


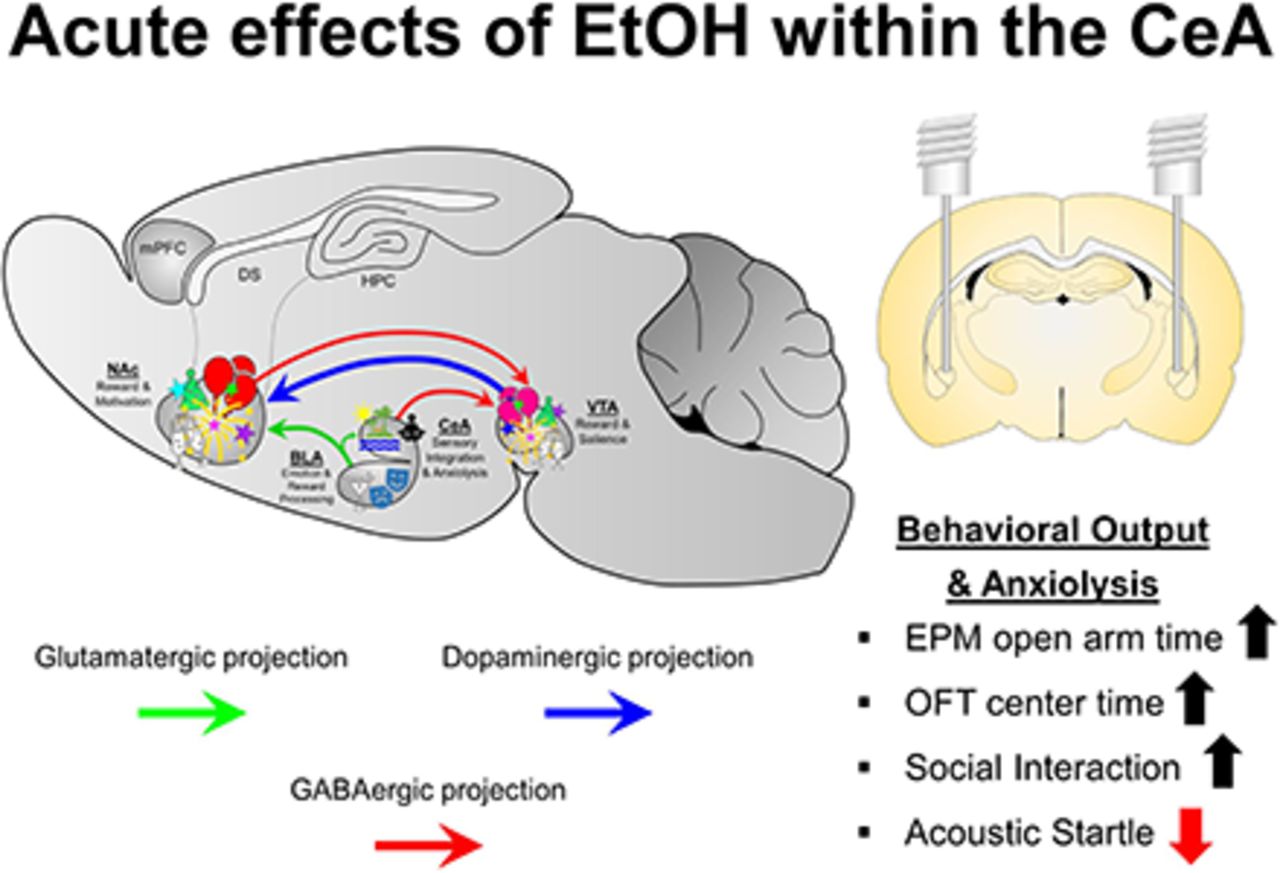
Visual Overview
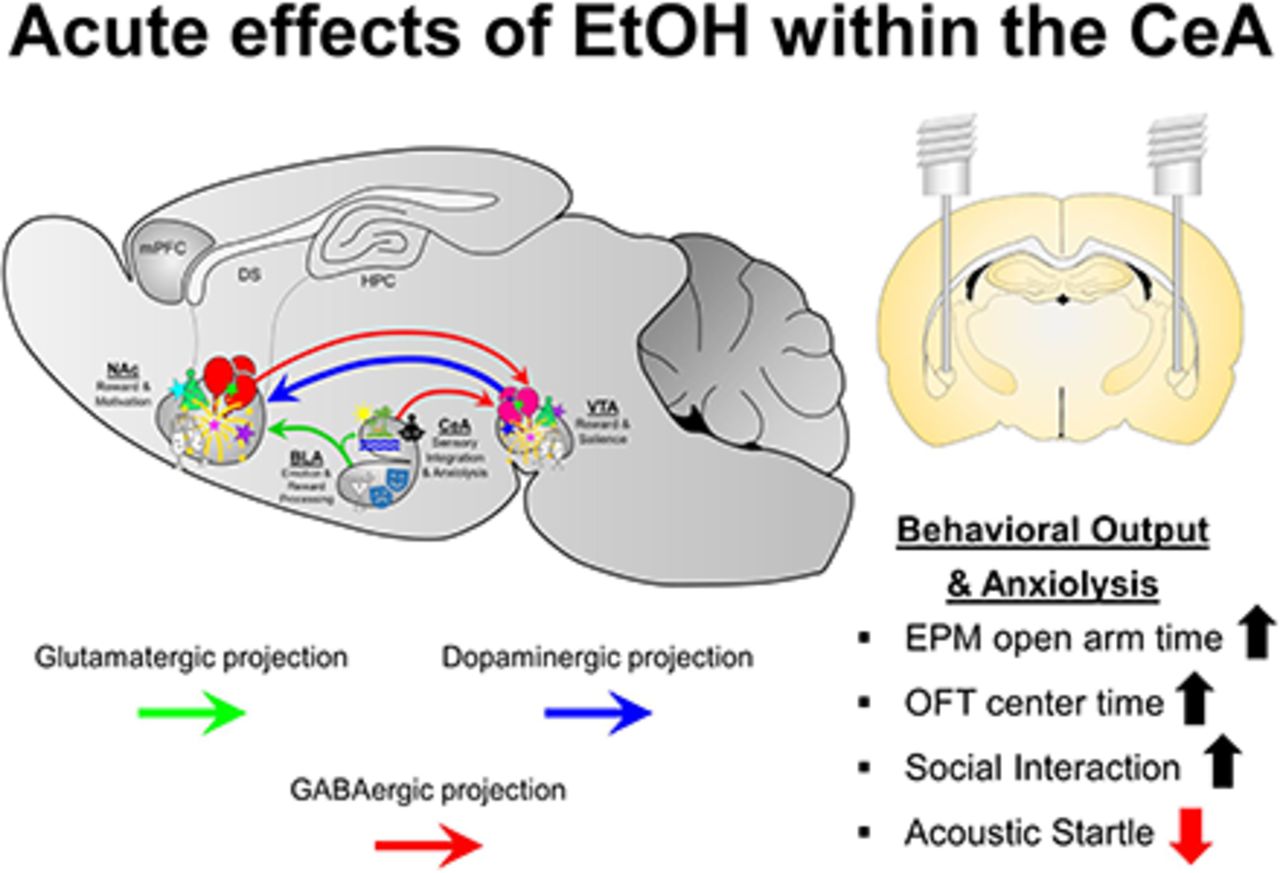
Abstract
In humans, alcohol is consumed for its rewarding and anxiolytic effects. The central nucleus of the amygdala (CeA) is considered a neuronal nexus that regulates fear, anxiety, and drug self-administration. Manipulations of the CeA alter ethanol (EtOH) consumption under numerous EtOH self-administration models. The experiments determined whether EtOH is reinforcing/anxiolytic within the CeA, whether selective breeding for high alcohol consumption alters the rewarding properties of EtOH in the CeA, and whether the reinforcing/anxiolytic effects of EtOH in the CeA are mediated by the neuropeptides corticotropin-releasing factor (CRF) and nociceptin. The reinforcing properties of EtOH were determined by having male Wistar and Taconic alcohol-preferring (tP) rats self-administer EtOH directly into the CeA. The expression of anxiety-like behaviors was assessed through multiple behavioral models (social interaction, acoustic startle, and open field). Coadministration of EtOH and a CRF1 antagonist (NBI35965) or nociceptin on self-administration into the CeA and anxiety-like behaviors was determined. EtOH was self-administered directly into the lateral CeA, and tP rats self-administered a lower concentration of EtOH than Wistar rats. EtOH microinjected into the lateral CeA reduced the expression of anxiety-like behaviors, indicating an anxiolytic effect. Coadministration of NBI35965 failed to alter the rewarding/anxiolytic properties of EtOH in the CeA. In contrast, coadministration of the nociceptin enhanced both EtOH reward and anxiolysis in the CeA. Overall, the data indicate that the lateral CeA is a key anatomic location that mediates the rewarding and anxiolytic effects of EtOH, and local nociceptin receptors, but not local CRF1 receptors, are involved in these behaviors.
