


Abstract
Women are at significantly greater risk of developing stress-related disorders such as depression. The increased risk begins during puberty and continues throughout life until menopause, suggesting a role for ovarian hormones in this increased susceptibility. Importantly, inflammation has been gaining momentum in its role in the pathogenesis of depression. Herein, clinical and preclinical studies have been reviewed to better understand how sex differences within the immune system may contribute to exaggerated risk of depression in females. First, studies that investigate the ability of psychologic stress episodes to engage the inflammatory systems both in the brain and periphery are reviewed with a special focus on sex-specific effects. Moreover, studies are discussed that identify whether imbalanced inflammatory milieu contributes to the development of depression in males versus females and whether these effects are regulated by estradiol. Importantly, we propose a locus coeruleus–norepinephrine–cytokine circuit as a conduit through which stress could increase stress susceptibly in females. Finally, the anti-inflammatory capacity of traditional and nontraditional antidepressants is investigated, with the goal of providing a better understanding of pharmacotherapeutics to enhance strategies to personalize antidepressant treatments between the sexes. The studies reviewed herein strongly support the need for further studies to elucidate whether females are especially sensitive to anti-inflammatory compounds as adjuvants to traditional therapies.
SIGNIFICANCE STATEMENT Women have hve an increased risk of developing stress-related disorders such as depression. In this review, literature from clinical and preclinical studies are integrated to define sex differences in stress-induced inflammatory responses as a potential source for the etiology of sex differences in depressive disorders. Moreover, the anti-inflammatory capacity of traditional and nontraditional antidepressants is reviewed to inform on potential pharmacotherapeutic strategies to personalize antidepressant therapy in a sex-dependent manner.
Introduction
Depression is one of the most debilitating diseases in the United States (Almeida, 2005) and, globally, is considered a significant source of disability (Reddy, 2010). Despite improvements in overall health and life expectancy in the United States, the incidence of depressive disorders has steadily increased since the 1990s (Murray et al., 2013), and recently, it has been reported to occur in 7.3% of adults and 12.7% of adolescents in the United States (Weinberger et al., 2018) and other countries, including Canada (Patten et al., 2006). Although reports from the World Health Organization identify that depression can be precipitated by medications, illnesses, substance use disorders, or genetics, research in humans and animals identify that exposure to stress remains a significant risk factor for the development of major depression (Krishnan and Nestler, 2008). However, it should be noted that rather than definitive mechanisms that promote depression in humans, our knowledge is largely limited to a list of factors that increase the risk of developing depression. Beyond stressful life experiences increasing risk of depression, there is also a robust sex difference whereby women are nearly twice as likely as men to suffer from depression (Kessler et al., 1993; Weissman et al., 1993; Marcus et al., 2005). In fact, this differential risk of depression is recognized globally in developed countries, suggesting that the female sex bias may be routed more from biologic sex differences rather than race, diet, education, or other factors (Rai et al., 2013). Despite the recognition that the incidence of depression is biased toward females, the etiology of this sex difference is unclear, which impedes progress for the development of precision medicine.
Despite evidence of exaggerated inflammation associated with major depression and several proof-of-concept studies demonstrating the efficacy of anti-inflammatory therapy as antidepressants, there remain several key questions that will be discussed in this Minireview using evidence from human and rodent studies. First, studies that investigate the ability of psychologic stress episodes to engage the inflammatory systems in both the brain and periphery will be reviewed. Moreover, studies identifying whether imbalanced inflammatory milieu contributes to the development of depression will be discussed. Throughout this Minireview, we will provide an introduction to available literature defining the sex differences in stress-induced inflammatory responses as a potential source for the etiology of sex differences in stress-related disorders. Finally, recent advances in strategies to treat depression will be reviewed, with the goal of providing a better understanding of pharmacotherapeutics resulting in modifications to immune levels in depression to enhance strategies for precision medicine.
Recruitment of the Innate Immune Response after Psychological Stress
Inflammatory Response to Experimental Models of Stress in Men and Women.
Meta-analysis of studies conducting controlled, acute psychologic stressors in humans identified consistent and robust increases in circulating inflammatory factors (Steptoe et al., 2007). The Trier social stress test (TSST) has been used for more than 2 decades as a way to reliably evaluate the response to social stressors in humans in a controlled laboratory setting (Kirschbaum et al., 1993). This model induces stress by placing participants in a situation in which they are required to make an interview-style presentation, followed immediately by an unexpected mental arithmetic test. This test is conducted in front of a panel of judges lacking any feedback or encouragement, eliciting a highly stressful experience (Kirschbaum et al., 1993; Allen et al., 2016). The TSST has been shown to elicit elevated levels of both IL-6 and TNF-α in healthy controls (Gaab et al., 2005). Moreover, patients suffering from depression exhibit significantly increased TSST-induced IL-6 levels above that of nondepressed controls (Pace et al., 2006). Another study using the TSST compared the immune responses between men and women, showing that women in the luteal phase of their menstrual cycle demonstrated increased lipopolysaccharide (LPS)-induced cytokine production in blood samples collected after stress, whereas men exhibited a reduction (Rohleder et al., 2001). A similar study focused on female participants who used oral contraceptives, comparing them with participants who were in the luteal phase of their menstrual cycle (Rohleder et al., 2003). The findings from this study showed that women who used contraceptives had higher levels of IL-6 after exposure to TSST. Contraceptive users also had a blunted glucocorticoid sensitivity when compared with women in the luteal phase of their menstrual cycle (Rohleder et al., 2003). It has also been demonstrated that women and men exhibit differing time courses of stress-induced cytokine release. For example, in a study using a mental arithmetic stress task, the paced auditory serial addition task (PASAT), male participants exhibited stress-induced IL-6 release that peaked 30 minutes after the PASAT but returned toward baseline by 60 minutes. IL-6 release in female participants, on the other hand,increased only only 60 minutes after the PASAT (Edwards et al., 2006). Taken together, the distinct inflammatory response to social stress in females may underlie the increased sensitivity women demonstrate to social stressors and may promote the increased risk of stress-related psychiatric disorders (Slavich and Irwin, 2014; Slavich and Sacher, 2019).
Inflammatory Response to Preclinical Models of Stress in Male and Female Rodents.
