


Abstract
Nephrotic syndrome (NS) is associated with metabolic perturbances including profound dyslipidemia characterized by hypercholesterolemia and hypertriglyceridemia. A major underlying mechanism of hypertriglyceridemia in NS is lipoprotein lipase (LPL) deficiency and dysfunction. There is emerging evidence that elevated angiopoietin-like protein 3 (ANGPTL3), an LPL inhibitor that is primarily expressed and secreted by hepatocytes, may be in part responsible for these findings. Furthermore, there is evidence pointing to the contribution of ANGPTL3 to the pathogenesis of proteinuria in NS. Therefore, we hypothesized that inhibition of hepatic ANGPTL3 by RNA interference will ameliorate dyslipidemia and other symptoms of NS and pave the way for a new therapeutic strategy. To this end, we used a subcutaneously delivered, GalNAc (N-Acetylgalactosamine)-conjugated small interfering RNA (siRNA) to selectively target and suppress liver Angptl3 in rats with puromycin-induced NS, which exhibits clinical features of NS including proteinuria, hypoalbuminemia, hyperlipidemia, and renal histologic abnormalities. The study demonstrated that siRNA-mediated knockdown of the liver Angptl3 relieved its inhibitory effect on LPL and significantly reduced hypertriglyceridemia in nephrotic rats. This was accompanied by diminished proteinuria and hypoalbuminemia, which are the hallmarks of NS, and significant attenuation of renal tissue inflammation and oxidative stress. Taken together, this study confirmed the hypothesis that suppression of Angptl3 is protective in NS and points to the possibility that the use of RNA interference to suppress hepatic Angptl3 can serve as a novel therapeutic strategy for NS.
SIGNIFICANCE STATEMENT The current standard of care for mitigating nephrotic dyslipidemia in nephrotic syndrome is statins therapy. However, the efficacy of statins and its safety in the context of impaired kidney function is not well established. Here, we present an alternate therapeutic approach by using siRNA targeting Angptl3 expressed in hepatocytes. As the liver is the major source of circulating Angptl3, siRNA treatment reduced the profound hypertriglyceridemia in a rat model of nephrotic syndrome and was also effective in improving kidney and cardiac function.
Introduction
Nephrotic syndrome (NS) is associated with significant hyperlipidemia and substantial alterations in lipid and lipoprotein metabolism (Vaziri, 2016a; Moradi and Vaziri, 2018). Serum cholesterol, triglycerides (TG), and the apolipoprotein B–containing lipoproteins, including very low-density lipoprotein (VLDL), intermediate-density lipoprotein cholesterol, and low-density lipoprotein cholesterol (LDL-C), are significantly elevated in NS (Joven et al., 1990). Although altered biosynthesis of fatty acids and cholesterol play a minor role in the pathogenesis of these abnormalities, their impaired clearance is a key cause of the NS-induced dyslipidemia (Kaysen et al., 1992; Agrawal et al., 2018). The latter results in significant elevations of serum triglycerides (Dewey et al., 2017) and triglyceride-rich lipoproteins including VLDL and intermediate-density lipoprotein (Kaysen and de Sain-van der Velden, 1999; Vaziri, 2016b). The major underlying cause of these abnormalities is lipoprotein lipase (LPL) deficiency and dysfunction (Garber et al., 1984; Vaziri, 2016a; Agrawal et al., 2018). LPL is the rate-limiting enzyme that mediates lipolysis of chylomicrons and VLDL and release of their fatty acids for uptake by adipocytes for storage of energy and the muscle and other tissues for production of energy. NS is associated with significant reduction of LPL activity and marked reductions of heparin-releasable and intracellular LPL protein (Liang and Vaziri, 1997a,b; Shearer and Kaysen, 2001; Sato et al., 2002; Vaziri et al., 2012). These abnormalities are present despite normal LPL mRNA expression in adipose tissue, skeletal muscle, and myocardium, therefore indicating the presence of post-transcriptional and translational mechanisms of NS-induced LPL dysfunction (Liang and Vaziri, 1997b; Sato et al., 2002).
