


Abstract
The tachykinin NK2 receptor plays a key role in gastrointestinal motor function. Enteric neurons release neurokinin A (NKA), which activates NK2 receptors on gastrointestinal smooth muscle, leading to contraction and increased motility. In patients with diarrhea-predominant irritable bowel syndrome, the NK2 receptor antagonist ibodutant had a greater therapeutic effect in females than males. The present study aimed to determine whether gender influences the expression and activity of NK2 receptors in human colonic smooth muscle. In vitro functional studies were performed to examine the contractile responses of colonic muscle strips to NKA and the selective NK2 receptor agonist [Lys5,MeLeu9,Nle10]NKA(4-10). Contractions were also measured in the presence of ibodutant to determine its antagonistic potency. The signal transduction pathways coupled to NK2 receptor activation were investigated using second messenger inhibitors. Western blot and fluorescent immunohistochemistry were conducted to determine the protein expression and localization of NK2 receptors. NK2 receptor–mediated contractility was greater in females compared with males. When against NKA, ibodutant was more potent in females. NK2 receptor expression increased with age in females, but not in males. Phospholipase C–mediated signaling was less prominent in females compared with males, whereas Ca2+ sensitization via Rho kinase and protein kinase C appeared to be the dominant pathway in both genders. The distribution of NK2 receptors in the human colon did not differ between the genders. Overall, gender differences exist in the expression and activity of NK2 receptors in colonic smooth muscle. These gender distinctions should be considered in the therapeutic development of NK2 receptor agents.
SIGNIFICANCE STATEMENT The tachykinin NK2 receptor has been identified as a therapeutic target for the treatment of bowel and bladder dysfunctions. The present study has revealed gender-related variations in NK2 receptor activity, signaling transduction pathways, antagonist potency, and changes in expression with age. These factors may underlie the gender differences in the treatment of diarrhea-predominant irritable bowel syndrome with NK2 receptor antagonists. Our findings highlight that gender differences should be considered in the therapeutic development of NK2 receptor agents.
Introduction
Tachykinins are a family of peptides that regulate a broad range of biologic activities, including smooth muscle contraction, vasodilation, inflammation, and nociception. The major naturally occurring tachykinins are substance P (SP), neurokinin A (NKA), and neurokinin B, and they exhibit preferential affinity for tachykinin NK1, NK2, and NK3 receptors, respectively (Nakamura et al., 2011). Tachykinin receptors primarily couple to the Gαq/11 class of G proteins, which activate phospholipase C (PLC), leading to the formation of inositol 1,4,5-triphosphate (IP3) and the subsequent release of Ca2+ from intracellular stores (Regoli et al., 1994; Mizuta et al., 2008). These receptors may also activate Gαs proteins, leading to increased cAMP levels (Nakajima et al., 1992; Regoli et al., 1994; Perdona et al., 2019).
In the gastrointestinal tract, SP and NKA are primarily released from myenteric and submucosal neurons and are involved in the modulation of motor and secretory events (Shimizu et al., 2008; Steinhoff et al., 2014). Among the three tachykinin receptor subtypes, NK2 receptors have an important role in regulating human colonic motility. Numerous pharmacological studies using human colonic smooth muscle strips have revealed that the contractile effects of endogenous tachykinins are mediated by NK2 receptors (Croci et al., 1998; Cao et al., 2000; Liu et al., 2002, 2009; Nakamura et al., 2011; Tanaka et al., 2012). This is in line with the high mRNA expression levels of the tachykinin receptor 2 gene (encoding NK2 receptors) in human colonic smooth muscle (Liu et al., 2011). Furthermore, NK2 receptor activation has been shown to be responsible for the tachykinergic component of excitatory neurotransmission, and altered NK2 receptor function has been implicated in colonic motility disorders, such as inflammatory bowel disease, diverticular disease, and slow transit constipation in both adults and children (Menzies et al., 2001; Mitolo-Chieppa et al., 2001; Liu et al., 2002; Stanton et al., 2003; Maselli et al., 2004; Auli et al., 2008; Burcher et al., 2008; Alvarez-Berdugo et al., 2015).
Given their key involvement in human colonic motor function, NK2 receptors have been considered as possible drug targets for the development of effective therapies for diarrhea-predominant irritable bowel syndrome (IBS) (Lecci et al., 2004; Corsetti et al., 2015; Szymaszkiewicz et al., 2019). Specifically, the NK2 receptor antagonist ibodutant emerged as a promising candidate because of its high selectivity and potency for human NK2 receptors, long duration of action, and good oral bioavailability (Cialdai et al., 2006; Meini et al., 2009; Santicioli et al., 2013). However, clinical trials uncovered a gender distinction in the effectiveness of ibodutant, whereby dose-dependent improvements in symptoms reached statistical significance over placebo in female patients with IBS only (Tack et al., 2017).
The gender difference observed in the therapeutic outcome of ibodutant in diarrhea-predominant IBS sparks an interest in the gender-specific variations surrounding colonic NK2 receptors. Gender differences in the efficacy of ibodutant have also been reported in a guinea pig colitis model, in which colonic contractions induced by colorectal distension were inhibited by ibodutant at lower doses in females than those required in males (Bellucci et al., 2016). Previously, our findings revealed a gender-related distinction in the density of [125I]NKA binding sites (corresponding to the number of NK2 receptors) in human colonic smooth muscle, with a significantly higher binding capacity observed in males compared with females (Burcher et al., 2008). However, [125I]NKA binding affinities and the expression levels of NK2 receptor mRNA did not differ between the genders (Burcher et al., 2008).
Further investigation is required to explain the existence of gender distinctions in the efficacy of ibodutant and the abundance of NK2 receptor binding sites. The present study aims to clarify these observations by uncovering any further gender-related differences in the expression and activity of NK2 receptors in human colonic smooth muscle. Although NK2 receptors primarily couple to the Gαq/11-PLC-IP3-Ca2+ transduction cascade to mediate smooth muscle contraction, such as in the bladder and airway (Mizuta et al., 2008; Der et al., 2019), this signal transduction does not appear to be the main pathway used by NK2 receptors in human colonic smooth muscle (Warner et al., 2000; O’Riordan et al., 2001). Therefore, this study also aims to uncover the signal transduction pathways coupled to NK2 receptor activation in the human colon.
Materials and Methods
Human Colon Specimens.
