


Abstract
Timely nerve restoration is an important factor for the successful regeneration of tissues and organs. It is known that axon regeneration following nerve injury is a multifactorial process that depends on the local expression of neurotrophins, including brain-derived neurotrophic factor (BDNF). Along with the survival of neurons, the active reorganization of the extracellular matrix is an important step for the growth of axons to their targets. Urokinase serine protease is part of the plasminogen activator system, which provides the vectoriality of the process of fibrinolysis and matrix reorganization, facilitating the growth of nerves to their targets. Based on this and in view of the results of our previous studies, we suggest that a combined bicistronic plasmid encoding the complementary proteins BDNF and urokinase may be beneficial in nerve regeneration. The ability of this bicistronic plasmid to stimulate nerve restoration was confirmed by in vitro stimulation of Neuro2a neurite growth and in vivo nerve conductivity and histology studies. To our knowledge, this is the first article that demonstrates the effectiveness of a bicistronic plasmid containing the human genes BDNF and urokinase plasminogen activator in the regeneration of the injured peripheral nerve. The results obtained demonstrate that plasmid vectors encoding several complementary-active therapeutic proteins may serve as a basis for developing prospective treatments for a wide range of multicomponent neural system disorders, such as nerve trauma.
SIGNIFICANCE STATEMENT This study is the first to show the effectiveness of using a bicistronic plasmid encoding complementary-active human protein brain-derived neurotrophic factor and urokinase plasminogen activator in the regeneration of the crushed peripheral nerve in a murine model.
Introduction
Blood vessels and nerve fibers are essential components of any tissue and provide for the normal development, functioning, and regeneration of tissues. However, tissue injuries sometimes lead to damage to blood vessels and nerve fibers. Following injury, both the regenerating blood vessels and nerve fibers begin to grow from the remaining stump. Unlike the regeneration of blood vessels, the regeneration of nerve fibers has rather restricted potential, which is caused by remote neuron cell bodies being localized predominantly within the central nervous system and the short-time profile of neurotrophic factor production (Boyd and Gordon, 2003). Contemporary research on tissue regeneration describes it as a complex multistep process that requires the participation of a variety of cellular and signaling components, which interchange and/or cooperate with each other during the regeneration progress (Li et al., 2016; Wang et al., 2017; Nimiritsky et al., 2019). This may explain the relatively moderate effects of monogenic gene therapy in clinical studies despite the promising results that were obtained in preclinical studies (Gupta et al., 2009; Makarevich et al., 2015). Despite advances in medicine, there are no effective therapeutic approaches to or treatments for nerve injury, and persistent disability is reported in 75% of all cases following severe nerve injury (Odinak and Zhivolupov, 2009; Shevelev, 2005).
The modern concept of successful regeneration highlights the necessity of multimodal and complex interventions and stimulation of the regenerating tissue (Li et al., 2016; Wang et al., 2017; Nimiritsky et al., 2019). With this in mind, the activation of several signaling cascades that drive different modalities and complement each other seems feasible. Among the therapeutic compounds that complement each other during injured nerve regeneration, neurotrophic factors and proteases are of particular interest. In this combination, a neurotrophic factor would be used to stimulate the survival and growth of nerve fibers, and a protease would be used to promote neurite growth cone migration via cleavage of the endoneurial channels from inhibitory molecules and fibrin deposits as well as to activate endogenous growth factors by proteolysis (Benga et al., 2017; Karagyaur et al., 2017).
It is known that the expression of neurotrophins in an injured nerve is not sustained for longer than a month. Short-term neurotrophin expression and a lack of integrity and passableness of the nerve trunk are the main challenges for successful nerve regeneration (Jia et al., 2014; Gordon T. and English, 2016) because of increased injured neuron death and a lack of neurite outgrowth. In contrast, prolonged neurotrophin secretion (e.g., via neurotrophin gene delivery) and adequate neurite channel clearance helps the regenerating neurites to reach their target organs. Brain-derived neurotrophic factor (BDNF) can be used as a stimulator for motor neuron regeneration according to previous research (Carlone et al., 1987; Karagyaur et al., 2015). A wide range of proteins are suitable complements to BDNF. However, proteases, including plasminogen activators that stimulate fibrinolysis and angiogenesis, take part in neurite channel clearance, and activate BDNF, are among the most attractive. Among these, urokinase plasminogen activator (uPA) particularly stands out. It was recently shown that uPA is necessary for the directed growth of axons via binding to its receptor (uPAR) on the surface of growth cones (Semina et al., 2016; Merino et al., 2017).
In our previous work, we showed that single-gene plasmids encoding hBDNF or muPA stimulated the recovery of an injured nerve (Karagyaur et al., 2015). Alongside existing research, this finding confirms the important role of BDNF and uPA in the process of neuron survival and neurite regeneration. Taking into account the complexity and multiple stages of the regenerative process, we surmised that a bicistronic genetic construct encoding both hBDNF and huPA could be beneficial for enhancing nerve regeneration. We decided to create the desired bicistronic construct in the form of a plasmid because plasmids are still the most safe and convenient genetic instruments for heterologous protein expression in medicine and basic science. Here, we show that a bicistronic plasmid construction encoding growth factors of complementary modalities (BDNF and uPA) stimulates nerve fiber restoration after nerve crush in a clinically relevant model of nerve injury.