SIGNIFICANCE STATEMENT Alcohol is consumed for the stimulatory, rewarding, and anxiolytic properties of the drug of abuse. The current data are the first to establish that alcohol is reinforcing and anxiolytic within the lateral central nucleus of the amygdala (CeA) and that the nociceptin system regulates these effects of alcohol within the CeA.
Introduction
Alcohol has multiple effects on the human body. At low doses, alcohol can have stimulatory effects, which have been associated with activation of the posterior ventral tegmental area (VTA) (Boehm et al., 2002). Further actions of alcohol include euphoria, anxiolysis, and sedation. The total intrinsic reinforcement value (valiancy) of alcohol likely incorporates these distinct biologic consequences of alcohol consumption, and discrete brain regions mediate specific action(s) of alcohol.
The central nucleus of the amygdala (CeA) is a neuronal nexus that mediates response to stress, negative affect processes, fear processes, and drug-related behaviors (Koob and Volkow, 2010). The CeA is primarily composed of GABAergic neurons and is part of the extended amygdala (Dong et al., 2001). The CeA innervates the bed nucleus of the stria terminalis in a topographically specific pattern derived from two primary pathways: the stria terminal and ansa penduncularis (Dong et al., 2001). The CeA also innervates regions of the hypothalamus and other cortical regions (Dong et al., 2001). The CeA–bed nucleus of the stria terminalis pathway is a critical regulatory system for fear and anxiety responses (Ciocchi et al., 2010; Davis et al., 2010). Neuropeptides have conflicting influences on fear and anxiety within the CeA. Corticotropin-releasing factor (CRF) is considered to be a proanxiety/prostress neuropeptide, whereas neuropeptide Y and nociceptin are thought to be anxiolytic/antistress neuropeptides (Karatsoreos, 2016).
Ethanol (EtOH) has differential acute and chronic effects within the CeA. EtOH effects within the CeA are complex and could be mediated by a number of factors that need to be properly examined. For example, multiple reports have indicated that EtOH acts to stimulate the release of GABA and CRF in the CeA (Roberto et al., 2003, 2004a,b, 2010; Cruz et al., 2012; Kallupi et al., 2014a,b). Other reports have indicated that EtOH acts to inhibit GABA release in the CeA (Silberman and Winder, 2013). Additional reports indicate that EtOH does not alter GABA levels in the CeA (Gilpin, 2012). A similar level of uncertainty exists for the effect of EtOH on glutamate levels in the CeA: reduction (Roberto et al., 2004a,b) or increase (Silberman and Winder, 2013; Silberman et al., 2015).
Despite the uncertainty of the method of action of EtOH in the CeA, the CeA mediates the self-administration of EtOH and other drugs of abuse. For example, optogenetic stimulation of the CeA enhances motivation for cocaine (Warlow et al., 2017), and muscimol + baclofen inactivation of the CeA decreases cue-induced methamphetamine seeking during withdrawal (Li et al., 2015). Lesions of the CeA alter voluntary EtOH consumption (Moller et al., 1997), and acute ethanol administration at doses as low as 0.25 g/kg induce neuronal c-fos immunoreactivity in the CeA (Morales et al., 1998). Optogenetic stimulation of basolateral amygdala (BLA) terminals in the CeA induces a reversible, anxiolytic effect, whereas inhibition of this circuit leads to increased anxiety-related behaviors (Tye et al., 2011).
Genetic predisposition to consume EtOH is associated with alterations to the response of stimuli within the CeA. Marchigian Sardinian alcohol-preferring rats have enhanced GABA signaling in the CeA, and CRF application in the CeA stimulates glutamate release, which is not observed in control rats (Herman et al., 2013, 2016). Alcohol-preferring (P) rats have reduced levels of neuropeptide Y (mRNA and protein) in the CeA (Pandey et al., 2005). Therefore, there may be different sensitivity to the effects of EtOH in the CeA predicated upon a genetic predisposition to consume high levels of EtOH.
EtOH is reinforcing directly within the mesolimbic dopamine system. EtOH and nicotine are directly self-administered into the posterior, but not anterior, VTA, and the sensitivity to EtOH and nicotine reward in the posterior VTA is enhanced in rats selectively bred for high alcohol consumption (Rodd-Henricks et al., 2000; Hauser et al., 2014, 2019). The nucleus accumbens also displays regional heterogeneity for the rewarding properties of EtOH and other drugs of abuse. EtOH and cocaine are directly self-administered into the nucleus accumbens shell (AcbSh) but not the core, and rats selectively bred for high alcohol consumption are more sensitive to the rewarding properties of these drugs of abuse (Engleman et al., 2009; Katner et al., 2011).
The initial experiments were designed to determine whether EtOH was reinforcing directly in the CeA. Serendipitously, after the data collection of the initial groups of animals, the current research team noticed a major difference in the behavior of the rats self-administering EtOH into the CeA from subjects of past intracranial self-administration (ICSA) experiments. In rats self-administering EtOH into the posterior VTA or AcbSh (an estimated 1400 rats), subjects were behaviorally activated and expressed a high level of locomotor activity upon return to their home cages. In contrast, in rats self-administering EtOH into the CeA, the subjects were extremely calm and malleable to handling by the research team. It was this observational detection that led to the hypothesis that the reinforcing property of EtOH in the CeA is based upon the anxiolytic effects of EtOH within this region.
The current experiments were conducted to determine: 1) whether EtOH is reinforcing within the CeA; 2) whether EtOH reward in the CeA is influenced by selective breeding for high alcohol consumption; 3) whether coadministration of a CRF1 antagonist or nociceptin would alter EtOH self-administration into the CeA; 4) the anxiolytic properties of EtOH within the CeA, posterior VTA, and AcbSh; and 5) whether coadministration of a CRF1 antagonist or nociceptin would alter the anxiolytic properties of EtOH within the CeA.
Materials and Methods
Animals.