Ascending and sigmoid colon segments were taken 10–20 cm away from the tumor site of patients with colorectal carcinoma undergoing surgical resection. Patients with obstructions or those who had received radiation therapy or chemotherapy were excluded from the study. The mean age of the specimens used was 58.5 years for females (age range: 31–85 years, n = 55) and 60.7 years for males (age range: 32–84, n = 43). There were no significant differences in the age of the female and male specimens used for the functional and expression studies. Specimens were collected from Hurstville Private, St George Private, and Prince of Wales Public and Private Hospitals. The research was performed in accordance with the Declaration of Helsinki and approved by the South Eastern Sydney Local Health District Human Research Ethics Committee, Sydney, NSW, Australia. The specimens were transported to the laboratory in ice-cold, carbogenated (95% O2 and 5% CO2) Krebs-Henseleit solution (NaCl 118 mM, KCl 4.7 mM, NaHCO3 25 mM, KH2PO4 1.2 mM, MgSO4 1.2 mM, CaCl2 2.5 mM, d-glucose 11.7 mM). Specimens were gently washed to remove fecal matter, and fat was trimmed. For the functional experiments, a portion of smooth muscle was dissected away from the other layers and stored in Krebs-Henseleit solution overnight at 4°C. The remaining tissue was processed for the expression studies as described below.
Functional Organ Bath Set-Up.
Colonic smooth muscle strips (each approximately 3 × 6 mm) were cut along the circular axis. Muscle strips were mounted under 1 g tension in 3.5-ml organ baths containing carbogenated Krebs-Henseleit solution at 37°C, as previously described (Liu et al., 2002). One end of each strip was fixed to a hook at the bottom of the bath, and the other end was connected to an isometric force transducer (Grass FTO3C, Warwick, RI). The change of muscle tension was recorded using the Polygraph computer program (E. Crawford, UNSW Sydney, NSW, Australia). Preparations were allowed to equilibrate for 30 minutes before exposure to antagonists and inhibitors and for 60 minutes before exposure to tachykinin agonists.
Contractile Responses to NK2 Receptor Agents.
During optimizing experimental conditions, it was shown that NKA- and [Lys5,MeLeu9,Nle10]NKA(4-10) (LMN-NKA)-induced human colonic contractile responses remained unaffected in the presence of the NK1 receptor antagonist CP9994 (100 nM) but were greatly reduced by the NK2 receptor antagonist SR 48968 (100 nM) (Supplemental Fig. 1, A and B). Furthermore, the selective NK1 receptor agonist [Pro9] SP (up to 10 µM), was virtually ineffective in contracting human colonic smooth muscle (Supplemental Fig. 1C). These results suggest that the influence of NK1 receptors is negligible. There was no evidence for the involvement of enteric neuron transmission, as atropine (1 µM) showed no effect to NKA and LMN-NKA (Supplemental Fig. 2). Thus, the subsequent experiments were conducted without the presence of an NK1 antagonist nor atropine.
A single concentration-response curve to NKA or the selective NK2 receptor agonist LMN-NKA (0.001–10 µM) was obtained from each strip, using 60-minute intervals to avoid desensitization. Washing occurred twice between each concentration of agonist. Acetylcholine (ACh, 10 mM) was used as an internal standard and applied at the start and end of each concentration-response curve, as per previously published protocols (Warner et al., 2000; Liu et al., 2002; Burcher et al., 2008). In parallel experiments, contractile responses were measured in the presence of the selective NK2 receptor antagonist ibodutant (either 0.01, 0.1, or 1 µM) preincubated for 30 minutes before the addition of agonist.
Contractile Responses in the Presence of Second Messenger Inhibitors.
NKA and LMN-NKA were fixed at the submaximal concentration of 0.3 μM and were added at 60-minute intervals. Muscle strips were washed twice between each application of agonist. Female muscle strip desensitization was not observed under control conditions, whereas slight desensitization occurred in male strips. For both control and treated muscle strips, results were expressed as a percentage of the average of two initial contractile responses under control conditions (before inhibitors were applied). In treated muscle strips, preincubation with second messenger inhibitors occurred for 30 minutes using increasing concentrations (3, 30, 100, 300 µM) of either U73122 hydrate (PLC inhibitor), 2-aminoethoxydiphenyl borate (2-APB, IP3 receptor inhibitor), GF109203X [protein kinase C (PKC) inhibitor], or Y-27632 dihydrochloride (Rho kinase inhibitor). The inactive analog of U73122 hydrate, U-73343 (100 µM), did not produce an effect on the agonist-induced contractions.
Neuronal Tachykinin Release by Electrical Field Stimulation.
Muscle strips were suspended between two platinum rings at 1 g tension, and preparations were allowed to equilibrate for 30 minutes. Electrical field stimulation (EFS) was conducted using cumulative frequencies (0.5, 5, 10, 50 Hz) of 1-millisecond duration, in 80-V trains lasting 10 seconds (Liu et al., 2009). Each frequency of EFS was given 10 minutes apart. Immediately after each stimulation, 300-μl bath fluid was collected and stored at −80°C, and an equal volume of Krebs-Henseleit solution was added back to each bath. The amount of released NKA and SP was later analyzed using radioimmunoassay kits (RK-046-15 for NKA and RK-061-05 for SP; Phoenix Pharmaceuticals, Burlingame, CA).
Protein Expression of NK2 Receptors by Western Blot.
Total protein was extracted from frozen human sigmoid colonic smooth muscle using lysis buffer containing 1 mM EGTA, 10 mM Tris-HCl, and protease inhibitor cocktail tablets (Sigma-Aldrich, St Louis, MO). Protein samples (20 µg) were denatured at 95°C for 8 minutes and loaded into 50-µl wells of 10% Mini-PROTEAN precast gels (Bio-Rad Laboratories, Hercules, CA). Electrophoresis was conducted at 100 V for 1.5 hours. Protein transfer onto polyvinylidene difluoride membranes was carried out using the Trans-Blot Turbo Transfer Pack and System (Bio-Rad Laboratories) at 25 V for 7 minutes. Immunodetection was achieved by incubating with the anti-NK2 receptor antibody (ATR-002; Alomone Laboratories, Jerusalem, Israel) at a dilution of 1:1000 for 1 hour at room temperature. The specificity of the NK2 antibody was verified using corresponding control peptides, the same as previously confirmed (Delvalle et al., 2018), as well as by conducting positive and negative control Western blots. The anti–glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (G9545, 1:50,000; Sigma-Aldrich) was used as the internal control. After washing, the membranes were incubated with anti-rabbit IgG (whole molecule) peroxidase secondary antibody (A4914, 1:5000; Sigma-Aldrich) for 1 hour at room temperature. Visualization of protein bands was performed using SuperSignal West Pico PLUS Chemiluminescent Substrate (34577; Thermo Scientific, Waltham, MA), and densitometry analysis was carried out using Image Studio Lite 5.2 (LI-COR Biosciences, Lincoln, NE).