Materials and Methods
Plasmids
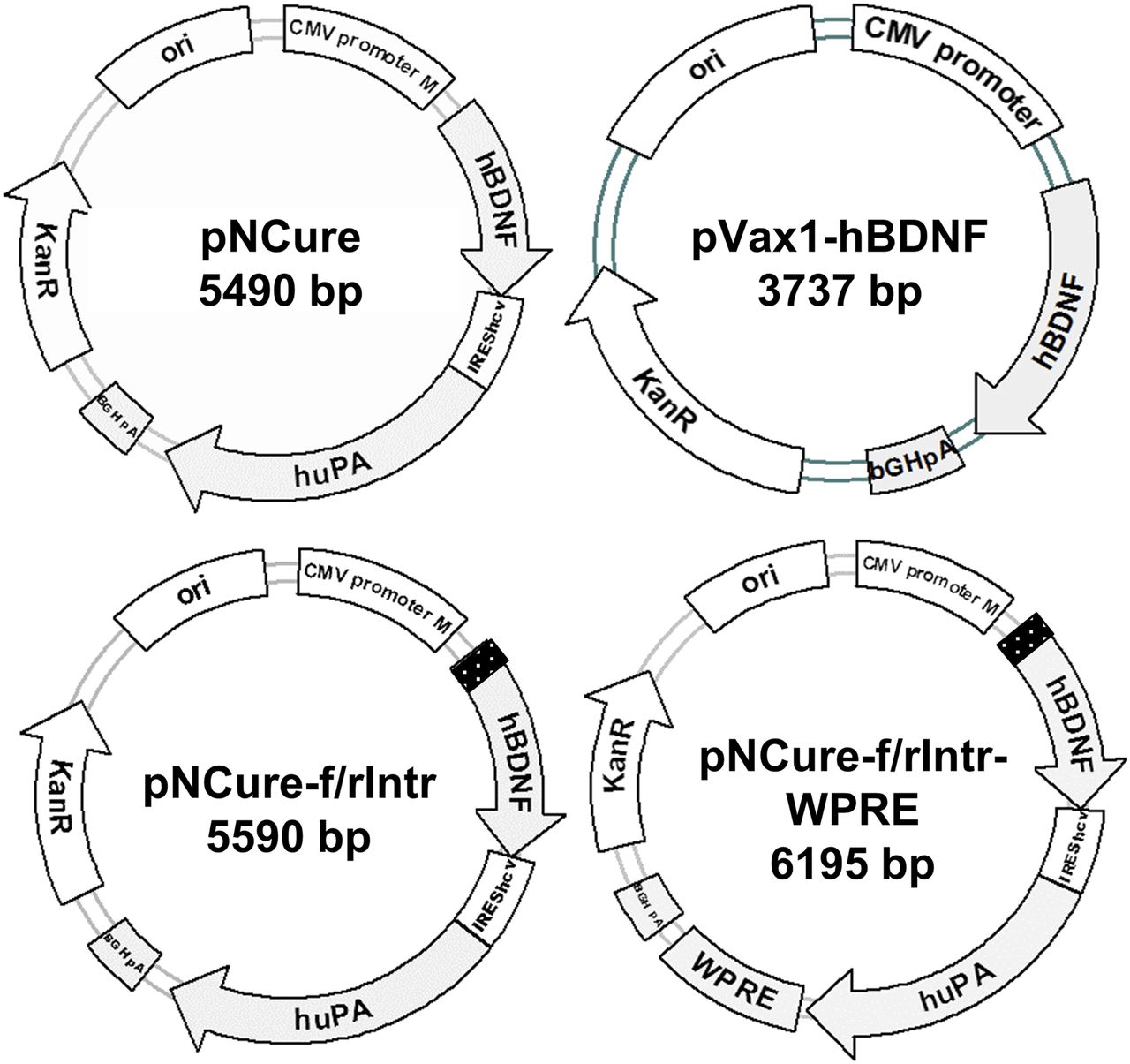
To stimulate nerve regeneration, a bicistronic vector encoding hBDNF and huPA cDNAs was created. As a backbone, a pVax1m plasmid was used, which was obtained by introducing an SV40 (Simian virus 40) enhancer sequence between a CMV promoter and a CMV enhancer in the pVax1 vector (V26020; Thermo Fisher Scientific) to increase plasmid uptake by the cell nucleus (Dean et al., 2005). hBDNF and huPA cDNAs were obtained by reverse-transcription polymerase chain reaction of total RNA extracted from human adipose–derived mesenchymal stromal cells, which were obtained from the biomaterial collection within the frame of the Moscow University Project “Noah’s Ark” (primer sequences are listed in Supplemental Table 1). hBDNF cDNA was introduced into the pVax1m vector subsequent to the CMV promoter and the Kozak consensus sequence (GCCACC) using NheI (New England Biolabs, #R0131S) and HindIII (New England Biolabs, #R0104S) restriction sites (Kozak, 1984). huPA cDNA (cloned using Acc65I (New England Biolabs, #R0599S) and EagI (New England Biolabs, #R0505S) restriction sites) was separated from hBDNF by HCV IRES (Lukavsky, 2009), assembled from synthetic oligonucleotides (polymerase chain reaction–amplified and cloned using HindIII and Acc65I restriction sites). According to the study by Jubin et al. (2000), HCV IRES provides for the production of the second protein, huPA, at a 10-fold lower molar rate than the first protein, hBDNF. Each cDNA carries its own start and stop codons (the start codon for huPA is built in the HCV IRES). The plasmid pVax1m-hBDNF-IRES-huPA was named “a plasmid for nerve cure” (pNCure). To increase the productivity of the plasmid, four additional variants of the plasmid were constructed with a synthetic intron and WPRE sequence (Fig. 1). The synthetic intron was designed de novo, assembled from synthetic oligonucleotides, and introduced between the CMV promoter and Kozak consensus sequence (NheI restriction site). Intron’s sequence is 5′-GTGAGTAACAGAGATCTAATACGCTTCTCTTGCTAAGCCTAACCTTTTCTGTCTTGTTTCTGCTTTCCCCCCCCCACAG-3′ (bold letters mark intron key motifs). It was not obvious which intron orientation (forward vs. reverse) would be advantageous in protein production, so both plasmid variants were stored (pNCure-f/rIntr). The WPRE sequence was cloned from pAAV-EF1a-tdTomato-WPRE-pGHpA (a gift from Botond Roska, plasmid 67527; Addgene) (Wertz et al., 2015) and introduced into pNCure-f/rIntr variants between the huPA stop codon and bovine growth hormone polyadenylation signal (XbaI (New England Biolabs, #R0145S) restriction site). The obtained plasmid variants were tested in parallel with the previously described pVax1-hBDNF plasmid (Karagyaur et al., 2015).

