


Abstract
Cinnamaldehyde (Cin), a bioactive cinnamon essential oil from traditional Chinese medicine herb Cinnamomum cassia, has been reported to have multipharmacological activities including anti-inflammation. However, its role and molecular mechanism of anti-inflammatory activity in musculoskeletal tissues remains unclear. Here, we first investigated the effects and molecular mechanisms of Cin in human synoviocyte cells. Then in vivo therapeutic effect of Cin on collagen-induced arthritis (CIA) also studied. Cell Counting Kit CCK-8 assay was performed to evaluate the cell cytotoxicity. Proinflammatory cytokine expression was evaluated using quantitative polymerase chain reaction and ELISA. Protein expression was measured by western blotting. The in vivo effect of Cin (75 mg/kg per day) was evaluated in rats with CIA by gavage administration. Disease progression was assessed by clinical scoring, radiographic, and histologic examinations. Cin significantly inhibited interleukin (IL)-1β–induced IL-6, IL-8, and tumor necrosis factor-α release from human synoviocyte cells. The molecular analysis revealed that Cin impaired IL-6–induced activation of Janus kinase 2 (JAK2), signal transducer and activator of transcription 1 (STAT1), and STAT3 signaling pathway by inhibiting the phosphorylation of JAK2, STAT1, and STAT3, without affecting NF-κB pathway. Cin reduced collagen-induced swollen paw volume of arthritic rats. The anti-inflammation effects of Cin were associated with decreased severity of arthritis, joint swelling, and reduced bone erosion and destruction. Furthermore, serum IL-6 level was decreased when Cin administered therapeutically to CIA rats. Cin suppresses IL-1β–induced inflammation in synoviocytes through the JAK/STAT pathway and alleviated collagen-induced arthritis in rats. These data indicated that Cin might be a potential traditional Chinese medicine–derived, disease-modifying, antirheumatic herbal drug.
SIGNIFICANCE STATEMENT In this study, we found that cinnamaldehyde (Cin) suppressed proinflammatory cytokines secretion in rheumatology arthritis synoviocyte cells by Janus kinase/signal transducer and activator of transcription pathway. The in vivo results showed that Cin ameliorated collagen-induced arthritis in rats. These findings indicate that Cin is a potential traditional Chinese medicine–derived, disease-modifying, antirheumatic herbal drug.
Introduction
Inflammation accounts for a wide range of progressive musculoskeletal diseases (Gallo et al., 2017), including osteoarthritis (Geyer and Schonfeld, 2018), rheumatology arthritis (RA) (Jia et al., 2015), and cervical spondylopathy (Kang et al., 1995). It is a complex process regulated by an array of inflammatory mediators and cytokines (Gibon et al., 2017). Cinnamomum cassia Presl is frequently used in traditional Chinese medicine to treat gastritis, blood circulation disturbances, and inflammatory disease (Gunawardena et al., 2015), such as Jing Shu Ke Li for treatment of cervical spondylopathy.
Cinnamaldehyde (Cin, 3-phenyl-2-propenal), as a major active constituent of Cinnamomum cassia, is widely used as flavoring agents in foodstuffs (Kim et al., 2007), with many pharmacological activities, including anti-inflammation (Huang et al., 2015), suppression of adipocyte differentiation (Huang et al., 2011), and osteoclastgenesis (Tsuji-Naito, 2008). Cin has been known to have anti-inflammatory activity in a variety of cells, such as human renal tubular cells (Huang et al., 2015), monocyte/macrophage (Chao et al., 2008), and gastrointestinal cells (Li et al., 2016). Furthermore, Cin exhibits inhibitory effects on proinflammatory cytokines in LPS-stimulated macrophages in vitro by suppressing intracellular signaling (Chao et al., 2008; Kim et al., 2010). Liao et al. (2012) found that Cin strongly suppress nitric oxide, tumor necrosis factor (TNF-α), and prostaglandin E2 productions in LPS-stimulated macrophages. Intraperitoneal injection of Cin with 5 mg/kg dose to carrageenan-induced mouse paw edema model improved the paw edema by suppressing proinflammatory cytokine expression and promoting the activities of catalase, superoxide dismutase, and glutathione peroxidase in the paw tissue (Liao et al., 2012). Further in vivo studies showed that Cin inhibits the activation of NF-κB in aged rat kidney tissues (Kim et al., 2007) and attenuates the advanced glycation end products-induced inflammation via JAK/STAT signaling in human renal tubular cells (Huang et al., 2015). In addition, Cin affects the biologic behavior of human colorectal cancer cells and induces apoptosis via inhibition of the phosphoinositide 3-kinase/Akt signaling pathway (Li et al., 2016). Overall, these findings depicted Cin as a potential therapeutic agent in inflammatory diseases. However, the possible anti-inflammatory and protective mechanisms of Cin on musculoskeletal cells are not fully understood.
Fibroblast-like synoviocytes (FLS) are located in the knee joint and responsible for most of the musculoskeletal diseases (Rahmati et al., 2016) by releasing large amounts of proinflammatory mediators, including interleukin (IL)-6, IL-8, and TNF-α (Sluzalska et al., 2017). IL-1β is one of the key cytokines stimulating FLS to produce IL-6 and IL-8; thus, many researchers use IL-1β–induced fibroblast-like synoviocytes for inflammation experiments (Sluzalska et al., 2017).
