


Abstract
The human organic cation transporter 2 (OCT2) mediates the first step of tubular secretion of most positively charged substances. We describe the role of plasma membrane cholesterol in OCT2 activity. Human embryonic kidney 293 cells overexpressing OCT2 (OCT2-HEK293) and wild-type HEK293 cells (WT-HEK293) were employed. Cellular cholesterol content, assessed by thin layer chromatography, was manipulated using empty methyl-β-cyclodextrin (mβcd) and cholesterol-presaturated mβcd (RAMEB). The effect of mβcd on OCT2 protein stability and oligomerization state was evaluated by immunofluorescence and Western blotting. Transport activity of OCT2 was measured using [3H]1-methyl-4-phenylpyridinium (MPP+). A 20-minute incubation with mβcd reduced the total cellular cholesterol content by 40% to 60% as compared with that in untreated cells, without altering the content of the other main lipid species. In this condition, OCT2-mediated uptake of MPP+ was reduced by ∼50%. When cells were coincubated with empty mβcd and RAMEB, the cholesterol content and OCT2-mediated uptake of MPP+ were comparable to those in untreated cells, suggesting that the mβcd effect on OCT2 activity was cholesterol dependent. In untreated cells, the MPP+ influx kinetics was allosteric, whereas in cells treated with mβcd, one binding site was observed. Our findings suggest that changes in cellular cholesterol content can dramatically alter OCT2-mediated transport, potentially resulting in abnormal tubular secretion and unexpected drug toxicity and drug–drug interactions.
SIGNIFICANCE STATEMENT Plasma membrane cholesterol is important for the allosteric properties of OCT2. From a pharmacologic standpoint, the variability in cholesterol content stemming from certain pathophysiologic conditions such as aging and acute kidney injury should be taken into account as additional source of interpatient pharmacokinetic/pharmacodynamic variability and unexpected toxicity profile of OCT2 substrates, which can escape preclinical and clinical development.
Introduction
The human organic cation transporter 2 (OCT2) is a Na+-independent, polyspecific cation transporter primarily located at the basolateral plasma membrane of renal proximal tubule cells. It mediates the first step of tubular secretion by translocating various substrates from the interstitium (blood side) into the proximal tubule cells. Substrates of OCT2 are several endogenous compounds, such as choline and its metabolite trimethylamine N-oxide, various neurotransmitters, widely used drugs such as cimetidine, oxaliplatin, cisplatin, gentamicin, and metformin, and fluorinated choline (fluorocholine), which is routinely used in positron emission tomography imaging of malignancies such as brain and prostate cancer (Koepsell, 2013; Gai et al., 2016; Visentin et al., 2017, 2018). The pivotal role of OCT2 in renal excretion of drugs is underscored by the guidances of drug regulatory agencies, which demand that each molecule in development be tested in vitro for inhibition of OCT2 transport activity to predict potential drug–drug interactions (http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2012/07/WC500129606.pdf; http://www.fda.gov/downloads/Drugs/GuidanceComplianceRegulatoryInformation/Guidances/UCM292362.pdf).
As an integral membrane protein with 12 predicted transmembrane domains spanning the lipid bilayer, OCT2 is embedded in the plasma membrane, in contiguity with the variety of lipid components of the surrounding microenvironment. In particular, plasma membrane cholesterol is a critical regulator of the activity of several transport proteins as well as signaling pathway receptors (Scanlon et al., 2001; Ringerike et al., 2002; Caliceti et al., 2012; Dickens et al., 2017). First, by interacting with phospholipids, cholesterol regulates the fluidity of the lipid bilayer, which can dictate the mobility of the carrier and, in turn, its binding to the substrate as well as its cycling (Cooper, 1978; Chong, 1994; Radhakrishnan and McConnell, 1999; Radhakrishnan et al., 2000; Huang, 2002; Andersen and Koeppe, 2007). Additionally, cholesterol can directly interact with transmembrane proteins at the level of cholesterol recognition/interaction amino acid consensus sequences (CRAC and CARC) or areas flanking a GXXXG sequence (Fantini and Barrantes, 2013).