Adult male Wistar (Envigo) and Taconic P (tP) rats weighing 300–400 g at the start of the experiment were used. Rats were maintained on a 12-hour reversed light/dark cycle (lights off at 0900 hours). The P rats used in the current experiments were from a substrain of P rats termed tP rats. Taconic P rats were separated from the Indiana University: School of Medicine (IUSM) stock of P rats in 2004 and maintained at Taconic Bioscience (Rensselaer, NY) until the rats were returned to IUSM in 2013. The tP rats were maintained for over 30 generations at Taconic Bioscience and represent a highly functional substrain of the P rat. The tP rats have been isolated from the standard P rats since their return to IUSM. Food and water were available ad libitum throughout the experiment, except during operant testing. The animals used in these experiments were maintained in facilities fully accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care. All research protocols were approved by the Institutional Animal Care and Use Committee and are in accordance with the guidelines of the Institutional Care and Use Committee of the National Institute on Drug Abuse, National Institutes of Health, and the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, Commission on Life Sciences, National Research Council 2011).
Surgery.
The details for stereotaxic surgery and postsurgery care are provided in published reports (e.g., Rodd-Henricks et al., 2000; Hauser et al., 2019). The coordinates (Paxinos and Watson, 1998) for the CeA were 2.3 mm posterior to bregma, 4.3 mm lateral to the midline, and 7.5 mm below the surface of the skull at a 0° angle to the vertical. The coordinates for the BLA, which served as an anatomic control, were 2.3 mm posterior to bregma, 5.1 mm lateral to the midline, and 7.8 mm below the surface of the skull at a 0° angle to the vertical. Coordinates for the posterior VTA were 5.8–6.1 mm posterior to bregma, 2.1 mm lateral, and 8.5 mm ventral from the surface of the skull at a 10° angle from the vertical. Coordinates for the AcbSh were 1.0–1.4 mm anterior to bregma, 1.2 mm lateral, and 8.0 mm ventral from the surface of the skull at a 10° angle from the vertical. A 28-gauge stylet that extended 0.5 mm beyond the tip of the guide was placed into the guide cannula between experimental sessions.
Intracranial Self-Administration Procedure.
The ICSA apparatus and standard procedures have been described in detail in published reports (e.g., Rodd-Henricks et al., 2000; Hauser et al., 2019). Briefly, each operant chamber has an active and inactive lever. Responding on the active lever results in a 100-nl infusion over 5-second time period. After each infusion, there is a 5-second timeout period in which responses are recorded but have no consequences. The inactive lever responses are recorded but have no consequences. The duration of each session was 4 hours, and sessions were conducted every other day. Experiments have determined that the diffusion of EtOH microinjected (100 nl/injection) into the brain was less than 0.1 mm from injection site (Gatto et al., 1994).
Microinjection Procedures for Discrete Brain Regions.
The equipment used for ICSA testing has been modified to microinject solutions into discrete brain regions (see Truitt et al., 2015; Deehan et al., 2018 for details). Briefly, using an electrolytic microinfusion transducer (EMIT) system, rats receive a 100-nl injection over 5-second time period followed by a 15-second timeout period (three injections per minute). Rats receive injections for a 10-minute time period (30 total microinjections).
Wistar and tP Male CeA EtOH ICSA Dose Response.
For the CeA EtOH dose-response experiment, animals were randomly assigned to one of seven groups (n = 6–10/group; n = 115 rats). A vehicle group received infusions of artificial cerebrospinal fluid (aCSF) (which consisted, in millimolars, of: 120 NaCl, 4.8 KCl, 1.2 KH2PO4, 1.2 MgSO4, 25 NaHCO3, 2.5 CaCl2, and 10 D-glucose with pH adjusted to 7.4 ± 0.1) for all seven sessions. The other groups received infusions of 75, 100, 150, 200, 250, or 350 mg% EtOH for the first four sessions. During the fifth and sixth sessions, all animals received infusions of aCSF. During the seventh session, rats were allowed to respond for their originally assigned infusate. As an anatomic control, another group of rats received infusions of 200 or 250 mg% EtOH into the BLA, as these were the concentrations most readily self-administered into the CeA. An anatomic control group had cannula implanted at the BLA. These Wistar male rats were allowed to self-administer 200 or 250 mg% EtOH (n = 9/group).
Effects of Coadministration of the CRF1 Antagonist NBI35965 on EtOH Self-Administration into the CeA.
Funds to maintain the tP line are strictly limited, and the rats are typically only available to conduct adolescent EtOH exposure research. Therefore, the tP rats were not used in subsequent experiments. Wistar male rats were allowed to self-administer 200 mg% EtOH for the first four sessions. During sessions 5 and 6, rats were randomly assigned to self-administer 200 mg% EtOH and 5 nM, 200 nM, 25 μM, or 50 μM concentration (n = 24; 5–7/group) of the CRF1 antagonist, NBI35965 hydrochloride (Bachem). During the seventh session, rats were allowed to respond for 200 mg% EtOH alone. Concentrations of NBI35965 used are within past research examining behaviors in the CeA, but the 25 and 50 μM concentrations are on the high end of typical dose-response curves (Kosoyan et al., 2005).
Effects of Coadministration of the Nociceptin on EtOH Self-Administration into the CeA.
Wistar male rats were allowed to self-administer 200 mg% EtOH for the first four sessions. During sessions 5 and 6, rats were randomly assigned to self-administer 200 mg% EtOH and 100 nM, 500 nM, 1 μM, or 5 μM concentration (n = 41; 9–12/group) of nociceptin trifluoracetate salt (Bachem). During the seventh session, rats were allowed to respond for 200 mg% EtOH alone. Past research has examined a total infusion of 0.5 or 1.0 nM nociceptin into the CeA on behavior (Ciccocioppo et al., 2014). ICSA is the microinjection of 100 nl of a concentration of a solution, and so the concentrations that were used in the current experiment are similar to past research.
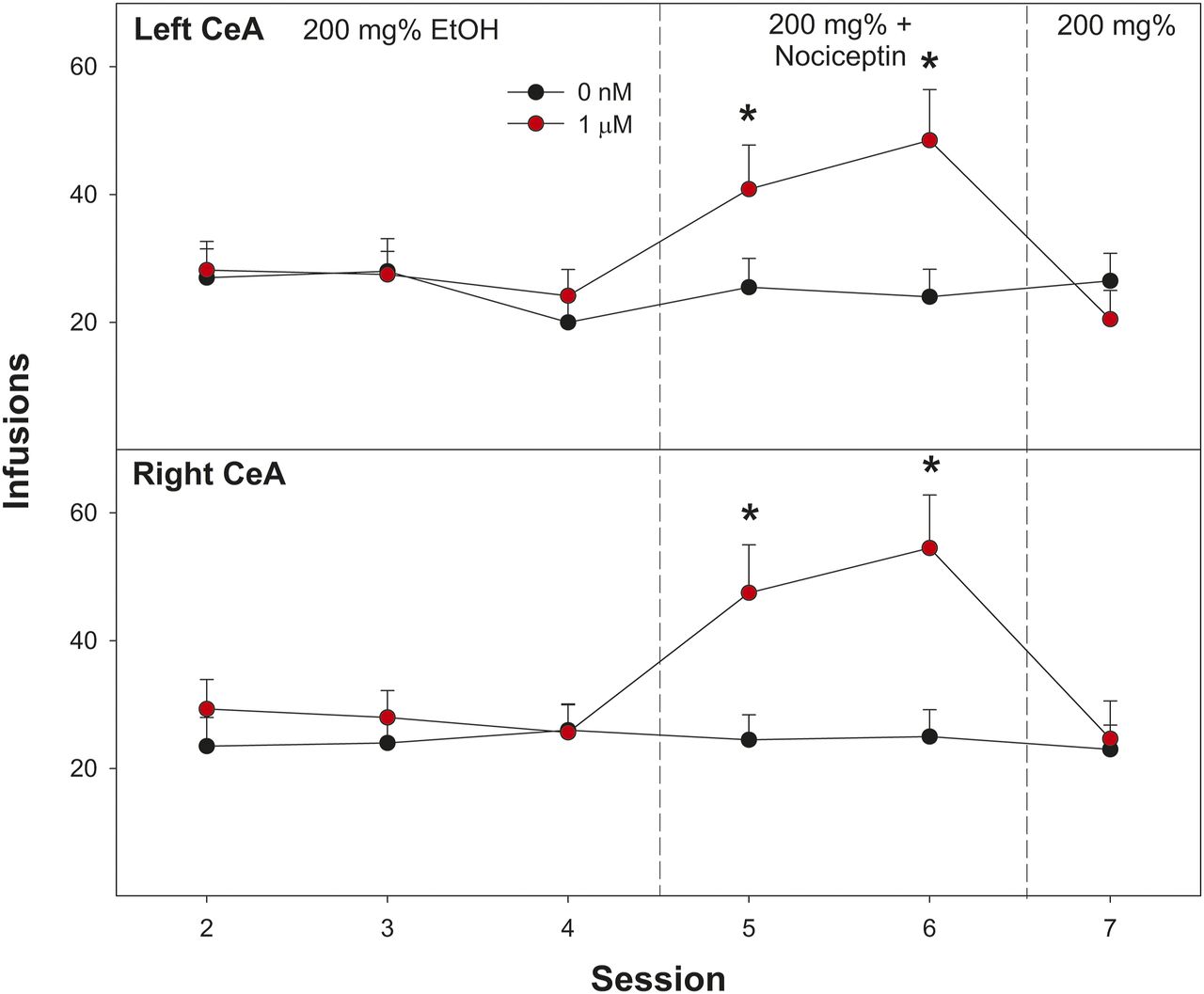
Determining Whether EtOH Self-Administration into the CeA Differs between Right and Light Hemisphere: Re-Examining Coadministration of 1 µM Nociceptin.
