines represent the best fit for each data point determined using the exponential model of demand (Hursh and Silberberg, 2008). (C and D) Mean and 95% confidence intervals (CIs) for demand intensity [(C) Q0] and elasticity [(D) α] parameters for each drug. Abscissa, estimated parameter value. Ordinate, drug available for self-administration. *95% CIs do not overlap with cocaine, #95% CIs do not overlap with MCAT.
Chronic Treatment
A clinical treatment regimen of chronic exposure to l-MA was modeled by continuously administering l-MA (0.1 mg/kg per hour, i.v.) or, for comparison, d-MA (0.032 mg/kg per hour, i.v.) or saline vehicle for 23 hours per day over approximately 100 consecutive days. Baseline levels of activity did not differ between the three chronic treatment groups [FTreatment(2,6) = 0.714; P = 0.53]; however, as expected, there were considerable individual differences among subjects (overall mean = 4364 counts, minimum = 57, maximum = 26,387). As such, total daily activity count was transformed to a percentage of each individual’s baseline level of activity for statistical analysis. Daily activity varied as an interaction between treatment and day across the chronic treatment period [FTreatmentXDay(36,108) = 2.13; P = 0.002; Fig. 4, A–C]. In the saline- and l-MA–treated groups, mean daily activity was primarily maintained within 20% of baseline levels throughout the entire treatment period. In contrast, daily activity was markedly decreased in the d-MA–treated group to about 50% of baseline levels. This decrease was evident in the first week of treatment, maintained for approximately 30 days, and then steadily returned to baseline after about 45 days of treatment. Baseline levels of activity in the d-MA–treated group were then maintained for the remainder of the treatment period.
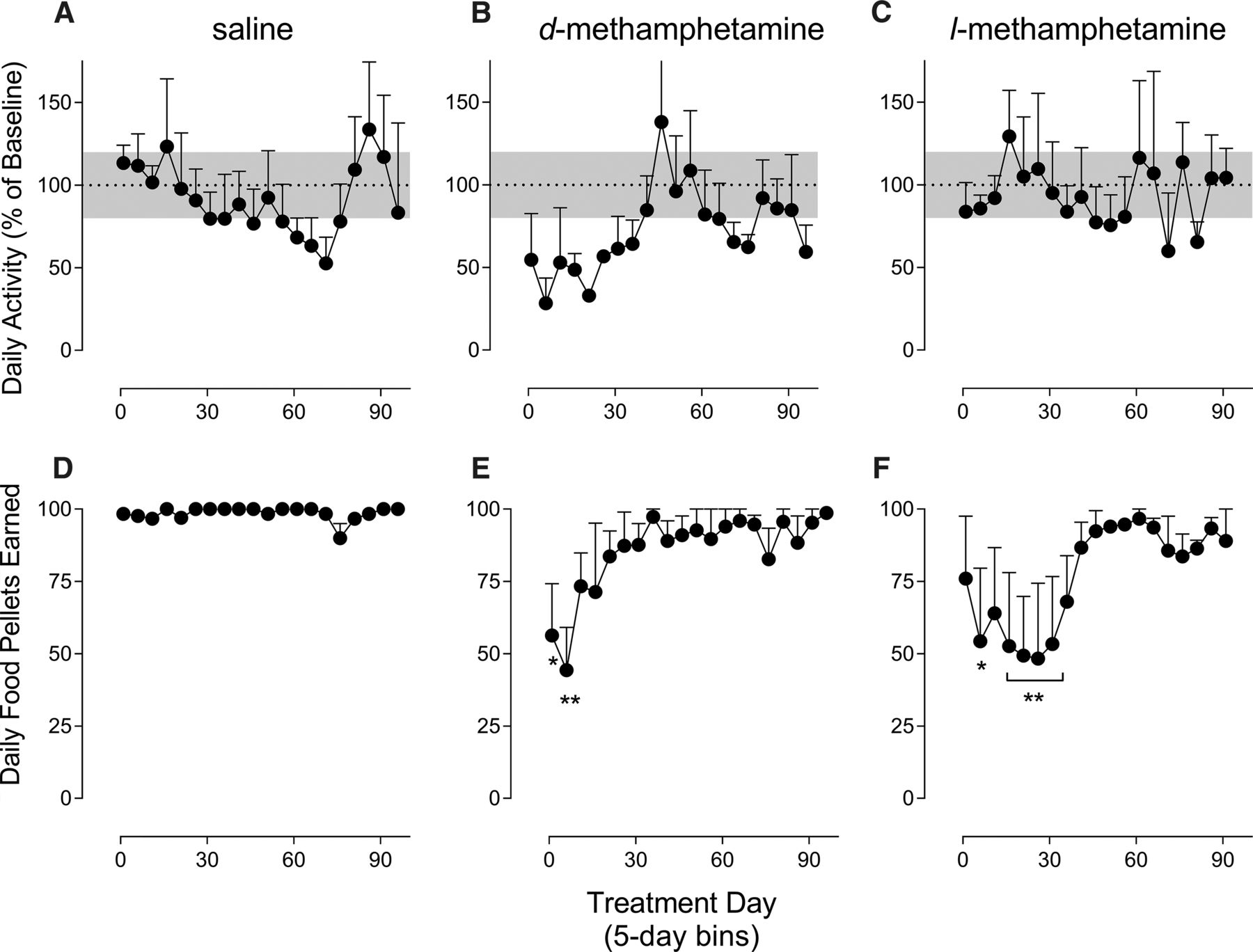
Effects of chronic 100-day treatment with saline, d-MA (0.032 mg/kg per hour), or l-MA (0.1 mg/kg per hour) on locomotor activity and food self-administration. Abscissa, treatment day. Ordinate, daily activity expressed as a percentage of pretreatment baseline or daily food pellets earned. (A–C) d-MA but not l-MA or saline resulted in multiweek suppression of behavioral activity by 20% of baseline (shaded gray area). (D–F) d-MA and l-MA transiently disrupted food-reinforced responding during chronic treatment, whereas the effect of d-MA demonstrated a faster onset and offset than l-MA. *P < 0.05 vs. saline; all data points represent mean ± S.E.M. of three subjects.
