


Visual Overview

Abstract
Here we describe for the first time the distinctive pharmacological profile for (3S)‐3‐(2,3‐difluorophenyl)‐3‐methoxypyrrolidine (IRL752), a new phenyl-pyrrolidine derivative with regioselective central nervous system transmission-enhancing properties. IRL752 (3.7–150 µmol/kg, s.c.) was characterized through extensive in vivo studies using behavioral, tissue neurochemical, and gene expression as well as microdialysis methods. Behaviorally, the compound normalized tetrabenazine-induced hypoactivity, whereas it was unable to stimulate basal locomotion in normal animals or either accentuate or reverse hyperactivity induced by amphetamine or MK-801. IRL752 induced but minor changes in monoaminergic tissue neurochemistry across noradrenaline (NA)- and dopamine (DA)-dominated brain regions. The expression of neuronal activity–, plasticity-, and cognition-related immediate early genes (IEGs), however, increased by 1.5-fold to 2-fold. Furthermore, IRL752 dose-dependently enhanced cortical catecholamine dialysate output to 600%–750% above baseline, whereas striatal DA remained unaltered, and NA rose to ∼250%; cortical and hippocampal dialysate acetylcholine (ACh) increased to ∼250% and 190% above corresponding baseline, respectively. In line with this cortically preferential transmission-promoting action, the drug was also procognitive in the novel object recognition and reversal learning tests. In vitro neurotarget affinity and functional data coupled to drug exposure support the hypothesis that 5-hydroxytryptamine 7 receptor and α2(C)-adrenoceptor antagonism are key contributors to the in vivo efficacy and original profile of IRL752. The cortical-preferring facilitatory impact on catecholamine (and ACh) neurotransmission, along with effects on IEG expression and cognition-enhancing features, are in line with the potential clinical usefulness of IRL752 in conditions wherein these aspects may be dysregulated, such as in axial motor and cognitive deficits in Parkinson disease.
SIGNIFICANCE STATEMENT This report describes the distinctive preclinical profile of (3S)‐3‐(2,3‐difluorophenyl)‐3‐methoxypyrrolidine (IRL752). Its in vivo neurochemical, behavioral, microdialysis, and gene expression properties are consistent with a cortically regioselective facilitatory impact on catecholaminergic and cholinergic neurotransmission accompanied by cognitive impairment–reversing features. The pharmacological characteristics of IRL752 are in line with the clinical usefulness of IRL752 in conditions wherein these aspects may be dysregulated, such as in axial motor and cognitive deficits in Parkinson disease.
Introduction
Traditionally, Parkinson disease (PD) is primarily associated with its well-known motor manifestations, such as slowness and poverty of movements, resting tremor, muscular rigidity, and postural dysfunction. It is however becoming increasingly recognized that several nonmotor symptoms are also prominently represented in PD, some of which may also temporally precede the motor impairments (Chaudhuri et al., 2015; Sauerbier et al., 2016). Among the nonmotor symptoms in PD, cognitive deficits are highly prevalent (e.g., Willis et al., 2012). Cognitive symptoms may present early (Aarsland et al., 2009), with their prevalence rising dramatically over the course of the disease. Cognitive impairment is also a strong and independent predictor of mortality in patients with PD (Grimbergen et al., 2009; Willis et al., 2012; de Lau et al., 2014), and among those that do survive more than 20 years, dementia is present in >80% of cases (Hely et al., 2008). The number of elderly people with PD is likely to grow as global life expectancy increases. As drug treatments for cognitive impairment associated with neurodegenerative disorders, such as PD, are nonexistent (apart from the modestly effective Alzheimer dementia agents), improvements in this area would represent a significant advance to address this key unmet medical need in PD care (see Seppi et al., 2011; Szeto and Lewis, 2016). It is clear that the lack of efficacious treatments for cognitive dysfunction in PD has deleterious effects on the patients’ quality of life, burdens the caregivers, and causes substantial health costs to society (Vossius et al., 2011; Leroi et al., 2012; Kowal et al., 2013).
The exact underlying biologic substrates for cognitive impairment and dementia in PD have yet to be pinpointed. Deficits in cortical catecholaminergic and cholinergic transmission as well as in the signaling in cortico-striatal neurocircuitries have, however, been implicated (Beeler et al., 2013; Shepherd, 2013; Schirinzi et al., 2016). Subcortical Lewy body– and Alzheimer disease–related neuropathology has also been reported to be associated with cognitive decline in PD (see Weintraub and Burn, 2011, and references cited within). Interestingly, axial motor symptoms relating to postural instability (balance) and falls are strongly related to cognitive deficits in PD (Pantall et al., 2018). Both of these symptom clusters respond poorly to current anti-Parkinson treatment (Hely et al., 2008). It is probable that cognitive deficits in different PD subjects have heterogeneous pathologic and neurochemical substrates (e.g., Goldman and Holden, 2014). This notwithstanding, it may be assumed that, broadly, novel treatments aiming at strengthening cortico-striatal connectivity through enhanced monoaminergic and cholinergic network function could become beneficial in this context. One of the aims of our ongoing Research and Development projects has therefore been to develop a molecule that would meet the aforementioned profile.
(3S)‐3‐(2,3‐difluorophenyl)‐3‐methoxypyrrolidine (IRL752; Fig. 1) is a novel compound discovered and developed in an in vivo systems pharmacology (see Waters et al., 2017) program aiming to generate novel agents with a preferential enhancing effect on cortical versus subcortical catecholamine [dopamine (DA) and noradrenaline (NA)] output. The current report describes the actions of this agent on regional neuronal transmission in vivo, behavior, cognition, and synaptic activity–related gene expression in the rat. Overall, the data are consistent with the characterization of IRL752 as a cortical-preferring catecholamine- and cognition-promoting agent. Its profile is suggestive of potential usefulness for symptoms and signs in PD associated with deficits of cortical function, such as cognitive impairment and postural/axial motor dysfunction.

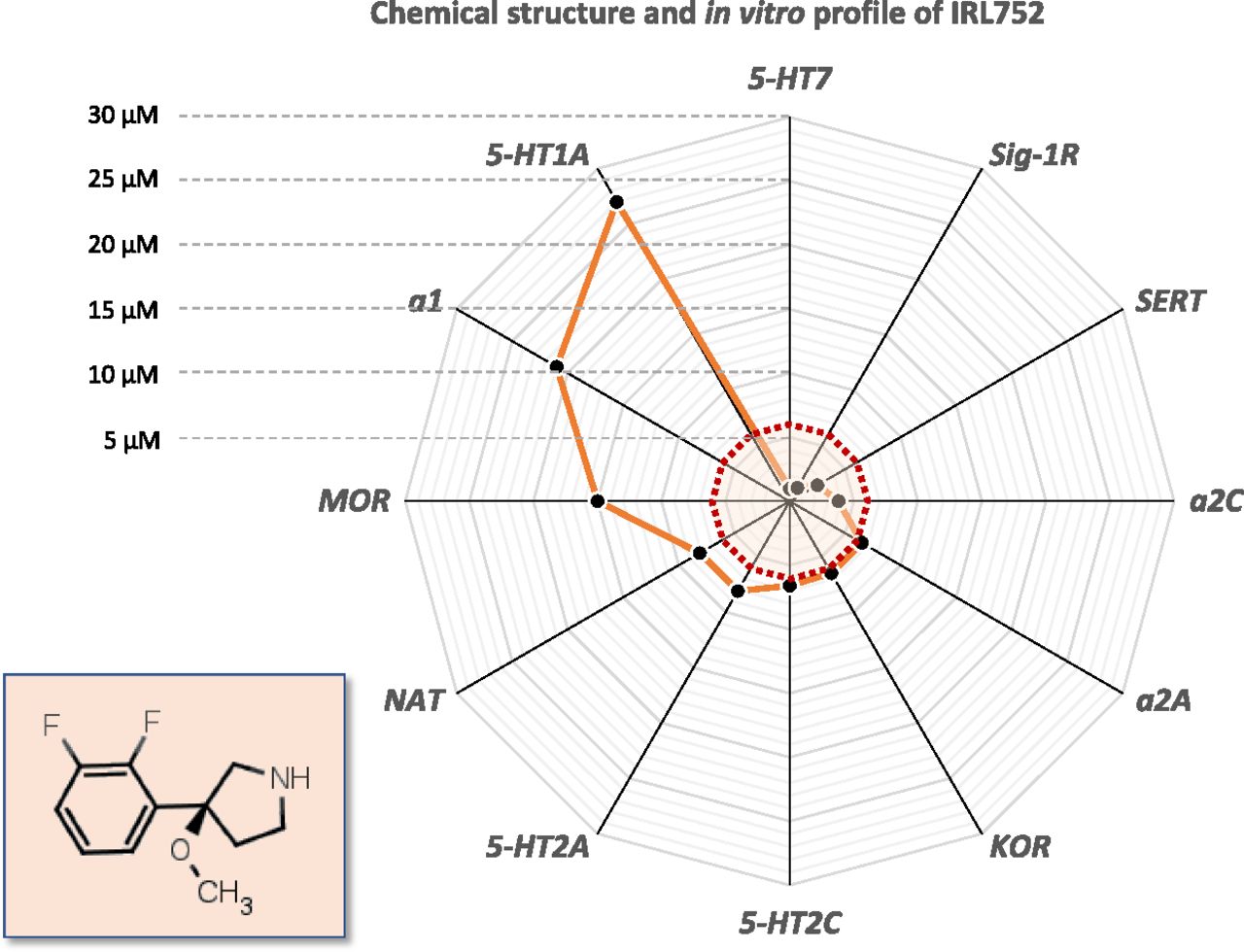
Chemical structure and radar diagram illustrating the target binding profile of IRL752. Shown is the chemical structure (inset) and target binding profile of IRL752, with Ki data plotted (filled circles) in a radar-type diagram with an excentric scale from 0 to 30 µM. Dotted, color-shaded area depicts estimated [IRL752]brain, free levels 60’ after administration of doses between 11 and 50 µmol/kg s.c. (see Results)—corresponding to the Ki concentration-range up to 6 µM, within which, i.a. the 5-HT7, σ1 receptor (Sig-1R), and α2C-adrenoceptor affinities are found: the closer to center, the higher affinity (see further, Table 1). KOR, κ opioid receptor; MOR, μ opioid receptor.