Angiopoietin-like protein 3 (ANGPTL3), secreted by the liver (Conklin et al., 1999), is an inhibitor of LPL, and endothelial lipase has a major role in lipid metabolism (Musunuru et al., 2010; Wu et al., 2020). Homozygous loss of ANGPTL3 function causes familial combined hypolipidemia characterized by low plasma levels of triglycerides (Dewey et al., 2017), high-density lipoprotein (HDL) cholesterol, and LDL-C and a decreased risk of coronary artery disease (Romeo et al., 2009; Musunuru et al., 2010; Minicocci et al., 2012; Robciuc et al., 2013; Stitziel et al., 2017). After an effort to replicate this advantageous human phenotype for treating dyslipidemia, several therapeutic modalities have been developed. Antibodies targeted toward circulating ANGPTL3 lower plasma TG, LDL-C, and HDL cholesterol in mice (Gusarova et al., 2015), monkeys (Gusarova et al., 2015), and humans (Dewey et al., 2017; Gaudet et al., 2017). RNA interference–mediated suppression of liver Angptl3 transcript has been shown to have a similar effect in several mouse models and humans (Graham et al., 2017; Xu et al., 2018). ANGPTL3 inhibition has thus emerged as a highly promising alternative therapeutic option for mitigating hyperlipidemia in various lipoprotein disorders (Arca et al., 2013; Kersten, 2019).
Recent studies have identified ANGPTL3 and ANGPTL4 as proteins that, by inhibiting LPL activity, play a significant role in the pathogenesis of NS-induced LPL deficiency (Clement et al., 2014; Vaziri and Moradi, 2014; Kersten, 2017). ANGPTL3 is a secreted protein that is mainly expressed in hepatocytes (Conklin et al., 1999). Recent studies have found that the Angptl3 knockdown via siRNA or use of knockout animal models markedly ameliorated these findings, improved structure and integrity of podocytes, and decreased proteinuria (Lin et al., 2013; Dai et al., 2015, 2019; Liu et al., 2015). NS, in addition to heavy proteinuria and glomerular disease (as evidenced by heavy proteinuria, glomerulosclerosis), also results in tubulointerstitial disease marked by inflammation and oxidative stress. The latter abnormalities are worsened and contributed to by the significant dyslipidemia of NS including profound hypertriglyceridemia. It is well known that free fatty acids can cause cellular toxicity and contribute to oxidative stress and inflammation (Hoffman and Levy, 1989; Han et al., 2013). Therefore, any therapy that would ameliorate glomerular damage and tubulointerstitial disease would also reduce oxidative stress and inflammation. Our evaluations were constructed to show not only that proteinuria was reduced but also that this salutary effect is accompanied by reduced markers of inflammation and oxidative stress.
Given these findings and that the liver is the major source of circulating ANGPTL3, we hypothesized that Angptl3 mRNA knockdown, specifically in the hepatocytes by using GalNAc-conjugated siRNA (Rajeev et al., 2015), may not only ameliorate NS-associated hypertriglyceridemia but also have a salutary effect on proteinuria and renal function. Therefore, we evaluated the impact of Angptl3 siRNA treatment in a rat model of puromycin-induced nephrotic syndrome.
Material and Methods
In Vivo.
All animal experiments were approved by the University of California Irvine Institutional Committee for the Use and Care of Experimental Animals. Male Sprague-Dawley rats with an average body weight of 180–200 g (Charles River Laboratories, Raleigh, NC) were used in this study. Animals were housed in a climate-controlled vivarium with 12-hour day/night cycles and access to food and water ad libitum. At the beginning and the final week of the study, the animals were placed in metabolic cages for a 24-hour urine collection, and systolic blood pressure was measured by tail plethysmography as described previously (Vaziri et al., 2002). NS was induced in a randomly selected group of animals via sequential intraperitoneal injections of puromycin aminonucleoside on day 1 (130 mg/kg) and day 14 (60 mg/kg) (Hoffman and Levy, 1989; Han et al., 2013). The rats assigned to the control group received saline injections. Subsequently, the NS animals were randomized to either weekly subcutaneous injections of 1× PBS (vehicle control) or 10 mg/kg of Angptl3 siRNA and followed for 4 weeks. The siRNA was designed and synthesized as published elsewhere (Nair et al., 2014) to target rat Angptl3 mRNA NM_001025065.1. It targets the sequence at position 265–287, 5′- AAAAAGACTGATCAAATATGTTG -3′, with a single mismatch at position 6.