Used plasmid variants encoding hBDNF and huPA.
Treatment
Animals.
The study was performed on mature male F1 CBA/C57Bl mice aged 8–10 weeks, with standard weight characteristics (20–23 g), obtained from SPF vivarium of Faculty of Medicine (Lomonosov Moscow State University). Animal housing and research procedures were conducted in compliance with Directive 201/63/EU.
Intramuscular Plasmid Injection.
In our previous work on the intramuscular delivery of plasmids, we used a quick injection of a plasmid solution at a single muscle point followed by electroporation. However, for therapeutic use, multiple points for plasmid injection are used (muscle infiltration) with no electroporation, as this causes massive necrosis of muscle tissue, of which approximately 10%–30% depends on electroporation voltage (Manthorpe et al., 1993; Karagyaur et al., 2015). Therefore, here, we used a more clinically relevant plasmid delivery protocol by substituting the single-injection electroporation regimen with multiple intramuscular injections and no electroporation. To test the efficacy of using multiple injections with no electroporation, a solution containing pVax1-b-Gal or pNCure-fIntr (as the most productive) plasmid (60 μl, 1 μg/μl) was injected at a single point or at six distinct points in the anterior tibial muscles of experimental mice (three muscles per each group). Furthermore, 72 hours after the injections were administered, the mice were sacrificed, and their anterior tibial muscles were isolated. To assess b-Gal expression, the appropriate muscles were preserved in Tissue-Tek O.C.T. Compound (Sakura) and sliced. Slices were fixed and stained for b-Galactosidase expression (http://www.ihcworld.com/_protocols/special_stains/x_gal.htm), and b-Gal–positive muscle fibers were counted. To assess how the injection of a plasmid into multiple sites of a muscle changed recombinant protein production, an explant culture (Jang et al., 2004) was prepared from the muscles in which pNCure-fIntr had been injected. These muscle explants were conditioned in DMEM High Glucose (SH30022FS; HyClone) with 2% fetal bovine serum (SH3007103; HyClone) and 1% penicillin-streptomycin-glutamine solution (PSG) (10378016; Thermo Fisher Scientific). The conditioned medium was concentrated 20× using a combination of desalting-lyophilization reconstitution and analyzed for recombinant hBDNF and huPA concentration as it was described above. The concentration procedure was necessary to adjust for the rather low productivity of the muscle explants and moderate sensitivity of the hBDNF and huPA ELISA systems.
Nerve Injury Model.
To assess the ability of the plasmid to stimulate peripheral nerve regeneration following nerve crush, a mouse model was used (Karagyaur et al., 2015). Forty mice were randomized into the following four groups (n = 10): a positive and a negative control group and two experimental groups (Supplemental Fig. 1). This number of experimental animals is the minimum needed to assess the pharmacological effects and conduct a statistical analysis of the results. Mice were anesthetized with avertin injections, and the common peroneal nerve (n. peroneus communis) was isolated from the surrounding tissue under sterile conditions and crushed using 1-mm–wide forceps with silicone-coated branches two times for 20 seconds. After the wound site was sewn, the plasmids [pVax1m 1 μg/μl (as control), pNCure-fIntr 1 or 2 μg/μl] were injected at six distinct sites in the anterior tibial muscles of mice from the Control, pNCure1, and pNCure2 groups. Plasmid injections were repeated at 4, 9, and 13 days after nerve crush (Supplemental Fig. 1).
Mice from the positive control group (B12) were given subcutaneous injections of a vitamin B12 solution (65 μg/kg) on the day of the trauma and then every 48 hours until the end of the experiment.
Measurements
HEK293 Transfection and ELISA of Conditioned Media.