Preclinical studies have also shed considerable light on immune system activation in response to social and psychologic stress in experimental models; therefore, this concept will only be briefly reviewed below (for reviews, see Bekhbat and Neigh (2018), Finnell and Wood (2018), Rainville and Hodes (2019)). For example, in response to foot shock (Blandino et al., 2013; Jones et al., 2018), an established psychological stressor whereby the animal receives brief, mild, and inescapable electric shocks, IL-1β gene expression in the paraventricular nucleus is robustly increased (Blandino et al., 2013). Moreover, the effect of foot shock on inflammatory cytokines also occurs in extrahypothalamic regions; immunohistochemical analysis demonstrates that foot shock can ultimately increase the levels of IL-1β protein within the hippocampus (Jones et al., 2018). Moreover, a recent study also evaluated sex differences in the inflammatory response to an LPS injection 24 hours after inescapable shock. Peripheral cytokine release was potentiated in females with a history of stress compared with stressed males or unstressed males or females (Fonken et al., 2018). In the hippocampus, both males and females exhibited enhanced stress-induced neuroinflammation, yet the priming was proposed to occur through different mechanisms (Fonken et al., 2018). Importantly, this inflammation-enhancing effect of stress is not limited to foot shock. Chronic mild stress (CMS) or chronic variable stress, an experimental stressor whereby the subject is placed under mild and ever-changing stress-provoking conditions (Hodes et al., 2015; Wang et al., 2018; Johnson et al., 2020), has been shown to cause an increase in gene expression for inflammatory factors such as IL-6, IL-1β, and TNF-α in the hippocampus (Wang et al., 2018; Liu et al., 2019). In studies directly identifying sex-specific effects of CMS on hippocampal inflammation, both males and females demonstrated increased proinflammatory cytokines, but males exhibited increases in mRNA induced nitrous oxide synthase and IL-1β, whereas females demonstrated increases in TNF-α and reductions in anti-inflammatory cytokines IL-10 and IL-1 receptor antagonist (Liu et al., 2019). Moreover, ratios of pro- to anti-inflammatory cytokines indicated that females were more prone to a proinflammatory environment. Moreover, chronic stress during adolescence resulted in increased expression of IL-1β mRNA within the hippocampus of adult females, but not males (Bekhbat et al., 2019). Furthermore, in males, the use of minocycline was shown to dampen the activation of stress-induced proinflammatory responses in the brain, indicating that these cytokines are sourced from microglia, the resident immune cells of the brain (Wang et al., 2018). CMS has also been shown to induce microglial activation and cytokine release within the prefrontal cortex, but this effect was most pronounced in males, despite both males and females demonstrating depressive-like and anxiety-like behaviors (Wohleb et al., 2018). Importantly, a separate study also identified that females are more sensitive to a subchronic regimen of chronic variable stress compared with males as measured by behavioral sensitivity (Hodes et al., 2015).
Similar to the TSST in humans, experimental models of social stressors are also commonly used to generate an ethologically relevant stress exposure for rodents. One of the most commonly used of the social paradigms is the resident-intruder social defeat test, originally developed by Miczek (1979). This paradigm induces stress by introducing an experimental male into the home cage of an older, more aggressive, territorial male. The social defeat paradigm, like the TSST, is well recognized to stimulate the immune response in rodents (Wohleb et al., 2012; Wood et al., 2015; Tang et al., 2018). Social stress–induced activation of immune cells in the brain causes a prolonged inflammatory response and release of IL-1β (Wohleb et al., 2012). Moreover, long-lasting increases in neuroinflammation have been identified in the locus coeruleus (LC), a major noradrenergic nucleus, of a susceptible subset of male rats, whereas this response was lacking in the behaviorally resilient subset (Wood et al., 2015; Finnell et al., 2017a). Conducting social defeat stress in female rats has been a challenge because male rats are not aggressive toward females, and female rats are not aggressive toward other females outside of the postpartum period [see review Finnell and Wood (2016)]. One study used a designer receptor exclusively activated by designer drug-based activation of the ventromedial hypothalamus to induce male aggressive behavior toward females and determined that this modified version of social defeat produced behavioral and peripheral/central immune dysfunction in females (Yin et al., 2019). Alternatively, another strategy to subject females and males to social stress is by exposing rats or mice to a witness or vicarious stress, whereby a male or female is forced to witness social defeat occurring between two males (Hodes et al., 2014; Patki et al., 2014; Finnell et al., 2017b, 2018; (Iñiguez et al., 2018). These studies have highlighted that witness stress engages a heightened immune response within the periphery and the central nervous system, and that this immune response is involved in sensitivity to this stressor. Future studies directly comparing the immune response to witness stress between males and females will be critical in understanding sex differences in social stress–induced inflammation.
Sex-Specific Effects of Stress-Induced Activation of Microglia: A Link to Noradrenergic Hyperarousal.
Stress-induced promotion of a proinflammatory milieu is suggested to occur by several specific mechanisms, including microglial activation and blood-brain barrier degradation [(Menard et al., 2017; Dudek et al., 2020) and as extensively reviewed in Finnell and Wood (2018)]. The mechanism we will focus on for the purpose of this Minireview is microglial activation because of the experimental evidence supporting a role for sex differences in stress-induced microglial activation. Microglial cells are capable of changing morphology between a proinflammatory (M1) and anti-inflammatory (M2) state (Hanisch and Kettenmann, 2007; Crain et al., 2013; Nelson et al., 2017). Microglia, when activated by stress, for example, shift morphology states from a ramified surveillant state to an ameboid, activated state, leading to the secretion of proinflammatory cytokines such as Il-1β, IL-6, and TNF to neighboring cells (Hanisch and Kettenmann, 2007; Nelson et al., 2017). Importantly, female rats exhibit more activated microglia as demonstrated by morphology beginning near the time of early puberty and continuing throughout adulthood (Schwarz et al., 2012). As we discuss later in this review, this timeframe coincides with the period of greatest risk of depression in females. In addition, the prefrontal cortex of unstressed female rats has demonstrated heightened basal activation of microglia evidenced by a primed phenotype (Bollinger et al., 2016). Interestingly, a primed or sensitized microglial phenotype is most commonly noted to occur after stress exposure (Frank et al., 2012, 2018; Fonken et al., 2018). Importantly, Bollinger et al. (2017) went on to determine whether acute or chronic restraint stress affected microglial morphology in a sex-dependent manner. In the orbitofrontal cortex, basolateral amygdala, and dorsal hippocampus, chronic stress reduced microglial activation (as evidenced by morphology changes) yet upregulated a greater number of immune factors in females compared with males. It will be important to determine whether similar effects on microglial morphology are also achieved with social stress; however, these findings provide evidence of sex differences in microglial morphology that may bias females toward greater neuroinflammatory responses in the context of stress.