The plasma membrane cholesterol content of the proximal tubule cells, similar to that of other tissues, is highly dynamic and subject to substantial changes under both physiologic and pathologic conditions. For instance, the cholesterol-to-phospholipid ratio increases up to 25% during senescence in rat kidney membranes (Grinna, 1977; Hegner, 1980; Pratz and Corman, 1985). Additionally, mice with acute kidney injury display a substantial (up to 50%) increase in the total membrane cholesterol level, depending on the underlying injury model (ischemia/reperfusion or obstructive nephropathy) (Zager et al., 1999).
These fluctuations in membrane cholesterol content may influence the process of tubular secretion. For instance, it has been previously shown that the transport of the organic cation tetraethanolamine bromide (TEA) in brush-border membrane vesicles is elevated by cholesterol enrichment (Nabekura et al., 1996). Our present work studies the effect of the chemical depletion of cholesterol on OCT2 activity in intact cells overexpressing human OCT2.
Materials and Methods
Reagents.
N-methyl-4-phenylpyridinium, acetate [N-methyl-3H]-N-methyl-4-phenylpyridinium ([3H]MPP+, specific activity: 81.3 Ci/mmol), and cholesterol [4-14C]-cholesterol ([14C]cholesterol, specific activity: 50.8 mCi/mmol) were purchased from PerkinElmer (Boston, MA). Nonlabeled MPP+, methyl-β-cyclodextrin (mβcd), cholesterol, sphingomyelin, phosphatidylcholine, phosphatidylserine, phosphatidylinositol, and phosphatidylethanolamine were provided by Sigma-Aldrich (St. Louis, MO). Cholesterol-presaturated mβcd (RAMEB) was provided by CycloLab Ltd. (Budapest, Hungary). Penicillin/streptomycin mixtures, Geneticin G-418, and Dulbecco’s modified Eagle’s medium (DMEM) culture medium were purchased from ThermoFisher Scientific (Waltham, MA). Biowest fetal bovine serum was provided by VWR (Dietikon, CH). Poly-d-lysine was purchased from Corning (Bedford, MA).
Cell Lines.
Wild-type human embryonic kidney 293 cells (WT-HEK293) were maintained in DMEM supplemented with 10% FBS, 100 U/ml penicillin, and 100 µg/ml streptomycin at 37°C in a humidified atmosphere of 5% CO2. Cells stably expressing the pcDNA3.1(+) plasmid containing the untagged coding sequence of the human OCT2 (OCT2-HEK293) were grown under selective pressure with Geneticin G-418 at the extracellular concentration of 400 µg/ml (Thévenod et al., 2013).
Plasma Membrane Integrity Assay.
To assess cell membrane integrity, WT-HEK293 cells were seeded on 12-well plates coated with 0.1 mg/ml poly-d-lysine at a density of 2 × 105 cells/well. After 48 hours the cells were washed, treated for 20 minutes at 37°C with mβcd dissolved in transport buffer (116.4 mM NaCl, 5.3 mM KCl, 1 mM NaH2PO4, 0.8 mM MgSO4, 5.5 mM d-glucose, and 20 mM HEPES/KOH, pH 7.4), then washed with PBS and finally resuspended in a 0.02% Trypan blue solution for cell counting.
Immunofluorescence of OCT2-HEK293 Cells.
The cells were seeded on chamber slides at a density of 4 × 104 cells per chamber, coated with 0.1 mg/ml poly-d-lysine. Seventy-two hours later, the cells were washed with cold PBS, fixed in 4% paraformaldehyde, and treated with 0.1% Triton X-100 in PBS for 15 minutes and with 0.1% Tween-20 in 1% bovine serum albumin/PBS for 30 minutes. The cells were then incubated at 4°C overnight with an antibody raised against the full-length OCT2 protein (Clone #640438; R&D Systems, Minneapolis, MN). After washing, the cells were mounted with 4ʹ,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA) and visualized under a fluorescent microscope (DMI6000B; Leica Microsystems, Wetzlar, Germany).
Lipid Extraction.
Lipid extraction was performed with a standard chloroform/methanol method (Folch et al., 1951). The cells were seeded at a density of 0.5–1 × 106 cells per dish on 10-cm dishes precoated with 0.1 mg/ml of poly-d-lysine. After 72 hours, the cells were gently scraped from the plate and spun down for 5 minutes at 900gav. The pellet was resuspended in 1 ml of PBS. One hundred microliters were lysed with 900 µl of 0.1% (w/v) Triton X-100 in deionized water for bicinchoninic acid protein determination.