To determine which plasmid variant provided maximal recombinant protein expression, HEK293 cultures were transfected with pNCure plasmid variants, pVax1-hBDNF, and the control plasmid pVax1m (in duplicates) using Lipofectamine2000 according to the manufacturer’s instructions. Transfected cells were cultivated in DMEM High Glucose (SH30022FS; HyClone) supplemented with 2% FBS (SH3007103; HyClone) and 1% PSG solution (10378016; Thermo Fisher Scientific). Seventy two hours after transfection, the conditioned media was collected and analyzed for hBDNF and huPA concentrations using the Human Free BDNF Quantikine ELISA Kit (DBD00; R&D) and the uPA (URK) Human ELISA Kit (ab119611; Abcam), respectively.
In Vitro Testing of the Ability of hBDNF and huPA to Stimulate Neurite Outgrowth in Mouse Neuro2a Cells.
The murine neuroblastoma cell line Neuro2a was used as a neurite outgrowth in vitro model. This cell line is known to differentiate (i.e., enable neurite outgrowth) in serum deprivation conditions (Semina et al., 2016; Namsi et al., 2018). The Neuro2a cell line was cultured in DMEM High Glucose (SH30022FS; HyClone) supplemented with 10% FBS (SH3007103; HyClone), 1% PSG solution (10378016; Thermo Fisher Scientific), and 1% minimal essential medium nonessential amino acids (11140050; Thermo Fisher Scientific).
HEK293 cultures were transfected with pVax1m, pVax1-hBDNF, and pNCure-fIntr plasmids. Neuro2a cells were seeded in a 24-well plate in low confluent (10%–15%). After 24 hours of cultivation of the cells in DMEM High Glucose with 10% FBS, the media was replaced with DMEM High Glucose containing 1% FBS to stimulate neurite outgrowth. Conditioned media from HEK293 cells transfected with pNCure-fIntr, pVax1-hBDNF, or control plasmid pVax1m was added to the Neuro2a cells (with final hBDNF concentration in wells “pNCure-fIntr,” “pVax1-hBDNF” about 50 ng/ml).
Some wells were used as positive (1% FBS) or negative (10% FBS) control groups to assess neurite outgrowth. Each group existed in triplicates. Cells were incubated for at least 72 hours. Pictures were taken with a bright light microscope equipped with a charge-coupled device camera. Neurites were counted in each group using ClickCounter2 software and normalized per number of cells.
Nerve Regeneration Assessment.
On the 18th day after injury, the effect of the multiple plasmid injections on functional and structural nerve recovery was assessed. Nerve conductivity and the number of axons were evaluated. Nerve conductivity measurements were performed as previously described (Karagyaur et al., 2015). The animals were sacrificed, and the injured nerves were isolated and placed in a Laily solution (135.0 mМ NaCl, 4.0 mМ KCl, 2.2 mМ CaCl2, 1.0 mМ MgCl2, 0.9 mМ NaH2PO4, 16.3 mМ NaHCO3, 11.0 mМ D-glucose, pH 7.2–7.4). Three series of compound nerve action potentials (CNAPs) were independently recorded for each nerve using a monopolar aspirating silver electrode, 10 mm distal from the injury site, connected to an analog-to-digital converter and output, and they were analyzed using PowerGraph Professional 3.3 software. The length of the latency period and the amplitude of the CNAPs were measured. The number of axons was evaluated on formalin-fixed frozen sections of a common peroneal nerve fragment, 3 mm distal to injury site, using immunofluorescent staining with rabbit anti-NF200 antibody (catalogue number 4142; Sigma) and morphometric analysis (Lopatina et al., 2011; Karagyaur et al., 2015).
Statistical Analysis
Statistical analysis was performed using SigmaPlot11.0 Software. Data were assessed for normality of distribution using the Kolmogorov–Smirnov test. Differences between the treatment and control groups were then analyzed using the Student’s t test or ANOVA on ranks Dunn’s test, depending on whether there was normal distribution or not. Data are expressed as mean ± S.D. or median (25%; 75%) depending on the test used. P < 0.05 was considered statistically significant.
Results
Plasmid Variant pNCure-fIntr Provides Maximum Expression of hBDNF and huPA in HEK293 Cells.