Localization of NK2 Receptors by Immunofluorescence.
Full-thickness human sigmoid colon samples were fixed in Zamboni’s solution, embedded in paraffin, sectioned, and mounted on poly-l-lysine–coated slides. The slides were deparaffinized with xylene, rehydrated with graded ethanol concentrations (100%, 70%), and washed in PBS (0.1 M, pH = 7.4). Antigen retrieval was performed at 110°C for 5 minutes using EnVision FLEX Target Retrieval Solution Low pH (Dako Omnis, Santa Clara, CA). Nonspecific binding sites were blocked by PBS containing 10% goat serum (G9023; Sigma-Aldrich) for 30 minutes. Slides were incubated overnight with primary antibodies diluted in Tris-buffered saline with 0.05% Triton (0.1 M, pH = 7.6) containing 2% serum. The anti-NK2 receptor antibody (ATR-002, 1:50; Alomone Laboratories) was double labeled with either the anti–α-smooth muscle actin (α-SMA) antibody (ab7817, 1:50; Abcam, Melbourne, VIC, Australia), the anti-cD117/c-kit antibody (AF332, 1:50; R&D Systems, Minneapolis, MN), the anti–HuC/D antibody (A21271, 1:200; Invitrogen, Carlsbad, CA), the anti–β-tubulin antibody (MMS435P, 1:100; Covance, Princeton, NJ), the anti-S100 antibody (ab4066, 1:50; Abcam), or the anti-GFAP antibody (ab4674, 1:100; Abcam). The next day, the slides were incubated with secondary antibodies diluted in Tris-buffered saline with 0.05% Triton for 2 hours at room temperature. The secondary antibodies used were goat anti-rabbit IgG Alexa Fluor 488 (ab150077; Abcam), goat anti-mouse IgG Alexa Fluor 594 (ab150116; Abcam), and goat anti-chicken IgG Alexa Fluor 488 (BA-9010; Vector Laboratories, Burlingame, CA). Slides were mounted in 4′, 6′-diamidino-2-phenylindole (DAPI; Invitrogen). Images were captured by the Neurolucida/Stereoinvestigator system (MBF Bioscience). Obvious visual differences were noted, but no quantitative statistical analysis was performed. Negative controls were conducted under the same conditions without primary antibodies and did not produce a signal.
Gene Expression of Second Messenger Molecules by Real-Time Polymerase Chain Reaction.
Human ascending and sigmoid colonic smooth muscle samples were frozen in RNAlater (R0901; Sigma-Aldrich). Total RNA was extracted by using the Trizol method (Invitrogen) and was purified by DNase treatment (3 U at 37°C, 20 minutes). Single-strand cDNA (sscDNA) was synthesized using the SuperScript III First-Strand Synthesis Supermix (Invitrogen) in a reaction mixture containing 2 µg RNA and 50 ng µl−1 random hexamers, at one cycle of 25°C for 10 minutes, 50°C for 50 minutes, and 85°C for 5 minutes. RNase H treatment occurred (2 U µl−1) at 37°C for 20 minutes. sscDNA was diluted to a working concentration of 500 ng µl−1.
Gene expression was determined by real-time PCR using the SYBR Select Master Mix (Applied Biosystems, Austin, TX). The primers were designed with Primer3, and primer sequences and PCR amplification efficiencies are listed in Supplemental Table 1. Each PCR reaction was performed in a volume of 10 µl containing 5 µl Master Mix, 10 µM forward and reverse primers, and 80 ng µl−1 sscDNA. The solutions were dispensed into 384-well plates by the Eppendorf epMotion 5075 Automated Pipetting System. GAPDH was used as a housekeeping gene. Inter-run comparisons were achieved using a designated calibrator (one colonic smooth muscle sample) in each real-time PCR assay. PCR amplification occurred for one cycle at 95°C (2 minutes), 40 cycles at 95°C (15 seconds), 60°C (15 seconds), and 68°C (20 seconds), followed by 10 minutes of gradual temperature elevation from 60 to 95°C for the melting curve analysis.
Data Analysis.
Data were analyzed using GraphPad Prism 7 (GraphPad Software, San Diego, CA), and statistical significance was achieved when P < 0.05. The results obtained from ascending and sigmoid colonic muscle strips did not differ, so data from both regions were combined in the analyses.
For the functional studies, data were expressed as tension (grams). Similar results were seen when the data were expressed as tension (grams) per strip weight (grams). However, ACh could not be used as a reliable internal standard because contractile responses varied between the experimental groups. Results are presented as means ± S.E.M. unless otherwise stated. Concentration-response curves were generated using nonlinear regression analysis to determine maximal contractile responses (Emax) and half maximal EC50 values. The data were analyzed by two-way ANOVA followed by Bonferroni multiple comparisons test. For the antagonist ibodutant, Schild plots were created, and pA2 values were taken as the x-intercept of the Schild regression analysis. The apparent pKB values of each individual antagonist concentration were also calculated using the equation: pKB = log (dose ratio − 1) − log [B], where [B] is the antagonist concentration (Arunlakshana and Schild, 1959; Kenakin, 2009). Data were analyzed using one-way ANOVA with Bonferroni’s multiple comparisons test.
For the NKA and SP release study, basal levels of NKA and SP were expressed as nanograms per strip weight (nanograms per gram), and comparisons of median values were made using Mann-Whitney analysis. EFS-induced release was expressed as a percentage of the basal level and presented as means ± S.E.M. Gender differences were assessed by two-way ANOVA with Bonferroni’s multiple comparisons test, whereas comparisons between basal and EFS-induced tachykinin levels were made at each EFS frequency using one-sample t test analysis.
For the expression studies, NK2 receptor protein levels were expressed as a percentage of GAPDH. The mRNA level of genes encoding second messengers was expressed as fold change normalized to GAPDH and relative to the calibrator, using the following equation: Fold change = 2−ΔΔCt, where ΔΔCt = [Ct(target) − Ct(GAPDH)]sample − [Ct(target) − Ct(GAPDH)]calibrator (Pfaffl, 2001). Median values were compared using Mann-Whitney analysis or the Kruskal-Wallis test for multiple comparisons. Spearman analysis was used for the correlation studies.