Microglial-induced increases in proinflammatory cytokines no doubt have various effects throughout the brain depending on the specific brain region and cell type. One brain nucleus through which proinflammatory cytokines are likely acting to promote stress sensitivity is the LC. IL-1β microinjection into the LC increased burst firing of LC neurons (Borsody and Weiss, 2002–2003), an LC response that occurs after a sensory stimulus (Aston-Jones and Bloom, 1981a,b). Moreover, both peripheral and intra-LC injection of LPS increased LC-NE firing, dependent upon IL-1 receptors within the LC (Borsody and Weiss, 2002–2003, 2004). Importantly, LC-norepinephrine (NE) dysfunction is linked to stress-related disorders such as depression and PTSD, and as discussed above, females already demonstrate distinct differences within the LC-NE system capable of promoting stress-related dysfunction. Combined with the knowledge that females demonstrate increased inflammatory responses to social stressors, as discussed above, this begs the question as to whether exaggerated inflammatory input to the LC could be serving as the conduit through which stress increases risk of stress-related disorders in females.
A recent study provided key information as to the mechanisms driving the elevated inflammation in the context of stress. NE regulates cytokine release from macrophages and microglia via binding to adrenergic receptors on these cell types. Finnell et al.(2019) challenged the contribution of the LC-NE system in male rats on stress-induced circulating and neuroinflammatory priming by using the compound N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine, an LC-selective noradrenergic neurotoxin. These studies demonstrated that suppressed NE release in N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine–treated rats had a significant impact on stress-induced inflammation in the plasma and brain (Finnell et al., 2019). Importantly, these findings clearly indicated that the LC-NE system was necessary for the stress-induced inflammatory priming known to occur after social defeat stress (Frank et al., 2015). The recognition that the LC-NE system is capable of regulating stress-related inflammation opens a path for future studies to understand possible treatment pathways. Although this study did not assess sex differences, it is noteworthy that there is robust sexual dimorphism within the LC-NE system. As is eloquently reviewed in Bangasser et al. (2016), sex differences in the LC-NE system may serve as a core pathophysiological mechanism that predisposes women to disorders that are characterized by hyperarousal, such as depression and PTSD. In brief, the LC of adult female rats is larger than that of adult males because of an increase in the number of NE-containing neurons in females (Luque et al., 1992) and because the dendrites of the LC are denser than those of males (Bangasser et al., 2011), potentially increasing synaptic connectivity with other stress-related brain regions. Moreover, estrogen increases NE levels in various stress-sensitive brain regions through ERα or ERβ (Lubbers et al., 2010). Both ER subtypes are prevalent in the LC (Shughrue et al., 1997) and are therefore well positioned to regulate NE release from the LC to stress-sensitive target regions. Within NE-containing nuclei, estradiol is capable of increasing NE synthesis by enhancing the expression of tyrosine hydroxylase, a rate-limiting enzyme critical to the synthesis of NE (Serova et al., 2002). Moreover, estradiol is also capable of impeding enzymatic degradation of NE via inhibiting catechol-O-methyltransferase (Jiang et al., 2003).
Link between Inflammation and Depression
Inflammation’s Link to Depression in Humans.
In humans with major depressive disorder (MDD), high levels of proinflammatory cytokines have been identified in subsets of patients. For example, proinflammatory cytokines have been found in high levels in both the cerebrospinal fluid and the serum of depressed patients (Zorrilla et al., 2001; Miller et al., 2002; Alesci et al., 2005; Motivala et al., 2005; Maes et al., 2009; Raison et al., 2013). Overall, meta-analyses lend support to the presence of increased inflammation in MDD; however, there are individual studies that report no such association (Fried et al., 2019). Furthermore, one meta-analysis suggests that this variability is due to different types of depression; Lamers et al. (2013) found the association between depression and inflammation only evident for atypical depression rather than melancholic depression. Nonetheless, alterations in cytokines in the central and peripheral nervous systems were highly correlated with the severity of the symptoms of depression (Zorrilla et al., 2001). Moreover, in patients with MDD, markers of activated microglia, such as translocator protein, have been shown to be elevated in serum and brain and represent a candidate marker of treatment outcome (Setiawan et al., 2015; Holmes et al., 2018; Richards et al., 2018). Beyond the correlational studies linking markers of inflammation and depression, inflammatory factors have also been shown to have a causal effect on MDD symptoms, such as cognitive dysfunction, sleep alterations, and regular fatigue, which are all associated with depression (Bower et al., 2002; Meyers et al., 2005; Motivala et al., 2005). Moreover, cytokine treatment of diseases that are unrelated to depression have a high probability of leading patients to develop depression and anxiety (Miller et al., 2009). Use of cytokines such as IL-2, interferon (IFN)-α, IFN-γ, and granulocyte macrophage colony stimulating factor for cancer treatment have been shown to cause depression in patients with no prior history (Denicoff et al., 1987; Renault et al., 1987; Capuron et al., 2001; Schmeler et al., 2009). Indeed, the use of antidepressants such as paroxetine counteracts the effects of depression induced by IFN-α treatment, further supporting the concept that cytokines can independently produce depressive symptoms (Musselman et al., 2001a). Furthermore, studies have shown that females may demonstrate greater risk for MDD after inflammatory challenges, such as IFN-α, compared with men (Fontana et al., 2002; Koskinas et al., 2002; Gohier et al., 2003). However, not all studies support this sex-biased interpretation, with some research reporting no sex differences in depressive symptoms after IFN-α administration (Bonaccorso et al., 2002; Kraus et al., 2003; Raison et al., 2005). Despite these inconsistent findings, a larger meta-analysis that included 26 studies concluded that females are significantly more likely to develop MDD after IFN-α administration (Udina et al., 2012).
There is also support for sex differences in markers associated with MDD. Various sex-dependent markers for MDD were identified from the serum of both male and female patients (Ramsey et al., 2016). Only three female-specific markers were identified for MDD. Insulin-like growth factor 5, involved in biologic processes including cell growth and glucose metabolism, and macrophage inflammatory protein-3β, a chemokine that regulates lymphocyte recirculation, were both reduced in MDD females. Interestingly, the third female-specific marker was a reduction in IL-2 receptor α. Although IL-2 receptor α has no functional response on its own, it is one of three critical subunits necessary for IL-2 receptor function. As such, these data may provide insight into adaptations that are occurring in females with MDD versus males. Furthermore, a systematic study performed by Labaka et al. (2018) showed that women with MDD had higher levels of IL-6, leptin, α-[11C]Methyl-L tryptophan, and S100B in their serum when compared with men with comparable depression. These specific markers are indicators of the presence of a high inflammatory response, further associating acute inflammation with MDD in a sexually dimorphic manner. Their findings also demonstrate a link between the severity of depressive symptoms with the levels of IL-1β, TNF-α, C-reactive protein, and brain-derived neurotropic factor present in individuals, which are common proinflammatory markers (Labaka et al., 2018).
Inflammation’s Link to Depressive-Like Behaviors in Experimental Models in Humans and Rodents.