Materials and Methods
Animals.
Behavioral, in/ex vivo neurochemistry, gene expression, and in vivo microdialysis studies were carried out in adult male Sprague-Dawley rats (Taconic; weight 290–320 g at the time of use in microdialysis and 220–250 g in neurochemistry and gene expression studies). Upon arrival from the supplier, the animals were housed two to four per cage under standard controlled laboratory conditions (temperature 20–22°C; humidity 40%–70%; lights on/off 0700/1900 hours; 20–25 air exchanges/h) with food (standard rat chow; R36 Lactamin or similar: protein 20%–25%, carbohydrates 40%–50%, fat 3%–5%) and tap water allowed ad libitum until the animals were used in the experiments. The cages were equipped with enrichment materials, such as wooden sticks, plastic tunnels, etc.
Cognition studies were carried in a total of two cohorts of 60 female Lister Hooded rats (Harlan, UK, for the Bradford study and Charles River, UK, for the Manchester study): 60 for novel object recognition (NOR) and 60 for the reversal learning (RL) study; weights were 220–260 g and 275–300 g, respectively, at the time of testing. The NOR study was performed at the University of Bradford, and the RL study was performed at the University of Manchester. All animals were housed in groups of 5 under standard laboratory conditions under a 12-hour light:dark cycle (lights on at 0700 hours). At Manchester, rats were housed in individually ventilated, two-tiered plastic cages (38 × 59 × 24 cm, GR1800 Double-Decker Cage; Tecniplast, UK). These cages contained paper sizzle nest, sawdust, and cardboard tunnels (Datesand Group, UK). In Bradford, rats were housed in standard, nonventilated, single-tiered caging, as described in Sutcliffe et al. (2007). The NOR cohort were allowed free access to food, whereas the RL cohort, prior to operant training and testing, was gradually food-deprived to approximately 90% of free-feeding body weight; reduced body weight was maintained by restricting the amount of food given to each rat per day (approx. 12 g/day). Testing was carried out in the light phase. Female rats were used for these studies because it has been shown that female rats show a greater response to NMDA receptor antagonists than males (Moy and Breese, 2002), an effect we can confirm (B.G., personal communication). Furthermore, we have shown that female rats perform better in certain cognitive tasks than males (Sutcliffe et al., 2007).
All studies were carried out in compliance with European Union Directives and Guidelines for the care and use of animals in experiments and were approved by the local Animal Ethics Committee (microdialysis: 182/10 and 23/14; behavior/neurochemistry/gene expression: 279/10, 140/11, and 91/14) on behalf of the Swedish Department of Agriculture. The cognition studies were conducted in accordance with the Animals Scientific Procedures Act (UK, 1986) and approved by each University’s Animal Welfare and Ethical Review Board. All efforts were made to keep the number of animals used to the minimum acceptable for robust statistical assessment of treatment outcomes.
Drugs.
IRL752 [(3S)‐3‐(2,3‐difluorophenyl)‐3‐methoxypyrrolidine × HCl] was synthesized in-house and dissolved in physiologic saline (0.9% w/v NaCl). The drug was injected subcutaneously in a volume of 5 ml/kg, either 4–5 minutes before start of locomotor activity recordings (i.e., 64–65 minutes before termination of the animals and subsequent sampling for neurochemical and gene expression analysis) or immediately before start of microdialysis sampling. For interaction studies, rats were pretreated with MK-801 (maleate, 0.2 or 0.7 mg/kg; Sigma-Aldrich, UK) or d-amphetamine (sulfate, 1.5 mg/kg; AKL, Sweden) dissolved in 0.9% NaCl and given intraperitoneally 90 or 10 minutes, respectively, before start of locomotor recordings (i.e., 150 or 70 minutes, respectively, before termination of the animals and subsequent sampling for neurochemical and gene expression analysis). Tetrabenazine (0.64 mg/kg, s.c.; Sigma-Aldrich) was given in 0.9% NaCl (5 ml/kg) together with IRL752 at 5 minutes before start of locomotor activity recordings (i.e., 65 minutes before termination of the animals and subsequent sampling for neurochemical and gene expression analysis).
For the cognition model studies, rats were pretreated with either 2.0 mg/kg phencyclidine (PCP) (Sigma) dissolved in 0.9% NaCl or vehicle (0.9% NaCl), our standard subchronic PCP (scPCP) dosing regimen shown to produce robust and stable cognitive deficits (for reviews see Neill et al., 2010; Cadinu et al., 2018). PCP was administered intraperitoneally twice daily for 7 days, and this was followed by a 7-day drug washout period (see Fig. 4 for the study design). IRL752 was dissolved in 0.9% NaCl and administered in a volume of 5 ml/kg, s.c., 30 or 45 minutes prior to testing. Risperidone (0.1 mg/ml; Sigma) was dissolved in a minimum volume of acetic acid, made up to volume with distilled water pH-adjusted to 6 with 0.1 M NaOH, and administered intraperitoneally in a volume of 1 ml/kg 60 minutes prior to testing. SKF38393 (Sigma) was dissolved in 0.9% NaCl and administered intraperitoneally in a volume of 1 ml/kg 60 minutes prior to testing. In all experiments, the appropriate vehicle treatment was used. All drug doses were calculated as base equivalent weight.
Doses of IRL752 were selected based on in-house data. The dose range of IRL752 in the RL test was reduced (4.7–47 µmol/kg) compared with the NOR test (15.6–140 µmol/kg) because IRL752 at the highest dose tested (140 µmol/kg) was found to significantly reduce responding such that only two rats completed the task (see Results below and Supplemental Material). Risperidone (0.1 mg/kg) and SKF38393 (6 mg/kg) were selected as positive controls at the doses tested in this study based on their efficacy to improve scPCP-induced cognitive deficits in previous studies in our laboratory (Grayson et al., 2007; McLean et al., 2009). These drugs would also be expected to represent indirect and direct, respectively, means of enhancing D1 receptor–mediated neurotransmission, as they are examples of agents with D2 (auto)receptor blocking and direct D1 receptor–stimulating properties.
In Vitro Target Binding and Functional Studies.
The in vitro binding and functional characterization of IRL752 were performed by CEREP (Celle l’Evescault, France; contracted by IRL AB for these studies) across a panel of 65 selected targets using standardized binding and/or functional assays (see https://www.eurofinsdiscoveryservices.com/cms/cms-content/services/in-vitro-assays/). Inhibition constants (Ki) were calculated using the Cheng-Prusoff equation [Ki = IC50/(1 + (L/Kd)), in which L = concentration of radioligand in the assay, and Kd = the equilibrium dissociation constant; affinity of the radioligand for the receptor]. A Scatchard plot was used to determine the Kd.
In/Ex Vivo Brain Monoaminergic Neurochemistry.
Post-mortem concentrations of several monoaminergic indices (DA, NA, 5-HT, DOPA, HVA, 3-MT, 5-HIAA) were determined in striatal, limbic, and cortical tissue; collected; and dissected 64 minutes after administration of IRL752 (3.7–100 µmol/kg, s.c.) to normal rats, immediately after a 60-minute motor activity recording session (see below). Tissue sample preparation and neurochemical content analysis of monoamines and their metabolites (standard high-performance liquid chromatography with electrical detection) were carried out as described elsewhere (Waters et al., 2017).
Behavior: In Habituated and Nonhabituated, Nonpretreated Rats and in Monoamine-Depleted, Hyperdopaminergic, or Hypoglutamatergic States.
The behavior of rats was recorded in sound- and light-attenuating motor activity–monitoring chambers equipped with infrared movement detectors (two rows, each consisting of 16 sensors placed 2.5 cm apart and placed in a 90° angle along the front and side of the floor of the cage; Digiscan Activity Monitor RZYCCM-16 TAO; Omnitech Electronics), thus allowing for high-resolution assessment of motor patterns across a 40 × 40 cm–sized arena using several different sampling frequencies (0.25–25 Hz) in parallel (for further details, see Waters et al., 2017). Only data from the 25-Hz sampling channel (distance traveled) are shown in the Results.