Drugs.
NKA was purchased from Abcam (ab120185), and LMN-NKA was custom synthesized by China Peptides (Pudong, Shanghai, China). Both were reconstituted in 0.01 M acetic acid with β-mercaptoethanol. ACh was purchased from Sigma-Aldrich (A6625) and reconstituted in water. Ibodutant was a gift from our collaborators at The Menarini Group (Florence, Italy) and was prepared in DMSO. The second messenger inhibitors 2-APB (1224), GF109203X (0741), and Y-27632 dihydrochloride (1254) were purchased from Tocris Bioscience (Noble Park, Victoria, Australia), and the inhibitor U73122 hydrate (U6756) was obtained from Sigma-Aldrich. The second messenger inhibitors were dissolved in DMSO. Aliquots of all reagents were stored at −20°C.
Results
Contractile Responses to NK2 Receptor Agonists.
The endogenous NK2 receptor ligand NKA and the selective NK2 receptor agonist LMN-NKA evoked prominent contractile responses in human colonic smooth muscle strips (Fig. 1). There were no gender differences in contractions produced by NKA (n = 11 females, 10 males) (Fig. 1A). On the other hand, LMN-NKA elicited contractile responses that were significantly greater in females (n = 11) compared with males (n = 11, **P = 0.0023, two-way ANOVA) (Fig. 1B). In line with this, the Emax of NKA did not vary between the genders (1.8 ± 0.27), whereas the Emax of LMN-NKA was significantly greater in females (2.0 ± 0.18) compared with males (1.2 ± 0.17, **P = 0.0029, unpaired t test). When comparing the Emax of NKA and LMN-NKA in males, values were close to statistical significance (P = 0.0742), suggesting that suppressed responses to LMN-NKA in male strips underlie the observed gender difference. There was no correlation between age and Emax for either agonist, in either gender. There were no gender differences in EC50 values. For NKA, the EC50 values and 95% confidence intervals were 65.63 nM (25–167) in females and 75.31 nM (28–200) in males. For LMN-NKA, the EC50 values and 95% confidence intervals were 87.06 nM (45–166) in females and 28.76 nM (10–78) in males.

Gender-related differences in NK2 receptor–mediated contractility. Contractile responses of female (n = 11) and male (n = 10 to 11) human colonic circular smooth muscle strips to (A) the endogenous NK2 receptor ligand NKA and (B) the selective NK2 receptor agonist LMN-NKA. Data are expressed as mean tension (grams) ± S.E.M. Significance by two-way ANOVA is indicated.
Potency of NK2 Receptor Antagonist Ibodutant.
Ibodutant antagonized the contractile responses elicited by NKA and LMN-NKA in the human colonic smooth muscle strips of females (n = 11) and males (n = 9–11) and produced a rightward shift of the agonist concentration-response curves (Fig. 2). Schild plots were generated to calculate pA2 values for ibodutant against both agonists, and its competitive antagonism property was confirmed, since ibodutant did not change the maximum contractilities of the agonists, and the slopes of Schild plots are not different from unity (Figs. 2 and 3; Table 1). The apparent pKB values of each individual antagonist concentration were also calculated and presented as mean values in Table 1.

Gender-specific distinctions in the potency of the selective NK2 receptor antagonist ibodutant. The effect of different concentrations of ibodutant on contractile responses induced by the endogenous NK2 receptor ligand NKA (left panel) and the selective NK2 receptor agonist LMN-NKA (right panel) in female (A and B) and male (C and D) human colonic circular smooth muscle strips. Data are expressed as a tension (grams), and curves are presented as a percentage of the mean maximum response for each concentration of ibodutant. n = 9–11 for each curve, and closed circles indicate means ± S.E.M.

The Schild plots generated to calculate pA2 values for ibodutant against the endogenous NK2 receptor ligand NKA (left panel) and the selective NK2 receptor agonist LMN-NKA (right panel) in female (A and B) and male (C and D) human colonic circular muscle strips (n = 11).
Summary of Schild plot results for ibodutant against NKA and the selective NK2 receptor agonist LMN-NKA in human colonic circular smooth muscle
pA2 values and slopes were generated from Schild plot and expressed as means ± S.E.M. All slopes were not significantly different from 1, and there was no difference between the slopes. Values in parentheses are the mean apparent pKB of individual ibodutant concentrations calculated using the equation pKB = log (dose ratio − 1) − log [B], where [B] is the antagonist concentration (Arunlakshana and Schild, 1959; Kenakin, 2009).
When against NKA, ibodutant displayed a significantly higher degree of antagonism in females (pA2 = 8.51 ± 0.38) compared with males (pA2 = 7.23 ± 0.21, *P < 0.05, one-way ANOVA) (Table 1). On the other hand, no gender differences were observed in the potency of ibodutant when against LMN-NKA (pA2 = 8.30 ± 0.36 for females and 8.26 ± 0.22 for males) (Table 1). Surprisingly, ibodutant appeared agonist-dependent in the male colon, being less potent against NKA compared with LMN-NKA (*P < 0.05, one-way ANOVA). These results were similar to the apparent pKB values calculated from the individual concentration of ibodutant (Table 1). There was no significant difference in the potency of ibodutant against the different agonists in female colonic muscle strips. The pA2 values of ibodutant obtained from our study were lower than that previously reported in the human colon (pKB = 9.1), and gender-related differences were not described (Santicioli et al., 2013). It is noted that in their study, the concentration-response curves to the NK2 receptor agonist [βAla8]NKA(4-10) were cumulatively constructed (Santicioli et al., 2013). Nevertheless, a 60-minute interval between each addition of the agonists was applied in our study to avoid desensitization. The different protocols may contribute to the discrepancies between the two studies.
Basal and EFS-Induced Release of NKA and SP.
NKA and SP released from human colonic smooth muscle strips were measured to uncover any gender differences that could be contributing to the observed distinctions in contractility (Fig. 4). The basal level of NKA did not vary between females (4.7 ± 2.0, n = 14) and males (2.8 ± 0.8, n = 15) (Fig. 4A). On the other hand, a higher amount of SP was spontaneously released under resting conditions from muscle strips of females (5.5 ± 1.0, n = 10) compared with those of males (2.6 ± 0.4, n = 9, *P = 0.0279, Mann-Whitney test) (Fig. 4B).