In this study, we focused on investigating the effects of Cin on IL-1β–induced inflammatory cytokine production in human MH7A cells derived from RA-FLS (Jia et al., 2015) and joint destruction in collagen-induced arthritis (CIA) animal model and its underlying mechanisms. Our data showed that Cin inhibited IL-1β–induced inflammatory cytokine production in MH7A cells mainly via the suppression of JAK/STAT pathways. We also demonstrated that Cin attenuated the severity of arthritis in rats with CIA. These results indicate that Cin may be a potential traditional Chinese medicine–derived, disease-modifying, antirheumatic drug.
Materials and Methods
Cell Culture.
Human rheumatoid fibroblast-like synoviocyte line MH7A were provided by Riken cell bank (Tsukuba, Japan). Cells were incubated in RPMI-1640 medium (Hyclone) with 10% FBS (Gibco, GrandIsland) and penicillin/streptomycin (1:100; Hyclone) at 37°C and 5% CO2. Primary synovial cells of patients with rheumatoid arthritis were from Peking University Shenzhen Hospital. The cells were incubated in Dulbecco’s modified eagle’s medium (Hyclone) with 10% FBS (Gibco) and penicillin/streptomycin (1:100; Hyclone) at 37°C and 5% CO2. This study was approved by the Medical Ethical Committee of Peking University Shenzhen Hospital and was performed according to the recommendations of the Declaration of Helsinki.
Cell Viability Assay (CCK-8 Assay).
MH7A cells were seeded into 96-well culture plates at 5 × 103 cells/well; after 24-hour incubation, cells were treated with or without cinnamaldehyde (Cin, the compound provided by Jing Fang [Anhui] pharmaceutical Limited Company, SinoPharm Group, the purity is 97.96%) at the concentrations of 20, 40, 60, 80, and 100 nM for 24 hours. Then, CCK-8 (Dojindo, Japan) was added and incubated at 37°C for 2 hours. The absorbance of CCK-8 was detected at 450 nm by a microplate reader (PerkinElmer).
Quantitative Real-Time Polymerase Chain Reaction.
MH7A cells were pretreated or untreated with various concentrations of Cin for 2 hours and then incubated for another 6 hours with or without 20 ng/ml of IL-1β (Peprotech). Total RNAs were isolated using the commercial total RNA miniprep kit (Axygen), according to the manufacturer’s instructions. Each sample was reverse transcribed using the cDNA synthesis kit (TaKaRa, China) according to the manufacturer’s protocol. The primer sequences were used for real-time PCR as displayed in Table 1. Real-time PCR analysis was performed using SYBR Green PCR Premix Ex Taq II reagents (TaKaRa) on a Light Cycler 480 II real-time system (Roche). The GAPDH was served as house-keeping gene for normalization.
Primer sequences for real-time PCR
ELISA.
MH7A cells and primary synovial cells were seeded at 5 × 106 cells/well into six-well plates; after 24 hours, MH7A cells were pretreated with various concentrations of Cin or not for 2 hours and then stimulated with or without IL-1β (20 ng/ml) for another 24 hours. Another group was stimulated with IL-1β (20 ng/ml) for 2 hours then treated by Cin for 24 hours. The culture medium was collected, and the concentrations of key cytokines were determined by ELISA with a commercial kit (eBioscience Technology Shenzhen, China), according to the manufacturer’s instructions.
Western Blot.
MH7A cells were pretreated with Cin, Jak 2 inhibitor (Santa Cruz), or were not pretreated for 2 hours, then stimulated with IL-1β (20 ng/ml) for 1 hour, and then cellular proteins were assayed by western blot analysis. Total proteins in the cell were lysed by lysis buffer (radio-immunoprecipitation assay; Thermo), and protein concentration was measured by BCA kit (Thermo). 50 μg of protein from each sample was separated by 10% SDS-PAGE and transferred to a polyvinylidene fluoride membrane (Millipore Co.). After blocking with 5% bovine serum albumin in Tris-buffered saline/Tween20, the membrane was incubated with specific antibody overnight at 4°C, followed by incubation with the secondary antibody. The band density was quantified by Image Laboratory (Bio-Rad). Primary antibodies for phospho-JAK2 (Tyr1007/1008), phospho-STAT1 (Tyr701), phospho-STAT3 (Tyr705), phospho-Akt (Thr308), phospho-ERK1/2 (Thr202/Tyr204), JAK2, STAT1, STAT3, Akt, ERK1/2, and GAPDH were purchased from Cell Signaling Technology (Beverly), and the antibodies for phospho-NF-κB-p65 (Ser468) and NF-κB-p65 were purchased from Signalway Antibody (College Park, Maryland). The corresponding densitometry analysis was performed using Image Laboratory (Bio-Rad) by calculating the average optical density in each field. The ratio of phosphorylated protein to total protein was qualified for comparison between different groups.
Determination of the Effect of Cin on CIA.