There was an abnormal finding for the 1 μM nociceptin group (Fig. 3). All rats were conducted through the ICSA procedure in groups of eight. By random assignment, the 1 μM nociceptin group was conducted last in the following experiment (in conjunction with the 200 nM NBI35965 group), and a different source of nociceptin was used. The studies examining the effects of EtOH microinjected into the CeA on anxiety-like behaviors employed bilateral microinjection procedures, whereas ICSA is a unilateral procedure. Therefore, the current experiment determined whether EtOH would be equivalently self-administered into the right and left CeA and whether coadministration of 1 μM nociceptin (new source) would similarly enhance EtOH self-administration into the CeA. Wistar male rats were implanted with guide cannula aimed at the right or left CeA (n = 8/side). All rats were allowed to self-administer 200 mg% EtOH for the first four sessions, and then rats were randomly assigned to self-administer 200 mg% EtOH (n = 2/side) or 200 mg% EtOH + 1 μM nociceptin (n = 6/side) for sessions 5 and 6. All rats self-administered 200 mg% EtOH during session 7.
Effects of EtOH Microinjections into the Posterior VTA, Nucleus Accumbens Shell, and CeA on Anxiety-Like Behaviors: Social Interaction.
Twenty-four hours before the first social interaction (SI) test, rats were habituated to the SI apparatus by placing the Wistar male rat into the chamber alone for 5 minutes under dim red lighting conditions. The SI test for anxiety-like behavior is unique in that it can be performed within subject as long as the cohort test animal is changed between test sessions. The protocol used for the SI test has been described previously ((Lungwitz et al., 2014); (Shekhar and Katner, 1995). In brief, SI testing consisted of placing the experimental rat into the SI box simultaneously with an age-, weight-, and sex-matched conspecific partner for a 5-minute test session. SI time was measured as the amount of time in seconds that the experimental rat spent engaging in nonaggressive physical investigation of the partner rat, which was defined by the experimental rat sniffing the partner rat (none of the experimental rats displayed aggressive behavior in these studies). Partner-initiated contact or investigation was independent of SI time, thus SI times were independent of the partner’s behavior (none of the partner rats used in these studies displayed avoidant or aggressive behavior). Partner rats were used for a maximum of four sessions in a single day, and these sessions were separated by at least 30 minutes. Each session was video recorded from above and subsequently scored using ODlog for Mac OS X version 2.6.1 by Macropod Software by a treatment-blind observer.
Rats were randomly assigned to receive bilateral EtOH microinfusions into the CeA, VTA, or AcbSh followed immediately by social interaction testing (only one region was tested per rat, and EtOH concentrations were counterbalanced in each region group). The social interaction testing took place over four consecutive days. On the first day, rats received aCSF, 150 mg% EtOH, 200 mg% EtOH, or 250 mg% EtOH bilaterally into the CeA or aCSF, 75 mg% EtOH, 150 mg% EtOH, or 200 mg% EtOH into the VTA or AcbSh. Groups were counterbalanced such that assignment to dose on a given day was randomly assigned. Each rat was exposed to each of the four conditions over the 4-day period specific to the region. Doses were chosen to use concentrations that are readily self-administered into the respective brain regions, and a subthreshold dose for each region that the rats did not readily self-administer was chosen.
Effects of EtOH Microinjections into the CeA on Anxiety-Like Behaviors: Acoustic Startle and Open Field.
A standard in the anxiety field is to confirm alterations in anxiety-like behaviors in multiple behavior protocols (c.f., Donner and Lowry, 2013). The current experiments examined the effects of microinjections of EtOH into the CeA on the expression of anxiety-like behaviors tested by acoustic startle and open field. Detailed methodology of the behavioral assays can be obtained through publication of coinvestigators (Truitt et al., 2007; Johnson et al., 2015). Briefly, Wistar males were administered 0 (aCSF), 150, or 200 mg% EtOH bilaterally into the CeA and then processed through a standard acoustic startle paradigm (n = 22; 7 to 8/group). Separate Wistar males were administered 0 (aCSF), 150, or 200 mg% EtOH bilaterally into the CeA and then processed through a standard open field protocol (n = 23; 7 to 8/group).
Effects of Coadministration of the CRF1 Antagonist NBI35965 on EtOH-Induced Anxiolysis in the CeA (SI).
The current experiment examined the effects of coadministration of NBI35965 on EtOH-induced anxiolysis when microinjected into the CeA. Rats (n = 8) were administered aCSF, 200 mg% EtOH, or 200 mg% EtOH + 5 nM, 200 nM, or 25 µM NBI35965 bilaterally into the CeA immediately prior to SI testing. The experiment was a within-subject design with test conditions counterbalanced.
Effects of Coadministration of Nociceptin on EtOH-Induced Anxiolysis in the CeA (SI).
The EMIT system has been previously used in this laboratory, as well as in other laboratories to identify the neuroanatomical substrates that underlie the reinforcing properties of many drugs of abuse ((McBride et al., 1999). The EMIT system used in the current study is identical to that described previously ((Ding et al., 2009) The EMIT system has been previously used in this laboratory as well as in other laboratories to identify the neuroanatomical substrates that underlie the reinforcing properties of many drugs of abuse (McBride et al., 1999). The EMIT system used in the current study is identical to that described previously (Ding et al., 2009).
The experiment examined the effects of coadministration of nociceptin and EtOH into the CeA on anxiety-like behaviors. Rats received microinjections of aCSF, 200 mg% EtOH, 200 mg% EtOH + 100 nM nociceptin, 500 nM nociceptin, or 1 μM nociceptin (n = 8–10). The experiments were within-subject designs, and test conditions were counterbalanced.
Effects of Administration of Nociceptin in the CeA on the Expression of Anxiety-Like Behaviors (SI).
The experiment examined the effects of administration of nociceptin into the lateral CeA on anxiety-like behaviors. Rats were used in a counterbalanced SI protocol. Rats received microinjections of aCSF, 500 nM nociceptin, 1 μM nociceptin, or 5 μM nociceptin (n = 8).