Gender-related differences in the neuronal release of tachykinin peptides. (A and B) Basal release of NKA and SP from female (n = 14 and 10, respectively) and male (n = 15 and nine, respectively) human colonic smooth muscle strips. Horizontal lines denote medians. Significance by Mann-Whitney test is indicated, where *P = 0.0279. (C and D) The release of NKA and SP in response to increasing frequencies of EFS. Data are expressed as a percentage of the basal amount released and are presented as means ± S.E.M. One-sample t test analysis of EFS-induced tachykinin release compared with basal level is indicated by *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.
The neuronal release of NKA and SP elicited by cumulative frequencies of EFS was then recorded and expressed as a percentage of the basal level. EFS-induced release of NKA (Fig. 4C) and SP (Fig. 4D) was significantly greater than basal amounts at 5 and 10 Hz for both genders (one-sample t test). At 50 Hz, there was a significant increase in the amount of NKA released from male smooth muscle only (*P = 0.0467, one-sample t test) (Fig. 4C). Interestingly, SP release in response to 0.5 Hz was significantly lower than basal levels in male strips (****P < 0.0001, one-sample t test), and a trend of decrease was also observed in female strips (P = 0.0934) (Fig. 4D). Overall, there were no significant differences in EFS-induced tachykinin release between the genders.
NK2 Receptor Protein Expression.
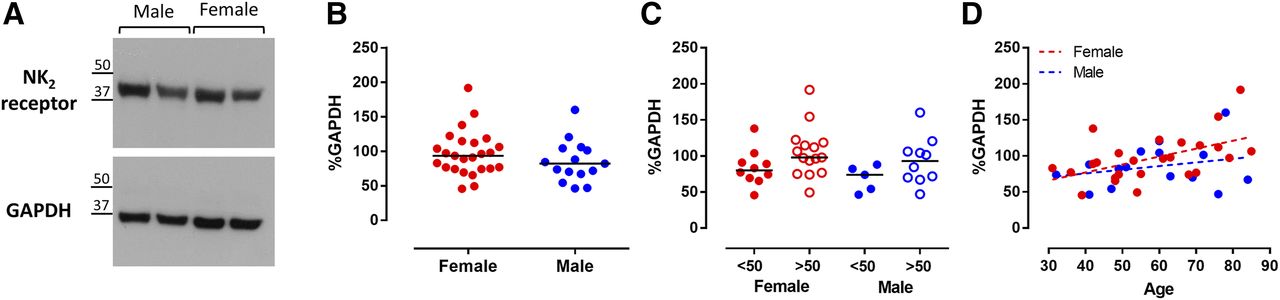
Western blot and densitometry analysis of human colonic sigmoid smooth muscle were performed to uncover any gender differences in the protein expression of NK2 receptors (Fig. 5). The NK2 receptor protein band was consistent with an ∼45-kDa protein size, as indicated by the manufacturer (Fig. 5A). Data are expressed as a percentage of GAPDH levels (∼36-kDa protein size). There was no difference in NK2 receptor protein expression between females (n = 25) and males (n = 15) (Fig. 5B). There was a noticeable trend increase in the expression of the NK2 receptor in specimens older than 50 years of age compared with younger ones (Fig. 5C). Correlation analysis revealed an increased NK2 receptor expression with age in females (r = 0.5023, *P = 0.0105, Spearman correlation test), but not in males (r = 0.127, P = 0.6499) (Fig. 5D).

Western blot and densitometry analysis of NK2 receptor protein expression in human colonic sigmoid smooth muscle. Results are expressed as a percentage of the loading control GAPDH. (A) NK2 receptor and GAPDH protein blots with specific bands corresponding to the expected molecular weight of ∼45 and ∼36 kDa, respectively. (B) NK2 receptor protein expression in female (n = 25) and male (n = 15) samples. Horizontal lines denote medians. (C) NK2 receptor protein expression in samples from patients younger (closed circles) or older (open circles) than 50 years of age. (D) Spearman correlation analysis of NK2 receptor expression and age in females (r = 0.5023, *P = 0.0105) and males (r = 0.127, P = 0.6499).
NK2 Receptor Localization Throughout the Smooth Muscle Layers.
Strong NK2 receptor immunoreactivity was detected throughout the circular and longitudinal smooth muscle layers of the human sigmoid colon (n = 6) (Fig. 6). Extensive colocalization with α-SMA (n = 2) indicates that NK2 receptors are present on smooth muscle cells (Fig. 6, A–C). On the other hand, NK2 receptors did not appear to be expressed on interstitial cells of Cajal (n = 2), as evidenced by double labeling with c-kit (Fig. 6, D–I). NK2 receptor staining was colocalized with the neuronal marker β-tubulin on some neuronal processes throughout the smooth muscle (Fig. 6, J–L). No obvious gender differences were observed in the distribution of NK2 receptors throughout the smooth muscle layers.