At the end of the study, body weight, tail arterial blood pressure, and 24-hour urinary protein excretion were measured. The animals were then fasted overnight and sacrificed under general anesthesia between the hours of 9 and 11 AM. Blood was collected using cardiac puncture, and subsequently, relevant tissues including kidney and heart were immediately removed and processed for histologic evaluation including electron microscopy and then frozen and stored at −70°C for biochemical analysis.
For euthanasia, the animals were placed into a sealed anesthesia induction chamber under 5% isoflurane (Piramal Clinical Care, Bethlehem, PA)/oxygen gaseous mixture to induce sedation, which was maintained at 2%–4%.
Urinary protein was measured using the Rat Urinary Protein Assay Kit, catalog number 9040 (Chondrex, Inc., Redmond, WA). Serum urea was measured by the QuantiChromTM Urea Assay Kit, catalog number DIUR-500 (BioAssay Systems, Hayward, CA). Urinary creatinine was measured by the QuantiChrom Creatinine Assay Kit, catalog number DICT-500 (BioAssay Systems). Serum cholesterol and triglyceride were measured by the Cholesterol Fluorometric Assay Kit, item number 10007640, and the Triglyceride Colorimetric Assay Kit, item number 10010303 (Cayman Chemical, Ann Arbor, MI). HDL cholesterol was measured by the HDL-Cholesterol Assay Kit, catalog number STA-394 (Cell Biolabs, Inc., San Diego, CA). Serum LDL was measured by the Rat LDL-Cholesterol Assay Kit, catalog number 79960 (Crystal Chem, Elk Grove Village, IL). Serum urea was measured using the Urea Assay Kit, catalog number DIUR-500, and serum albumin was measured using the QuantiChromTM BCP Albumin Assay Kit, catalog number SIAP-250 (BioAssay Systems). All kits were used according to the manufacturer instructions. Serum creatinine was measured by capillary electrophoresis using the PA800 Plus Pharmaceutical Analysis System (Beckman Coulter) from the George M. O’Brian Kidney Research Core at University of Texas Southwestern Medical Center.
Histologic Analysis.
The kidneys were removed after sacrifice and fixed with 10% formalin solution. Histologic sections were stained with hematoxylin and eosin and Masson trichrome. Briefly, 30 glomeruli from each trichrome-stained section were evaluated for glomerulosclerosis. Any degree of glomerulosclerosis was counted as positive, and the total number of affected glomeruli were counted counted for each animal. The mean number of sclerotic glomeruli was calculated for each experimental group. Tubulointerstitial damage marked by inflammatory cell infiltration, fibrosis, tubular dilatation, and atrophy was evaluated semiquantitatively according to the extension of the damaged area in the renal cortex: 0, normal; grade 1, <10%; grade 2, 10%–25%; grade 3, 25%–50%; grade 4, 50%–75%; and grade 5, 75%–100%.
Electron Microscopy.
Samples were fixed in 4% glutaraldehyde for 3 to 4 hours; tissues were then cut into sections of 1.2–2.0 mm3 and fixed in osmium tetroxide for 1 hour. After rinsing with distilled water, the tissues were dehydrated using 50%, 75%, 95%, and 100% ethanol and then placed in propylene oxide followed by 1:1 of propylene and embedded in Epon media. Sections measuring approximately 1000 μm were stained with methylene blue to identify the appropriate segments to be used, and subsequently 100 μm sections were stained with uranyl acetate and lead acetate and examined using a Tecnai Spirit electron microscope at 80 KV.
Western Blot Analyses.