Findings from experimental stressors in humans also support the link between inflammation and depression. As discussed in Recruitment of the Innate Immune Response after Psychologic Stress, the experimental social stressor in humans, the TSST, activates cytokine release in healthy controls. However, patients suffering from depression exhibit significantly higher TSST-induced IL-6 levels than healthy controls (Pace and Miller, 2009). It is also noteworthy that the inflammatory response to the TSST is positively related to depressive symptom severity, even well below clinical thresholds for depression. For example, Fagundes et al. (2013) measured the IL-6 response to a TSST paradigm in subjects with depressive symptoms within the 75th and 25th percentile on the Center for Epidemiologic Studies Depression Scale, representing a score of 10 and 2, respectively, in which 16 is the clinical threshold for depression. This study identified that the subject population that fell within the 75th percentile for depressive symptoms exhibited greater TSST-induced IL-6 release as measured in plasma compared with those in the 25th percentile. Taken together, these studies support a strong association between social stress–induced cytokine release and severity of depressive symptomatology.
Experimental stressors in rodents also demonstrate similar trends whereby the impact of stress on later indices of depression-like behavior are strongly dependent on the levels of cytokines in the central and peripheral nervous systems (Hodes et al., 2014; Finnell et al., 2017b, 2019; Menard et al., 2017). Similar to humans, mice and rats exposed to repeated social stress exhibit individual differences in susceptibility to developing a depressive-like phenotype (Krishnan et al., 2007; Hodes et al., 2014; Wood et al., 2015; Finnell et al., 2017a). Moreover, the stress-susceptible phenotype is characterized by enhanced circulating (Hodes et al., 2014) and brain inflammation (Wood et al., 2015; Finnell et al., 2017a), a profile that is not observed in resilient rodents [for reviews on this topic, see (Finnell and Wood, 2018; Tsyglakova et al., 2019)]. These studies have been insightful in revealing the central and peripheral mechanisms contributing to susceptibility. Studies in mice identified that the susceptible phenotype was characterized by high levels of monocyte-derived IL-6 in serum and that neutralizing IL-6 in various ways rescued the phenotype (Hodes et al., 2014). In rats, studies have confirmed the neuroinflammatory contribution to a susceptible phenotype, identifying that IL-1β is elevated long-term in the stress-sensitive noradrenergic LC (Wood et al., 2015; Finnell et al., 2019). Importantly, blocking activation of the IL-1 receptor in brain (Wood et al., 2015) or utilizing high doses of resveratrol that reduced LC cytokine expression (Finnell et al., 2017a) was sufficient to rescue the anhedonic phenotype in susceptible rats. Taken together, these studies have provided support for a causal role of inflammation in a stress-susceptible, depressive-like phenotype in preclinical studies.
Impact of Ovarian Hormones on Behavioral and Immune Responses to Stress
Exaggerated Inflammatory Responses to Endotoxin and Stress Challenges in Women.
Several clinical reports indicate that women are more likely than men to be diagnosed with depression (Weissman and Klerman, 1992; Gallo et al., 1993; Kessler et al., 1993; Hankin et al., 1998), and establishing the basis for these differences provides a framework for understanding how these disparities emerge and potential avenues for therapeutic intervention. In fact, there is a 2-fold increased risk for depression in women, but only during certain age ranges; the elevated risk emerges at the onset of puberty, persists into adulthood (Kessler et al., 1993; Hankin et al., 1998; Nolen-Hoeksema, 2001), and is no longer evident after menopause (Kessler et al., 1993; Hankin et al., 1998). These findings strongly suggest that ovarian hormones may contribute to elevated susceptibility to stress in females. Although there is a lack of controlled studies determining whether inflammation plays a role in this elevated risk for depression, there are several lines of clinical evidence that suggest women are more sensitive to inflammatory stimuli. Two key studies have provided considerable insight into sex differences in the inflammatory and depressogenic effects of endotoxin administration (LPS). In a study published by Engler et al. (2016), a low dose of LPS (0.4 ng/kg) elicited depressive symptoms in both males and females, yet the cytokine response was distinct; women exhibited robust increases in proinflammatory IL-6 and TNF-α compared with men, whereas men responded with approximately 3 times higher serum levels of the anti-inflammatory cytokine IL-10. In a similar double-blind randomized placebo-controlled study, the acute response to low-dose endotoxin challenge of 0.8 ng/kg LPS, twice the dose of that administered in Engler et al., was also determined. Women exhibited a dramatic induction of depressive behaviors, whereas men did not, despite exhibiting similar levels of endotoxin-induced IL-6 and TNF-α (Moieni et al., 2015). It is noteworthy that Moieni et al. did not measure anti-inflammatory cytokines; thus, it begs the question as to whether IL-10, for example, was elevated selectively in men, thereby protecting against the depressogenic effects of this dose of LPS. Taken together, these findings could support the notion that the cytokine response to LPS in women is more sensitive to lower doses and, as doses escalate, that males and females demonstrate differential cytokine profiles that bias females toward proinflammatory and depressive responses, whereas males may be biased toward anti-inflammatory protective pathways. These studies highlight the need to better understand sexual dimorphism in inflammatory responses, especially considering that low endotoxin administration may elicit a similar immune response to that of social stress exposure.
As might be expected, greater exposure to life stressors of an interpersonal nature is associated with a significantly increased risk for depression in many populations, including adolescent girls. Moreover, women exhibit greater immune responses after stressful experiences and also display greater depressive symptoms when compared with controls (Miller and Cole, 2012; Giletta et al., 2018; Sullivan et al., 2018). Strikingly, one study went on to identify that the association between social stress exposure and increased risk of depression was only true for girls that exhibited a strong cytokine reaction to a laboratory-based social stressor (Slavich et al., 2020). This important study indicates that inflammation may be a critical factor linking stress exposure with increased risk of depression in females. Finally, beyond stress exposure, diseases of an inflammatory nature, such as cardiovascular disease, can also lead to a higher predisposition of the development of depression (Bucciarelli et al., 2019). In all, there is considerable support for the notion that women have a greater overall inflammatory response to endotoxin and social stressors, which may play a role in the significantly increased risk of depression evident in the female population.
Role of Ovarian Hormones in Stress-Related Inflammatory and Behavioral Responses.