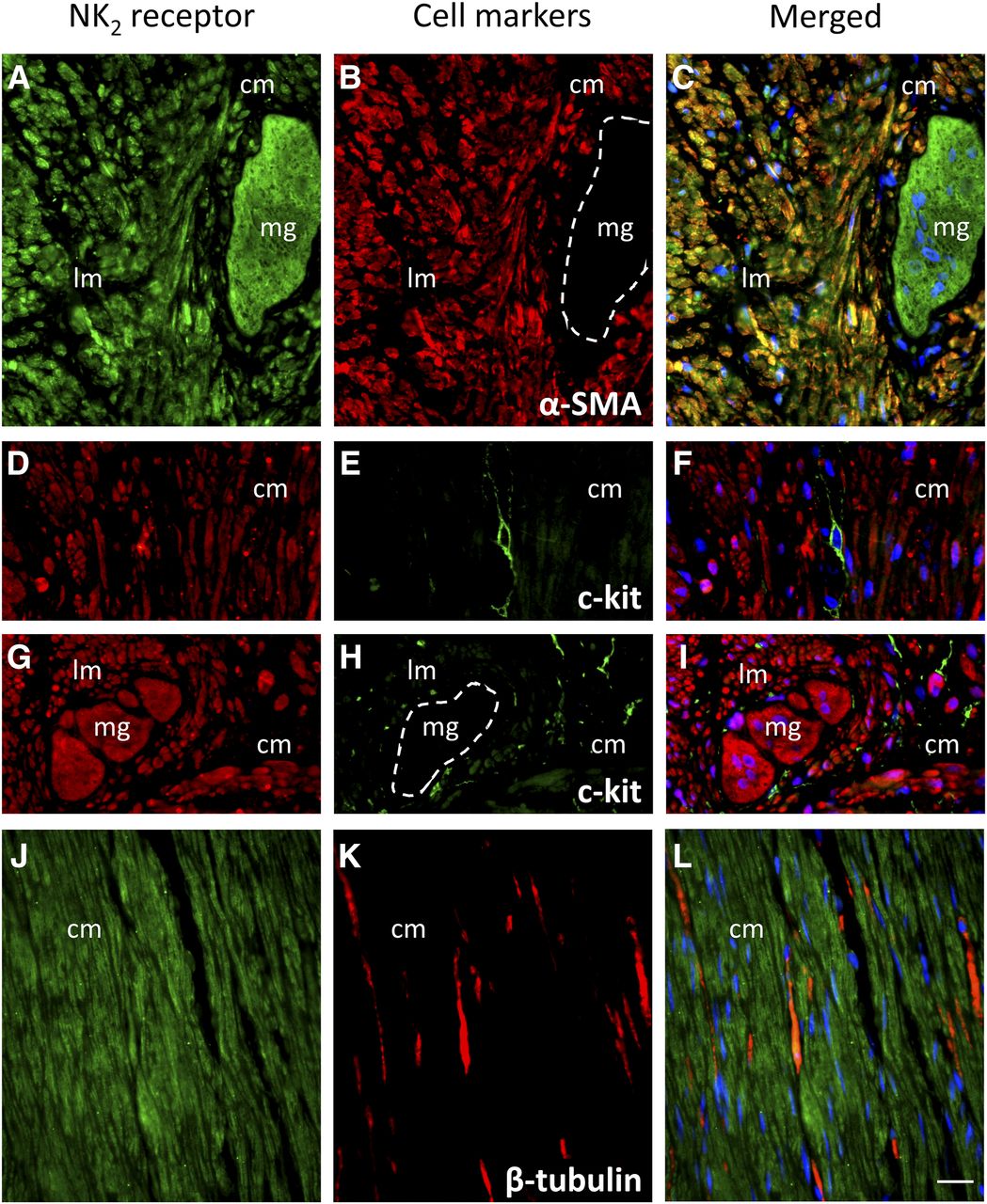
Fluorescent immunoreactivity of the NK2 receptor (left panel) in the smooth muscle layers of the human sigmoid colon. The cell markers used (middle panel) are α-SMA (B, n = 2), the interstitial cells of Cajal marker c-kit (E and H, n = 2), and the neuronal marker β-tubulin (K, n = 6). The merged images (right panel) contain the nuclear marker DAPI (blue). Staining is present on myenteric ganglia (mg), circular muscle (cm), and longitudinal (lm) muscle. Scale bar, 20 µm. Representative images were taken from female specimens aged 62 years old (A–C and J–L) and 46 years old (G–I) and a male specimen aged 49 years old (D–F).
NK2 Receptor Localization within the Myenteric Ganglia.
Dense NK2 receptor immunoreactivity was expressed within the myenteric ganglia of the human sigmoid colon (n = 6) (Fig. 7). Although weak colocalization with the neuronal marker HuC/D was observed, strong NK2 receptor staining was detected on the structures surrounding the HuC/D positive cells (n = 2) (Fig. 7, A–C). Similarly, double labeling with β-tubulin (n = 6) suggests that NK2 receptors are present on a limited number of enteric neurons (Fig. 7, D–F). On the other hand, NK2 receptor staining was extensively colocalized with the glial cell markers S100 (Fig. 7, G–I, n = 6) and GFAP (Fig. 7, J–L, n = 2) within the myenteric ganglia, as well as on glial processes throughout the muscle layers. There were no obvious gender differences in the localization of NK2 receptor expression within the myenteric ganglia.

Fluorescent staining of the NK2 receptor (left panel) within the myenteric ganglia of the human sigmoid colon. The cell markers used (middle panel) are the nerve cell body marker HuC/D (B, n = 2), the neuronal marker β-tubulin (E, n = 6), and the glial cell markers S100 (H, n = 6) and GFAP (K, n = 2). The merged images (right panel) contain the nuclear marker DAPI (blue). Staining is present on myenteric ganglia (mg), circular muscle (cm), and longitudinal (lm) muscle. Scale bar, 20 µm. Representative images were taken from a female specimen aged 62 years old (A–F) and a male specimen aged 49 years old (G–L).
NK2 Receptor–Mediated Signaling Pathways.
Contractile responses of human colonic smooth muscle strips to NKA and LMN-NKA were examined in the presence of various second messenger inhibitors to determine whether the gender differences observed in contractility could be attributed to receptor signaling (Fig. 8). Following the blockade of PLC by U73122, contractile responses produced by NKA in male strips (n = 11) were significantly attenuated (***P = 0.0005, two-way ANOVA) (Fig. 8A). NKA-induced contractions in female strips (n = 9) were also reduced, but to a lesser extent (**P = 0.0058), and a gender difference was observed at the highest concentration (300 μM) of inhibitor (*P = 0.0138, two-way ANOVA) (Fig. 8A). Similar observations were made using LMN-NKA as the agonist (Fig. 8B). Contractions elicited by LMN-NKA in both male (n = 10) and female (n = 9) strips were reduced by PLC inhibition (****P < 0.0001 and **P = 0.0078, respectively, two-way ANOVA), and overall, this inhibitory effect was more pronounced in males compared with females (***P < 0.0008, two-way ANOVA) (Fig. 8B).

Gender-related variations in NK2 receptor–mediated second messenger signaling pathways. Contractile responses of female (red) and male (blue) human colonic circular smooth muscle strips to NKA (left panel) and LMN-NKA (right panel) at 0.3 μM, either alone (dashed lines) or in the presence of the PLC inhibitor U73122 (A and B), the inositol IP3 receptor inhibitor 2-APB (C and D), the PKC inhibitor GF109203X (E and F), or the Rho kinase inhibitor Y-27632 dihydrochloride (G and H) at 3, 30, 100, and 300 μM. Data are expressed as a percentage of the contractile response before pharmacological inhibition and are presented as means ± S.E.M. (n = 5–11 for each curve). Significance by two-way ANOVA is indicated by a single symbol for P < 0.05, two for P < 0.01, three for P < 0.001, and four for P < 0.0001 (* for before vs. after inhibition within each gender; † for females vs. males).