Subjects generally earned the maximal number of food pellets available each day (i.e., 100) prior to the onset of the 100-day treatment period. Food-maintained responding, however, varied among the treatment conditions as a function of day of treatment [FTreatmentXDay(36,108) = 1.912; P = 0.006; Fig. 4, D–F]. Saline-treated subjects earned near the maximal number of daily food pellets available during the entire treatment period. In contrast, both d-MA and l-MA treatment decreased the number of pellets earned, but with different patterns of disruption. Food-maintained responding in d-MA–treated monkeys decreased to approximately 50% of the maximal number available at the 5- and 10-day time points (both P < 0.02). Responding then increased to 75% over the next two time points before returning to near baseline levels of food intake for the remainder of the treatment period, but with more variability than saline-treated subjects. Like the d-MA group, food-maintained responding was decreased to about 50% of the maximal number of pellets available in l-MA–treated subjects; however, this disruption occurred gradually and was observed later in treatment than the disruption seen with d-MA. That is, responding decreased to about 75% of baseline levels over the first 5 days of treatment (P = 0.21) but then further decreased to 50% between days 5 and 10, which lasted approximately 30 days (P < 0.02; except for days 10–15, P = 0.06). Food intake then steadily returned to baseline by about day 45 of treatment, which was maintained for the remainder of the treatment period.
Blood samples were collected prior to and at approximately 30, 60, 90, and 100 days of the chronic treatment period to examine changes in CBC/chemistry profile consequent to long-term treatment with d- or l-MA. There was no evidence for changes in any CBC measure between any of the treatment groups (all F values < 1.8; P > 0.05). Measures of alanine transaminase [FDay(4,24) = 8.383; P = 0.0002], albumin [FDay(4,24) = 3.47; P = 0.023], and bicarbonate [FDay(4,24) = 4.808; P = 0.0054] showed time-dependent changes; alanine transaminase and bicarbonate levels increased, and albumin levels decreased in all groups during the treatment period.