Estradiol is poised to regulate microglial function, with estradiol receptors present on macrophages and microglia (Mor et al., 1999). In the absence of stress, estradiol has been shown to have dual effects on inflammation under certain conditions and is largely dose-dependent (Straub, 2007). For example, low levels of estradiol, but not progesterone or high levels of estradiol, increased IFN-γ in mitogen-activated human mononuclear leukocytes in whole blood (Matalka, 2003). Preclinical studies have sought to understand the role of this sex hormone on microglial inflammatory responses, yet findings are equivocal. There is a strong body of evidence that estradiol exerts anti-inflammatory effects on microglia after cerebral ischemia (Rahimian et al., 2019). Yet even in studies that report evidence of general microglial inhibition by estradiol, some report elevated proinflammatory cytokines concurrently in the same samples (Baker et al., 2004). Moreover, estradiol may serve a distinct role for microglia beyond ischemic events that are highly sensitive to the route of administration. For example, one study points to the critical importance of route of exposure of estradiol in differentiating a pro- versus anti-inflammatory effect on microglia from a healthy brain (Loram et al., 2012); when female rats were ovariectomized and administered estradiol in vivo followed by microglial isolation, microglia exhibited a dose-dependent increase in IL-1β production. Alternatively, when microglial cultures were isolated from ovariectomized females and exposed to estradiol ex vivo, estradiol produced an anti-inflammatory effect (Loram et al., 2012). In addition, ovarian hormones also exacerbate social stress–induced inflammatory responses, as evidenced by studies that have exposed females to repeatedly (five daily) witnessing brief social defeat episodes between two males in the resident-intruder paradigm (witness stress). For example, re-exposure to the resident’s cage in the absence of the aggressive resident rendered intact females to respond with a robust increase in proinflammatory cytokines above that of intact females with a history of nonstress control. In comparison, ovariectomized females with a history of witness stress responded with less cytokine release than that of ovariectomized rats with a history of control (Finnell et al., 2018). It should also be noted that the same paradigm was tested in males in a separate study from the same laboratory, and males with a history of witness stress did not exhibit increased inflammatory responses to being re-exposed to the resident’s cage (Finnell et al., 2017b).
Beyond the estradiol regulation of microglia, the HPA axis is another potential source of variation in immune stress responses between sexes (Miller and Raison, 2016; Bekhbat and Neigh, 2018). Cortisol is a well recognized, potent anti-inflammatory hormone. Notably, women and female rodents generally exhibit enhanced stress-induced activation of the HPA axis [for a comprehensive review on this topic, see Goel et al. (2014)]. However, women in the follicular phase of their menstrual cycle (and thus comparable progesterone levels to men) exhibit suppressed HPA axis reactivity in response to the TSST when compared with men, as demonstrated by decreased Adrenocorticotropic hormone and cortisol levels (Stephens et al., 2016). Further, this study confirmed that sex hormones were correlated with cortisol levels, thereby linking sex hormones with stress-induced cortisol responses in men and women. In support of these clinical findings, sex differences in corticosterone regulation of repeated stress-induced inflammation in rodents is also well recognized (Barnard et al., 2019). Taken together, these clinical and preclinical studies provide evidence whereby, through modulating release of the anti-inflammatory hormone cortisol in the context of social stress or through direct regulation of microglia via estrogen receptors, sex hormones may be promoting exaggerated inflammatory responses in females and anti-inflammatory responses in males.
Just as the role of ovarian hormones on inflammation is highly equivocal, so too are the effects of sex hormones on depressive-like behaviors in rodent models (Table 1). In females, in addition to the overall increased risk of depression between puberty and menopause, times of hormonal fluctuation such as postpartum and perimenopausal periods represent times of further enhanced risk of depression (Soares and Zitek, 2008). Moreover, when postmenopausal women are treated with estradiol daily for 3 months, ratings of negative mood and anxiety are elevated following the TSST as compared with placebo treated volunteers (Newhouse et al., 2010). Thus, in vivo studies have investigated the role of ovariectomizing rats, thus removing estradiol and progesterone, in a middle-aged model and found that long-term ovariectomy (4 months) enhanced a depressive-like phenotype after chronic unpredictable stress (Mahmoud et al., 2016). Although these findings deviate from human literature suggesting that fluctuations, rather than long-term removal of ovarian hormones, are what increase susceptibility, these studies concluded that ovarian hormones served a protective role in rats exposed to chronic stress. Other studies in unstressed adult female rats found similar effects when the process of ovariectomizing increased immobility in the FST compared with intact and sham rats (Li et al., 2014). However, body weight (and likely greater fat distribution) was increased by ovariectomy, and general locomotor activity was reduced in the ovariectomized group (Li et al., 2014); thus, these potential confounds of the behavioral findings cannot be ruled out. Moreover, although this and other studies (Rachman et al., 1998; Li et al., 2014) also determined that estradiol replacement could rescue the effects of ovariectomy, estradiol administration occurred by seven daily injections rather than cyclic administration (once every 4 days), which mimics the normal cycle. Alternative to studies reporting a behaviorally protective effect of estradiol, there is also considerable evidence from studies supporting that estradiol exacerbates stress sensitivity. For example, when gonadally intact and ovariectomized female mice were exposed to chronic unpredictable stress, ovariectomy conferred protection against stress-induced immobility in the FST (LaPlant et al., 2009). In addition, Shansky et al. (2004) identified that intact cycling females were more sensitive to mild natural stressors and pharmacological stressors compared with males and ovariectomized females. When intact females were further separated based on estrous cycle stage, it was clear that the heightened sensitivity occurred during proestrus and could be reinstated in ovariectomized rats after estradiol replacement (Shansky et al., 2004). Moreover, studies testing the effects of ovarian hormones on susceptibility to social stressors have reported similar findings. When testing the behavioral responses to witness stress, intact females were exquisitely sensitive to this test, demonstrating anxiety-like responses during the stressor and development of depressive-like responses the following week as measured by the sucrose preference and FST (Finnell et al., 2018). Strikingly, ovariectomized females were resistant to these behavioral adaptations during and after social stress. Moreover, estradiol administration reinstated the behavioral response during witness stress in ovariectomized females, yet administration was also daily via a time-release pellet and thus not recapitulating the cycle (Finnell et al., 2018). In all, these data speak to the clear divergent effects that ovarian hormones may produce on stress susceptibility, potentially dependent upon age of animals, length of time past ovariectomy, dosing regimen for estradiol replacement, and type of stress paradigm (or lack of stress exposure). More controlled studies are needed to determine under what conditions ovarian hormones are behaviorally protective versus deleterious.
Impact of ovarian hormones on behavior and stress susceptibility
Impact of Antidepressant Therapy on Inflammation
As discussed earlier, stress dramatically activates inflammatory processes such as microglial activation and cytokine release and is known to contribute to stress-induced MDD (and behavioral dysfunction in rodents). Moreover, growing evidence supports the notion that exaggerated activation of the innate immune system may play a significant role in the pathogenesis of MDD and anxiety disorders in patients. In fact, several studies have linked treatment resistance to increased proinflammatory cytokines or microglial activation (Maes et al., 1997; Musselman et al., 2001b; Alesci et al., 2005; Motivala et al., 2005; Miller et al., 2009; Raison et al., 2013; Setiawan et al., 2015; Kiraly et al., 2017). As such, there are several important therapeutic considerations that we have investigated in the following section: 1) whether current efficacious treatment of stress-related psychiatric disorders impact cytokine expression, 2) whether anti-inflammatory therapies may serve as antidepressants in certain patient populations, and 3) whether there are known sex differences in these therapies.