CIA was induced in Lewis female rats (8 weeks old) by bovine type II collagen (Chondrex, Redmond, WA) in acetic acid (2 mg/ml), which was emulsified in an equal volume of Incomplete Freund’s Adjuvant. Inject 0.2 ml (200 mg collagen/rat) of the emulsion subcutaneously at the base of the tail. A booster injection has been given on day 7 after the initial immunization with 0.1 ml of the emulsion subcutaneously in the tail (Brand et al., 2007). When swollen joints appeared at day 14 after the first injection, rats (six per each group) were given gavage administration of either Cin (75 mg/kg per day, CIA-Cin) (Weng et al., 2019), methotrexate (MTX, 0.5 mg/kg per 3 days, CIA-MTX) as positive drug or sodium carboxymethylcellulose (CMC-Na, CIA-CMC-Na) as negative control for 21 days. Another six rats were used as the normal control group. The animals were housed under special pathogen-free conditions at the animal facility. The animal experiments were approved by the Animal Ethics Committee of Chinese Academy of Sciences Shenzhen Institute of Advanced Technology (SIAT-IRB-150303-YGS-CWX-A0114).
Paw Volume Assessment.
The hind paw volume was measured every week from the first immunization. Place a beaker containing a 10% soap solution of a known density on a top-loading balance and tare the balance to zero. Light anesthesia may facilitate this process. Immerse the limb in the solution to the level of the anatomic hair line. After retraction of the paw, the container is weighed, subtracted from the start weight and corrected for fluid density.
Micro-Computed Tomography Assessment of Arthritis.
The right paws and ankle joints were scanned and reconstructed into a three-dimensional structure by the SKYSCAN 1176 micro-CT (Bruker, Belgium). The settings recommended by the manufacturer with exposure of 60 kV and 400 μA was used. Bone volume (BV) and bone surfaces (BS) of the tarsal bones were then computed to evaluate the structural changes. The region of interest (ROI) included the region between the metatarsal and tarsal bones. To evaluate the surface topology of the periarticular bone (indicating focal erosion on the bone surfaces) the ratio BS:BV was used (Antill et al., 2016; Zheng et al., 2018).
Measurement of Serum Cytokine Production in Rats.
Rat serum was obtained by centrifugation of blood in serum separator Microtainer tubes after cardiac puncture. The levels of cytokines and chemokines were measured by multiplex immunoassay using MILLIPLEX MAP Rat Cytokine/Chemokine magnetic panel as per manufacturer’s instructions (Millipore). We analyzed protein levels of IL-1β. Species cross reactivity was evaluated by the manufacturer (http://www.abacus-als.com/media/Milliplex_2014.pdf).
Histopathology Assessment.
After 21 days of treatment, the hind limbs were removed and fixed with 10% formalin, decalcified for 6 weeks in 10% EDTA, dehydrated and embedded in paraffin. The sections were stained with hematoxylin and eosin (H&E) for light microscopy (Chow et al., 2018). The sections were scored for changes in synovial proliferation, inflammation, cartilage damage, and bone erosion using the following scoring system: 0, normal; 1, mild; 3, moderate; and 4, severe (Hayer et al., 2016).
Statistical Analysis.
Data were presented as mean ± S.D. The in vitro data were from at least three independent experiments. The sample size was six per group for in vivo evaluation. Statistical analysis was performed using GraphPad Prism 7.0 (GraphPad Software Inc., San Diego, CA). The significance of differences between groups was determined using Student’s unpaired t test. One-way ANOVA followed by Fisher’s post hoc was used to evaluate differences between experimental groups. A value of P < 0.05 was considered to be statistically significant.
Results
Cin Effects on Cell Viability.
To test whether Cin (Fig. 1A) exerted anti-inflammation effects below a cytotoxic threshold, we performed CCK-8 assay to detect the influence of Cin on MH7A cell growth. MH7A cells were cultured in various concentrations ranking from 0 to 100 nM of Cin for 24 hours (Fig. 1B). No cytotoxic effects were found below the 100 nM, a dose that caused an 8.74% reduction in viability (P < 0.01). We therefore selected 40, 60, and 80 nM dosages for Cin in subsequent experiments testing its potential therapeutic utility against RA.

Effect of Cin on cell viability of MH7A cells by CCK-8. (A) The chemical structure of Cin. (B) CCK-8 results showed that Cin did not have toxic effects at concentrations lower than 100 nM. **P < 0.01 compared with the control group (Ctrl).
Cin Treatment Causes IL-6, IL-8, and TNF-α Level Decreased in IL-1β-Stimulated MH7A Cells and Primary Synovial Cells.
Using the in vitro IL-1β–stimulated MH7A cell and primary synovial cells of inflammation, we next examined whether Cin influenced the accumulation of proinflammatory cytokines IL-6, IL-8, and TNF-α. MH7A cells were pretreated with Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml). After 6 hours, the mRNA levels of IL-6, IL-8, and TNF-α were measured by quantitative PCR. The results showed that IL-1β significantly upregulated IL-6, IL-8, and TNF-α gene expressions to 181.8, 1167, and 10.62 folds, respectively (P < 0.001 for all). Cin at 40, 60, and 80 nM dose-dependently and significantly inhibited the gene expression of IL-6 to 58.08%, 46.87%, and 31.91%, respectively (P < 0.001 for all), whereas Cin at 60 and 80 nM dose-dependently and significantly inhibited the gene expression of IL-8 to 78.28% and 66.3%, respectively (P < 0.001 for both). Furthermore, Cin at 40, 60, and 80 nM dose-dependently and significantly inhibited the gene expression of TNF-α to 76.38%, 64.14%, and 53.38%, respectively (P < 0.001 for all, Fig. 2A).