Motor activity recordings started 4 minutes after injection of IRL752 (3.7–100 µmol/kg, s.c.) and covered 0–60 minutes thereafter. Motor activity studies were carried out in a variety of conditions, including during drug effects on the spontaneous nonhabituated locomotion/exploration as well as after habituation to the novel environment. In addition, the effect of different doses of IRL752 was assessed in acutely monoamine-depleted, hyperdopaminergic, and hypoglutamatergic behavioral states, as induced by the vesicular monoamine transport inhibitor tetrabenazine (0.64 mg/kg, s.c., 4 minutes before recording), the DA-releasing agent d-amphetamine (1.5 mg/kg, i.p., 10 minutes before recording), or the glutamate NMDA receptor antagonist MK-801 (dizocilpine; 0.2 or 0.7 mg/kg, i.p., 90 minutes before recording), respectively.
Cognitive Behavioral Tests: NOR and RL.
The NOR test was performed as previously described in detail elsewhere (Grayson et al., 2007; Snigdha et al., 2010). Briefly, rats (N = 60; 50 subchronically treated with the NMDA receptor antagonist PCP and 10 treated with vehicle) were habituated in cage groups to an empty test box for 1 hour on day 1. After an additional 3-minute habituation period on day 2, rats were given two 3-minute trials separated by a 1-minute intertrial interval in the home cage. In the first (acquisition) trial, animals were placed in the test box and allowed to explore two identical objects (A1 and A2). In the second (retention) trial, animals were placed in the test box with one duplicate familiar object from the acquisition phase (to avoid olfactory trails) and one novel object. On the day of testing, rats were randomly assigned to six treatment groups (n = 9 to 10 per group) to receive an acute treatment with IRL752 (15.6, 47, and 140 µmol/kg, s.c., 30 minutes prior to testing), risperidone (0.1 mg/kg, i.p.), or vehicle. Behavior was filmed and scored by a trained experimenter who was blinded to the treatment groups. Total object exploration time (defined as the duration of time animals spent licking, sniffing, or touching the object but not including time spent standing or sitting on or leaning against the object) was recorded for each of the familiar and novel objects in the acquisition and retention trials; locomotor activity (defined as movement, measured by the number of lines crossed in both trials) was also monitored. Any animal that failed to explore one or both of the objects in the acquisition trial was excluded from the final analysis.
For the RL test, rats (N = 60) maintained at 90% free-feeding weight were trained to perform an operant reversal learning task and then tested using a method previously described in detail (Neill et al., 2016). During the PCP treatment phase, animals were not trained in the RL task to avoid an association between drug treatment and cognitive performance. On the day of testing, rats were randomly assigned to six treatment groups (n = 9 to 10 per group) to receive an acute treatment with IRL752 (4.7, 14, or 47 µmol/kg, s.c., 45 minutes prior to testing), the DA D1 receptor agonist SKF38393 (6 mg/kg, i.p.), or vehicle before testing in the reversal learning task. We tested a lower dose range here because of behavioral disruption observed at the highest dose of 140 µmol/kg in NOR and efficacy at the lowest dose tested, 15.6 µmol/kg, to avoid behavioral impairment and to determine the minimal effective dose. The percentage of correct responses was calculated in each phase. In the initial phase of the reversal task session, the reward contingency was the same as the previous day, and the test was terminated at 5 minutes or when the animal had earned 20 pellets of food, whichever occurred first. After the initial phase, the rat remained in the test chamber for a 2-minute time-out period in which the house light was extinguished to serve as a cue that the rule was about to change. In the reversal phase, the reward contingency was reversed, and the test was repeated as described above; again this lasted for 5 minutes or the time taken to earn 20 pellets.
In/Ex Vivo Gene Expression.
The expression of mRNAs encoding glutamate decarboxylase 67, glutamate dehydrogenase, proenkephalin as well as the immediate early genes (IEGs) activity-regulated cytoskeleton-associated protein (Arc), c-fos, early growth response protein 1 (egr1), neuronal PAS domain protein 4 (Npas4), brain-derived neurotrophic factor (BDNF), a postsynaptic density scaffolding protein (Homer1), and neuronal pentraxin-2 protein (Nptx2) was assessed by quantitative polymerase chain reaction in tissue homogenates of the striatum, hippocampus, frontal cortex, and limbic regions (containing nucleus accumbens, most parts of the olfactory tubercle, ventral pallidum, and amygdala) collected 65 minutes after administration of IRL752 (11–100 µmol/kg, s.c.) to normal rats immediately subsequent to completion of the behavioral activity–monitoring recording sessions described above. Tissue samples were stored at −80°C until further processing. For analysis details, see Supplemental Material.
In Vivo Microdialysis.
The effect of IRL752 on extracellular levels of monoamines and their metabolites (DA, NA, 5-HT, DOPA, 3-MT, HVA, and 5-HIAA) and of acetylcholine (ACh) was studied by means of dual-probe in vivo microdialysis methodology, as previously described (Ponten et al., 2010; Jerlhag et al., 2012). In brief, rats were anesthetized with isoflurane, and homemade (see Waters et al., 1993) I-shaped microdialysis probes (20,000 kDa cut off with an o.d./i.d. of 310/220 µm, HOSPAL; Gambro, Lund, Sweden) were stereotaxically (Paxinos and Watson, 2009) implanted into the prefrontal cortex (PFC) (AP +3.2, ML −0.7, DV −4.0; membrane length, m.l. = 2.5 mm) and striatum (AP +1.0, ML −2.6, DV −6.2; m.l. = 3.0 mm) or, alternatively, into the dorsal (AP −4.0, ML −2.5, DV −4.4; m.l. = 2.0 mm) or ventral hippocampus (AP −6.0, ML +4.8, DV −8.0; m.l. = 4.0 mm) and nucleus accumbens (AP +1.85, ML −1.6, DV −7.8; m.l. = 2.0 mm). Probes for monitoring the levels of ACh after IRL752 were placed in the PFC (AP +3.0, ML −0.7, DV −4.0; m.l. = 3.0 mm) and the ventral hippocampus (AP −5.2, ML +4.8, DV −8.0; m.l. = 3.0 mm). The rats were housed individually in their home cages for 36–48 hours before the dialysis experiments. The microdialysis was performed during daytime under freely moving conditions with the rats connected to a microperfusion pump (perfusion speed 2 µl/min) via a swivel system through which 40-µl samples were collected every 20 minutes. After at least 100 minutes perfusion to establish balanced fluid exchange and baseline dialysate levels of analytes (typically between 10.00 hours and noon), IRL752 was administered subcutaneously (injection volume 5 ml/kg); vehicle controls received corresponding 0.9% NaCl injection. Dialysate samples were then collected every 20 minutes for the consecutive 3 hours, and dialysate contents of monoamines and corresponding metabolites (see above) or of ACh were analyzed by high-performance liquid chromatography/electrical detection or liquid chromatography/tandem mass spectrometry techniques, respectively, according to methods described elsewhere (Jerlhag et al., 2012; Waters et al., 2017). At termination, the brains were removed rapidly and fixated in Accustain solution (Sigma-Aldrich) for verification of probe placement.
Central Nervous System Drug Exposure after Systemic or Local Administration.
The in vivo central nervous system (CNS) exposure of IRL752 was gauged in two preliminary experiments after either systemic or local intracortical perfusion of the drug. In the first experiment, the drug was administered to two rats at a dose of 16.7 µmol/kg, s.c., and PFC and striatal dialysates were sampled and analyzed for IRL752 concentrations for up to 3 hours thereafter. In a second experiment (n = 2), the drug was locally applied in consecutively increasing concentrations (1, 10, and 100 µM; 3 × 20 minutes perfusion/concentration level) to the PFC via the dialysis perfusion medium (2 µl/min), during which the effects on NA and DA output were simultaneously followed in the dialysates; levels of IRL752 were not analyzed in this experiment.
Statistics.
In/ex vivo neurochemistry.
Neurochemical results are presented as percent of control mean ± S.E.M. (n = 4 per group). Statistical significance versus corresponding controls was assessed using one-way ANOVA followed by Dunnett’s test (two-sided). Probability levels <0.05 were considered significant.
Motor activity.
Motor activity data are presented as accumulated motor activity counts 0–60 minutes, percent of corresponding vehicle controls, and mean ± S.E.M. (n = 4 to 5/group). Statistical significance was assessed using one-way ANOVA followed by Dunnett’s test (two-sided) versus controls or other relevant comparison treatment group. Probability levels <0.05 were considered significant.
In/ex vivo gene expression.
Gene expression results are presented as percent of control mean ± S.E.M. (n = 5/group). Statistical significance versus corresponding controls was assessed using one-way ANOVA followed by Dunnett’s test (two-sided). Probability levels <0.05 were considered significant.
Cognitive behavioral tests.