Cytoplasmic extracts of the renal tissue were prepared as described previously (Sakurai et al., 1996). The proteins of interest in the cytoplasmic fractions of the kidney tissue were measured by Western blot analysis as previously described (Kim and Vaziri, 2010; Aminzadeh et al., 2012) using the following antibodies. Rabbit against rat NADPH-oxidase 4 (NOX4) (catalog number ab133303) and transforming growth factor-β (TGF-β) (catalog number ab215715) were purchased from Abcam (Cambridge, MA). Rabbit antibody against monocyte chemoattractant protein-1 (MCP-1) (catalog number NBP1-07035) were purchased from Novus Biologicals. Nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) (catalog number ab16502) was obtained from Cell Signaling. Mouse antibody against α-smooth muscle actin (catalog number 701457) was obtained from Invitrogen. Mouse antibody against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (catalog number TH270379) was purchased from Invitrogen and used for the measurement of loading control reference.
Lipoprotein Lipase Activity.
Lipoprotein lipase activity in hepatic, muscle, and white adipose tissues was determined using an assay kit purchased from Abcam (catalog number ab204721) following the manufacturer’s protocol. Briefly, 50 mg of tissue samples were homogenized with 200 µl of ice-cold PBS. Samples were then centrifuged at 4°C at 10,000g for 10 minutes, and supernatant was collected in a new tube. In total, 50 μl of samples was added to a 96-well plate followed by 50 µl of substrate (in DMSO, included in the kit). The plate was incubated at 37°C for 10 minutes, protected from light, and measured using a microplate reader at Excitation/Emission = 482/515 nm every 10 minutes for 1 hour.
Real-Time Polymerase Chain Reaction Analysis.
Total RNA was isolated from liver tissue using the miRNeasy kit (Qiagen) following manufacturer’s protocols. In total, 1 μg of RNA from each sample was reverse-transcribed using the High-Capacity Reverse Transcription Kit (Invitrogen). Quantitative real-time PCR was performed on the cDNA using the Roche LightCycler and the LightCycler 480 master mix (Roche). All experimental samples were analyzed using TaqMan assays (Invitrogen) to detect Angptl3 (Rn01433283_m1) and normalized with the expression level of a reference gene, Gapdh (4352339E).
Statistical Analysis.
Data are presented as means ± S.D. One-way ANOVA and multiple comparisons were performed using GraphPad Prism 8.4 (GraphPad Software, San Diego, CA). Tukey’s post-test was used to determine differences between the groups. Grubbs test was used to analyze potential outliers. P values less than 0.05 were considered significant.
Results
Angptl3 siRNA Treatment Reduced Liver Transcript Angptl3 Levels and Correspondingly Increased LPL Activity.
To study the effects of Angptl3 siRNA on NS, we used a previously established rat model of puromycin-induced NS. The rats were subcutaneously treated with siRNA targeting Angptl3. Compared with the control group, the placebo-treated nephrotic group exhibited a trend for increased hepatic tissue Angptl3 gene expression. Angptl3 siRNA administration significantly decreased hepatic tissue Angptl3 mRNA expression by 86.2% ± 7.4% (Fig. 1A). This level of Angptl3 transcript knockdown, using this particular siRNA duplex, has routinely shown significant decrease in circulating ANGPTL3 in various rodent models (Wu et al., 2020). As expected, this led to a disinhibition of LPL observed by an increase in LPL activity in the serum (Fig. 1B). In addition, Angptl3 siRNA treatment significantly increased LPL activity in tissues including hepatic, muscle, and adipose tissues (Fig. 1, C–E) when compared with the vehicle-treated controls.

Impact of Angptl3 siRNA treatment on Angptl3 transcript levels and LPL activity. Liver Angptl3 transcript levels and LPL activity in the indicated tissues were measured to assess the efficacy of Angptl3 siRNA treatment in vehicle-treated controls (CTL), nephrotic syndrome, and Angptl3 siRNA–treated NS + siRNA animals. (A) Angptl3 transcript levels were decreased in the Angptl3 siRNA–treated group (n ≥ 8). Lipoprotein lipase activity was reduced in nephrotic animals and recovered in Angptl3 siRNA treatment group (NS + siRNA) in (B) serum (n ≥ 8), (C) hepatic (n ≥ 6), (D) muscle (n ≥ 7), and (E) adipose tissues (n ≥ 7). Data are presented as means ± S.D., *P < 0.05; **P < 0.01; ***P < 0.001, ****P < 0.0001
Angptl3 siRNA Treatment Attenuated Hyperlipidemia and Proteinuria.