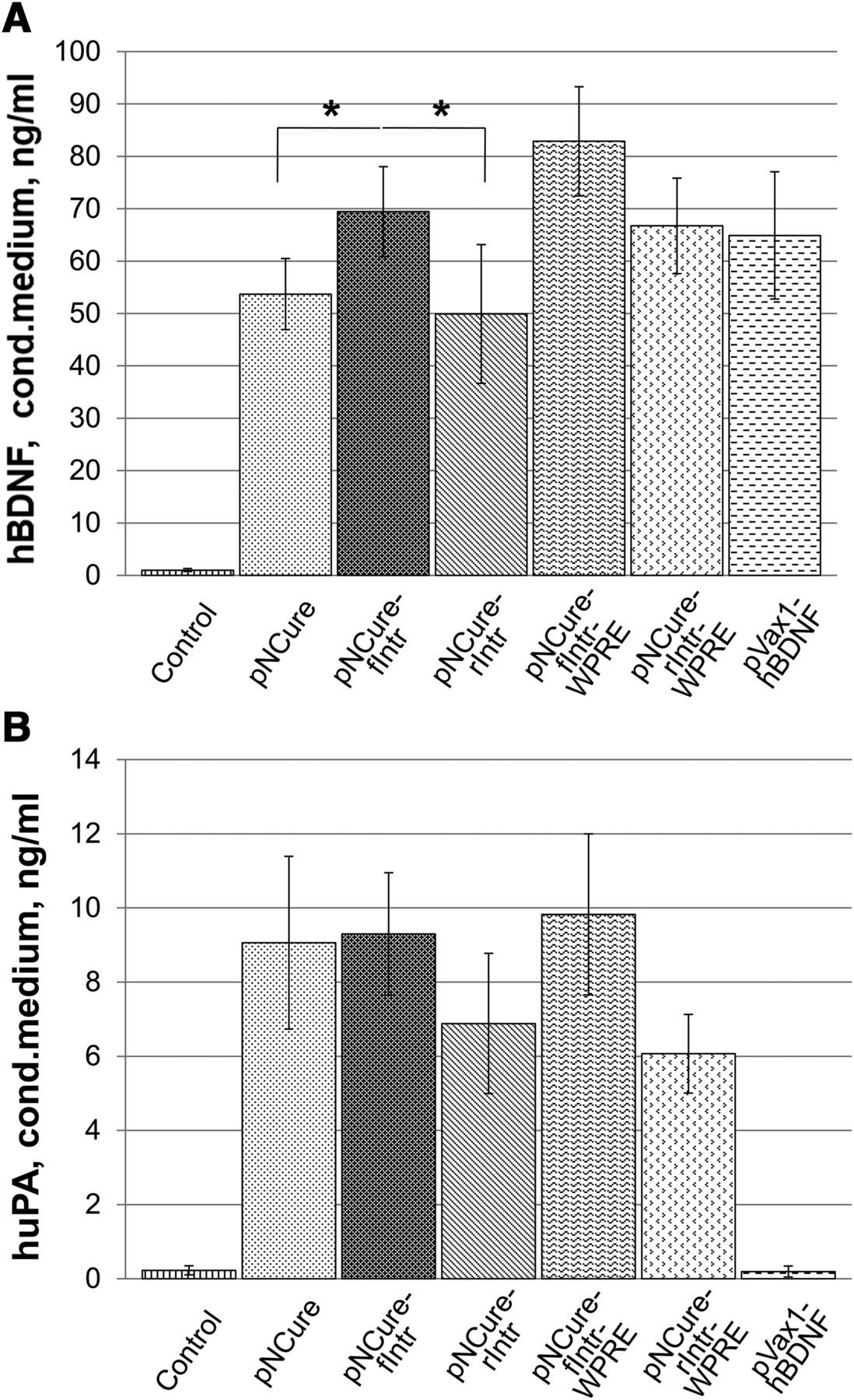
Analysis of the expression of recombinant hBDNF and huPA in HEK293 cells showed that the plasmid pNCure variant containing the synthetic intron sequence fIntr produced significantly higher hBDNF than the basic pNCure plasmid (69.3 ± 9.1 vs. 53.3 ± 7.3 ng/ml; n = 3, *P < 0.05). However, huPA production did not differ significantly between the pNCure-fIntr and pNCure plasmid variants (9.2 ± 1.4 vs. 9.0 ± 2.5 ng/ml; n = 3) (Fig. 2). The addition of the WPRE element pNCure-f/rIntr-WPRE did not significantly increase the production of recombinant proteins. However, it did increase the size of the plasmid by 10%. For this reason, we decided not to use variants of the pNCure plasmid containing WPRE in this study. The effect of the pNCure-fIntr plasmid on hBDNF expression was almost equivalent to that of pVax1-hBDNF, as previously described (Karagyaur et al., 2015).

pNCure plasmid variants provide an acceptable level of hBDNF (A) and huPA (B) expression in culture of HEK293. Data are represented as mean ± S.D. (n = 3). *P < 0.05.
pNCure-fIntr Plasmid Stimulates Neurite Outgrowth in a Neuro2a Cell Model.
The conditioned media stimulated neurite outgrowth in all groups compared with negative control group (DMEM supplemented with 10% FBS). However, the most prominent effect was observed in the pVax1-hBDNF and pNCure-fIntr groups, in which 29.4% ± 2.5% and 34.3% ± 3.5% of cells had neurites, respectively. The difference was significant compared with the negative control, positive control, and pVax1m groups, in which 8.2% ± 1.4%, 22.9% ± 2.7%, and 18.3% ± 2.1% of cells had neurites, respectively (n = 20, *P < 0.05) (Fig. 3). The number of neurites in the positive control and pVax1m groups differed significantly from the negative control group (n = 20, **P < 0.05). No significant difference was observed between the pVax1-hBDNF and pNCure-fIntr groups, but a tendency toward better neuritogenesis was noted in the pNCure-fIntr group.

A conditioned media of HEK293 cells transfected with pVax1m-hBDNF and pNCure-fIntr plasmids stimulates neurite outgrowth in a Neuro2a cell model. On the pictures are Neuro2a cell cultures 72 hours following neuritogenesis induction. (A) negative control (DMEM 10% FBS); (B) positive control (DMEM 1% FBS); (C–E) conditioned media from HEK293 cells, transfected with pVax1m, pVax1m-hBDNF, or pNCure-fIntr plasmids (original magnification, 100×). The graph plot demonstrates the percentage of cells with neurites in each group. Data are represented as mean ± S.D. (n = 20). *P < 0.05 vs. negative control, positive control, and pVax1m group; **P < 0.05 vs. negative control group.