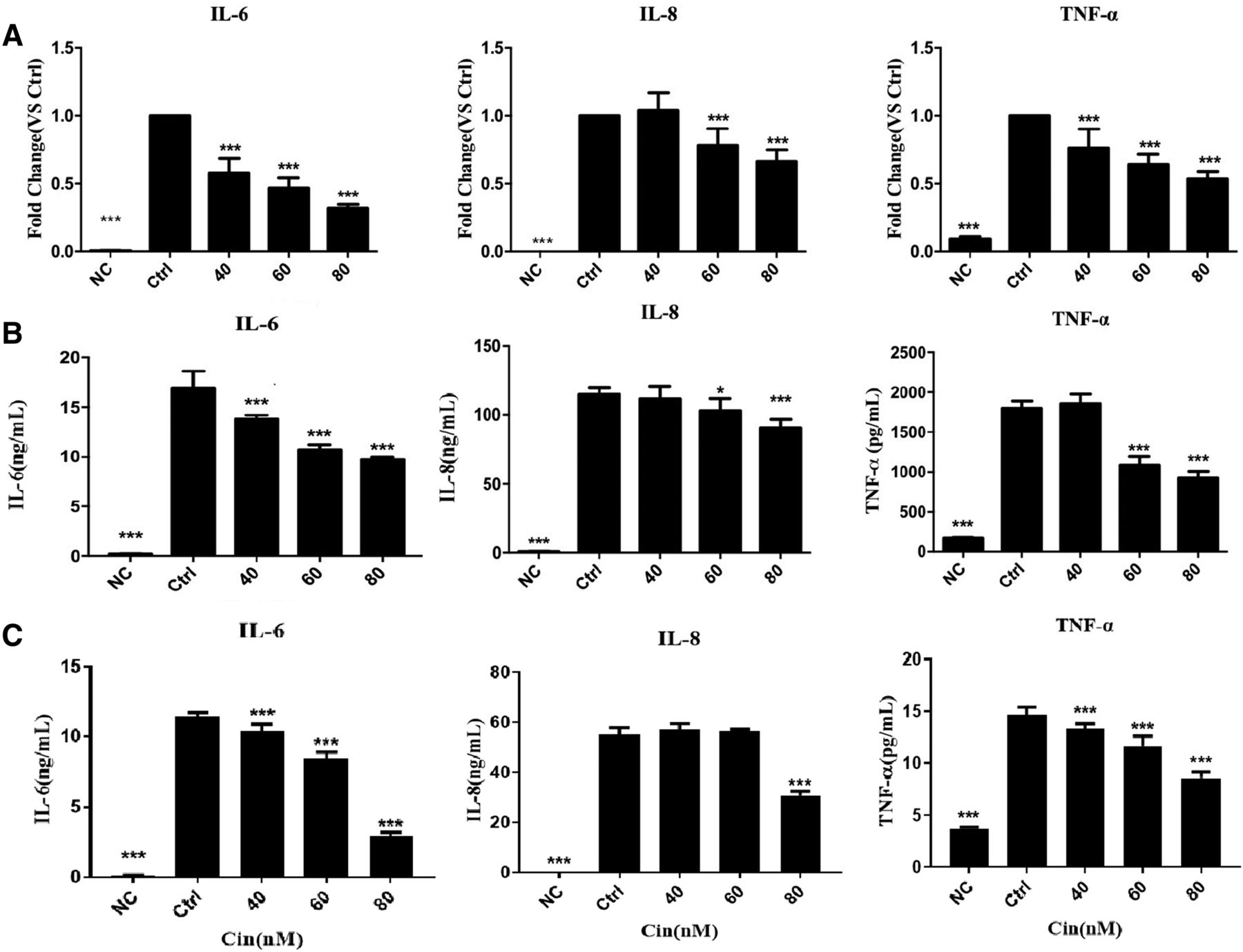
Anti-inflammation effects of Cin in MH7A. Real-time PCR (A) and ELISA. The result demonstrated that Cin dose-dependently inhibited the production of IL-6, IL-8, and INF-α in IL-1β–stimulated MH7A cells. (B) Group for pretreated with Cin first, then stimulated by IL-1β. (C) Group for stimulated by IL-1β first, then treated with Cin. *P < 0.05; ***P < 0.001 compared with the control group (Ctrl, IL-1β treated alone).
After the Cin treatment, whether stimulated by IL-1β previously and after, the protein levels of IL-6, IL-8, and TNF-α were measured in the culture medium by ELISA in MH7A (Fig. 2, B and C). IL-1β significantly stimulated the releases of IL-6, IL-8, and TNF-α protein to 88.58, 180.4, and 10.3 folds, respectively (P < 0.001 for all). Cin at 40, 60, and 80 nM dose-dependently and significantly inhibited the release of IL-6 to 81.42%, 63.2%, and 57.41%, respectively (P < 0.001 for all). Cin at 40 and 80 nM significantly inhibited the release of IL-8 to 89.4% (P < 0.05) and 78.93% (P < 0.001). Cin at 60 and 80 nM dose-dependently and significantly inhibited the release of TNF-α to 60.19% and 51.22%, respectively (P < 0.001 for both). Cin also suppressed the level of proinflammatory factors in primary synovial cells, which is consistent with the results of MH7A cell lines, significantly inhibited the release of IL-6 and IL-8, and the expression of TNF-α was too low to be detected (Supplemental Fig. 1).