Discussion
The present study compared the abuse-related effects of l-MA with stimulant drugs of abuse—cocaine, d-MA, and MCAT—and assessed the effects of chronic l-MA treatment. l-MA has been shown to produce sustained decreases in cocaine and d-MA self-administration (Kohut et al., 2016; Bardo et al., 2019) and shares several behavioral and pharmacological effects with other monoamine releasers that have been proposed as candidate medications for cocaine and other stimulant use disorders (Herin et al., 2010; Czoty et al., 2016; Kohut and Bergman, 2016, 2017; Siemian et al., 2017; Xue et al., 2019). However, unlike previous candidates, l-MA is considered to have less prominent dopaminergic and stimulant-like behavioral effects, which may make it a more suitable agonist medication. The present study found that l-MA was self-administered to a lesser extent and showed lower reinforcing effectiveness than cocaine, d-MA, and MCAT in nonhuman primates with histories of stimulant self-administration. Further, chronic l-MA treatment was well tolerated, producing minimal disruptions in daily activity and transient effects on food-maintained responding. These results support l-MA’s potential as an agonist therapy for stimulant use disorder.
The present finding that cocaine, d-MA, and MCAT maintained high rates of self-administration behavior and were similarly reinforcing is consistent with previous studies. For example, self-administration of MCAT in baboons was comparable in peak injections earned and total intake with cocaine (Kaminski and Griffiths, 1994), and progressive-ratio breakpoints were similar between d-MA and cocaine in rhesus monkeys (Lile et al., 2013). These findings are not surprising given the well documented abuse liability (schedule I or II compounds) and prominent dopaminergic mechanism of action of these compounds (Rothman and Baumann, 2003; Simmler et al., 2013). In contrast, l-MA engendered approximately half the number of injections at the peak dose and was 10- to 30-fold less potent than cocaine, d-MA, and MCAT. These data in rhesus monkeys with histories of stimulant self-administration extend previous findings in drug-naïve rats showing similar potency and effectiveness differences between d-MA and l-MA self-administration (Yokel and Pickens, 1973; Bardo et al., 2019). A previous study in rhesus monkeys, however, found that l-MA maintained rates of self-administration comparable to cocaine using a cumulative dosing procedure (Winger et al., 1994). Although this may appear to be at odds with the data presented here, procedural variations between the studies may explain these differences. Whereas Winger et al. (1994) used a self-administration procedure in which total injections were limited to 20 during 25-minute components, the present study permitted 100 minutes of unlimited access. Inspection of the cumulative records from the present study revealed that although cocaine, MCAT, and d-MA were robustly self-administered throughout the session, only l-MA engendered patterned responding for about 25 minutes before becoming slowed and infrequent. Importantly, these data suggest that although l-MA maintains self-administration behavior, it may be limited in a way not seen with dopamine-preferring psychomotor stimulants.
Simple FR schedules of reinforcement, such as those used in the present study, have been employed to study the relative abuse potential of investigational compounds but are not designed to directly measure reinforcing effectiveness (Kohut and Bergman, 2016, 2018; de Moura et al., 2021). Behavioral economic analysis applied to simple FR schedules of reinforcement is a powerful complement, providing a quantitative metric of demand for a reinforcer that can be compared across studies with various reinforcers, doses, and treatment regimens (Strickland and Lacy, 2020). Such claims are supported by results from the current study, as the elasticity of consumption for each drug studied was found to be dose-independent, highlighting the potential utility of this approach for generalizations of reinforcer essential value across factors that influence drug intake (Bickel et al., 1990; Strickland and Lacy, 2020). Maximum intensity, however, showed dose-dependent differences; although the maximum intensity for l-MA overlapped with the other stimulants at the peak dose, a 1/2-log increase in dose resulted in a lesser maximum intensity for l-MA compared with MCAT. This divergence is to be expected, as maximum intensity is sensitive to differences in size of commodities, such as reinforcer magnitude or dose (Hursh and Silberberg, 2008).