Histology.
At the termination of all experiments, 1% bromophenol blue (0.5 µl) was injected into the infusion site(s). Subsequently, the animals were given a fatal dose of nembutal and then decapitated. Brains were removed and immediately frozen at –80°C. Frozen brains were equilibrated at –15°C in a cryostat microtome and then sliced into 40-µm sections. Sections were then examined by a trained observer for verification of the injection site using the rat brain atlas of Paxinos and Watson (1998).
Statistical Analysis.
Only rats with cannula placements within the lateral CeA (−2.0 to −2.6 bregma) were included for study. Dependent measures for the ICSA experiments were average number of infusions (first four sessions) and number of active lever responses. Multifactor ANOVAs were performed on each data set. The dependent measures for the assessment of anxiety-like behaviors included time spent engaging in social activity (SI), startle amplitude (N/kg), and time spent in center/center crosses (ppen field). Similar multifactor ANOVAs were performed on the data sets. Post hoc comparisons were always Tukey’s b.
Results
Wistar and tP Male CeA EtOH ICSA Dose Response.
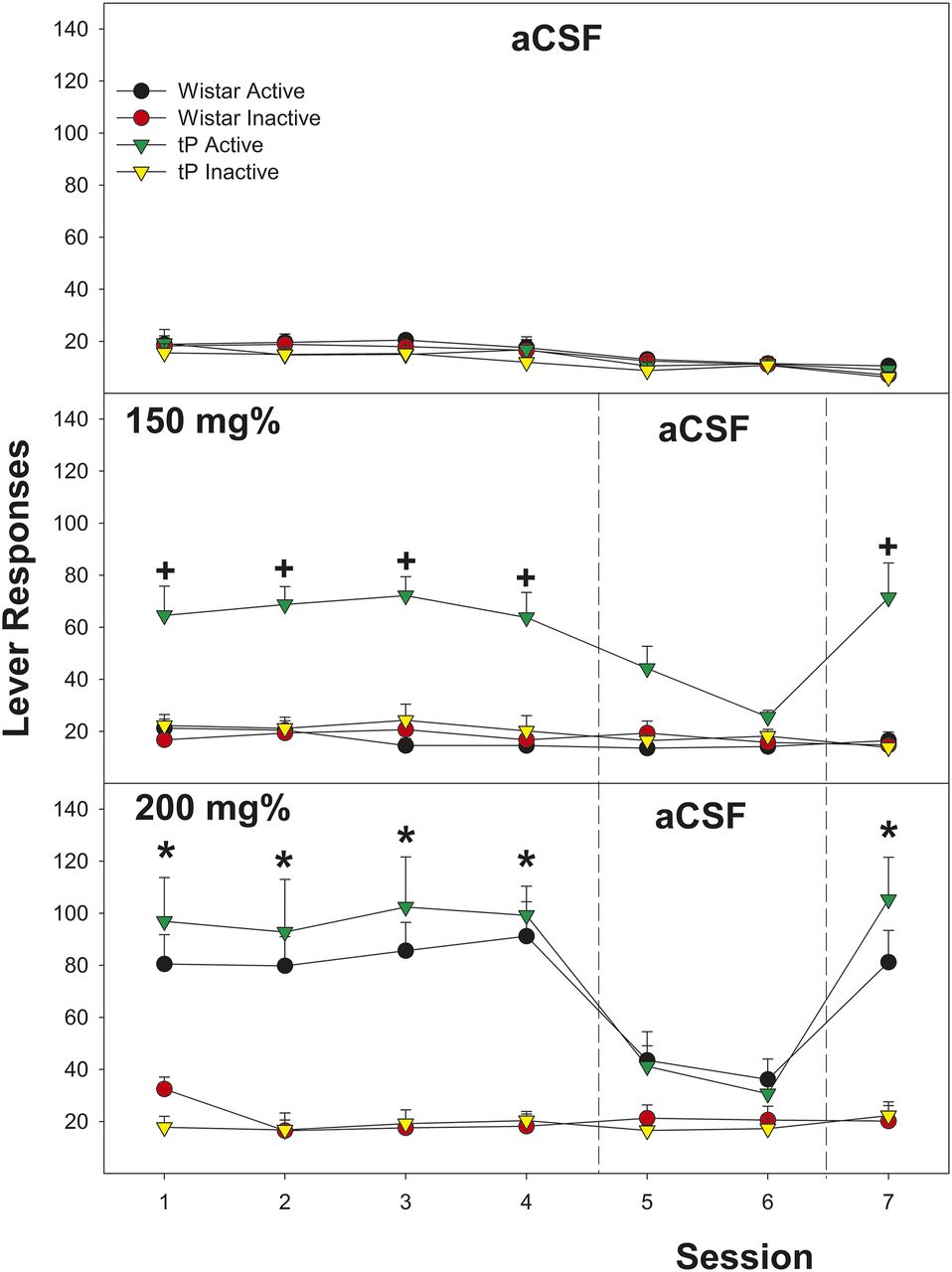
The overall analysis indicated that there was a significant rat line × EtOH concentration (F6,101 = 4.07; P < 0.001) for the dependent variable average number of infusions (first four sessions; Fig. 1). The significant interaction term was reduced by examining average number of infusion for each rat line. In Wistar male rats, there was a significant effect of EtOH Concentration (F6,52 = 29.2; P < 0.001). Post hoc comparisons indicated that male Wistar rats self-administering 200 and 250 mg% EtOH self-infused significantly more than all other groups. In tP male rats, there was also a significant effect of EtOH concentration (F6,46 = 18.8; P < 0.001). Post hoc comparisons indicated that tP rats self-administering 150, 200, and 250 mg% self-infused more than all other groups. Contrasting the average number of infusions between rat lines indicated a significant difference in the 150 mg% concentration (F1,15 = 23.2; P < 0.001).

Depicts the mean (±S.E.M.) average number of infusions self-administered directly into the CeA during sessions 1–4 in Wistar and tP male rats. +Indicates significantly higher self-administration than aCSF and tP males > Wistar males. *Indicates significantly higher self-administration than aCSF controls.
Examining the effects of EtOH concentration and rat line and the repeated measure of session on the number of active lever presses revealed a significant three-way interaction term (Fig. 2; F = 3.75; P < 0.001). In Wistar male rats, there was a significant EtOH concentration × session interaction term (F36,312 = 1.6; P = 0.021). When EtOH was available in the infusate (Sessions 1–4 and 7), there was a significant effect of EtOH dose (F values > 16.04; P values < 0.001). During extinction (Sessions 5 and 6), there was no significant effect of EtOH dose (P values > 0.08). In addition, Wistar rats self-administering 200 and 250 mg% EtOH discriminated between the active and inactive lever (P < 0.0001), whereas all other groups did not show this discrimination (P values > 0.31). Similar results were observed in tP rats, except that significant levels of responding on the active lever were observed in rats self-administering 150, 200, and 250 mg% EtOH. Paralleling the results of average infusion, the significant three-way interaction term was based upon tP rats self-administering 150 mg% EtOH responding more than Wistar rats self-administering the same EtOH concentration (P < 0.001). The effect size (Cohen’s d; {µ1 − µ2}/pooled S.D.) for genetic predisposition to consume alcohol (150 mg% concentration {29.5 − 9.6}/s) indicated a large effect size (0.95).

Depicts the mean (±S.E.M.) number of lever responses in Wistar and tP male rats self-administering aCSF, 150 mg% EtOH, or 200 mg% EtOH. +Indicates significantly higher self-administration than aCSF and tP males > Wistar males. *Indicates significantly higher self-administration than aCSF controls.