Cin Suppresses JAK/STAT Signaling in IL-1β–Induced MH7A Cells.
The JAK/STAT cascade is involved in the regulation of numerous developmental and homeostatic processes, including inflammatory responses (Ahmad et al., 2015). To further characterize the role of JAK/STAT signaling in anti-inflammation of Cin treatment, we detected the activation degrees of signal protein. MH7A cells were pretreated with Cin or not for 2 hours, and then stimulated with IL-1β (20 ng/ml) for 15, 30, or 60 minutes, JAK/STAT pathway was determined by western blot analysis.
Cin decreased the ratio of p-JAK2:JAK2 in a dose-dependent manner (Fig. 3A1), and 80 nM of Cin significantly decreased the ratio to 52.3% compared with the IL-1β stimulated only group (P < 0.01) (Fig. 3A2). The ratio of p-STAT1:STAT1 and p-STAT3:STAT3 increased by 2.5- and 2.0-fold at 60 minutes after IL-1β stimulation, respectively (Fig. 3B), and Cin at 80 nM dramatically decreased the ratio of p-STAT1:STAT1 and p-STAT3:STAT3 to 42.3% and 49.5% compared with the IL-1β stimulated only group (P < 0.01 for both) (Fig. 3B). Compared the effects of 80 nM of Cin and 20 μM of Jak 2 inhibitor, at 60 minutes after IL-1β stimulation, 80 nM of Cin could decrease the expression of p-JAK2, p-STAT1, and p-STAT3, whereas Jak 2 inhibitor decreased the expression of p-STAT1 and p-STAT3 but not p-JAK2 (Fig. 3C).

Representative western blot images and quantitative results (ratio of phosphorylated protein to total protein or GAPDH) to demonstrate the effect of Cin on JAK2, STAT1, and STAT3 activation in IL-1β–induced MH7A cells. (A) MH7A cells were pretreated with 40, 60, and 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 60 minutes. (B) MH7A cells were pretreated with 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 15, 30, and 60 minutes. (C) MH7A cells were pretreated with 80 nM of Cin, 20 μM of Jak 2 inhibitor or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 60 minutes. *P < 0.05; **P < 0.01, the comparison was performed between IL-1β–induced and drug treatment group and IL-1β–induced only group at the same time point. P < 0.05; ##P < 0.01, the comparison was performed between the IL-1β–induced group at different time points with the group without IL-1β induction. &&P < 0.01, the comparison was performed between the uninduced groups with or without Jak2 inhibitor treatment.
Cin Promotes Phosphorylation of Akt but Slightly Decreases Phosphorylation of NF-κB in IL-1β–Induced MH7A Cells.
Inhibition of NF-κB transcriptional activity is an important way for anti-inflammation in osteoarthritis and RA. Akt and ERK signaling pathways could be the main upstream of NF-κB signaling cascades (Dan et al., 2008; Sun et al., 2017). We observed that the NF-κB transcriptional activity was affected by Cin in MH7A cells. MH7A cells were pretreated with or without Cin for 2 hours, followed by stimulation with IL-1β (20 ng/ml) for 15, 30, or 60 minutes. Cin at 60 and 80 nM significantly increased the ratio of p-AKT:AKT compared with the IL-1β stimulated only group (P < 0.01 for both) (Fig. 4A). The ratio of p-AKT:AKT was the highest at 15 minutes after IL-1β stimulation, and Cin at 80 nM significantly increased the ratio of p-AKT:AKT at each time point (Fig. 4B). The total protein of ERK1/2 decreased with IL-1β stimulation, and Cin had no effect on the ratio of p-ERK1:2:ERK1/2 (Fig. 4). The ratio of p-NF-κB-p65:NF-κB-p65 did not change obviously with IL-1β stimulation, but 80 nM of Cin could slightly decrease the ratio of p-NF-κB-p65:NF-κB-p65 (P > 0.05).

Representative western blot images and quantitative results (ratio of phosphorylated protein to total protein) demonstrated the effect of Cin on AKT, ERK, and NF-κB activation in IL-1β–induced MH7A cells. (A) MH7A cells were pretreated with 40, 60, and 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 60 minutes. (B) MH7A cells were pretreated with 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 15, 30, and 60 minutes.P < 0.05; **P < 0.01, the comparison was performed between IL-1β–induced and Cin treatment group and IL-1β–induced only group at the same time point.Representative western blot images and quantitative results (ratio of phosphorylated protein to total protein) demonstrated the effect of Cin on AKT, ERK, and NF-κB activation in IL-1β–induced MH7A cells. (A1) MH7A cells were pretreated with 40, 60, and 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 60 minutes. (A2) MH7A cells were pretreated with 80 nM of Cin or not for 2 hours and then stimulated with IL-1β (20 ng/ml) for 15, 30, and 60 minutes. (B1 & B2) Corresponding densitometric analysis of A1 & A2. *P< .05 compared with the control group.The *P < 0.05; **P < 0.01, the comparison was performed between IL-1β–induced and Cin treatment group and IL-1β–induced only group at the same time point.
Cin Administration Attenuates the Arthritis Severity in CIA Rats.