All data are expressed as the mean ± S.E.M. (n = 9 to 10). NOR data were analyzed via two-way ANOVA with factors of drug and exploration time of the two objects (two identical objects in the acquisition phase and novel and familiar objects in the retention phase) and via one-way ANOVA (line crossings). Time spent exploring the objects was analyzed using paired Student’s t test, and post hoc analysis was carried out after a one-way ANOVA followed by least significant difference t test (line crossings). RL data were arcsine transformed and then analyzed via one-way ANOVA to detect the main effect of drug treatment in the initial and reversal phases of the task. When a significant effect (P < 0.05) was detected, a post hoc least significant difference t test was performed to compare treatment groups with the appropriate control group.
In vivo microdialysis.
In vivo microdialysis data are expressed as percent of baseline values averaged from the levels of three consecutive samples collected prior to drug administration. Statistical significance was assessed by means of a three-way ANOVA for repeated measures, with time, treatment, and region as factors. Probability levels <0.05 were considered significant.
Results
In summary, IRL752 displayed its highest in vitro affinities for 5-HT and NA-related targets. The compound also induced dose-dependent and regioselective alterations in brain monoamine transmission indices and gene expression. Behaviorally, IRL752 had no significant effect on acute hyperdopaminergic or hypoglutamatergic motor responses but reversed deficits resulting from hypomonoaminergic function. The compound also proved effective in correcting subchronic PCP-induced impairments in models of cognitive function.
In Vitro Target Binding and Functional Profile of IRL752.
The in vitro profiling of IRL752 showed that the compound displayed its highest affinity at human 5-HT7 receptors (Ki = 980 nM) and Sigma-1 receptor (σ1) receptors (1.2 µM), whereas the affinity for 5-HT2A and 5-HT2C receptors was 8.1 and 6.6 µM, respectively (Fig. 1; Table 1). IRL752 also displayed affinity for the 5-HT transporter (SERT; Ki = 2.5 µM) and to a lesser extent the NA transporter (NAT; Ki = 8.1 µM). With respect to α-adrenoceptor subtypes, IRL752 was most potent at α2C and α2A (Ki = 3.8 and 6.5 µM, respectively), whereas the affinity for the α1 receptor was ∼3–6 times less potent (Ki = 21 µM). Affinity for the rat κ receptor was 6.5 µM. IRL752 was inactive (<20% binding at 10 µM) for the remainder of targets assessed (n = 55; see Supplemental Material).
IRL752 IC50/Ki values for selected human target proteins
For assay details, see https://www.eurofinsdiscoveryservices.com/cms/cms-content/services/in-vitro-assays/.
IRL752 was further investigated in vitro in cellular functional assays designed to detect intrinsic activity at human recombinant α2A, α2C, 5-HT2A, 5-HT2C, and 5-HT7 receptors. The compound did not show any agonist-like properties at the α2-adrenoceptor and 5-HT7 targets (up to 100 µM; unpublished data) but displayed antagonist effects at the α2A, 5-HT2A, 5-HT2C, and 5-HT7 receptors (see Table 2).
Antagonist effects in cellular functional assays (human receptors)
For assay details, see https://www.eurofinsdiscoveryservices.com/cms/cms-content/services/in-vitro-assays/.
IRL752 Effects on In/Ex Vivo Brain Monoamine Neurochemistry.
IRL752 (3.7–100 µmol/kg, s.c.) resulted in small, although regioselective, effects on post-mortem rat brain tissue levels of monoamines and their corresponding metabolites. Thus, the compound induced a significant dose-dependent reduction (max ∼15%) of cortical NA and of striatal 3-MT. There was also a trend to dose-related but nonsignificant reductions in limbic and striatal DA and in cortical DOPA after IRL752 but only marginal, inconsistent changes in HVA, 5-HT, and 5-HIAA across brain regions (Figs. 2 and 3). Levels of normetanephrine were not detectable in any of the regions analyzed (too-small peaks and/or obscured by nearby peaks) and are therefore left out of Fig. 3.

In/ex vivo experiment design overview. Vehicle or IRL752 was given 4 or 5 minutes prior to locomotor and behavioral monitoring recordings (see further Materials and Methods), and 60 minutes later this was followed by termination and dissection of brain tissue for subsequent analysis of regional levels of monoamine neurochemical indices and gene expression patterns. Doses and times are given in the overview boxes.
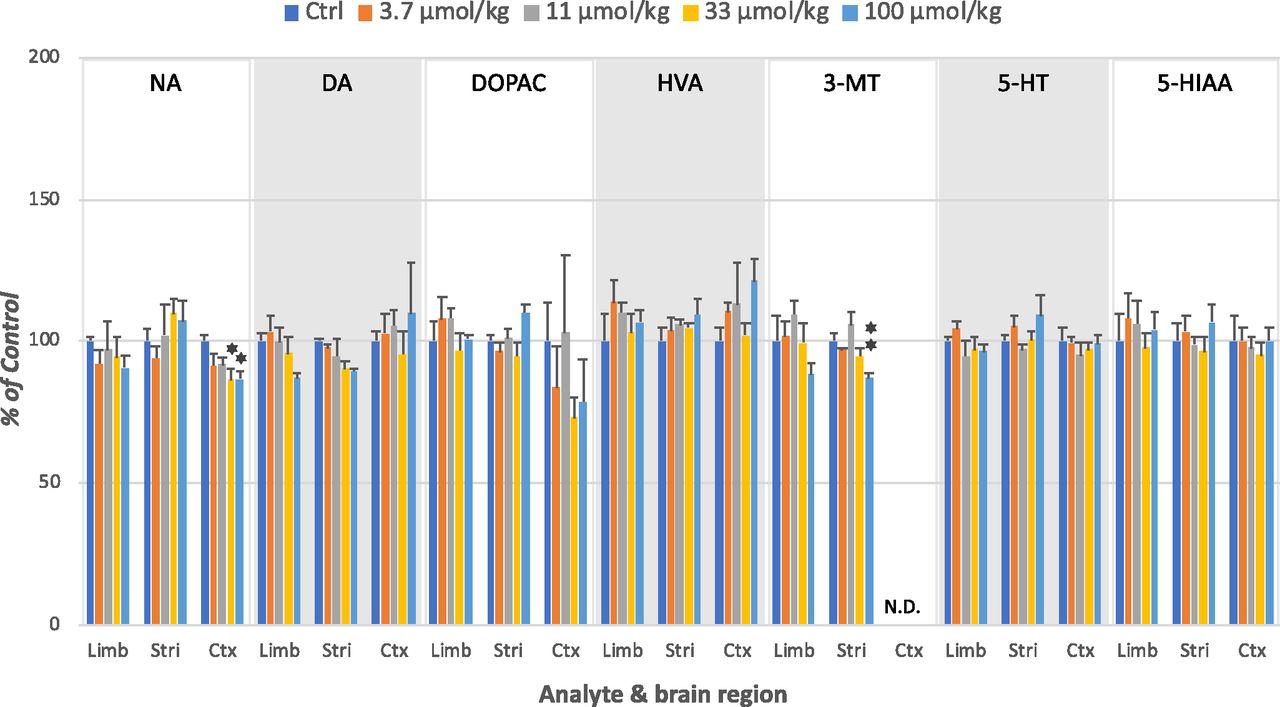
Effects of IRL752 on monoaminergic neurochemistry in different rat brain regions. Shown is the effect of IRL752 (0.8–21.3 mg/kg = 3.7–100 µmol/kg, s.c.) in the rat limbic (Limb), striatum (Stri), and cortical (Ctx) regions 65 minutes after drug administration. Monoaminergic indices measured were: NA, DA, DOPA, HVA, 3-MT, 5-HT, and 5-HIAA. Shown are the mean ± S.E.M. (n = 4) expressed as percentage of control (saline; NaCl 0.9% w/v) group means. Statistics: one-way ANOVA followed by Dunnett’s test (two-sided) vs. corresponding controls: *P < 0.05; **P < 0.025. Ctrl, control; N.D., not detectable.

Cognition experimental design overview. Subchronically PCP-treated animals were treated with either vehicle, IRL752, or reference agents 30, 45, or 60 minutes prior to cognitive model test sessions. Doses and pretreatment times for the different treatments and cognitive test models are given in the overview boxes.
Behavioral Assessment of IRL752.
The effects of IRL752 on locomotor activity in rats were assessed under different experimental conditions. To obtain a broad behavioral characterization, IRL752 was studied in otherwise untreated (acutely or after habituation) rats but also after tetrabenazine, d-amphetamine, or dizocilpine (MK-801) to assess the activity of the compound in acute hypomonoaminergic, hyperdopaminergic, and hypoglutamatergic brain states, respectively.
IRL752 effect on spontaneous motor activity in the untreated or habituated state.