The remaining cell suspension was mixed with 3 ml of chloroform:methanol (2:1) solution spiked with 14C-cholesterol, serving as an internal standard. The amount of 14C-cholesterol added as the internal standard did not exceed 1% of the total cholesterol content measured in untreated cells. After 20 minutes of shaking, samples were centrifuged for 5 minutes at 3000gav for phase separation. The lower phase containing the lipid fraction was dried under a faint nitrogen flux at 30°C. Finally, the lipid pellet was resuspended in 600 µl of ice-cold chloroform. A 50-µl aliquot was used for assessing radioactivity by liquid scintillation counting.
Thin Layer Chromatography.
For analysis of the cholesterol and phospholipid content, aliquots from the extracted lipids were loaded on high-performance thin layer chromatography (HPTLC) Silica Gel 60 plates with a concentrating zone (Merck KGaA, Darmstadt, Germany) using an automated Camag thin layer chromatography (TLC) sampler ATS4 and separated by one-dimensional TLC. Cholesterol was resolved in 62.3% n-hexane, 18.3% n-heptane, 18.4% diethyl ether, and 1% acetic acid. Phospholipids were resolved in 26.6% methyl acetate (v/v), 26.6% 1-propanol (v/v), 26.6% chloroform (v/v), 10.6% methanol (v/v), and 0.05% KCl (w/v). Staining was performed in 9.6% ortho phosphoric acid (v/v) and 3% copper acetate (w/v), and then the plate was dried at 120°C for 30 minutes.
The bands were scanned at 366 nm, and absolute quantification performed from a serial dilution of the main lipid species separated in parallel. The values were then normalized for the [14C]cholesterol recovery yield and the protein content.
Cell Surface Labeling.
Nearly confluent OCT2-HEK293 cells were washed twice with PBS and treated with 1 mg/ml of EZ-Link Sulfo-NHS-LC-biotin in PBS on ice for 1 hour. This nonpermeable reagent alkylates the accessible Lys residues. The cells were then washed twice and incubated for 30 minutes with hypotonic buffer (0.5 mM Na2HPO4 and 0.1 mM EDTA at pH 7.0) containing protease inhibitors. The cells were then scraped from the plates and centrifuged at 16,000gav for 15 minutes at 4°C. The pellet was lysed in 200 μl of lysis buffer (0.1% SDS, 1% Triton X-100, 1 mM EDTA, 150 mM NaCl, and 20 mM Tris, pH 7.4) containing protease inhibitors and rotated for 2 hours at 4°C, then spun down at 16,000gav at 4°C for 15 minutes.
A portion of the supernatant was used for bicinchoninic acid protein determination and for total lysate analysis. The remaining portion was mixed with streptavidin-agarose beads (50 μl/sample) overnight at 4°C. The beads were washed twice in lysis buffer and twice in lysis buffer supplemented with 2% (w/v) SDS. After the final wash, the bead-bound proteins were stripped; the samples were incubated at 95°C for 5 minutes in Laemmli buffer containing 1.5% (w/v) dithiothreitol (DTT) and loaded directly onto polyacrylamide gels. The total lysate samples were denaturated using the same conditions.
SDS Gel Electrophoresis and Western Blotting.
Surface labeling protein samples and the respective total lysates were resolved on 8% (w/v) polyacrylamide gels. To assess the OCT2 oligomerization state, total lysate samples were denatured for 15 minutes at 50°C in Laemmli buffer with or without 1.5% DTT (w/v), then were resolved on a gel with a 6% to 12% (w/v) gradient of polyacrylamide. Proteins were electroblotted onto nitrocellulose membranes (GE Healthcare, Piscataway, NJ). The membranes were blocked with 5% (w/v) nonfat dry milk in PBS supplemented with 0.1% (w/v) Tween-20 and were incubated overnight at 4°C with anti-OCT2 (Clone #640438; R&D Systems), followed by probing with a horseradish peroxidase-conjugated secondary antibody. Blots were developed with SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Scientific) and Fusion FX7 (Vilber Lourmat, Eberhardzell, Germany). As a control, the sample blots were probed with anti-pan-actin.
Transport Studies in Intact Cells.