EtOH (200 and 250 mg%) was not self-administered into the BLA. The average number of infusions for these two groups was equivalent to that observed during aCSF self-administration (Fig. 1, insert). In addition, there were four Wistar rats with cannulas implanted into the medial CeA. In these rats, self-administration of 150 (1), 200 (2), or 250 (1) mg% EtOH was not observed (average infusion – 9.2 ± 2/session).
Effects of Coadministration of the CRF1 Antagonist NBI35965 on EtOH Self-Administration into the CeA.
Coadministration of NBI35965 did not alter the self-administration of 200 mg% EtOH into the CeA (Fig. 3; top panel). Statistically, there was no effect of NBI concentration on responding for 200 mg% EtOH on the number of infusions (F18,120 = 0.48; P = 0.53).
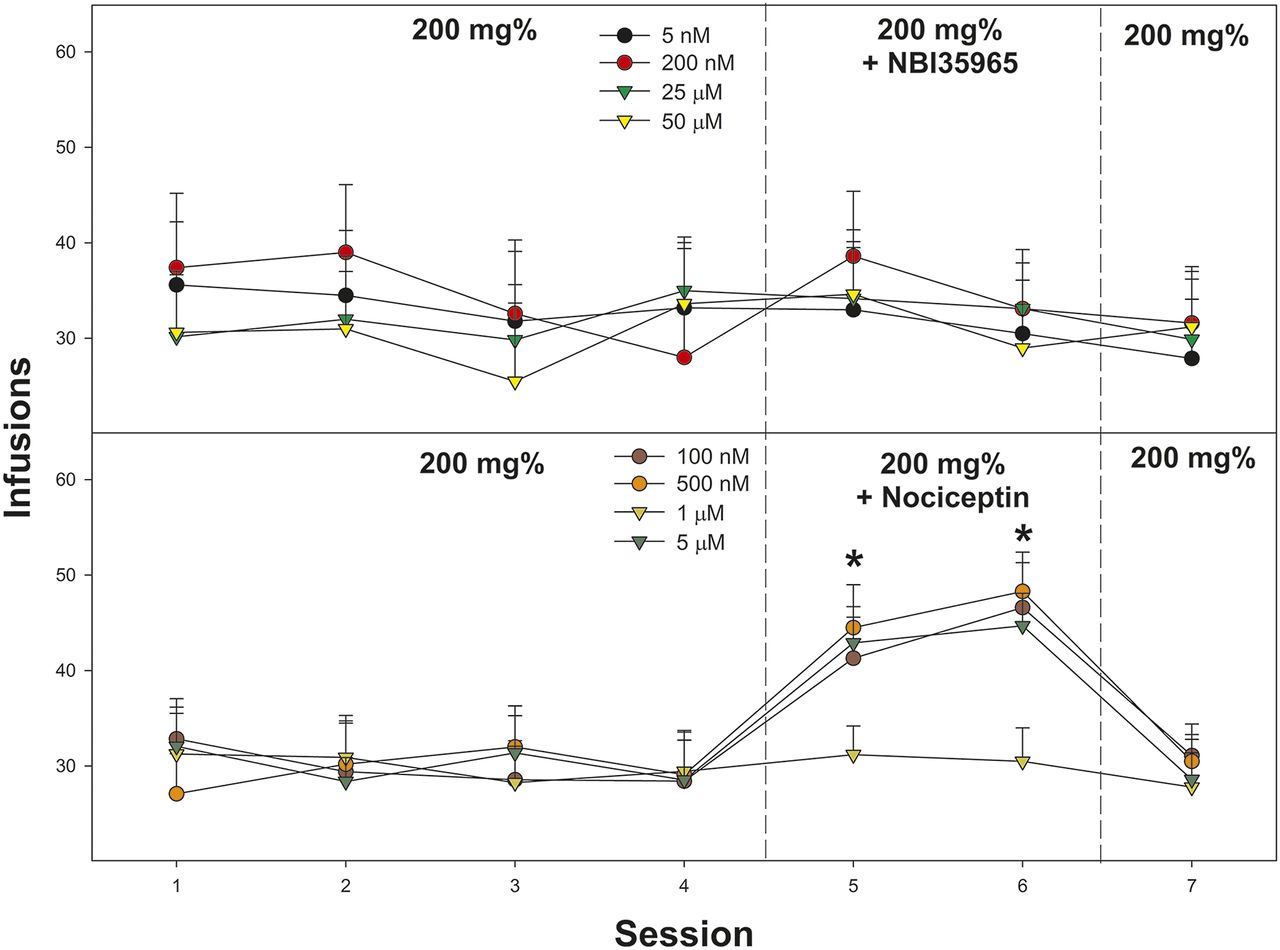
Depicts the mean (±S.E.M.) number of lever responses in Wistar male rats self-administering 200 mg% EtOH and the CRF1 antagonist NBI35965 (top) or nociceptin (bottom). *Indicates that rats coadministering 200 mg% EtOH + 100 nM nociceptin, 500 nM nociceptin, or 5 µM nociceptin self-infused more than when only 200 mg% EtOH alone (sessions 1–4).
Effects of Coadministration of the Nociceptin on EtOH Self-Administration into the CeA.
Coadministration of nociceptin enhanced the self-administration of 200 mg% EtOH into the CeA (Fig. 3; bottom panel). Statistically, there was a significant effect of nociceptin concentration on responding for 200 mg% EtOH on number of infusions (F18,248 = 3.5; P < 0.001). Individual repeated-measure ANOVAs performed on active lever responses indicated that in Wistar male rats, self-administering 200 mg% during sessions 1-4, there was a significant increase in responding during nociceptin coadministration for the 100 nM, 500 nM, and 5 µM groups (sessions 5 and 6; F4,62 values > 11.2; P values < 0.001). Paradoxically, there was no significant effect on number of infusions in rats coadministering 1 µM nociceptin (P = 0.74). The overall power analysis indicated that π = 0.68. The lower π is the result in the variability in the response (lack of an effect of 1 µM nociceptin).
Determining Whether EtOH Self-Administration into the CeA Differs between Right and Light Hemisphere: Re-Examining Coadministration of 1 µM Nociceptin.
Examining the average number of self-infusion of 200 mg% EtOH into the right and left lateral CeA indicated no significant effect of hemispherical location (Fig. 4; F1,14 = 0.57; P = 0.914). To determine the effects of coadministration of 1 µM nociceptin (sessions 5 and 6), the two 200 mg%–alone groups (both right and left CeA) were collapsed to create a single control group (n = 4). A mixed-factor ANOVA was performed with three groups (EtOH alone, EtOH + nociceptin left CeA, EtOH + nociceptin right CeA) and within subject factor of session (4–6). Overall, there was a significant session × group interaction (F4, 124 = 2.7; P < 0.001). Individual ANOVAs performed on individual sessions indicated no group difference during session 4 (P = 0.84) but significant differences during sessions 5 and 6 (F2,14 values > 13.7; P values < 0.001). Post hoc comparisons indicated that Wistar rats self-administering 200 mg% EtOH + 1 µM nociceptin into the right or left CeA self-infused more than rats self-administering only 200 mg% EtOH into right/left CeA (Tukey’s b). The overall power analysis indicated that π = 0.72. The effect size (Cohen’s d) was medium (d = 0.51).

Depicts the mean (±S.E.M.) number of lever responses in Wistar male rats self-administering 200 mg% EtOH into the left (top panel) and right (bottom panel) CeA and 1 µM nociceptin (sessions 5 to 6). *Indicates that rats coadministering 200 mg% EtOH and 1 µM nociceptin self-infused more than rats self-administering 200 mg% EtOH alone.