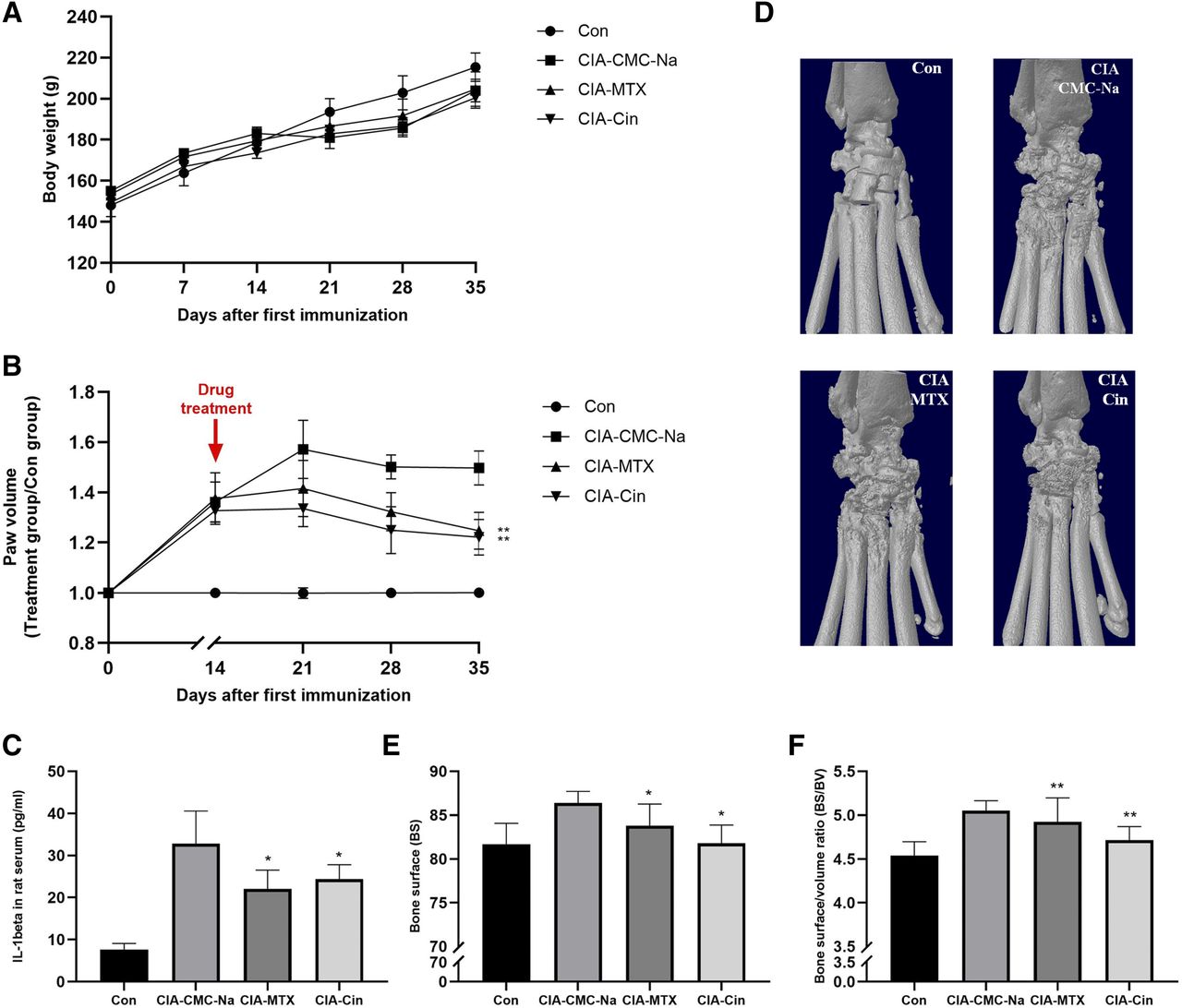
We next evaluated the in vivo effects of Cin on the synovial inflammation of CIA rats. Compared with the negative control group, a slight body weight loss was noted in the CIA model rats (P > 0.05) (Fig. 5A). As shown in Fig. 5, from day 14 onwards, there was a continuous increase of paws and ankle volume. Administration with Cin caused a significant reduction in paw volume (30%) (Fig. 5B). In agreement with the inhibition of paw volume, treatment with Cin had a marked inhibitory effect on the systemic expression of proinflamatory cytokine IL-1β (Fig. 5C). However, the expression of IL-6 and TNFα was too low to be detected in serum (unpublished data).

Cin attenuated the symptoms of CIA rats and joint destruction. (A) The body weight. (B) Hind paw volume was recorded every 7 days after drug treatment. (C) The serum IL-1β levels decreased in the CIA-treated rats with Cin or MTX group. (D) On day 21 after being drug treated, rats were sacrificed. 3D reconstructions of the micro-CT analysis from CIA rats using the CTAn software. The bone radiologic destruction parameters BS (E) and BS:BV ratio (F) were attributed from the micro-CT analysis. The ROI included the region between the metatarsal and tarsal bones. N = 6. *P < 0.05; **P < 0.01, the comparison was performed between treatment and vehicle control group.