To assess the effects of the Angptl3 siRNA treatment, we looked at the well established markers of NS. Puromycin-treated nephrotic animals exhibited all the hallmarks of NS and dyslipidemia as observed by heavy proteinuria; significantly increased serum cholesterol, LDL, and triglyceride levels; and decreased serum albumin concentrations. In addition, the nephrotic group had modest increases in systolic blood pressure and serum urea concentration and reduced creatinine clearance (Table 1). Treatment with Angptl3 siRNA in NS animals resulted in a remarkable and statistically significant lowering of serum triglyceride levels (307.5 ± 88.9 vs. 89.7 ± 78.4 mg/dl). This was accompanied by partial improvement of hypoalbuminemia and increased creatinine clearance. In accordance with these markers of improved kidney function, the Angptl3 siRNA–treated animals also showed decreased proteinuria at week 4 post–puromycin treatment, when the maximum effect of the puromycin is observed in this model (Fig. 2). In addition, there was a modest but significant improvement in arterial blood pressure and decreased left ventricular hypertrophy in the Angptl3 siRNA–treated group, indicating potential for a modest betterment of cardiac function, probably as a direct effect of improved kidney function.
Impact of Angptl3 siRNA treatment on NS markers
Angptl3 siRNA treatment attenuated hypertriglyceridemia and improved pathologic kidney markers in puromycin-treated NS animals. Data are presented as means ± S.D.

Impact of Angptl3 siRNA treatment on urinary protein excretion kidney dysfunction in NS causes increased protein excretion and is a hallmark of the disease. Urinary protein was measured in vehicle-treated controls (CTL), nephrotic syndrome, and Angptl3 siRNA–treated NS + siRNA animals. (A) Time course of 24-hour urinary protein excretion post–puromycin treatment showing increased urinary protein excretion, which was mitigated by the treatment with Angptl3 siRNA. (B) Individual animal data of 24-hour urinary protein at week 4 post–puromycin treatment. Data are presented as means ± S.D. (n ≥ 8). *P < 0.05 vs. CTL, †P < 0.05 NS + siRNA vs. N, **P < 0.01, ****P < 0.0001
Angptl3 siRNA Treatment Partially Protected against Pathologic Renal Injury in NS Rats.
We next observed the effect of Angplt3 siRNA treatment on kidney injury. Puromycin treatment induced sclerosis of the glomeruli. Angptl3 siRNA treatment partially attenuated the glomerulosclerosis score (Fig. 3, A and B). Further evaluation by electron microscopy confirmed moderate podocyte swelling and effacement in the NS group (Fig. 3C(b,e)) when compared with controls (Fig. 3C(a,d). When treated with Angptl3 siRNA, the podocyte swelling and effacement were reduced (Fig. 3C(c,f). Puromycin treatment also induced tubulointerstitial injury seen with tubular atrophy, tubular dilation, and interstitial fibrosis. However, there was no resolution of the tubulointerstitial abnormalities between the vehicle- and siRNA-treated NS animals (Fig.3D) . This is consistent with mild renal injury observed in this animal model and possible renal recovery after week 4 when the effects of puromycin are waning.