Overall, it appears that the anomalous finding in the previous experiment was based upon the quality of the nociceptin stock solution. This solution was discarded and not used in any other experiments.
Effects of EtOH Microinjections into the Posterior VTA, Nucleus Accumbens Shell, and CeA on Anxiety-Like Behaviors: Social Interaction.
The amount of baseline social interaction was greater in rats with cannula implanted into the pVTA (unpublished data). This was not the result of the microinjection procedure because during a mock injection procedure, all animals displayed high level of social interaction. To control for the high level of social interaction behaviors in the pVTA rats, we selected to use the data transformation procedure of examining % change from the average aCSF value instead of z-score transformations (statistical analysis of z-scores produced identical results). A repeated-measures ANOVA was run to analyze the effects of EtOH infused bilaterally into the CeA on percent change from the average aCSF infusion social interaction score. Results revealed a significant effect of dose on total interaction time, (F3,18 = 3.680, P < 0.05; Fig. 5). Post hoc analysis revealed a significant difference between all three EtOH doses and the aCSF average. No significant social interaction differences were seen between any of the infused EtOH doses and the aCSF average when infused bilaterally into the pVTA or accumbens shell (P values > 0.64). The overall power analysis indicated that π = 0.87. The effect size (Cohen’s d) for the 150 mg% EtOH concentration to produce anxiolysis between brain regions tested was large (d = 0.82).

Depicts the mean (±S.E.M.) percent change from aCSF controls for social interaction in Wistar male rats microinjected with EtOH into the posterior VTA (top panel), nucleus accumbens shell (middle panel), or the central amygdala (bottom panel). *Indicates significantly higher than aCSF. +Indicates significantly higher than all other comparison groups.
Effects of EtOH Microinjections into the CeA on Anxiety-Like Behaviors: Acoustic Startle and Open Field.
A mixed-factor ANOVA was performed with a between subject factor of EtOH concentration and a repeated measure of audio stimulation. The overall analysis revealed a significant EtOH concentration × audio stimulation interaction term (F12, 115 = 10.16; P < 0.001; Fig. 6; left panel). Individual ANOVAs performed at each db tested revealed significant effects of EtOH concentration during the testing of 100, 105, 110, 115, and 120 db (F2,19 values > 9.93; P values < 0.01). Post hoc comparisons revealed that at the 100, 105, 110, 115, and 120 db test level, rats administered 150 or 200 mg% EtOH displayed less startle reactivity than aCSF controls, and 200 mg% EtOH rats responded less than the 150 mg% group.

Depicts the mean (±S.E.M.) startle amplitude (right panel) and open field–dependent measures (center time and center crosses) in Wistar male rats microinjected with EtOH bilaterally into the CeA. *Indicates that all three EtOH concentrations are significantly different from each other. +Indicates that the 150 and 200 mg% EtOH groups are significantly different from the aCSF control group.
A one-way ANOVA was run to analyze the effects of bilateral EtOH infusion into the CeA on total time spent in the center portion of the open field. There was a significant effect of EtOH concentration (F2,15 = 5.722, P = 0.02; Fig. 5; right panel). Post hoc comparisons indicated that Wistar male rats administered 150 or 200 mg% EtOH into the CeA spent more time in the center portion of the open field than did aCSF controls. A similar analysis performed on the number of center crosses indicated a significant effect of EtOH concentration (F2,15 = 6.686, P = 0.009). Post hoc comparisons indicated that Wistar male rats administered 150 or 200 mg% EtOH into the CeA spent more time in the center crosses than did aCSF controls, and 150 mg% was greater than the 200 mg% group.
The overall power analysis indicated that π = 0.79. The effect size (Cohen’s d) was medium (d = 0.66).
Effects of Coadministration of the CRF1 Antagonist NBI 35965 on EtOH-Induced Anxiolysis in the CeA (SI).
Coadministration of the CRF1 antagonist NBI35965 did not alter EtOH-induced anxiolysis in the CeA. Statistically, there was a significant effect of drug condition on the expression of SI (F3,21 = 23.2; P < 0.001). Post hoc comparisons indicated that rats bilaterally administered 200 mg% EtOH and 200 mg% EtOH + 5 nM NBI35965, 200 nM NBI35965, or 25 µM NBI35965 displayed a comparable increase in SI expression compared with aCSF controls (Fig. 7; left panel). The overall power analysis indicated that π = 0.84. The effect size (Cohen’s d) was medium (d = 0.70).

Depicts the mean (±S.E.M.) social interaction time in Wistar male rats microinjected with 200 mg% EtOH and the CRF1 antagonist NBI35965 (right panel) or nociceptin (left panel) directly into the CeA. *Indicates significantly higher than aCSF controls. +Indicates significantly higher than aCSF controls and 200 mg% EtOH group.
Effects of Coadministration of Nociceptin on EtOH-Induced Anxiolysis in the CeA (SI).
Statistically, there was a significant effect of drug condition on the expression of SI (F4,39 = 17.7; P < 0.001). Post hoc comparisons indicated that rats bilaterally administered 200 mg% EtOH expressed more SI behaviors than aCSF controls, and rats administered 200 mg% EtOH + 100 nM nociceptin, 500 nM nociceptin, or 1 μM nociceptin expressed significantly more SI behaviors than aCSF controls and rats administered only 200 mg% EtOH (Fig. 7; right panel). The overall power analysis indicated that π = 0.85. The effect size (Cohen’s d) was medium (d = 0.51).
Effects of Administration of Nociceptin in the CeA on the Expression of Anxiety-Like Behaviors (SI).
There was no significant effect of nociceptin concentration on the expression of SI (P = 0.82). Specifically, SI behavior after aCSF (18.3 ± 8.3 seconds) was not different compared with 500 nM (15.6 ± 6.4 seconds), 1 μM (16.7 ± 5.5 seconds), or 5 μM (20.6 ± 7.7 seconds) nociceptin (unpublished data).
Discussion
The data indicate that EtOH has both anxiolytic and rewarding properties within the lateral portion of the CeA (Figs. 1⇑⇑⇑⇑⇑–7). Previous reports have indicated that selective breeding for high alcohol consumption is associated with alterations within the CeA (Pandey et al., 2005; Herman et al., 2013, 2016). The current data indicate that there is a leftward shift in the dose-response curve for the tP rats compared with Wistar (the foundation stock of P rats) subjects (Figs. 1 and 2). Coadministration/infusion of a CRF1 antagonist had no observed effect on EtOH self-administration or EtOH-induced anxiolysis in the CeA (Figs. 3 and 7). In contrast, concurrent exposure to nociceptin and EtOH enhanced EtOH self-administration and EtOH-induced anxiolysis in the CeA (Figs. 3, 4, and 7).
EtOH is self-administered directly into the posterior VTA (Hauser et al., 2019) and AcbSh (Engleman et al., 2009). The effects of EtOH in the CeA are unique because equivalent microinjections of EtOH into the CeA induced anxiolysis, which was not observed after EtOH microinjections into the posterior VTA and AcbSh (Fig. 5). In the anxiety field, assessing the actions of compounds in multiple animal models of anxiety-like behavior is required to assert validity of any results (e.g., Donner and Lowry, 2013). The current data provide convergent evidence that EtOH microinjected into the CeA is anxiolytic in the standard animal models of anxiety assessment (Figs. 5 and 6).