Cocaine, d-MA, and MCAT showed comparable reinforcing effectiveness in nonhuman primates as demand elasticity values were similar for peak and peak + 1/2-log doses. This result is consistent with findings in rats in which several schedule I monoaminergic drugs of abuse demonstrated equal or greater reinforcing effectiveness compared with cocaine or racemic MA (Huskinson et al., 2017; Gannon et al., 2018). Importantly, l-MA exhibited the lowest reinforcing effectiveness in the present study, as demand elasticity was greater for l-MA (i.e., less resistant to increases in price) than cocaine, d-MA, and MCAT. Thus, the lower abuse-related behavioral profile of l-MA compared with monoaminergic stimulants with prominent dopaminergic effects spans across fixed ratio and behavioral economic self-administration procedures.
The lower rates of l-MA self-administration and greater demand elasticity are unlikely to be explained by pharmacokinetic differences, as d- and l-MA have similar onset, elimination half-life, and distribution in brain (Melega et al., 1999; Mendelson et al., 2006; Fowler et al., 2007; Mendelson et al., 2008; Li et al., 2010). Rather, the different pharmacodynamic profiles at monoamine systems—specifically dopamine and norepinephrine—between d- and l-MA likely underlie these differences. Cocaine, d-MA, and MCAT each show a preference or near equal potency for either dopamine transporter inhibition or DA release compared with NE, whereas l-MA shows the greatest (∼15-fold) NE:DA selectivity ratio of these compounds (Rothman et al., 2001; Simmler et al., 2013; Kohut et al., 2013; 2017). Thus, the role of more prominent noradrenergic mechanisms by l-MA is likely a key contributor to the differences in reinforcing effects reported here. Few studies have been conducted with NE-preferring releasers, but previous reports have shown drugs that primarily increase NE in the synapse, via NE transporter inhibition, do not support self-administration in rhesus monkeys (Woolverton, 1987) or increase measures of drug reward in conditioned place preference procedures in rodents [Martin-Iverson et al., 1985; but see Weinshenker and Schroeder (2007) for review]. Rather, evidence suggests that elevated NE signaling within the mesolimbic pathway promotes aversive states (Schank et al., 2008; Jones et al., 2009, 2010; Vranjkovic et al., 2014), an effect which may limit drug intake. In support of this, attenuating cocaine’s noradrenergic effects by genetic knockout of norepinephrine transporter in mice enhances cocaine reward in conditioned place preference (Xu et al., 2000; Sora et al., 2001). Further, cocaine, l-MA, and d-MA each produce subjective ratings of “drug-liking,” but l-MA’s drug-liking effects are weaker, and l-MA fails to elicit increases in positive mood compared with d-MA in human MA users (Rush and Baker, 2001; Mendelson et al., 2006). Further, l-MA’s self-reported drug effects have been shown to dissipate quicker than d-MA in experienced drug users [Mendelson et al., 2006; see also Kohut et al. (2017) for cocaine-like discriminative stimulus effects in monkeys]. These data, along with the differences observed in within-session pattern of responding shown in cumulative records, may suggest that the 15-fold greater release of NE over DA produced by l-MA serves to limit l-MA self-administration after a threshold of NE activity is reached. Once that threshold is met, the increased brain NE levels produce an aversive state that diminishes the DA-mediated reinforcing effects. This effect is not seen with standard stimulants because they are typically nonselective NE/DA releasers/uptake inhibitors and synaptic NE and DA levels are highly coupled. Thus, l-MA’s favorable profile as an agonist medication is based, in part, on its reinforcing effects, which would maintain compliance, but that excessive use may be limited by the emergence of aversive effects at higher doses.