Multiple Intramuscular Injections of a Plasmid Are as Efficient as a Single-Point Injection.
In our previous studies, we used a single intramuscular injection for therapeutic gene delivery to stimulate repair of the damaged nerve (Karagyaur et al., 2015). However, the approach used was not relevant in terms of clinical application. Hydrodynamic shock and electroporation are not used in clinical settings, as they cause pain and significant traumatic effects. Multiple intramuscular injections of small amounts of a plasmid solution (known as muscle chipping) are more clinically relevant, but it was unclear whether they would provide the acceptable level of recombinant protein expression. Using the marker plasmid (pVax1-b-Gal), we showed that muscle chipping provides plasmid delivery to muscle fibers of equal efficacy to a per saltum injection accompanied by hydrodynamic shock. Injections at six different points of the anterior tibial muscle provided transfection of 68.3 ± 12.7 muscle fibers (β-galactosidase positive), whereas a per saltum injection of a pVax1-b-Gal plasmid solution accompanied by hydrodynamic shock resulted in transfection of only 52.8 ± 11.5 muscle fibers (the difference is not significant, n = 3) (Fig. 4).

Multiple intramuscular injections of small volumes of plasmid solution (muscle chipping) provide the same amount of plasmid DNA delivery as a single injection accompanied by hydrodynamic shock (b-galactosidase staining) (original magnification, 100×, n = 3). Data are represented as mean ± S.D. A,B - muscle cryosections stained for b-galactosidase. C - an average number of b-galactosidase positive muscle fibers per muscle cryosection. D,E - the content of hBDNF and huPA in conditioned medium obtained from muscle explants (ELISA).
The analysis of hBDNF and huPA in a concentrated conditioned culture medium from muscle explants revealed that muscle chipping with small amounts of the plasmid pNCure-fIntr provided the same level of recombinant protein production. Six injections of pNCure-fIntr (muscle chipping) resulted in 136.5 ± 18.4 pg/ml hBDNF and 12.3 ± 1.7 pg/ml huPA, whereas a single intramuscular injection of pNCure-fIntr accompanied by hydrodynamic shock resulted in 107.3 ± 12.5 pg/ml hBDNF and 9.6 ± 2.0 pg/ml huPA in a culture medium. The difference between two groups was not significant, but an increase in recombinant protein expression in the muscle chipping group was noted (n = 3) (Fig. 4).
Because the effectiveness of the clinically relevant method of muscle chipping with the plasmid solution was confirmed experimentally, this delivery method was used for evaluating of the therapeutic efficacy of a constructed bicistronic plasmid vector.
A Bicistronic Plasmid Encoding hBDNF and huPA Stimulates Post-Traumatic Functional and Structural Nerve Recovery.
Crushing of the common peroneal nerve caused nerve dysfunction, as evidenced by a decrease in CNAP amplitude and CNAP latent period extension (Supplemental Fig. 2). Intramuscular injections of pNCure-fIntr stimulated the functional and structural recovery of the crushed nerve. On the 18th day of the experiment, CNAP amplitude was 1.7- and 2.2-fold higher in animals from the pNCure1 and pNCure2 groups compared with the Control group animals, which received injections of an empty pVax1m vector [0.475 (0.338; 0.612) and 0.627 (0.506; 0.832) mV versus 0.285 (0.245; 0.353) mV, respectively (n = 10, ***P < 0.001 vs. control)] (Fig. 5). A comparable increase in CNAP amplitude was also observed in the B12 control group [0.457 (0.329; 0.573) mV (n = 10; *P < 0.05 vs. control)]. CNAP amplitudes between the pNCure1 and pNCure2 groups did not differ significantly (P = 0.174).

The pNCure-fIntr plasmid stimulates nerve conduction recovery. (A) CNAP amplitude; (B) CNAP latency. The maximum stimulating effect was observed after pNCure-fIntr injection at a dosage of 120 μg/mouse. The average CNAP amplitude and latency of an intact nerve corresponds to the dashed lines. Data are represented as median with 25% and 75% (percentile) (n = 10). *P < 0.05 and ***P < 0.001 vs. control.
The latency period of the CNAP of the damaged nerve (conductivity indicator) did not differ significantly in all groups, but a tendency to its diminishing with dose increments was observed in the pNCure1 and pNCure2 groups (Fig. 5). The latency was 0.715 (0.622; 0.893) ms in the pNCure1 group and was 0.507 (0.45; 0.887) ms in the pNCure2 group, whereas in Control and B12 groups, the latency was 0.861 (0.68; 0.983) and 0.771 (0.7; 0.94) ms, respectively.
An analysis of nerve cryosections stained for markers of mature axons NF200 revealed that the pNCure-fIntr plasmid stimulated the recovery of the nerve structure. Both the number and size of the positively stained structures in nerves from the pNCure1 and pNCure2 groups was larger compared with nerves from the Control group animals. On the 18th day of the experiment, nerve sections from the pNCure1 and pNCure2 groups contained 573 (489; 619) and 554 (518; 633) positively stained structures (nerve fibers), respectively (n > 5, *P < 0.05 vs. Control), whereas in Control and B12 groups, the number of positively stained structures was 276 (234; 311) and 461 (407; 545) (n > 5, *P < 0.05 vs. Control) (Fig. 6). No significant difference between the pNCure1, pNCure2, and B12 groups was observed, though a tendency toward a higher number of restored nerve fibers was noted in the pNCure1 and pNCure2 groups.