In an initial study, IRL752 at doses of 3.7–100 µmol/kg had but a modest, nonsignificant impact on spontaneous locomotor activity in nonpretreated rats (max 50% ± 35% increase; Table 3). In a second study (unpublished data), doses from 33 to 300 µmol/kg were likewise without significant effect on spontaneous locomotor activity, indicating no major effects of the substance on psychomotor function in otherwise untreated animals. To check for any potential delayed impact on rat behavior, the effect of IRL752 (33 µmol/kg, s.c.) was also monitored 0–180 minutes postdosing but revealed no stimulation of locomotor activity by IRL752 compared with vehicle controls run in parallel (unpublished fata). Neither had IRL752 (11–100 µmol/kg, s.c.) had any consistent effect on locomotor activity during 0–60 minutes postdrug administration in rats habituated to their environment and expressing very low baseline activity (Table 3), thus indicating a lack of stimulant properties even when the sensitivity of the model was enhanced to capture subtle drug-induced behavioral activation.
Summary of effects of IRL752 on rat motor activity under different cotreatment conditions
Data expressed as accumulated motor activity counts 0–60 min, percent of corresponding vehicle controls, mean ± S.E.M. (n).
IRL752 effect on locomotor activity in d-amphetamine and MK-801 pretreated rats.
Across the dose range tested in nonpretreated rats (i.e., 11–100 µmol/kg, s.c.; see above), IRL752 did not influence d-amphetamine (1.5 mg/kg, i.p.)-induced hyperactivity (∼300% of non-IRL752 controls) in rats (Table 3). Similarly, IRL752 in the same dose range mildly and nonsignificantly, at best, affected locomotor hyperactivity in MK-801 (0.2 or 0.7 mg/kg, i.p.; ∼300% and 950% of non-IRL752 controls) pretreated rats (Table 3). Thus, whereas IRL752 at the dose of 11 µmol/kg tended to attenuate MK-801 (0.7 mg/kg)-induced hyperlocomotion, an additional study with IRL752 across 3.7–33 µmol/kg, s.c., did not further substantiate this observation (unpublished data). Similarly, IRL752 (100 µmol/kg, s.c.) only tended to partly—again nonsignificantly (P < 0.2)—reduce the behavioral activation induced by 0.2 mg/kg MK-801 (Table 3).
IRL752 effect on locomotor activity in tetrabenazine pretreated rats.
Tetrabenazine (0.64 mg/kg, s.c.) administration resulted in a 50% ± 5% inhibition of locomotor activity versus vehicle controls. This reduction was significantly reversed (to 91% ± 10% of vehicle control levels) by IRL752 (100 µmol/kg; Table 3).
IRL752 Effects in Cognitive Behavioral Tests.
The effects of IRL752 on cognitive functioning were assessed in the NOR and RL paradigms with regard to capacity to attenuate deficits induced by subchronic treatment with the NMDA receptor antagonist PCP. These tests are considered to reflect visual recognition memory, problem solving, and reasoning (Neill et al., 2010).
IRL752 effect on NOR.
There was no significant interaction between treatment and object exploration during the acquisition phase (two-way ANOVA). In addition, there were no significant differences in the exploration time of two identical objects for any group (Fig. 5a). In the retention phase, there was a significant interaction between treatment and object exploration (F5, 99 = 5.2; P < 0.001). Vehicle-treated rats spent significantly more time exploring the novel object compared with the familiar object (P < 0.05), whereas rats treated with scPCP showed deficits in the ability to discriminate between the novel and familiar objects (Fig. 5b). The scPCP-induced impairment in NOR was reversed by the two lower doses of IRL752 (15.6 µmol/kg, P < 0.01; 47 µmol/kg, P < 0.05) and by the reference D2 receptor antagonist risperidone 0.1 mg/kg (P < 0.001). The highest dose of IRL752 (140 µmol/kg) did not significantly attenuate the scPCP-induced impairments in NOR. Data did not demonstrate any significant reduction in line crossings in either phase of the task when compared with vehicle (see Supplemental Material). However, only two rats completed the task at the highest dose of 140 µmol/kg; the remaining eight rats failed to complete the test because of a reduction in exploratory behavior.

Effects of IRL752 in the NOR test. Shown is the effect of IRL752 (3.33–30 mg/kg = 15.6–140 µmol/kg, s.c., 30 minutes prior to testing) or risperidone (0.1 mg/kg i.p., 60 minutes prior to testing) on scPCP-induced impairment of novel object recognition in the rat. (a) Exploration time (s) of two identical objects (A and B) in the acquisition phase after acute treatment; (b) exploration time (s) of a familiar object and a novel object in the 3-minute retention trial. All data are expressed as the mean ± S.E.M. (n = 2–10 per group) and were analyzed by Student’s paired t test; significant differences between time spent exploring the familiar and novel object *P < 0.05; **P < 0.01; ***P < 0.001. sc, subchronic.
IRL752 effect on RL.
In the initial phase of the reversal learning task, there was no effect of drug treatment on the percentage of correct responses (Fig. 6, left). In the reversal phase of the task, there was a significant effect of drug treatment on correct responses (F5, 58 = 7.1; P < 0.001). scPCP treatment resulted in significantly impaired performance compared with vehicle (P < 0.001; Fig. 6, right). This impairment was dose-dependently and significantly attenuated by IRL752 to the level of vehicle control performance. Thus, although the lowest dose (4.7 µmol/kg) appeared ineffective compared with scPCP only, the middle dose (14 µmol/kg) approached significant attenuation levels (P = 0.055), and the highest dose (47 µmol/kg, P < 0.001) fully abolished the scPCP-induced impairment of RL (Fig. 6, right). Treatment with the reference D1 dopamine receptor agonist SKF38393 (6.0 mg/kg) also significantly reversed scPCP-induced impairments in this task (P < 0.01). There were no significant effects of any drug treatment on the total number of lever presses in this experiment (see Supplemental Material).
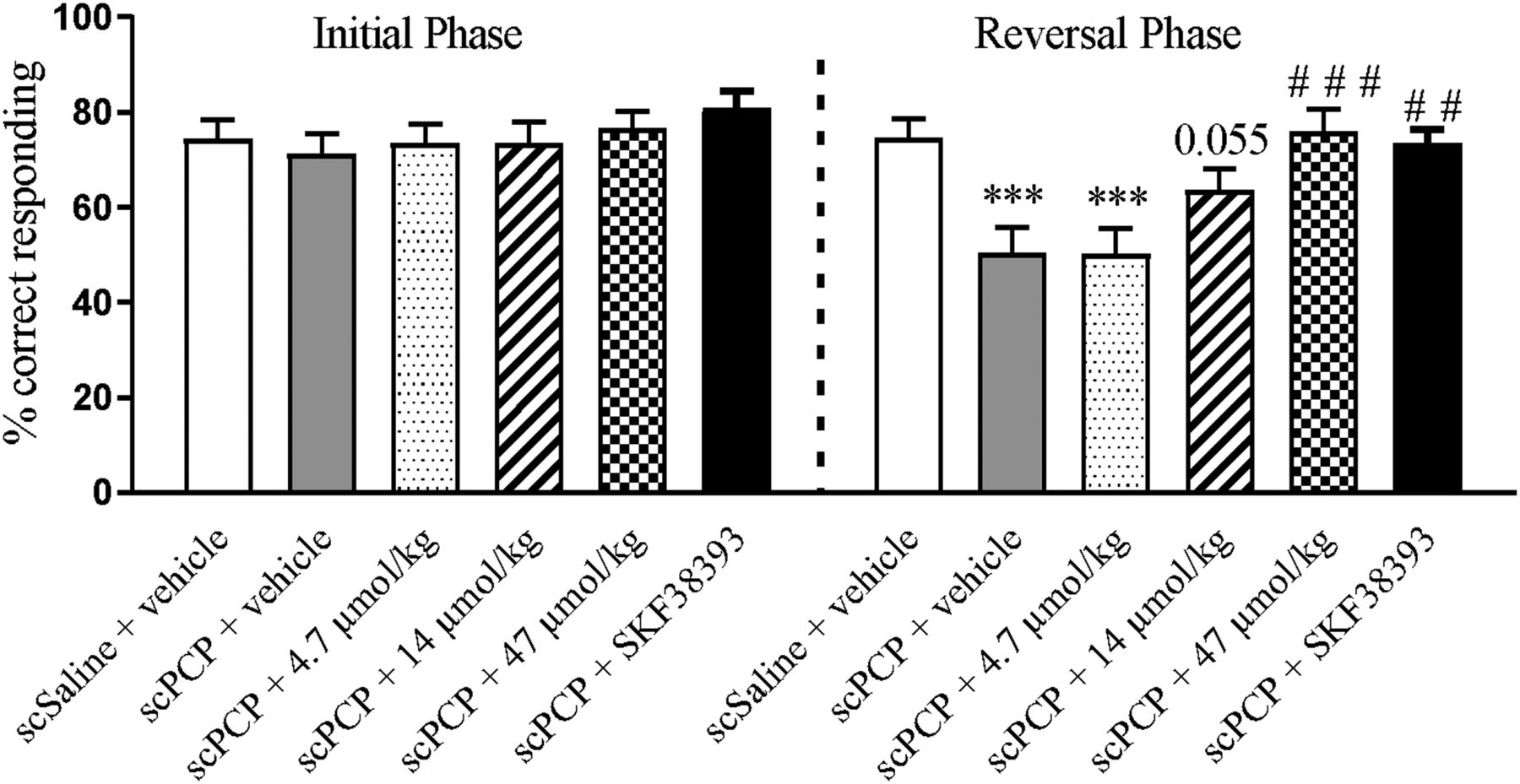
Effects of IRL752 in the RL test. The effect of acute treatment with IRL752 (1.0–10 mg/kg = 4.7–47 µmol/kg, s.c, 45 minutes prior to testing) and SKF38393 (6 mg/kg, i.p, 60 minutes prior to testing) on scPCP-induced impairment on performance in the reversal learning task. Data are shown as mean ± S.E.M. % correct responding (n = 9 to 10) and were analyzed by ANOVA and post hoc Dunnett’s test. Significant reduction in percent correct responding in the reversal phase compared with the vehicle group ***P < 0.001. Improvement in responding compared with scPCP alone in the reversal phase; P = 0.055; ##P < 0.01; ###P < 0.01. sc, subchronic.