Uptake of radiolabeled compounds was measured using a protocol designed for uptake determination in cells (Schroeder et al., 1998; Visentin et al., 2015). WT-HEK293 and OCT2-HEK293 cells were seeded at a density of 2.5 × 105 cells/dish on 35-mm dishes coated with 0.1 mg/ml poly-d-lysine (Corning). After 72 hours, the cells were washed and equilibrated in transport buffer (116.4 mM NaCl, 5.3 mM KCl, 1 mM NaH2PO4, 0.8 mM MgSO4, 5.5 mM d-glucose, and 20 mM HEPES/KOH, pH 7.4) at 37°C, then the buffer was aspirated and transport buffer containing the radiolabeled substrate was added. Uptake was stopped by quick aspiration followed by extensive washing with ice-cold transport buffer.
The cells were solubilized and the intracellular radioactivity was assessed by liquid scintillation counting. Protein content was determined by the bicinchoninic acid protein assay (Interchim, Montluçon, France). OCT2-independent uptake was determined in WT-HEK293 cells and subtracted from the total uptake to quantify the OCT2-mediated transport. Uptake is expressed as picomoles of substrate per milligram of protein.
Statistical Analysis.
Statistical comparisons were performed using GraphPad Prism (version 5.0 for Windows; GraphPad Software, San Diego, CA). Comparisons between two groups were performed with the two-tailed Student’s unpaired t test. For multiple comparisons, one-way ANOVA followed by Bonferroni’s post hoc test was employed.
Results
Acute Exposure to mβcd Removed Cholesterol without Changing the Content of the Other Major Classes of Lipids nor Affecting Cell Membrane Integrity.
Cyclodextrins are torus-shaped cyclic oligosaccharides with a hydrophobic inner cavity that attracts and encapsulates nonpolar substances in close proximity. Among cyclodextrin molecules, methyl-β-cyclodextrin (mβcd) preferentially accepts and scavenges cholesterol (Zidovetzki and Levitan, 2007). WT-HEK293 cells exposed to mβcd for 20 minutes showed a dose-dependent cholesterol depletion (Fig. 1A). At the highest extracellular concentration of mβcd tested, the total cholesterol content of WT- and OCT2-HEK293 cells was reduced by ∼60% and 40% respectively, as compared with the respective untreated cells (Fig. 1B).

Cholesterol content and membrane integrity of WT- and OCT2-HEK293 upon 20-minute incubation with mβcd. (A) Representative TLC plate from WT-HEK293 cells. (B) Quantification of total cholesterol. Data are relative to the cholesterol content in untreated cells. (C) Percentage of cells impermeable to Trypan Blue dye. All results represent the mean ± S.D. from three independent experiments.
Because most of the cellular cholesterol resides within the plasma membrane, it can be assumed that total cholesterol content reflects that of the plasma membrane (Lange et al., 1989). Because cholesterol has a time- and concentration-dependent effect on membrane organization and permeability (Raffy and Teissie, 1999), the effect of mβcd-induced cholesterol depletion on membrane permeability was also assessed.
WT-and OCT2-HEK293 cells were exposed for 20 minutes to increasing extracellular concentrations of mβcd and then mixed with Trypan blue solution for cell counting. Figure 1C shows that the majority of cells were impermeable to Trypan blue, suggesting preserved membrane integrity. Although mβcd is considered relatively specific for cholesterol, some studies have reported the depletion of other lipids as well (Ottico et al., 2003; Giocondi et al., 2004; Monnaert et al., 2004). After a 20-minute incubation of WT- and OCT2-HEK293 cells with mβcd at an extracellular concentration of 10 mM, no obvious depletion of the other major classes of lipids was observed (Fig. 2).

Phospholipid content of WT- and OCT2-HEK293 upon 20-minute incubation with mβcd. (A) Representative TLC plate of the main lipid species from WT-HEK293 cells. (B–F) Quantification of total sphingomyelin (SM), phosphatidylcholine (PC), phosphatidylserine (PS), phosphatidylinositol (PI), and phosphatidylethanolamine (PE) content. Data are relative to the respective content in untreated cells and represent the mean ± S.D from three independent experiments.
Mβcd Exposure Impaired MPP+ Influx Mediated by OCT2.