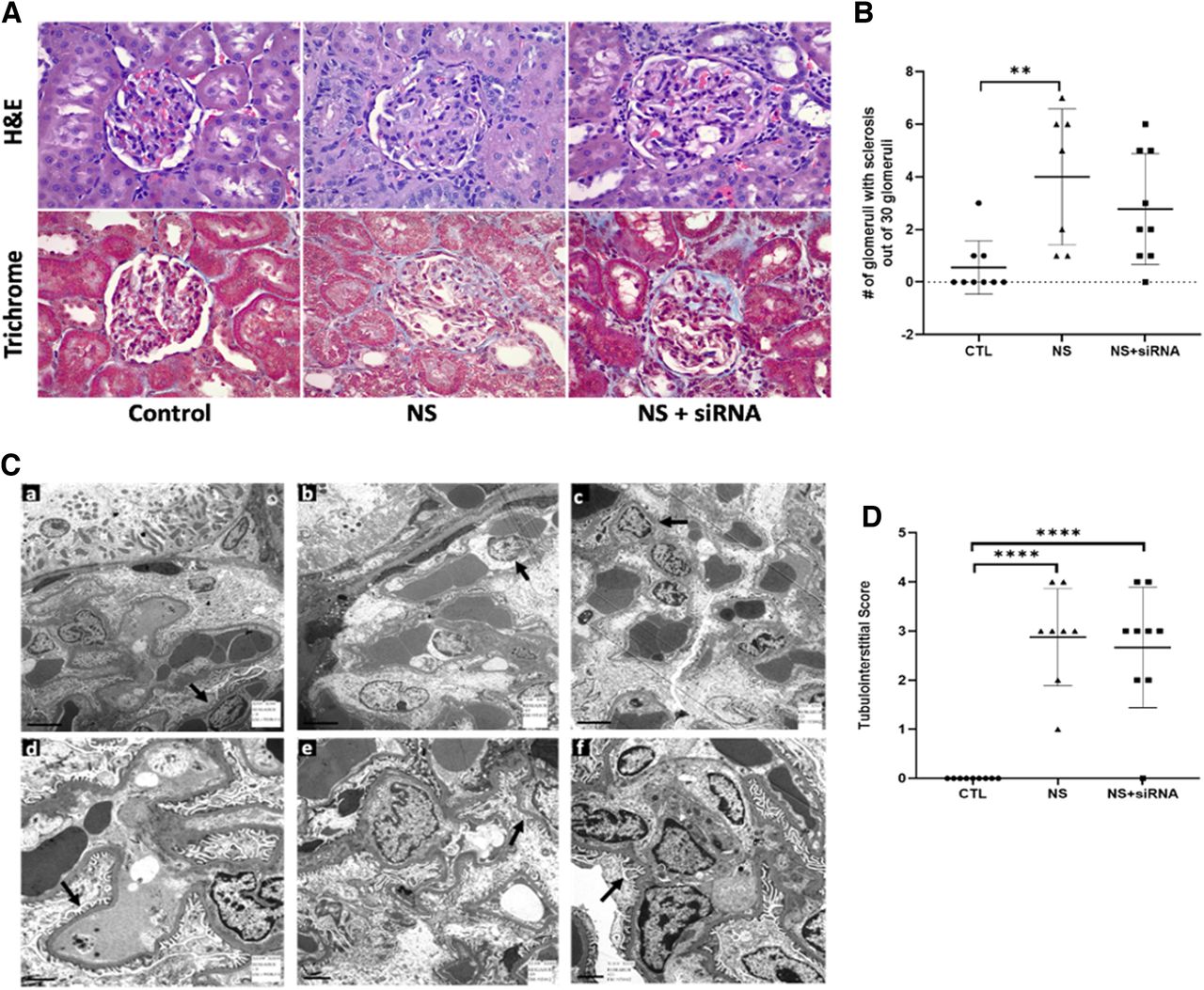
Impact of Angptl3 siRNA treatment on glomerulosclerosis and podocyte morphology. NS causes pathologic injury to kidney tissue. Tissue architecture was assessed by histologic staining and electron microscopy methods in vehicle-treated controls (CTL), nephrotic syndrome, and Angptl3 siRNA–treated NS + siRNA animals. (A) Photomicrographs show representative glomeruli in the 3 groups. (B) NS was associated with increased glomerulosclerosis, and treatment with Angptl3 siRNA improved this finding. Data are presented as means ± S.D. (n ≥ 7), **P < 0.01. (C) Electron microscopy revealed moderate podocyte swelling and effacement, which was reduced upon Angptl3 siRNA treatment. (a) Normal podocytes in control group (arrow). (b) Moderate podocyte swelling in NS group (arrow). (c) Mild podocyte swelling in NS + siRNA group (arrow). (d) Normal foot processes in control group (arrow). (e) Foot process effacement in approximately 20% of foot processes in NS group (arrow). (f) Foot process effacement in 5% of foot processes in NS + siRNA group (arrow). n = 3 animals analyzed per group. (D) Tubulointerstitial score was increased in NS animals; however, there was no improvement with Angptl3 siRNA treatment at the time point tested. Data are presented as means ± S.D. *P < 0.05; **P < 0.01; ****P < 0.0001.