IRL752 Effects on In/Ex Vivo Brain Tissue Gene Expression.
The most consistent effects on gene expression seen after IRL752 (11–100 µmol/kg, s.c.) were dose-related increases (∼1.5- to 2-fold those of corresponding controls) in the rapid IEGs Arc, c-fos, EGR1, and Npas4 mRNA, primarily in limbic, striatal, and cortical regions; the delayed IEGs BDNF, Homer1, and Nptx2 remained essentially unchanged across all brain areas, as did the hippocampal expression versus controls of the genes monitored (Fig. 7). No significant changes versus controls were found in the mRNA expression levels of glutamate decarboxylase, glutamate dehydrogenase, and proenkephalin, regardless of brain region (unpublished data).

Effects of IRL752 on gene expression in different brain regions, measured by quantitative polymerase chain reaction. Shown are the gene expression (mRNA levels) effects of IRL752 (2.4–21.3 mg/kg = 11–100 µmol/kg, s.c.) in the rat limbic region (Limb), striatum (Stri), frontal cortex (Ctx) and hippocampus (Hipp) 65 minutes after administration; controls received corresponding vehicle injection (saline; NaCl 0.9% v/w). Genes monitored were: Arc, c-fos (cellular DNA-binding proteins encoded by the c-fos genes), egr1, Npas4, BDNF, Homer1 (a postsynaptic scaffolding protein), and Nptx2. Genes are divided into “Rapid” and “Delayed” onset IEGs, as shown in the graph. Results are presented as percentage of controls (mean ± S.E.M.; n = 5 animals/treatment group). Statistics: one-way ANOVA followed by Dunnett’s test (two-sided), *P < 0.05; **P < 0.025; ***P < 0.01 vs. corresponding controls. Data are sorted by gene, then by region, and then by ascending dose from left to right. Ctrl, control.
IRL752: In Vivo Microdialysis.
The dose-response relationship for IRL752 with respect to the efflux of catecholamines in the PFC as compared with the striatum is illustrated in Figs. 8 and 9.
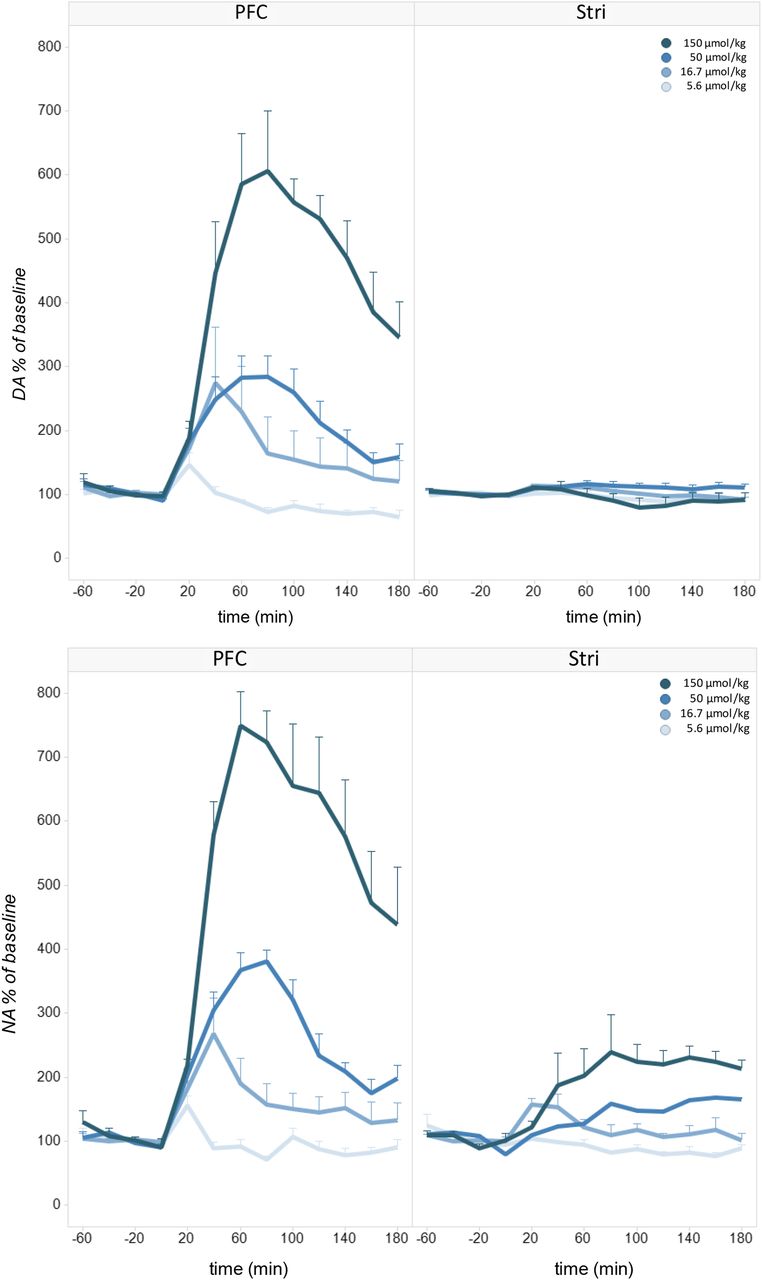
IRL752 time course of effects on rat brain regional dialysate levels of DA (top panels) and NA (bottom panels). Effect of IRL752 (5.6–150 µmol/kg, s.c.; dose color scheme from light to dark blue) on dialysate DA and NA in the rat PFC (n = 4–7; left panels) and striatum (Stri) (n = 3–6; right top panel, and n = 4 and n = 1–3 for 5.6–16.7 and 50–150 µmol/kg, respectively; right bottom panel) 0–180 minutes after administration. Shown are the mean dialysate levels/20-minute sample ±S.E.M. expressed as percentage of baseline values (average of the three fractions collected 40 to 0 minutes before dosing).

IRL752 time-course effects on rat brain regional dialysate levels of serotonin (5-HT). Effect of IRL752 (5.6–150 µmol/kg, s.c.; dose color scheme from light to dark blue) on dialysate 5-HT in the rat PFC (n = 2–7; left panel) and striatum (Stri) (n = 2–6; right panel), 0–180 minutes after administration. Shown are the mean dialysate levels/20-minute sample ±S.E.M. expressed as percentage of baseline values (average of the three fractions collected 40 to 0 minutes before dosing).
IRL752 (5.6–150 µmol/kg, s.c.) resulted in a dose- and time-related and regionally selective increase in extracellular monoamines and their metabolites. Thus, there was a dose-dependent increase in DA in the PFC peaking at ∼600% of baseline at 60–80 minutes after administration of the top dose [significant effect of 1): F4, 13 = 20.5, P = 0.000016, 2) region: F1, 13 = 58.1, P = 0.000004, 3) dose*time: F36, 117 = 6.4, P < 0.00001, 4) dose*region: F4, 13 = 16.2, P < 0.00001, and 5) dose*time*region: F36, 117 = 6.3, P < 0.00001; Fig. 8], whereas the levels of the transmitter remained essentially unchanged in typical DA-dominated brain areas like the striatum and the nucleus accumbens (Table 4). The regioselectivity of IRL752 extended also to NA in that the PFC levels of this amine increased to a maximum of ∼750% of baseline at 60 minutes postadministration of the top dose, whereas striatal NA peaked at only ∼250% of baseline [significant effect of 1) dose: F4, 8 = 37.7, P = 0.00003, 2) region: F1, 8 = 66.5, P = 0.00004, 3) dose*time: F36, 72 = 8.76, P < 0.00001, 4) dose*region: F4, 8 = 23.2, P = 0.00019, and 5) dose*time*region: F36, 72 = 6.3, P < 0.00001; Fig. 8]. The main DA metabolite, DOPA, increased only slightly in the PFC and remained unchanged in the striatum (unpublished data).
Comparison of the peak effects of IRL752 on extracellular amines in different brain regions
Shown is the peak effect of IRL752 (50 µmol/kg = 10.7 mg/kg, s.c) expressed as % of baseline in dorsal hippocampus, ventral hippocampus, nucleus accumbens, PFC, and striatum.