d-MA–induced disruptions in food-reinforced responding during 100 days of chronic dosing were immediate and lasted for about a week, similar to those seen with 28-day treatment of d-amphetamine in nonhuman primates and 7-day d-MA treatment in drug-naïve rats (Negus and Mello, 2003; Bardo et al., 2019). Utilizing a 0.1 mg/kg per hour l-MA treatment regimen, which is sufficient to decrease cocaine self-administration in rhesus monkeys (Kohut et al., 2016), l-MA produced a gradual disruption in food-reinforced responding, with tolerance to these effects emerging after 4 weeks of treatment. This finding is intriguing because 5–10 days of treatment with the same l-MA dose produced no significant effect on food-reinforced responding in primates with a recent cocaine self-administration history (Kohut et al., 2016). Rather, the present results, in subjects that were drug-free for several months prior to chronic l-MA treatment, more closely align with recent results from drug-naïve rats showing that l-MA potently decreased food-maintained responding (Bardo et al., 2019). Taken together, these findings suggest that cross-tolerance between the disruptive effects on food-reinforced behavior from l-MA and cocaine occurs, which could be clinically relevant for body weight changes seen in cocaine dependence and abstinence in humans (Cowan and Devine, 2008; Ersche et al., 2013). Given the overlapping but unique effects of l-MA compared with d-MA in behavioral responses outlined earlier, the divergent temporal pattern in the disruption of food-reinforced responding by d- and l-MA could similarly suggest different mechanisms. It is worth noting that changes in daily chow consumption or body weight were not observed during the treatment period. Previous studies suggest that the effects of NE on satiety are mediated in part by α-adrenergic receptors in the hypothalamus, and functionality of these receptors is documented to be altered by chronic cocaine administration (Wellman, 2000; Baumann et al., 2004). However, these suggestions are speculative, and additional studies designed to investigate the underlying pharmacological mechanisms responsible for d- and l-MA–induced decreases in food-maintained responding are needed to fully address this question.
Similar to the vehicle-treated group, chronic l-MA treatment did not alter daily activity across the entirety of the 100-day treatment period. This finding suggests l-MA may be better tolerated than d-MA, as d-MA suppressed activity for several weeks after treatment onset.d-MA’s more pronounced suppression of daily activity may be related to its prominent dopaminergic effects, as dopamine agonists have been shown to increase stationary posture in primates (Rosenzweig-Lipson et al., 1994). Further, no appreciable changes in any standard biomarkers measured through CBC or metabolic panels were observed during treatment with l-MA or d-MA, which corroborates previous reports in humans whereby acute l-MA administration was considered safe with limited cardiovascular risk (Mendelson et al., 2006, 2008; Dufka et al., 2009). These data highlight that, under these experimental conditions, chronic treatment with l-MA does not produce overt changes in standard health biomarkers, despite concern for the effects of monoaminergic drugs on metabolic and peripheral processes (Bakir and Dunea, 1996; Matsumoto et al., 2014).
Conclusions
The present study demonstrates that l-MA exhibits a lesser abuse liability profile relative to common stimulants (i.e. cocaine, d-MA, and MCAT). Further, a dose of l-MA known to decrease cocaine self-administration was well tolerated during chronic dosing with fewer unwanted effects compared with d-MA. This, combined with l-MA’s shared cocaine and d-MA–like subjective effects, suggest that it may be a promising candidate as an agonist therapy that would maintain treatment compliance. Future work aimed at clarifying the pharmacological mechanisms underlying l-MA’s distinct behavioral profile and the extent of tolerance to its therapeutic effects over extended treatment will be crucial for optimizing its use as an agonist therapy for stimulant use disorders.
Acknowledgments
The authors thank Olga Smirnova, Nathaniel Osiris, and Kevin Costa for assistance with conducting these studies. The authors thank Drs. Fernando B. de Moura and Alison Wakeford for helpful comments on an earlier version of the manuscript and Dr. Jack Bergman for thoughtful discussions about medications for substance use disorder.
Authorship Contributions
Participated in research design: Blough, Kohut.
Conducted experiments: Jacobs, Kohut.
Contributed new reagents or analytic tools: Blough.
Performed data analysis: Jacobs, Kohut.
Wrote or contributed to the writing of the manuscript: Jacobs, Blough, Kohut.
Footnotes
- Received February 2, 2021.
- Accepted May 10, 2021.
This work was funded by National Institutes of Health National Institute on Drug Abuse [Grants DA002519, DA039306, DA12970, and DA048150].
None of the authors have actual or perceived conflicts of interests with the contents of the article.
Abbreviations
- CBC
- complete blood count
- CUD
- cocaine use disorder
- DA
- dopamine
- d-MA
- d-methamphetamine
- FR
- fixed ratio
- inj
- injection
- l-MA
- l-methamphetamine
- MCAT
- methcathinone
- NE
- norepinephrine
- TO
- time-out
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}