The effects of all treatments on bone erosion in the periarticular bone was analyzed via micro-CT evaluation of the periarticular bone in ankle joints and the distal end of left tibiae with a quantitative histomorphometric imaging method. Micro-CT assessment of the ankle joints of the CIA control group revealed that bone erosions usually involved all small tarsal bones. Micro-CT three-dimensional images showed that the articular surfaces of tarsal bones were very irregular, and the talocalcaneal joint space was frequently obliterated (Fig. 5D). Qualitative data showed that in the Cin group, the surface area of the tarsal bone decreased and BS:BV decreased significantly (P < 0.05). The results in the MTX group were similar to that in the Cin group (Fig. 5, E and F).
We also conducted histologic examinations to evaluate the joint pathology in rats. H&E staining showed that a large number of inflammatory cells infiltrated and bone destruction in localized areas of the CIA control group (Fig. 6A; Supplemental Fig. 2). As expected, Cin-treated rats showed a significant reduction of pathologic disease severity as supported by lower synovial proliferation (Fig. 6B), inflammation (Fig. 6C), cartilage damage (Fig. 6D), and bone surface erosion (Fig. 6E) than those in the CIA group. Similarly, MTX also alleviated the histopathological arthritic damage in the CIA joints (Fig. 6, B–E). These data suggested that Cin could alleviate inflammation and prevent from bone destruction.

The histopathological severity was assessed and calculated histologic findings. (A) The specimens with removed arthritic paws were stained with H&E (original magnification, 100×). The histopathological severity in terms of synovial proliferation (B), inflammation (C), cartilage damage (D), and bone surface erosion (E) were assessed and calculated. The ROI included the region between the metatarsal and tarsal bones. N = 6. *P < 0.05; P < 0.01, the comparison was performed between treatment and vehicle control group.
Discussion
In the present study, the anti-inflammatory effect of Cin were explored in a rat model of collagen-induced arthritis and in IL-1β treated human rheumatoid fibroblast-like synoviocytes line MH7A, respectively. Cin exhibited the inhibitory potential against the proinflammatory cytokines via the suppression of JAK/STAT signaling pathway in in vitro assay. Moreover, systemic administration of Cin exerted an antiarthritic effect as evidenced by a decrease in paw swelling and significant reduction in histologic changes in the articular joints of rats. We also demonstrated that systemic induction of Cin therapy significantly reduced the serum levels of proinflammatory cytokine IL-1β.
Cytokine concentrations correlated with clinical symptoms, inflammatory indicators, disease activities, and serum biomarkers (Yuan et al., 2018). Thus, the proinflammatory productions of TNF-α, IL-6, and IL-8 were measured after Cin treatment to assess the anti-inflammatory outcome. The releases of cytokines, especially TNF-α, IL-1β, IL-6, and IL-8, were critically involved in the inflammation process (Swärd et al., 2012; Cassuto et al., 2018). IL-1β possesses a broad spectrum of proinflammatory properties and is usually used for including inflammation in vitro (Castejónet al., 2017; Feng et al., 2017). It could induce the synthesis of proinflammatory cytokines (such as TNF-α and IL-6), chemokines (such as IL-8), and activated macrophages (Castejónet al., 2017; Feng et al., 2017). Previous studies showed that Cin (80 μM) inhibited the secretion of IL-1β and TNF-α within LPS or lipoteichoic acid stimulated murine J774A.1 macrophages (Chao et al., 2008). In our study, Cin at the concentrations of 60 and 80 nM significantly suppressed IL-1β–activated inflammatory cytokine production levels in human synoviocyte cell line MH7A. It also inhibited IL-6 and IL-8 cytokines levels in primary synovial cells. The TNFα was not detectable in this primary cell, which may be due to the different cell resources as well as the cells state and the limited primary sample size (Guo et al., 2016). Consistent with the cell ELISA results, Cin led to a decrease in serum cytokine levels of IL-1β in CIA rats, suggesting that Cin might provide beneficial effects by specific down-regulating the synthesis of IL-1β.
The JAK/STAT cascade has been shown to be involved in the regulation of numerous developmental and homeostatic processes, including inflammatory responses (Ahmad et al., 2015). IL-1β binding to their receptors can induce the phosphorylation of receptor-associated JAK in chondrocytes, which in turn transduces the intracellular signal by further phosphorylating and activating STATs (Lim and Kim, 2011). The STATs are activated by phosphorylation at the tyrosine residue, and then dimerized, translocated to the nucleus and bind DNA, initiating the transcription of target genes (Adach et al., 2009; Qi and Yang, 2014).
Available literature reported that Cin attenuated advanced glycation end products-induced the JAK/STAT signaling in human renal tubular cells (Huang et al., 2015). We observed that, with the IL-1β stimulation, JAK2 was promptly phosphorylated while p-STAT1 and p-STAT3 phosphorylated. The results indicated that in the IL-1β–induced inflammatory cascade, JAK2, is at the upstream. Both Cin and Jak 2 inhibitor decreased the expression of p-STAT1 and p-STAT3 phosphorylation in human synoviocyte MH7A cells, whereas Cin also suppressed JAK2 phosphorylation, which hinted that Cin was effective on the upstream of JAK2. However, the direct target of Cin has not been found yet. In summary, Cin may become a new small-molecule inhibitor of JAK/STAT signaling to attenuate the inflammatory response.