The pNCure-fIntr plasmid stimulates nerve structure recovery. On the pictures are nerve stump cryosections 3 mm distally from the crush site (on the 18th day after crush) immunostained with axon-specific anti-NF200 antibody (original magnification, 50×). The graph plot demonstrates the number of NF200-positive structures per cryosection in each group. The average number of axons of an intact nerve corresponds to the dashed line. Data are represented as mean ± S.D. (n > 5). *P < 0.05 vs. control.
Discussion
Successful tissue regeneration requires restoration of its blood and nerve supply. Though a blood vessel can be regrown from a number of endothelial cell islets in a multifocal mode, a damaged nerve fiber regenerates only from its proximal stump along the nerve bed and is highly sensitive to neurotrophin expression (Benga et al., 2017; Karagyaur et al., 2017). The lack of effective specific drugs and the abundance of disabilities caused by nerve injury means that new therapeutic approaches should be sought. Regeneration is known to be a complex and multistage process, and this changes the paradigm of the current proregenerative therapy (Li et al., 2016; Wang et al., 2017; Nimiritsky et al., 2019). It is becoming increasingly clear that it is necessary to influence the different stages of the regenerative process to obtain the desired outcome. This can be achieved by combining medicines (e.g., gene therapy molecules) with different modalities.
In this paper, we studied the efficacy of a combined gene therapy with drug encoding brain-derived neurotrophic factor and urokinase plasminogen activator on the stimulation of damaged nerve fiber recovery in a model of peripheral nerve injury. The combination of a growth factor and a protease appeared to be beneficial; BDNF stimulated neuron survival and neurite growth (Ebendal, 1992), whereas uPA is a key molecule in fibrin degradation (the main inhibitor of neurite growth after trauma), neurite navigation, matrix remodeling, and growth factor maturation (Siconolfi and Seeds, 2001). For proper folding, most proteins are synthesized in preforms and then maturate via protease-mediated cleavage (matrix metalloproteinases, plasmin, plasminogen activators). BDNF is one such molecule. Moreover, its proform, proBDNF, has greater affinity to the alternative neurotrophin receptor p75 that activates neuron death program (MacPhee and Barker, 1997). Mature BDNF stimulates neuron survival and neurite elongation via TrkB (Tropomyosin receptor kinase B) receptor activation (Ebendal, 1992), and earlier research has shown that plasmin, activated by uPA or tPA (tissue plasminogen activator), causes maturation of BDNF and other growth factors (Rifkin et al., 1997).
As the aim of this study was to create a gene therapy drug for treating nerve injury in humans, human orthologs of BDNF and uPA were encoded in pNCure plasmid. The BDNF sequence is highly conservative among the species, and its human ortholog can be successfully used and tested in murine models (Lucini et al., 2018). However, uPA functions by interacting with the receptor uPAR, which has strong species specificity (Lin et al., 2010), and this makes the usage and testing of the human uPA ortholog in murine models impossible. Despite this, the ability of hBDNF and muPA to stimulate the regeneration of the crushed murine nerve was demonstrated in our previous studies (Karagyaur et al., 2015). Moreover, here, we showed that a bicistronic plasmid encoding hBDNF and huPA expressed in human cell line HEK293 has a tendency to promote better neuritogenesis in Neuro2a cells in vitro. One possible explanation is that the expression of huPA in human cell lines allows the activation of huPA via its binding to uPAR on the HEK293 cell surface, which leads to the conversion of plasminogen to plasmin and further growth factor activation. Previously, it was shown that the plasminogen/plasmin system plays an important role in the activation of many proteins, including growth factors (Rifkin et al., 1997). Unexpectedly, the pVax1m-conditioned media also provided significant neurite outgrowth. We propose that this could be caused by the basic productivity of cytokines and growth factors by the HEK293 cell line.
To increase the productivity of the basic pNCure plasmid, some short regulatory elements were introduced. A short synthetic intron, inserted before the Kozak consensus sequence and start codon, significantly increased protein expression, whereas the addition of the WPRE element after the stop codon did not. As they increased the size of the plasmid by 10% and had an insignificant influence on protein expression, the variants of pNCure containing WPRE were not used in this study. The results of the study demonstrated that the effect of the pNCure plasmid variants constructed here on hBDNF production did not differ dramatically from the previously studied pVax1-hBDNF plasmid despite the addition of the second gene urokinase plasminogen activator and an increase in plasmid size of 20%.