Since activation of PLC triggers separate transduction pathways involving IP3 receptors and PKC, the effect of inhibiting these signaling proteins was also examined. Blockade of IP3 receptors by 2-APB did not affect contractions elicited by NK2 receptor activation in either gender (n = 6 females, five males) (Fig. 8, C and D). On the other hand, PKC inhibition by GF109203X produced concentration-dependent reductions of NK2 receptor–mediated contractile responses in both female (n = 6) and male (n = 6) strips (****P < 0.0001 for both genders and agonists, two-way ANOVA) (Fig. 8, E and F). During smooth muscle contraction, the Gαq/11-PLC cascade may occur alongside Gα12/13-Rho kinase signaling, so the involvement of this latter pathway was also examined. In both genders (n = 9 for each), contractile responses evoked by NKA were significantly diminished by Rho kinase blockade with Y-27632 in a concentration-dependent manner (****P < 0.0001, two-way ANOVA) (Fig. 8G). Similarly, inhibition of Rho kinase caused a concentration-dependent attenuation of LMN-NKA–induced contractions in both female (n = 9) and male (n = 10) strips (****P < 0.0001; ***P < 0.0004, respectively, two-way ANOVA) (Fig. 8H). No gender-related distinctions were observed for PKC and Rho kinase inhibition (Fig. 8, E–H).
Since gender differences were observed for PLC inhibition, real-time PCR was conducted to uncover any gender variations in the expression of genes encoding the PLCβ family of isoenzymes, as well as the Gαq family of G proteins (Fig. 9). There were no observed differences in the mRNA expression of G proteins Gαq, Gα11, and Gα14 (Fig. 9, A–C) or PLCβ 1–4 isoenzymes (Fig. 9, D–G) between male (n = 25–27) and female (n = 26 to 27) human colonic smooth muscle samples.

Gene expression of the Gαq family of G proteins (A–C) and the PLCβ isoenzymes (D–G) in female (n = 26 to 27) and male (n = 25–27) human colonic smooth muscle. Results are normalized to GAPDH expression and presented as fold change relative to the inter-run calibrator. Horizontal lines denote medians.
Discussion
The present study has uncovered several gender differences in NK2 receptor activity and expression in the human colon. NK2 receptor–mediated contractility is more pronounced in females compared with males, and this is accompanied by a greater potency of the NK2 receptor antagonist ibodutant, a higher basal release of SP, and an increased NK2 receptor protein expression with age. In addition, the involvement of PLC-mediated signaling appears to be less prominent in females compared with males. The present findings also appreciate the importance of NK2 receptor–mediated Ca2+ sensitization pathways via PKC and, for the first time, Rho kinase in the human colon.
Male colonic smooth muscle exhibited reduced responsiveness to the selective NK2 receptor agonist LMN-NKA. This finding is in contrast to our previous report, in which contractile responses induced by LMN-NKA in human colonic muscle strips were similar in both genders (Burcher et al., 2008). However, in that earlier study, the data were expressed as a percentage of the maximum response to ACh, so this may underlie the discrepancy. On the other hand, there were no gender differences in contractility produced by the natural NK2 receptor ligand NKA in the current study, and this is consistent with our previous findings (Burcher et al., 2008). The differences observed for NKA and LMN-NKA could be due to the activation of NK1 receptors by NKA, since natural tachykinins preferentially, but not exclusively, bind to all three tachykinin receptor subtypes (Nakamura et al., 2011).
The NK2 receptor antagonist ibodutant displays a significantly higher degree of antagonism in females compared with males when against NKA. This is in line with the results obtained from human clinical trials and may underlie the gender discrepancies reported for the efficacy of ibodutant in patients with diarrhea-predominant IBS (Tack et al., 2017). Conversely, the potency of ibodutant against LMN-NKA did not vary between genders, and this is consistent with previous findings using [β-Ala8]NKA(4-10) as the selective NK2 receptor agonist (Santicioli et al., 2013). Ibodutant has been shown to display a high affinity for human NK2 receptors in [125I]NKA binding studies using transfected cells (pKi = 10.1) (Cialdai et al., 2006) and colonic smooth muscle (pKi = 9.9) (Santicioli et al., 2013). Its affinity values at NK1 (pKi = 6.1) and NK3 (pKi = 6.2) receptors were shown to be four orders of magnitude lower, confirming the selectivity of ibodutant for NK2 receptors (Cialdai et al., 2006). Although the mechanisms underlying agonist-dependent differences in the gender-specific antagonistic activity of ibodutant remain unclear, they may be occurring to different extents in males and females. Nevertheless, the potency of ibodutant against NKA, which we have observed to be greater in females, would more closely reflect the physiologic setting.
Interestingly, the antagonistic activity of ibodutant at the NK2 receptor appears agonist-dependent in males with a higher potency against LMN-NKA compared with NKA. NK2 receptors can couple to both Gαq/11 and Gαs proteins, and different ligands can stabilize distinct NK2 receptor states, leading to the activation of different downstream pathways (Palanche et al., 2001; Maillet et al., 2007; Valant et al., 2009). Thus, NKA- and LMN-NKA-dependent conformations of the NK2 receptor may be responsible for the agonist-specific effect of ibodutant in the male colon.
In terms of tachykinin peptide release, basal levels of NKA were similar in both genders, whereas a higher amount of SP was released from female colonic smooth muscle. There were no gender-related variations in EFS-induced tachykinin release. In a previous study, KCl-induced NKA and SP levels were higher in colonic smooth muscle from female guinea pigs subjected to colitis compared with controls, although this difference was not seen in males (Bellucci et al., 2016). The overall amount of SP released was lower compared with NKA in guinea pig colonic smooth muscle (Bellucci et al., 2016), and this is consistent with the current findings. It appears that alternative splicing of the tachykinin precursor 1 gene (encoding SP and NKA) favors NKA production in colonic enteric neurons. The reduced level of SP at lower frequency of EFS (0.5 Hz) may be due to an autoregulatory mechanism, whereby SP exerts a feedback inhibition of its own release by activating NK1 receptors on enteric nerve terminals (Lomax et al., 1998).