Angptl3 siRNA Treatment Improved the Dysregulation of Inflammatory, Oxidative Stress, and Fibrotic Pathways Seen in NS Kidneys.
Molecular pathways of inflammation, oxidative stress, and fibrosis are upregulated in NS and contribute to kidney dysfunction. We next sought to examine the effect of Angptl3 siRNA on some of these molecular components of pathogenesis. Compared with the control group, the NS rats exhibited increased NF-κB protein abundance in the renal tissue (P < 0.01), pointing to activation of the inflammatory pathway. Angptl3 siRNA treatment reversed upregulation of NF-κB in the kidney tissues of NS rats (Fig. 4A). Compared with the control group, the NS group had significant increases in protein abundance of MCP-1 and NOX4 in their renal tissue. Upregulations of the renal tissue MCP-1 and NOX4 were ameliorated with Angptl3 siRNA treatment (Fig. 4, B and C). Furthermore, renal tissue from the NS animals exhibited upregulations of the profibrotic proteins TGF-β (P < 0.01) and α-smooth muscle actin (P < 0.01), which were markedly attenuated with siRNA treatment (Fig. 4, D and E).

Impact of Angptl3 siRNA treatment on renal injury pathways. NS triggers kidney injury by activating pathologic cellular pathways. Markers of inflammation, oxidative stress, and fibrosis were assessed using Western blotting in vehicle-treated controls (CTL), nephrotic syndrome, and Angptl3 siRNA–treated NS + siRNA animals. Protein abundance of (A) NF-κB (n = 6), (B) MCP-1 (n = 10), (C) NOX4 (n = 6), (D) TGF-β (n = 8), and (E) α-smooth muscle actin (n = 10) showed resolution of injury pathways upon siRNA treatment. Data are presented as means ± S.D. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.
Discussion
Heavy proteinuria, which is a hallmark of NS, is also associated with various metabolic complications including hypoalbuminemia and dyslipidemia (Vaziri et al., 2003; Vaziri, 2016a). One of the defining characteristics of NS-associated dyslipidemia is hypertriglyceridemia and significantly elevated serum levels of triglyceride-rich lipoproteins (Agrawal et al., 2018). This hyperlipidemia contributes to an increase in cardiovascular disease, and adults with NS have an increased risk for myocardial infarction (Ordonez et al., 1993; Agrawal et al., 2018). Despite their subpar efficacy on triglyceride lowering, statins are used to treat hyperlipidemia in NS (Wheeler, 2001). A major cause of hypertriglyceridemia has been shown to be LPL deficiency and dysfunction, which has been attributed to many different factors, chief among them being elevated levels of ANGPTL3 and ANGPTL4 proteins (Mace and Chugh, 2014; Vaziri and Moradi, 2014; Jiang et al., 2019). These proteins have an inhibitory effect on LPL, and their elevated level leads to impairment of fatty acid and TG clearance and significant hypertriglyceridemia. The latter point was demonstrated in a study by Chugh and colleagues, who showed that increased circulating levels of ANGPTL4 caused significant LPL inhibition leading to hypertriglyceridemia, although elevated renal levels of this protein caused decreased urinary protein excretion (Clement et al., 2014). These observations have led to the intriguing possibility of utilizing Angptl3 targeting therapies to prevent dyslipidemia of NS and thus ameliorate symptoms of NS (Mace and Chugh, 2014; Jiang et al., 2019). In accordance with this idea, other investigators have shown that deletion or decreased expression of Angptl3 ameliorates proteinuria and protects renal structure and function in animal models of nephrotic syndrome (Dai et al., 2015, 2019; Liu et al., 2015). In addition, the feasibility of an siRNA-mediated approach to mitigate NS is highly supported by other successful and safe siRNA treatments for dyslipidemia, such as the proprotein convertase subtilisin/kexin type 9 inhibitor inclisiran, which was developed using similar siRNA technology (Fitzgerald et al., 2017; Ray et al., 2020).