In our previous study (Karagyaur et al., 2015), we showed that a single intramuscular injection of hBDNF-encoding plasmid, followed by an electroporation procedure, provided hBDNF expression in murine tissues for 7–10 days and stimulated the repair of a damaged nerve. However, the approach used was not relevant to clinical applications; hydrodynamic shock and electroporation are not used in clinical settings because they cause pain and traumatic effects. Furthermore, it should be kept in mind that recombinant proteins in human tissues have to be expressed for weeks or even months to complete the regeneration process. The small size and high regenerative potential ensure effective recovery of a crushed nerve, even with weak expression of neurotrophic factors in mice but not in humans. The clinical use of gene-therapeutic drugs requires repeated injections and prolonged protein expression. Here, we showed that the clinically relevant method of multiple intramuscular plasmid injections is as efficient as the previously used single-site injection. Additional tests showed that repeated multiple intramuscular injections of a bicistronic plasmid still provided local protein expression and did not increase their ectopic expression (Karagyaur et al., in press). Because the effectiveness of this delivery method was confirmed experimentally, it was used in this study for bicistronic pNCure-fIntr plasmid intramuscular delivery in a murine nerve injury model.
Based on our previous experience of in vivo testing of hBDNF- and muPA-encoding plasmids, the results of previous in vitro testing, and the absence of relevant human nerve injury models, we decided to test the activity of a pNCure-fIntr bicistronic plasmid in the familiar murine nerve crush model. We were not expecting expressed huPA to work as well in mice as it did in humans.
In vivo testing in mice showed that the bicistronic pNCure-fIntr plasmid stimulated the conductivity restoration of a damaged nerve. The amplitude of CNAP in mice treated with the pNCure-fIntr plasmid was greater than in the control group. Because CNAP’s amplitude in direct ratio characterizes the number of nerve fibers involved in nerve impulse conduction (Fugleholm et al., 2000; Vleggeert-Lankamp, 2007), we concluded that the pNCure-fIntr plasmid stimulated neurite post-traumatic survival and outgrowth in vivo. This data confirmed the results of in vitro testing and was supported by immunohistochemical studies of nerve structure (Fig. 6). The latent period (i.e., the time interval from the moment of stimulation to the moment of CNAP registration) that characterizes the speed of nerve conductance in inverse ratio was not restored significantly in the pNCure-fIntr group, though some improvement was observed. This indicates that the pNCure-fIntr plasmid did not significantly increase the myelination of restored nerve fibers. The most prominent effect of the pNCure-fIntr plasmid was observed at the higher dose of 120 μg (Fig. 5).
Because the concept of the experiment indicated multiple plasmid injections that could not last less than 18 days, relatively sparing method of damage to the peripheral nerve and high endogenous regenerative potential of the mice leveled the statistically significant difference during the latent period between the experimental and control groups. This was confirmed by the magnitude of the CNAP latent period in the experimental and control groups, in which the values were close to those for the intact nerve (0.6 ms).
Vitamin B12 was used as a positive control for in vivo studies because it is one of the few drugs registered in Russia for treating nerve injuries, and it shares ras/erk and Akt signal cascades with the neurotrophines.
We found that the bicistronic plasmid pNCure-fIntr, encoding the human orthologs BDNF and uPA, was able to stimulate neuritogenesis in vitro and post-traumatic peripheral nerve conductivity and structure restoration in vivo. This apparently occurred because of the activation of neuron survival and growth of the crushed axons.
To our knowledge, this is the first study that has tested a bicistronic plasmid construction encoding a neurotrophin and plasminogen activator for enhancement of the multicomponent and multistage process of nerve fiber regeneration.
As treatment outcomes for many neurologic and neurosurgery disorders are poor, and as BDNF and uPA are important growth factors in nerve regeneration, this successfully tested bimodal plasmid, in combination with an appropriate delivery system, could serve as the basis for developing new prospective treatments for a wide range of multicomponent neural system disorders, such as strokes and neurodegenerative disorders.
Authorship Contributions
Participated in research design: Karagyaur, Semina, Balabanyan, Stambolsky, Tkachuk.
Conducted experiments: Karagyaur, Rostovtseva, Semina, Klimovich, Popov.
Contributed new reagents or analytic tools: Karagyaur, Semina, Balabanyan.
Performed data analysis: Karagyaur, Rostovtseva, Semina, Klimovich, Balabanyan, Makarevich, Stambolsky, Tkachuk.
Wrote or contributed to the writing of the manuscript: Karagyaur, Semina, Balabanyan, Makarevich, Stambolsky, Tkachuk.
Footnotes
- Received July 29, 2019.
- Accepted December 17, 2019.
The study was supported by the Russian fund for basic research, project 18-015-00535. This study was conducted using biomaterial collected within the frame of Moscow University Project “Noah’s Ark” and equipment purchased as a part of Moscow State University Program of Development.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- BDNF
- brain-derived neurotrophic factor
- b-Gal
- b-Galactosidase
- CMV
- citomegalovirus
- CNAP
- compound nerve action potential
- DMEM
- Dulbecco’s modified Eagle’s medium
- hBDNF
- human BDNF
- HCV
- hepatitis C virus
- HEK293
- human embryonic kidney 293 cell line
- huPA
- human uPA
- IRES
- internal ribosome entry site
- muPA
- murine uPA
- pNCure
- a plasmid for nerve cure
- pNCure-f/rIntr
- a plasmid for nerve cure with a synthetic intron (in forward or reverse orientation) before hBDNF start codon
- PSG
- penicillin-streptomycin-glutamine solution
- uPA
- urokinase plasminogen activator
- uPAR
- uPA receptor
- WPRE
- woodchuck hepatitis virus posttranscriptional regulatory element
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}