Anti-Inflammatory Effects of Antidepressant Compounds.
Inflammation is greatly associated with the pathogenesis of depression (Hannestad et al., 2011; Gao et al., 2019). Moreover, patients treated with antidepressants show lower plasma levels of proinflammatory cytokines (Hannestad et al., 2011); however, it is unclear whether the anti-inflammatory effects of these drugs contribute to the antidepressant efficacy. Data presented by Hannestad et al. report that antidepressant treatment appears to lower the levels of IL-1β in the serum of patients treated for MDD while having little to no effect on the levels of TNF-α and IL-6 present in the serum. Moreover, although it is unclear as to whether the anti-inflammatory effects of antidepressants contribute to their antidepressant efficacy, it is striking that some studies report that cytokine changes predict the therapeutic response. For example, in a study testing the tricyclic amitriptyline, patients with MDD exhibited greater TNF-α plasma levels than nondepressed controls, and a reduction in TNF-α levels after 6 weeks of treatment predicted those who responded to the drug, whereas nonresponders lacked a significant anti-inflammatory effect after treatment (Lanquillon et al., 2000). Unfortunately, to our knowledge, it is not known whether sex differences exist in the anti-inflammatory effects of antidepressant compounds. However, given the significant number of sex differences in the pharmacokinetics of antidepressants based on differences in the rates of gastric emptying, amount of adipose tissue, and estrogenic regulation of cytochrome P450, as reviewed in LeGates et al. (2019), it would be expected.
The impact of antidepressants on inflammation is also supported by preclinical findings. For example, the selective serotonin reuptake inhibitor imipramine has been shown to decrease levels of microglial reactivity in rats exposed to an endotoxin stress (LPS) and a social stressor (repeated social defeat), while also attenuating the expression of TNF-α and IL-1β, although levels of IL-6 did not appear to be affected by this drug (Ramirez et al., 2015). In rats exposed to chronic social isolation stress, treatment with the antidepressant fluoxetine and the antipsychotic clozapine reduced the amount of TNF-α present in the hippocampus (Todorović and Filipović, 2017). The ability of antidepressants to inhibit microglial function has also been supported by findings in microglial cell culture. For example, three different antidepressants were shown to inhibit IFN-γ–induced microglial production of IL-6 and nitric oxide: the selective serotonin reuptake inhibitor fluvoxamine, the noradrenaline reuptake inhibitor reboxetine, and the nonselective monoaminergic reuptake inhibitor imipramine (Hashioka et al., 2007). Alternatively, this study also demonstrated that lithium chloride (a mood stabilizer), enhanced IL-6 release yet inhibited nitric oxide. The effects of antidepressants on proinflammatory components can vary depending on the drug and the cytokine being studied, indicating that different antidepressants may hold a certain degree of anti-inflammatory effect but likely cannot be considered an overall treatment of stress-induced inflammation (Hashioka et al., 2007).
With the recognition of treatment resistance in up to 30% of patients with MDD (Al-Harbi, 2012), the use of alternative therapies is essential. (R,S)-Ketamine is an N-methyl-d-aspartate antagonist that is not a traditional antidepressant but does exhibit antidepressant efficacy in patients with MDD (Berman et al., 2000) and has been shown to buffer learned fear expression in rodent models (McGowan et al., 2017). Moreover, (R,S)-ketamine serves as a prophylactic against stress-induced depressive-like behaviors in rodent models (Brachman et al., 2016; Mastrodonato et al., 2018, 2020). In mice in which LPS was the inflammatory precursor to the depressive-like behavior, (R,S)-ketamine prior to exposure to LPS attenuated learned fear if treatment occurred 1 week before re-exposure to a stressful condition (Mastrodonato et al., 2020). However, if treated 1 month or 1 day before the stress exposure, (R,S)-ketamine was ineffective, indicating that the timing of the treatment is also essential in the efficacy of the drug (McGowan et al., 2017). Along with the attenuation of depressive-like behaviors, the use of (R,S)-ketamine has also been identified to exhibit anti-inflammatory properties. (R,S)-Ketamine was shown to inhibit LPS-induced serum levels of TNF-α and IL-1β (Clarke et al., 2017). Additionally, in adult rats that had been stressed through maternal deprivation at a young age, treatment with (R,S)-ketamine reduced serum levels for TNF-α, IL-1, and IL-6 (Réus et al., 2015). Although the efficacy of (R,S)-ketamine as an antidepressant is understood, much is yet to be explored as to its effects on inflammation, as well as possible sex differences that could be present in the use of this drug as treatment of MDD; however, a few studies in rodents do suggest that females may be more sensitive to the antidepressant-like effects of ketamine compared with males (Carrier and Kabbaj, 2013; Franceschelli et al., 2015).
Antidepressant-Like Effects of Anti-Inflammatory Compounds.
Considering the evidence that inflammatory molecules are capable of producing a depressive phenotype, anti-inflammatory compounds have been tested for their antidepressant efficacy in clinical trials. Etanercept, a TNF-α antagonist, was tested in patients with psoriasis and found to produce significant antidepressant efficacy above that of placebo (Tyring et al., 2006). Moreover, another TNF-α antagonist, infliximab was tested in another population of patients with MDD and was also shown to exhibit antidepressant efficacy (Raison et al., 2013). However, depressive symptoms were only reduced for the subset of patients that presented with high baseline levels of inflammation prior to treatment. This suggests that subpopulations of depressed patients with elevated inflammatory profiles may benefit from anti-inflammatory compounds as an anti-inflammatory or for use as an adjuvant to traditional treatment.
Studies evaluating sex differences in the antidepressant efficacy of anti-inflammatory compounds are limited. One such study evaluated sex differences in the behavioral response to chronic unpredictable stress in male and female mice (Wohleb et al., 2018). Although both sexes demonstrated the development of depressive- and anxiety-like behaviors, only male mice exhibited increased microglial activation in the prefrontal cortex. Therefore, viral-mediated knockdown of neuronal colony stimulating factor 1 to inhibit the activation of microglia in the prefrontal cortex was only tested in males (Wohleb et al., 2018) and did produce an antidepressant and anxiolytic effect. Sex differences in drug pharmacokinetics and controlled anti-inflammatory studies from other diseases, on the other hand, have shed light on how men and women respond differently to the same therapy. For example, it has long been recognized that aspirin, an effective anti-inflammatory compound, has distinctly different pharmacokinetics in males versus females; the drug is cleared more slowly and has a longer half-life in women, owing to the greater bioavailability in women than in men (Ho et al., 1985). In a randomized clinical trial of IFN-β for multiple sclerosis, women responded better to IFN-β than men (Secondary Progressive Efficacy Clinical Trial of Recombinant Interferon-Beta-1a in MS (SPECTRIMS) Study Group, 2001). Moreover, other findings in patients with rheumatoid arthritis reported that after anti-TNF compounds were administered, only female patients exhibited increased plasma adiponectin (Nagashima et al., 2008), a molecule responsible for its anti-inflammatory effects (Mangge et al., 2010). It should also be noted that women respond to TNF-α antagonists with shorter half-lives, highlighting the critical point that, for pharmacological treatments (Desroches et al., 2010), serum concentrations should be used to tailor the dosing regimen for optimal therapeutic outcomes. This also begs the question as to whether women, who have greater inflammatory responses to social stressors and greater autoimmune disorder risk, may benefit from anti-inflammatory therapy as antidepressants or adjuvants to traditional therapy to a greater extent than men.