The current findings indicate that the CRF1 receptor does not mediate EtOH reward or anxiolysis in the CeA. Despite decades of speculation about CRF agents being potential therapeutics for the treatment of alcohol use disorder (AUD) and addictions to other drugs of abuse (e.g., Markou et al., 1998), repeated clinical studies have failed to observe any benefits of CRF antagonists (c.f., Shaham and de Wit, 2016). For example, the CRF1 antagonist Verucerfont failed to alter alcohol craving and other measures of alcohol intake in treatment-seeking women considered to have anxious AUD (Schwandt et al., 2016). Increases in CRF in the CeA are associated with increases in the expression of numerous anxiety- and fear-related behaviors (c.f., Holsboer and Ising, 2008). Therefore, the increase in CRF produced by EtOH exposure in the CeA (Roberto et al., 2010) would suggest an anxiogenic action. This is counterintuitive to the known anxiolytic effect of EtOH in humans and the observed anxiolytic action of EtOH microinjected into the lateral CeA (Figs. 5 and 6).
The medial and lateral CeA are heterogeneous for the expression of neuropeptides. Neurons expressing the CRF1 receptor are abundant in the medial CeA, and there are significantly fewer CRF1 receptors containing neurons in the lateral CeA (Justice et al., 2008; Herman et al., 2013). Recent data have indicated that neurons in the medial CeA that express the CRF1 receptor are GABAergic with specific segregation properties, coexpression of other neuropeptides, and dendritic spine characteristics, but there is less CRF1 receptor expression in the lateral CeA (Wolfe et al., 2019). Therefore, the failure to observe the effects of coadministration of the CRF1 antagonist on EtOH reinforcement and anxiolysis in the lateral CeA could be based upon the limited level of CRF1 receptors in that region.
There are subpopulations of neurons within the lateral and medial CeA for somatostatin, neurotensin, tachykinin 2, neuropeptide Y, encephalin, dynorphin, cholecystokinin, and other neuropeptides (Chieng et al., 2006; McCollough et al., 2018). A recent light-sheet microscopy experiment (IDISCO+) was used to identify prepronociceptin-expressing neurons within the CeA (Hardaway et al., 2019). The findings indicated a regional heterogeneity for the expression of prepronociceptin-expressing neurons with the highest concentration of nociceptin occurring in the Anterior-Posterior zone used for ICSA/anxiolysis testing in the current experiments (Hardaway et al., 2019). Prepronociceptin-expressing neurons in the lateral CeA had extremely weak coexpression with CRH and other neuropeptides (Hardaway et al., 2019). Optogenetic stimulation of prepronociceptin-expressing neurons in the lateral CeA produced reward-like behaviors but did not alter food consumption (Hardaway et al., 2019).
The biologic basis for the ability of the neuropeptide nociceptin to enhance both EtOH reward and anxiolysis in the CeA is unknown. Nociceptin inhibits GABA release in the CeA (Kallupi et al., 2014a,b). Nociceptin can also directly inhibit presynaptic CRF receptors within the CeA (Ciccocioppo et al., 2014). In the lateral CeA, EtOH increases glutamate levels (Silberman and Winder, 2013; Silberman et al., 2015). CRF1 receptors act presynaptically to reduce the release of glutamate in the CeA (Varodayan et al., 2017). Nociceptin microinjections into the CeA do not alter anxiety-liked behavior in the elevated plus-maze (Ciccocioppo et al., 2014) or in social interaction testing (unpublished data). Therefore, the ability of nociceptin to enhance EtOH reward and anxiolysis in the CeA could be predicated on several factors that need to be elucidated in future research.
The neurochemical response (microdialysis) to systemic EtOH in the CeA has not been extensively studied. The sole study examining systemically administered EtOH on neurochemical levels in the CeA indicated that EtOH increased both dopamine and serotonin levels (Yoshimoto et al., 2000). Reversed microdialysis perfusion of EtOH into the medial CeA (estimated from stereotaxic coordinates) has been shown to increase GABA release (Roberto et al., 2004a, 2010). To date, there has been no reported assessment of glutamate release in the lateral/medial CeA after systemic or local administration of EtOH using microdialysis or voltammetry.
Brain regions’ response to EtOH and other drugs of abuse can be classified as displaying regional heterogeneity. The VTA and nucleus accumbens both show regional heterogeneity for EtOH self-administration (Rodd-Henricks et al., 2000; Engleman et al., 2009). Winder and colleagues (2013; 2015) have postulated that the divergent observations of the effects of EtOH in the CeA could be based upon subregion specific effects (Silberman and Winder, 2013; Silberman et al., 2015). It is possible that the actions of EtOH in the lateral CeA are opposite to those observed by well-established researchers in other areas (medial) of the CeA (Roberto et al., 2010). In our hands, EtOH was reinforcing/anxiolytic in the lateral CeA, but we did not see EtOH self-administration in the BLA or in the limited number of subjects with cannula placement in the medial CeA (Fig. 1 insert). Multiple detailed regional analyses of EtOH effects in the CeA are needed to advance the alcohol research field.
A common misconception is that EtOH reinforcement and anxiolysis are two separate processes in the lateral CeA. To put it bluntly, anxiolysis can be reinforcing. Preclinically, anxiolytic agents/procedures can condition a place preference and alter brain stimulation parameters (Gerhardt et al., 1982) or establish the development of goal-directed behaviors to generate an anxiolytic state (Yadin et al., 1993). In humans, a large portion of patients with AUD report the continuation of alcohol consumption for the rewarding “relief” (anxiolysis) obtained (Grodin et al., 2019). Therefore, it may be impossible to separate the reinforcing/anxiolytic properties of EtOH within the lateral CeA because they are the same thing.
The CeA has been postulated as a neuronal nexus that mediates response to stress, negative affect processes, fear processes, and drug-related behaviors (Koob and Volkow, 2010). The effects of EtOH within the CeA appear to be complex under drug naïve conditions. The current results indicate that EtOH is both anxiolytic and reinforcing in the lateral CeA. The current tentative hypothesis is that the biologic basis for these effects of EtOH in the CeA is through activation/release of the glutamate, which may act at α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid and N-methyl-d-aspartate receptors to regulate behaviors (c.f., Gilpin et al., 2015). Chronic EtOH exposure can produce neuroadaptations in the CeA. Rats made EtOH-dependent have 4-fold higher basal extracellular GABA levels, alterations in GABAA receptor subunits, and alterations in GABAB function in the CeA, as compared with controls (Morrow et al., 1992; Roberto et al., 2004a, 2008). Therefore, an already complex neurologic circuit mediating the actions of EtOH could be further expanded by a history of alcohol use. Future studies need to determine the EtOH reward and anxiolysis in the CeA in dependent rats.
Acknowledgments
None of the authors of this manuscript have conflicts of interest to report.
Authorship Contributions
Participated in research design: Knight, Hauser, McBride, Rodd
Conducted experiments: Knight, Hauser, Waeiss, Molosh, Johnson, Truitt, Shekhar, Rodd
Contributed new reagents or analytic tools: Bell
Performed data analysis: Waeiss, McBride, Bell
Wrote or contributed to the writing of the manuscript: Knight, Hauser, Molosh, Johnson, Truitt, McBride, Shekhar, Rodd
Footnotes
- Received August 16, 2019.
- Accepted April 15, 2020.
This research was supported by National Institutes of Health National Institute on Alcohol Abuse and Alcoholism [Grants AA07611, AA022287, AA07462, and AA020908].
Abbreviations
- AcbSh
- nucleus accumbens shell
- aCSF
- artificial cerebrospinal fluid
- AUD
- alcohol use disorder
- BLA
- basolateral amygdala
- CeA
- central nucleus of the amygdala
- CRF
- corticotropin-releasing factor
- EMIT
- electrolytic microinfusion transducer
- EtOH
- ethanol
- ICSA
- intracranial self-administration
- P
- alcohol-preferring
- pVTA
- posterior VTA
- SI
- social interaction
- tP
- Taconic P
- VTA
- ventral tegmental area
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}