The impact of IRL752 on dialysate 5 HT was generally less marked; also, although dose- and time-dependent, it did not display significant regioselectivity (dose: F4, 11 = 7.47, P = 0.0037, region: F1, 11 = 0.46, P = 0.51, dose*time: F36, 99 = 3.25, P = 0.000002, and dose*region: F4, 11 = 0.12, P = 0.97; see Fig. 9; Table 4). The level of the main metabolite of serotonin, 5-HIAA, was not significantly changed over the dose range tested (unpublished data).
The effect of IRL752 (50 µmol/kg, s.c.) on dialysate monoamines across several different brain regions was also compared. Overall, the data show that IRL752 impacts the PFC and ventral hippocampal output of DA and NA to a larger extent than in other brain areas (Table 4).
IRL752 (50 µmol/kg, s.c.) elevated the dialysate levels of ACh to a maximum of ∼250% and ∼190% of corresponding baseline in the PFC and ventral hippocampus, respectively, in the 80’ postinjection samples (Fig. 10). Resembling the effects seen on catecholamines (above), there was a near-significant effect of region (F1, 5 = 6.15, P = 0.056) and dose*time*region (F9, 45 = 2.00, P = 0.061). Separate two-way ANOVA (repeated-measures) in the PFC showed significant effects of time (F13, 65 = 3.13, P = 0.001) and of dose*time (F13, 65 = 2.05, P = 0.03), whereas only the time effect proved significant in the ventral hippocampus (F13, 52 = 2.58, P = 0.008).

IRL752 time-course effects on ACh in rat PFC and ventral hippocampus dialysates. IRL752 was given at 50 µmol/kg s.c., and PFC (left panel) or ventral hippocampus (right panel) dialysate levels of ACh were monitored for 3 hours post–drug administration. Shown are the mean dialysate levels/20-minute sample ±S.E.M. expressed as percentage of baseline values (average of the three fractions collected from 40 to 0 minutes before dosing), mean ± S.E.M. (n = 2–4).
IRL752: CNS Drug Exposure—Levels and Functional Effect.
Preliminary experiments aiming to establish the relation between brain exposure of IRL752 and functional effects were carried out using microdialysis sampling and reverse dialysis perfusion of the drug.
IRL752 (16.7 µmol/kg, s.c.) resulted in peak concentrations of the drug in cortical and striatal dialysates, amounting to ∼400 nmol/l 40–60 minute after administration (see Supplemental Material). With an estimated probe recovery of IRL752 of ∼10%–12%, this corresponds to brain extracellular levels of the drug in the range of ∼3 to 4 µM. This is in fairly good agreement with an estimation of unbound brain concentrations around ∼2 µM at this dose, which was obtained by a different approach and derived from data on unbound peak plasma concentrations of IRL752 after 11 and 50 µmol/kg, s.c, assuming no active transport across the blood-brain barrier (unpublished data). Taking a conservative approach, the approximate free-CNS exposure would thus range between 1 and 6 µM in the 11–50 µmol/kg s.c. dose interval (Fig. 1). Local reverse dialysis perfusion of IRL752 (1, 10, and 100 µM; 3 × 20 minute/concentration level) via the probe into the PFC resulted in increased catecholamine output: 10 µM to ∼250% (DA) and ∼350% (NA), 100 µM to ∼360% (DA), and ∼650% (NA) of baseline levels (Supplemental Material).
Discussion
The in vivo profile of the novel phenyl-pyrrolidine IRL752 is consistent with its characterization as a multitarget, regioselective cortical neurotransmission enhancer with potential clinical usefulness in, for example, Parkinson disease dementia (PD-D) or other cognitive insufficiencies. The neurochemical properties of IRL752 suggest a concerted impact on PFC catecholamine and cholinergic neurotransmission and synaptic activity–related gene expression and a capability to reverse motor hypofunction and impairments in models of cognitive failure but without major effect in either normal or drug-induced hyperactive behavioral states. Taken together, the findings suggest a capacity to strengthen multiple systems believed to be impaired in motor and cognitive deficiency manifestations.
The pharmacological profile of IRL752 differs from other drugs known to modulate cortical monoamine transmission (e.g., monoamine reuptake inhibitors, psychostimulants, antidepressants, or antipsychotics). In addition to the striking cortical-over-basal-ganglia catecholamine transmission-promoting selectivity profile of IRL752, the brain tissue neurochemical data suggest that the compound does not to any major extent activate or inhibit negative feedback mechanisms on neurotransmission in monoaminergic projection areas. Moreover, its behavioral activity features indicate that IRL752 lacks stimulatory or inhibitory effects on motor function in normal states while reversing behavioral inhibition in tetrabenazine-treated rats—an experimental animal model of deficient catecholamine transmission, like in PD. Also, IRL752 does not affect d-amphetamine–induced hyperactivity and only mildly and inconsistently attenuated MK-801 (dizocilpine)-induced hyperactivity, which is thus indicative of low impact on acute hyperdopaminergic and hypoglutamatergic states.
Interestingly, IRL752 displayed clear-cut procognitive effects in the NOR and RL tests, reversing the cognitive impairment induced by scPCP administration. It has been shown that scPCP abolishes the increase in PFC DA during a NOR retention trial, which is suggested to underlie the accompanying cognitive deficiency seen after this treatment (McLean et al., 2017). In this context, the ability of IRL752 to enhance extracellular levels of DA and NA in the PFC but not in (the DA-dominated) striatal and nucleus accumbens areas is particularly noteworthy. The cortical catecholamine increases were likewise accompanied by increases in ACh in this region and (to a lesser extent) in the ventral hippocampus. The neurochemical signature as well as the behavioral profile of the compound is thus distinct from that of psychostimulants (e.g., phencyclidine and d-amphetamine) (Hertel et al., 1995) and indicates a disparate biologic substrate for the transmission-promoting effect of IRL752. The PFC is generally considered to be of key importance for cognitive function, with a pivotal role repeatedly being ascribed to intact catecholaminergic neurotransmission in this region (Arnsten, 1998; Robbins and Arnsten, 2009; Arnsten, 2011; Robbins and Cools, 2014; Millan et al., 2016). The association of ACh loss with deterioration of cognitive processes is likewise well-established. Indeed, there is ample evidence that executive and other cognitive functional impairment in PD-D may be associated with PFC DA as well as forebrain cholinergic deficits (Bohnen et al., 2003; Hilker et al., 2005; Narayanan et al., 2013; Gratwicke et al., 2015). Taken together, it is therefore suggested that IRL752 with its balanced and regionally selective transmission-strengthening properties across catecholamine and ACh systems may find clinical usefulness in conditions like PD-D.
The observed IRL752-induced changes in gene expression pattern are consistent with the above idea. Thus, the compound caused dose-dependent increases in the expression of mRNA for the rapid IEGs Arc, c-fos, egr1, and Npas4. The corresponding transcription factors and effector proteins are known to be linked to neuronal plasticity, recent neuronal and synaptic activity, and associated procognitive action (e.g., Okuno, 2011; Sun and Lin, 2016). For example, Arc is involved in the consolidation of memory and in connected processes, such as long-term potentiation, and is triggered intra-arterially by synaptic NMDA receptor–related mechanisms (Bramham et al., 2010) associated with enhanced monoamine transmission (Millan et al., 2016). Npas4 is uniquely activated by neuronal activity versus extracellular factors, is expressed only in neurones, and affects synaptic connections in both excitatory and inhibitory neurons, thereby providing another important IEG link between neuronal activity, learning, and memory (Sun and Lin, 2016).
The distinctive in vivo pharmacological profile of IRL752 likely reflects a joint impact on more than one target. Based on the in vitro characterization of IRL752, it is hypothesized that a combination of antagonism at 5-HT7 and α2-adrenoceptors may be primarily underlying the potent regioselective increase in PFC monoamine-dependent neurotransmission. Preliminary CNS IRL752 exposure data concur with this prediction. Although antagonism of 5-HT7 receptors has been demonstrated to contribute to enhanced cortical DA efflux, the neuronal localization of any such action remains to be determined (see Wesołowska and Kowalska, 2008; Blattner et al., 2019). Selective blockade of α2C-adrenoreceptors relative to (mainly postsynaptic; Erdozain et al., 2019) α2A-adrenoceptors also seems to promote DA in the PFC (see Uys et al., 2017a, and references cited within), whereas α2A-adrenoceptor antagonism may be important for boosting ACh output (Zaborszky et al., 2004). Although a potential contributory effect from inhibition of SERT cannot be entirely ruled out based on currently available data, the in/ex vivo neurochemical profile of IRL752 clearly differs from typical SERT inhibitors. Finally, the σ1 receptor affinity of IRL752 may additionally imply an effect on cell survival mechanisms and neuroplasticity (e.g., Nguyen et al., 2015). Notably, all the aforementioned targets are believed highly relevant to the functioning of cognition circuits and processes (e.g., Nikiforuk, 2015; Millan et al., 2016; Maurice and Goguadze, 2017; Uys et al., 2017a).