Similar to findings in clinical reports, there is also considerable evidence to support an antidepressant-like effect of anti-inflammatory compounds in preclinical studies in rodents. For example, treatment via central administration of an IL-1 receptor antagonist to male rats prior to daily exposure to social defeat inhibited the development of anhedonia in the sucrose preference test (Wood et al., 2015). Similarly, using chronic unpredictable stress, IL-1 receptor antagonist or IL-1 receptor knockout mice blocked the development of anhedonia that occurred after stress (Koo and Duman, 2008). Moreover, inhibiting IL-6 during repeated social defeat stress also produces an antidepressant-like response in mice (Hodes et al., 2014). Although, to our knowledge, studies have not been conducted to assess sex differences in anti-inflammatory efficacy within stress models, the vast sexual dimorphism of the inflammatory system suggests that these anti-inflammatory compounds will likely exhibit differential effects in males versus females.
Flavonoids are metabolites commonly derived from a variety of plant life, such as fruits, vegetables, herbs, grains, and red wine (Erdman et al., 2007); they have been studied for their potential therapeutic properties, especially in their role as anti-inflammatory agents (Serafini et al., 2010; Farzaei et al., 2019). For example, avicularin, a quercetin derivative, decreased levels of proinflammatory cytokines and subsequent depressive-like behaviors in rodents exposed to chronic, unpredictable, mild stress (Shen et al., 2019). In addition, daily treatment with resveratrol prior to repeated social defeat in male rats proved anti-inflammatory (Finnell et al., 2017a). Moreover, resveratrol produced an antidepressant-like effect by blocking anhedonia in susceptible rats, but only at doses that were efficacious in blocking neuroinflammation within the LC. It is important to note that some flavonoids affect the regulation of many biologic processes, including that of hormones such as estrogens (Kao et al., 1998; Rathee et al., 2009). Certain subgroups of flavonoids, such as flavones, inhibit the production of estrogen, acting as a competitive inhibitor of the aromatases that serve as the precursor of the hormone (Kao et al., 1998). Understanding these sex differences in the potential off-target effects of these compounds is crucial when taking into consideration the treatment of women versus men. Another major pitfall of these compounds is that they are not selective for one system and have many effects beyond reducing inflammation. Zheng et al. (2013) identified that the antidepressant-like effect produced by the flavonoid used in their studies occurred through the compound’s actions within the D1/D2 dopamine receptors, rather than anti-inflammation alone.
Putative Anti-Inflammatory Effects of CRF Antagonists.
It has long been recognized that subsets of depressed patients exhibit elevated levels of the stress-related neuropeptide corticotropin-releasing factor (CRF) (Owens and Nemeroff, 1991). Considerable evidence has now accumulated pointing toward inherent sex differences within the CRF system as a likely mechanism leading to enhanced stress susceptibility and thus increased rates of depression in the female population (Bangasser and Valentino, 2012; Bangasser and Wiersielis, 2018). Although research is far from elucidating the etiology of this health disparity, the molecular and cellular substrates identified in the CRF system that are differentially regulated in females versus males may consist of the most promising lines of evidence to understand exaggerated stress susceptibility in females (Bangasser and Valentino, 2012). It is notable that there is a bidirectional stimulatory relationship between cytokines and CRF (Dentino et al., 1999; Kageyama and Suda, 2009). Acute, short-term administration of CRF elicits anti-inflammatory effects through prostaglandin E2 synthesis (Tsatsanis et al., 2007), and although this study did not evaluate sex differences, female cells are known to release less prostaglandin E2 upon stimulation compared with male cells (Batres and Dupont, 1986). Alternatively, upon prolonged exposure to CRF (greater than 6 hours), a proinflammatory effect ensues, as measured by induction of TNF-α gene transcription (Wang et al., 2003; Tsatsanis et al., 2007). Importantly, a CRF1 antagonist is also capable of blocking the effect of cytokines on stress-related behaviors (Knapp et al., 2011). Thus, the exaggerated CRF system observed in women (Brady et al., 2009; Bangasser and Valentino, 2012) may contribute to the enhanced inflammatory responses to stressors in females (Cizza et al., 2009; Prather et al., 2009) and subsequent increased risk of suffering from depressive disorders.
Conclusions
With the long-recognized significant health disparity in MDD, i.e., women exhibiting twice the risk as men, it is surprising that we are not further in our understanding of how to personalize antidepressant treatments between the sexes (Kornstein et al., 2000). The fact that the majority of basic research studies have been conducted only in male subjects no doubt contributes to our lack of clarity on this subject. In the studies that do address this issue, vast sex differences have been identified in the pharmacokinetics of antidepressant therapies.
The female LC-NE system is well recognized to exhibit distinct morphological and neurochemical differences compared with males that predispose females to exaggerated LC-NE activity [as reviewed in Bangasser et al. (2016)]. As an extension of this knowledge, we propose that an LC-NE-cytokine circuit (Fig. 1) may serve as a conduit to increase stress susceptibility in females. Social stress–induced activation of the LC stimulates circulating NE release within the plasma and is necessary for the inflammatory priming that occurs after stress (Finnell et al., 2019). As a result, enhanced LC-NE activity in females can lead to hypersecretion of NE and, ultimately, increased circulating inflammation. Combined with evidence that peripheral inflammatory challenges serve to increase LC burst firing in an IL-1–dependent manner (Borsody and Weiss, 2004), we hypothesize that the exaggerated stress-induced inflammatory response evident in females may contribute to a feed-forward mechanism resulting in heightened IL-1β release in the periphery and exaggerated accumulation in the LC compared with males (Wood et al., 2010, 2015; Finnell et al., 2017a). This pathway is already primed to increase proinflammatory cytokine release in the face of stress and, given the knowledge that stress-induced LC-NE activity and cytokine responses are sensitized in females, suggests that this pathway may be hyperactive in stressed females versus males. Taken together, with the increasing appreciation for the role that inflammation may play in the pathogenesis of patients with MDD and the increased inflammatory stress responsivity observed in women, this favors the evaluation of anti-inflammatory therapies. In support of this, autoimmune disorders are also biased toward females and are comorbid with MDD, further highlighting the role that inflammatory cytokines may play in the pathogenesis of these stress-related psychiatric disorders. The studies reviewed herein strongly support the need for further studies to elucidate whether females are exquisitely sensitive to anti-inflammatory compounds as adjuvants to traditional therapies.