In the current study, we found that administration of a hepatocyte-targeted Angptl3-specific siRNA formulation in rats with puromycin-induced NS was associated with a significant decrease in serum TGs but no change in serum cholesterol or LDL levels. This is consistent with the fact that LPL does not play a major role in elevated cholesterol levels in NS, as the latter is mainly driven by LDL receptor deficiency and elevated serum proprotein convertase subtilisin/kexin type 9 levels (Jin et al., 2014; Liu and Vaziri, 2014; Vaziri, 2016a). We also found a significant improvement in LPL activity in the fat, muscle, and liver tissues of animals with NS treated with Angptl3 siRNA, confirming the mechanism through which Angptl3 inhibition improves TG metabolism. Interestingly, treatment with Angptl3 siRNA was also associated with an improvement in serum albumin concentration and a significant decrease in urine protein excretion, supporting the amelioration of proteinuria in these animals. Given the evidence that elevated fatty acid and triglyceride levels may cause renal damage and in light of the significant lowering of serum TGs after Angptl3 siRNA treatment, the improvement in proteinuria could be the result of reduced lipotoxicity (Nosadini and Tonolo, 2011; Martinez-Garcia et al., 2015; Escasany et al., 2019).
Interestingly, it has been shown that renal expression of Angptl3 is significantly upregulated in the setting of podocyte injury and nephrotic syndrome. Furthermore, altered expression of ANGPTL3 in the glomeruli of animals with nephrotic syndrome has been found to be associated with increased proteinuria and foot process effacement (Liu et al., 2015). Lin et al. (2013) further reported that exposure of podocytes to ANGPTL3 results in accelerated puromycin-induced loss of podocytes via their detachment and apoptosis, which was most likely mediated by the rearrangement of the F-actin. In fact, there was also a small improvement in serum markers of renal function (creatinine clearance), although the mild nature of renal injury in the early stages of this animal model limits the interpretation of these findings. Histologic analysis of renal tissue showed that animals treated with Angptl3 siRNA had decreased glomerulosclerosis on light microscopy and reduced podocyte effacement on electron microscopy. We also found decreased abundance of the markers of inflammation and fibrosis in the renal tissue of nephrotic animals treated with Angptl3 siRNA when compared with vehicle-treated controls.
However, this current study does not delineate additional mechanisms that could contribute to the protection of renal injury. These findings will need to be further delineated in future mechanistic studies. In summary, the data presented here support the hypothesis that inhibition of liver Angptl3 expression by an siRNA could ameliorate symptoms of NS and highlight the potential of RNA interference as a therapeutic alternative for NS.
Acknowledgments
The siRNA used in this study was generated at Alnylam Pharmaceuticals. We thank the various Alnylam personnel in the siRNA screening, validation, and synthesis groups for kindly providing us with this siRNA.
Authorship Contributions
Participated in research design: Vaziri, Haslett, Moradi, Soundarapandian.
Conducted experiments: Zhao, Goto, Khazaeli, Liu, Farahanchi.
Performed data analysis: Khanifar, Farzaneh.
Wrote or contributed to the writing of the manuscript: Zhao, Vaziri, Haslett, Moradi, Soundarapandian.
Footnotes
- Received July 29, 2020.
- Accepted December 4, 2020.
Y.Z. and M.G. contributed equally to this work.
This work was supported by Alnylam Pharmaceuticals by a grant to N.D.V.
M.S. and P.H. are employees and stockholders of Alnylam Pharmaceuticals. No author has an actual or perceived conflict of interest with the contents of this article.
Abbreviations
- Angptl3
- angiopoietin-like protein 3
- GAPDH
- glyceraldehyde 3-phosphate dehydrogenase
- HDL
- high-density lipoprotein
- LDL
- low-density lipoprotein
- LDL-C
- low-density lipoprotein cholesterol
- LPL
- lipoprotein lipase
- MCP-1
- monocyte chemoattractant protein-1
- NF-κB
- nuclear factor κ-light-chain-enhancer of activated B cells
- NOX4
- NADPH-oxidase 4
- NS
- nephrotic syndrome
- siRNA
- small interfering RNA
- TGF-β
- transforming growth factor-β
- VLDL
- very low-density lipoprotein
- Copyright © 2021 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}