Multiple Target Action of IRL752.
Pharma drug development typically remains strictly (single…) target-focused. There is growing appreciation and optimism, however, that a more comprehensive approach might eventually prove more successful (Hutson et al., 2017). IRL752 was discovered using a multivariate in vivo phenotypic pattern (in vivo systems pharmacology) analysis, including neurochemical and behavioral effects broadly, benchmarked against a wide range of known, clinically characterized CNS agents (Waters et al., 2017). Although from a single neuroreceptor target point of view, IRL752 may be viewed as moderately potent, the relative lack of selectivity and potency may in fact turn out to be an advantage. First, needless to say, no single drug target works in isolation but is integrated with multiple other components in systems, networks, and circuits to adjust (patho)physiologically disrupted function. Secondly, factors like the sensitivity of, and tone at, the target, as well as occupancy-response relation, vary greatly among brain areas, transmitter circuits, and synapses, underscoring that “the brain is no democracy” and thereby providing an opportunity for regioselectivity in drug action. Thirdly, drug combination treatments to modulate the activity of more than one neuronal target are in fact very common in the treatment of symptoms associated with CNS disorders, whether they are of psychiatric and/or other cognitive background (e.g., Mojtabai and Olfson, 2010). Finally, together with the above, it may be hypothesized that the joint contribution of moderate modulatory activities at two or more neurotargets in a network by a single agent may result in superior therapeutic characteristics compared with combination attempts using several different potent and selective drugs (see, e.g., Csermely et al., 2005; Tun et al., 2011). Indeed, approved CNS drugs or (fixed-dose) combinations with intended polypharmacological (“enriched”) target profiles are becoming increasingly common in recent years (e.g., vortioxetine, lurasidone, brexpiprazole) in endeavors to both improve efficacy and circumvent side-effect issues (Peters, 2013).
Summary and Conclusions.
This paper describes for the first time the novel, distinctive pharmacological action profile of a new compound, IRL752. The behavioral characteristics of IRL752 are consistent with a capacity to normalize impaired catecholamine function while avoiding overactivation of CNS systems and circuits involved. In vitro target affinity, functional response, and accompanying drug exposure data suggest that at least 5-HT7 receptor and α2(C)-adrenoceptor antagonism are likely key contributors to the in vivo effects of IRL752. Interestingly, in comparison with other agents with clinically proven or purported procognitive properties, IRL752 displays similarities but also clearly distinguishing features (Table 5). Its 1) cortically regioselective, facilitatory impact on catecholamine neurotransmission, without significant feedback activation; 2) increases in ACh transmission and cognitive function-associated IEG indices; and 3) demonstration that IRL752 effectively reverses cognitive deficits in rodent tests of recognition memory and of cognitive flexibility/problem solving are in line with clinical usefulness of the drug in conditions wherein these aspects may be dysregulated, such as in PD axial motor and cognition deficits.
Summary overview of microdialysis, behavioral, and gene expression effects of IRL752 and selected procognitive comparators
Sources for comparator data cited: Atomoxetine: Bymaster et al., 2002; Swanson et al., 2006; Tzavara et al., 2006; Cain et al., 2011; IRL AB, unpublished data. Methylphenidate: Gerasimov et al., 2000; Bymaster et al., 2002; Marsteller et al., 2002; Berridge et al., 2006; Tzavara et al., 2006; Banerjee et al., 2009; Seu and Jentsch, 2009; Rowley et al., 2014; IRL AB, unpublished data. Clozapine: Hertel et al., 1997; Westerink et al., 2001; Ichikawa et al., 2002; Shirazi-Southall et al., 2002; Devoto et al., 2003; Shilliam and Dawson, 2005; Abdul-Monim et al., 2006; Grayson et al., 2007; Robbins et al., 2008; IRL AB, unpublished data. Cariprazine: Neill et al., 2016; Kehr et al., 2018; Huang et al., 2019; Waters et al., 2020; IRL AB, unpublished data. Vortioxetine: Mørk et al., 2013; Pehrson et al., 2013, 2018; du Jardin et al., 2014; Wallace et al., 2014; IRL AB, unpublished data. Atipamezole: Kauppila et al., 1991; Haapalinna et al., 1998; Tellez et al., 1999; Devoto et al., 2003; Lapiz and Morilak, 2006; Bondi et al., 2010; Mervaala et al., 1993; IRL AB, unpublished data. Idazoxan: Devauges and Sara, 1990; Coull et al., 1996; Hertel et al., 1997; Tellez et al., 1999; Swanson et al., 2006; Uys et al., 2017b; IRL AB, unpublished data. Fluparoxan: Tellez et al., 1999; Millan et al., 2000; Borthwick, 2017; IRL AB, unpublished data. Disclaimer: Permutations of experimental conditions and differences in set-ups between laboratories preclude direct head-to-head comparisons of efficacies and responses among agents. This applies broadly but may be particularly apparent with regard to cognitive behavioral testing.
In support, IRL752 improved postural dysfunction and axial motor symptoms (UDPRS motor scores) in a recent phase IIa study in patients with PD (Svenningsson et al., 2020). Cognitive deficits in PD are strongly associated with such motor impairments; pharmacologically boosting executive as well as attentional functional strategies will likely counter the motor shortcomings and prevent falls (e.g., (Taylor et al., 2008). Notably, significant improvement in the patients who were treated with IRL752 was also seen with respect to the motivation/initiative domain in the UDPRS assessment, a finding substantiated by greatly decreased scores for apathy/indifference and caregiver distress in the neuropsychiatric inventory. In conclusion, the preclinical profile of IRL752 supports its potential usefulness in the treatment of cognitive and axial motor deficits in PD. Data from phase I and IIa clinical trials with IRL752 indicate that it is safe and well-tolerated both in healthy volunteers and in patients with PD. Further exploration of IRL752 efficacy with respect to improved balance, reduced falls, and markers of improved cognition will be carried out in planned phase IIb trials.
Acknowledgments
We thank Anette Nydén, Håkan Schyllander, Maria Gullme, Sverker von Unge, and Fredrik Pettersson for synthesizing the IRL752 batches used in the study. The skillful technical assistance of Anna Carin Jansson, Marianne Thorngren, and Katarina Rydén Markinhuhta in performing the experiments described in this report is gratefully acknowledged.
Authorship Contributions
Participated in research design: Hjorth, S. Waters, N. Waters, Tedroff, McLean, Grayson, Neill, Sonesson.
Conducted experiments: Fagerberg, Edling, Svanberg, Ljung, Gunnergren, McLean, Grayson, Idris.
Performed data analysis: Hjorth, S. Waters, McLean, Grayson, Neill.
Wrote or contributed to the writing of the manuscript: Hjorth, S. Waters, N. Waters, Tedroff, Svensson, Fagerberg, Edling, Svanberg, Ljung, Gunnergren, McLean, Grayson, Idris, Neill, Sonesson.
Footnotes
- Received April 2, 2020.
- Accepted June 25, 2020.
No financial support was received for the studies reported. Financial disclosures: S.W., N.W., J.T., P.S., A.F., M.E., B.S., E.L., J.G., and C.S. are full- or part-time employees of Integrative Research Laboratories Sweden (IRL AB) and own stock in the company. S.H. is a consultant to IRL AB and owns stock in the company. In the past 3 years, S.H. has received honoraria for consultancy from Otsuka, Lundbeck, and Recordati. J.C.N., B.G., S.L.M., and N.F.I. conducted the cognition test studies reported in the manuscript via the Contract Research Organisation b-neuro, covered by a fee-for-service contract between University of Manchester (UoM) and IRL AB. In the past 3 years, J.C.N. has also received honoraria for consultancy from Janssen and Lundbeck.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- σ1
- Sigma-1 receptor
- ACh
- acetylcholine
- AP
- anterior-posterior
- Arc
- activity-regulated cytoskeleton-associated protein
- BDNF
- brain-derived neurotrophic factor
- CNS
- central nervous system
- DA
- dopamine
- DV
- dorsal-ventral
- EGR1
- early growth response protein 1
- 5-HIAA
- 5-hydroxyindoleacetic acid
- 5-HT
- 5-hydroxytryptamine
- HVA
- homovanillic acid
- IEG
- immediate early gene
- IRL752
- (3S)‐3‐(2,3‐difluorophenyl)‐3‐methoxypyrrolidine
- IRL AB
- Integrative Research Laboratories Sweden AB
- ML
- medial-lateral
- 3-MT
- 3-methoxytyramine
- NA
- noradrenaline
- NAT
- NA transporter
- NMDA
- N-methyl-D-aspartate
- NOR
- novel object recognition
- Npas4
- neuronal PAS domain protein 4
- Nptx2
- neuronal pentraxin-2 protein
- PCP
- phencyclidine
- PD
- Parkinson disease
- PD-D
- PD dementia
- PFC
- prefrontal cortex
- RL
- reversal learning
- scPCP
- subchronic PCP
- Copyright © 2020 The Author(s).
This is an open access article distributed under the CC BY Attribution 4.0 International license.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}