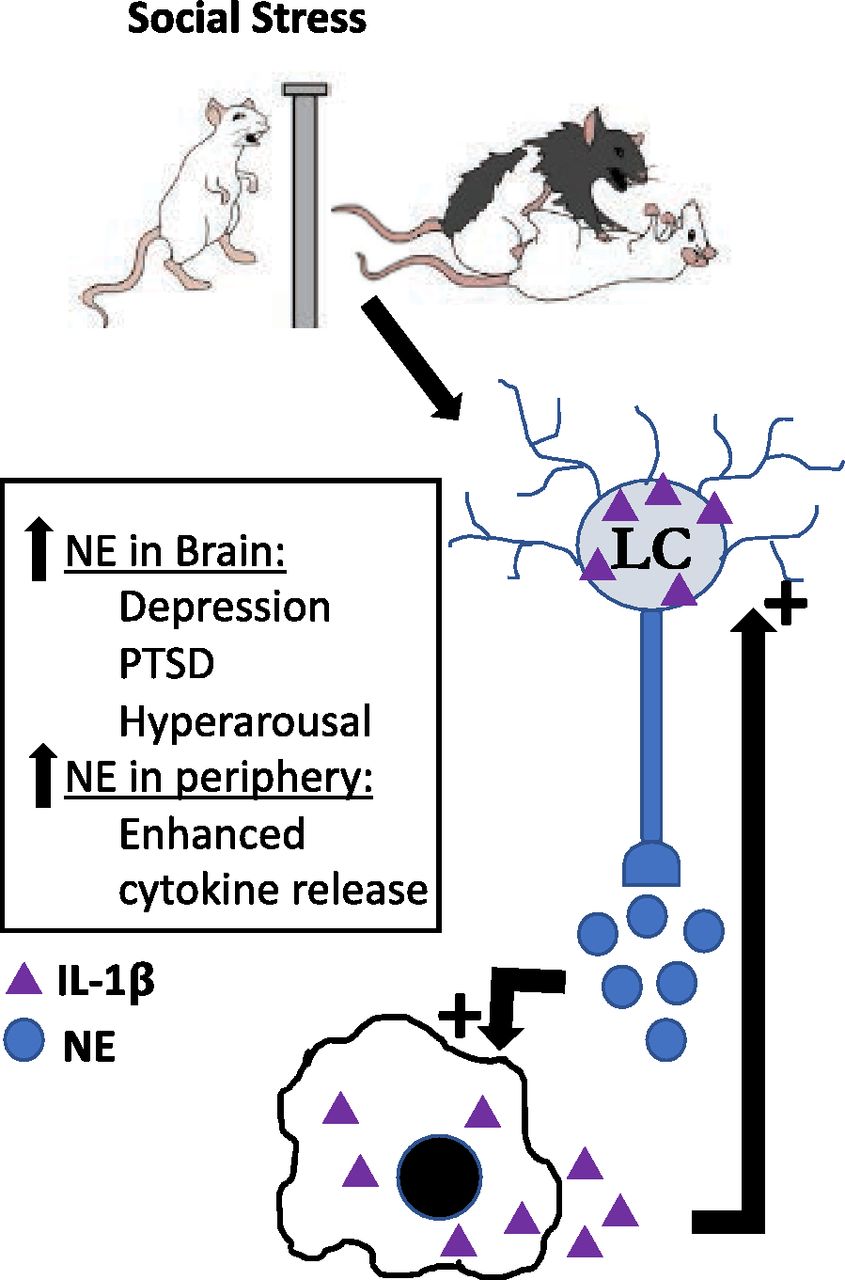
LC-NE-cytokine circuit as a conduit through which stress could increase stress susceptibly in females. The female LC-NE system exhibits distinct morphological and neurochemical differences compared with males, predisposing females to exaggerated LC-NE activity [as reviewed in Bangasser et al. (2016)]. As an extension of this understanding, we propose that an LC-NE-cytokine circuit may serve as a conduit through which stress functions to increase stress susceptibility (and increased risk of depression) in females. Social stress–induced activation of the LC stimulates circulating NE release within the plasma. This LC-induced NE release stimulates the inflammatory priming that occurs after stress (Finnell et al., 2019). Thus, enhanced LC-NE activity in females can lead to NE hypersecretion and ultimately increased circulating inflammation. Combined with evidence that peripheral inflammatory challenges serve to increase LC burst firing via IL-1 within the LC (Borsody and Weiss, 2004), we hypothesize that the exaggerated stress-induced inflammatory response evident in females may contribute to a feed-forward mechanism resulting in LC-NE–induced elevations of IL-1β release in the periphery as well as exaggerated accumulation within the LC compared with males (Wood et al., 2010, 2015; Finnell et al., 2017a).
Authorship Contributions
Performed data analysis: Martinez-Muniz, Wood.
Wrote or contributed to the writing of the manuscript: Martinez-Muniz, Wood.
Footnotes
- Received May 7, 2020.
- Accepted July 28, 2020.
This work was supported by National Institutes of Health National Institute of Mental Health [Grant MH113892], the Brain & Behavior Research Foundation, National Alliance for Research in Schizophrenia and Depression Young Investigator Award [26809], and the Department of Veterans Affairs [I21 BX002664].
Abbreviations
- CMS
- chronic mild stress
- CRF
- corticotropin-releasing factor
- ER
- Estrogen Receptor
- FST
- forced swim test
- HPA axis
- hypothalamic-pituitary-adrenal axis
- IFN
- interferon
- IL
- interleukin
- LC
- locus coeruleus
- LPS
- lipopolysaccharide
- MDD
- major depressive disorder
- NE
- norepinephrine
- PASAT
- paced auditory serial addition task
- PTSD
- Post traumatic stress disorder
- TSST
- Trier social stress test
- TNF
- tumor necrosis factor
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
Jump to section
- Article
- Abstract
- Introduction
- Recruitment of the Innate Immune Response after Psychological Stress
- Link between Inflammation and Depression
- Impact of Ovarian Hormones on Behavioral and Immune Responses to Stress
- Impact of Antidepressant Therapy on Inflammation
- Conclusions
- Authorship Contributions
- Footnotes
- Abbreviations
- References
- Figures & Data
- Info & Metrics
- eLetters