The transcription factor NF-κB plays a crucial role in the inflammation processes of many tissues (Ray et al., 1995; Ju Hwang et al., 2019). Inhibition of NF-κB transcriptional activity is an important way for anti-inflammation in osteoarthritis and RA (Park et al., 2016; Jiang et al., 2017). The NF-κB protein dimer is complexed with the inhibitory proteins, IκBs (IκBα, IκBβ) in the inactivated state and is sequestered in the cytoplasm (Hong, 2017). After the stimulation of proinflammation factors, such as LPS, TNF-α, or IL-1β, the inhibitory proteins IκBα is rapidly phosphorylated by IKKα/β, and ubiquitinated, rapidly degraded by the proteasome, allowing NF-κB to release from IκB and translocate to the nucleus and initiate transcription by binding to numerous specific gene promoter elements to stimulate inflammation (Gallo et al., 2017). In our study, although these proinflammatory factors were all significantly increased after IL-1β stimulated, phosphorylation of an important transcription factor in the inflammation processes, NF-κB, was not changed obviously after IL-1β stimulation. Compared with our previous work (Jia et al., 2015), it is found that the cells used in this study were in an inflammation state originally. However, the decrease of these cytokines with statistical analysis suggests that Cin is effective for anti-inflammation in human synoviocytes-MH7A cells.
Recently, numerous studies have shown that the Akt and ERK signaling pathways play essential roles in the pathogenesis of inflammation by phosphorylation of IκB (Yang et al., 2016; Wang et al., 2017). It is reported that hemolysate increases the phosphorylation of Akt and ERK1/2 promotes the degradation of IκBα and subsequently increases the nuclear translocation of NF-κB (Yang et al., 2016). Thus, Akt and ERK signaling pathways could be the main upstream of NF-κB signaling cascades (Dan et al., 2008; Sun et al., 2017). Our results showed that Cin could increase the phosphorylation of Akt at Thr308 but decrease the phosphorylation of NF-κB, which explained the slight effect of Cin on suppression of NF-κB phosphorylation and hinted that there were other regulators at the upstream of NF-κB in IL-1β–induced human synoviocyte MH7A cells.
The most important goal in RA therapy is to maintain normal joint function via prevention of bone destruction (Li et al., 2017; Tanaka, 2019). In the present study, we investigated Cin could attenuate the severity of CIA, in addition to suppressing inflammatory disease activity and preventing bone destruction. Blocking of IL-1 is a cartilage- and bone-protective therapy in destructive arthritis. Activated RA-FLSs are the major source of proinflammatory cytokines and inflammatory mediators in the synovial tissue. Therefore, the inhibition of inflammatory cytokines can significantly reduce RA-FLSs cartilage invasiveness (Joosten et al., 1999; Ganesan and Rasool, 2017). Our results showed that Cin markedly attenuated IL-1β level and arthritis symptom in CIA rats. Meanwhile, bone erosion in the RA control group was markedly more extensive than that in the Cin treatment group. Combined with the results of in vitro study, Cin might prevent bone destruction via inhibited fibroblast synovial cells producing IL-1β.
Our findings indicated that Cin significantly inhibited release of the proinflammatory cytokine both the rats with CIA and human synoviocyte MH7A cell, partly modulating through the JAK/STAT pathway. Importantly, Cin markedly ameliorated arthritis in rats and conferred joint protection when used therapeutically. Taken together, these findings suggest that Cin may be a potential therapeutic agent for chronic inflammatory disorders such as RA.
Acknowledgments
We thank Dr. Qing-Wen Wang (Peking University Shenzhen Hospital) for providing primary synovial cells from the rheumatoid arthritis patients.
Authorship Contributions
Participated in research design: Qin, X.-L. Wang.
Conducted experiments: Cheng, Zhong, Meng, Zheng.
Performed data analysis: Cheng, Zhong, Meng, Zhang, Y. Wang, X.-L. Wang.
Wrote or contributed to the writing of the manuscript: Cheng, Zhong, Qin, X.-L. Wang.
Footnotes
- Received September 25, 2019.
- Accepted February 3, 2020.
↵1 W.-X.C. and S.Z. contributed equally to this paper.
This work was supported by Sino-Swiss collaborative project from Ministry of Science and Technology and the Swiss National Science Foundation (2015DFG32200 and 156362), National Nature Science Foundation of China (81773964), and Shenzhen Science and Technology Research Funding (JCYJ20180302150101316 and 20170502171625936).
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- Akt
- RAC-alpha serine/threonine-protein kinase
- BS
- bone surfaces
- BV
- bone volume
- CIA
- collagen-induced arthritis
- Cin
- cinnamaldehyde CMC, Carboxymethyl cellulose
- CT
- computed tomography
- ERK, extracellular signal-regulated kinases; FLS
- fibroblast-like synoviocytes
- GAPDH
- glyceraldehyde-3-phosphate dehydrogenase
- H&E
- hematoxylin and eosin
- IL
- interleukin
- JAK
- Janus kinase
- LPS
- lipopolysaccharide
- MTX
- methotrexate
- NF-κB
- nuclear factor κ-light-chain-enhancer of activated B cells
- PCR
- polymerase chain reaction
- RA
- rheumatology arthritis
- ROI
- region of interest
- STAT
- signal transducer and activator of transcription
- TNF-α
- tumor necrosis factor α
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}