NK2 receptor protein expression increases with age in females, supporting the evidence that changes in sex hormone levels influence tachykinin peptide and receptor expression (Villablanca and Hanley, 1997; Pinto et al., 2009). Sex hormones have also been shown to influence intestinal smooth muscle contractions. In human colonic circular muscle strips, carbachol-evoked contractions were reduced in the presence of estrogen, and this effect was prevented by estrogen receptor antagonism (Hogan et al., 2009). Similarly, progesterone inhibited spontaneous and EFS-induced contractions in mice colonic circular muscle strips and decreased fecal output in two diarrheal mouse models (Li et al., 2012). Since sex hormones affect tachykinin expression and colonic contractile responses, they may also be contributing to the age-related changes of NK2 receptor protein expression in females. In other words, NK2 receptor expression and activity may be regulated by sex hormones.
NK2 receptor immunoreactivity was densely localized on colonic circular and longitudinal smooth muscle but was absent on interstitial cells of Cajal, reinforcing previous immunofluorescence findings in the human colon (Renzi et al., 2000; Warner et al., 2000; Jaafari et al., 2007; Nakamura et al., 2011). This suggests that NK2 receptors mediate colonic contractions by direct activation of smooth muscle cells, without regulating intestinal pacemaker cell activity.
Strong NK2 receptor staining was also detected throughout the myenteric ganglia, suggesting that apart from mediating direct myogenic effects, NK2 receptors also play a neuroregulatory role in colonic motor pathways. Although weak NK2 receptor colocalization with neuronal markers was observed, intense NK2 receptor labeling was found on myenteric glial cells. Glial cells were traditionally considered to provide passive support to neurons, but an abundance of evidence now exists linking impaired glial function to altered intestinal motility (Aube et al., 2006; Nasser et al., 2006; McClain et al., 2014). Interestingly, tachykinin receptor agonists can induce glial cell activation in the mouse colon (Broadhead et al., 2012). Furthermore, NKA has been shown to activate neuronal and glial networks during neuroinflammation and neurodegeneration (Delvalle et al., 2018). However, this effect is via NK2 receptors on surrounding sensory neurons and nerve varicosities, since NK2 receptors were undetectable on mouse enteric glial cells (Delvalle et al., 2018). This earlier study used the same NK2 receptor antibody as in the current study, indicating a species difference in the expression of NK2 receptors on enteric glial cells.
U73122 showed a more prominent effect in males compared with females. This does not appear to be associated with the gene expression of PLC, since the mRNA expression levels of all PLCβ isoenzymes were similar in both genders. The mechanism underlying the gender distinction is unclear but may involve differences in the protein levels of phosphorylated PLCβ isoenzymes, which have not been determined in the current study. In addition to PLC inhibition, U73122 has been shown to inhibit the Ca2+ pump on the sarcoplasmic reticulum to deplete Ca2+ (MacMillan and McCarron 2010).
Interestingly, NK2 receptor–mediated contractile responses in the human colon do not appear to be related to IP3 receptor activation. In accordance, inositol phospholipid hydrolysis stimulated by LMN-NKA has been previously shown to be much weaker in human colonic smooth muscle compared with rat bladder positive controls (Warner et al., 2000). In addition, NKA-evoked contractions of human colonic smooth muscle were unaffected by intracellular Ca2+ depletion (O’Riordan et al., 2001). Thus, NK2 receptor–induced intracellular Ca2+ release by IP3 receptor activation is a minor pathway in mediating human colon contractility.
NKA and LMN-NKA evoked contractile responses that were significantly diminished by PKC and Rho kinase inhibitors, suggesting that Ca2+ sensitization contributes to NK2 receptor–mediated contractility of the human colon. Ca2+ sensitization is the phenomenon by which smooth muscle contractions can be sustained independently of Ca2+, by PKC- and Rho kinase–mediated inhibition of myosin light-chain phosphatase, which prevents the dephosphorylation of myosin light chain (Puetz et al., 2009; Perrino, 2016). Previously, the contribution of PKC and Rho kinase to Ca2+ sensitization evoked by muscarinic receptor activation has been demonstrated in membrane-permeabilized rat colonic muscle strips (Takeuchi et al., 2004, 2007). This is the first report demonstrating the role of PKC and Rho kinase signaling in NK2 receptor–activated smooth muscle contractions of the human colon.
In conclusion, the present study has revealed an increased NK2 receptor activity in females, which may underlie the gender differences in the treatment of diarrhea-predominant IBS. These gender-related differences may be due to the influence of ovarian hormones, since the expression of NK2 receptors increases with age in females. These findings highlight that gender differences should be considered in the therapeutic development of NK2 receptor agents. The pronounced localization of NK2 receptors on enteric glial cells in the human colon suggests that aside from direct myogenic activation, NK2 receptors may also have an important neuroregulatory role. Furthermore, the current study indicates the importance of both Gαq/11-PKC and Gα12/13–Rho kinase signaling pathways in NK2 receptor–mediated contractile responses in the human colon.
Acknowledgments
We would like to thank Dr. Maria Altamura and Dr. Stefania Meini of Menarini Ricerche S.p.A., Firenze, Italy, for the supply of ibodutant and the discussion of ibodutant results. We also thank Elizabeth Burcher for providing intellectual input and Haylee Guy for assistance with specimen collections.
Authorship Contributions
Participated in research design: Drimousis, Liu.
Conducted experiments: Drimousis, Markus.
Contributed to new reagents or analytic tools: Perera, Phan-Thien.
Performed data analysis: Drimousis, Liu.
Wrote or contributed to the writing of the manuscript: Drimousis, Murphy, Zhang, Liu.
Footnotes
- Received February 26, 2020.
- Accepted July 28, 2020.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- ACh
- acetylcholine
- 2-APB
- 2-aminoethoxydiphenyl borate
- DAPI
- 4′, 6′-diamidino-2-phenylindole
- EFS
- electrical field stimulation
- Emax
- maximal contractile responses
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- IBS
- irritable bowel syndrome
- IP3
- inositol 1,4,5-triphosphate
- LMN-NKA
- [Lys5,MeLeu9,Nle10]NKA(4-10)
- NKA
- neurokinin A
- NK receptors
- neurokinin receptors for tachykinin peptides
- PCR
- polymerase chain reaction
- pKB
- dissociation constant
- PKC
- protein kinase C
- PLC
- phospholipase C
- α-SMA
- α-smooth muscle actin
- SP
- substance P
- sscDNA
- single-strand cDNA
- Copyright © 2020 by The Author(s)
This is an open access article distributed under the CC BY-NC Attribution 4.0 International license.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}