Figure 3A illustrates the time course of MPP+ uptake over 15 minutes at an extracellular concentration of 0.5 μM at pH 7.4 in WT- and OCT2-HEK293 cells. The increase in intracellular MPP+ rapidly deviated from linearity, reaching equilibrium within 5 minutes. The time course over a shorter interval showed that MPP+ uptake as a function of time was linear over 15 seconds, reflecting the unidirectional flux of MPP+ into the cells. The transport rate of MPP+ in OCT2-HEK293 cells was ∼36 times higher than that in the WT-HEK293 cells (slope, 0.36 ± 0.01 vs. 0.01 ± 0.02 pmol mg−1 s−1) (Fig. 3B).

OCT2-mediated MPP+ transport. (A and B) Time course of MPP+ at the extracellular concentrations of 0.5 µM in WT- and OCT2-HEK293 cells. Representative of two independent experiments. (C) OCT2-mediated MPP+ influx at the extracellular concentrations of 0.5 µM after 20-minute incubation with mβcd. Data were corrected for uptake in WT-HEK293 cells and represent the mean ± S.D. from three independent experiments. (D) OCT2-mediated MPP+ influx rate at the extracellular concentration of 1 µM in the presence of mβcd. Coincubation with nonlabeled MPP+ was used as positive control. Uptake values in WT-HEK293 were subtracted and results expressed as the mean ± S.D. from three independent experiments.
Based on the time course, the effect of a 20-minute preincubation with mβcd on OCT2-mediated MPP+ transport was assessed over a 5-second interval at an MPP+ extracellular concentration of 0.5 μM. In Fig. 3C, it can be seen that MPP+ influx was significantly lower in the mβcd-treated cells in comparison with that in the untreated cells (36.4 ± 5.9 vs. 22.2 ± 3.3 pmol mg−1 minute−1).
To rule out any direct effect of mβcd on OCT2, a cis-inhibition assay was performed. Figure 3D shows that mβcd coincubation had no effect on MPP+ OCT2-mediated influx (100.0 ± 14.9 vs. 110.8 ± 36.9 pmol mg−1 minute−1). For the cis-inhibition control, the cells were exposed to nonlabeled MPP+ at an extracellular concentration of 1 mM.
Incubation with mβcd-RAMEB Mixture Did Not Alter Total Cholesterol Levels and OCT2-Mediated MPP+ Influx.
Mβcd may have multiple nonspecific effects in addition to cholesterol depletion (Ormerod et al., 2012). To assess whether the effect of mβcd on OCT2-mediated MPP+ influx was indeed the result of cholesterol depletion, “cholesterol-matched” experiments were performed. To determine the cholesterol equilibrium point, different ratios of empty mβcd and RAMEB were tested in WT- and OCT2-HEK293 cells.
It can be seen that cells exposed for 20 minutes to a 1:1 mβcd:RAMEB mixture showed similar total cholesterol content to that of control cells (Fig. 4, A and B). Figure 4C shows that the MPP+ influx mediated by OCT2 in cells exposed to 1:1 mβcd/RAMEB was comparable to that in control cells, suggesting that the transport of MPP+ was reduced only when membrane cholesterol was depleted.

Cholesterol content and OCT2-mediated MPP+ influx after 20-minute exposure to mβcd-RAMEB mixtures. (A) Representative TLC plate of the cholesterol content in WT-HEK293 cells after incubation with the indicated ratios of mβcd and RAMEB. (B) Cholesterol content quantification. Data are relative to the cholesterol content in untreated cells and represent the mean ± S.D from three independent experiments. (C) Uptake of MPP+ (1 µM) after 20-minute incubation with mβcd or 1:1 mβcd/RAMEB. Data were corrected for uptake in WT-HEK293 cells and represent the mean ± S.D. from two independent experiments performed in duplicate.
Mβcd Did Not Alter Cell Surface Expression of OCT2.
The impact of cellular cholesterol manipulation on OCT2 protein levels and localization was assessed by Western blotting of the plasma membrane OCT2, pulled down by lysine residue labeling (Fig. 5A) and by immunofluorescence and (Fig. 5B). It can be seen that the treatment with mβcd at an extracellular concentration of 10 mM did not appear to affect the surface expression of OCT2. The selective labeling of the plasma membrane protein fraction was confirmed by the negative probing for actin (Fig. 5A).
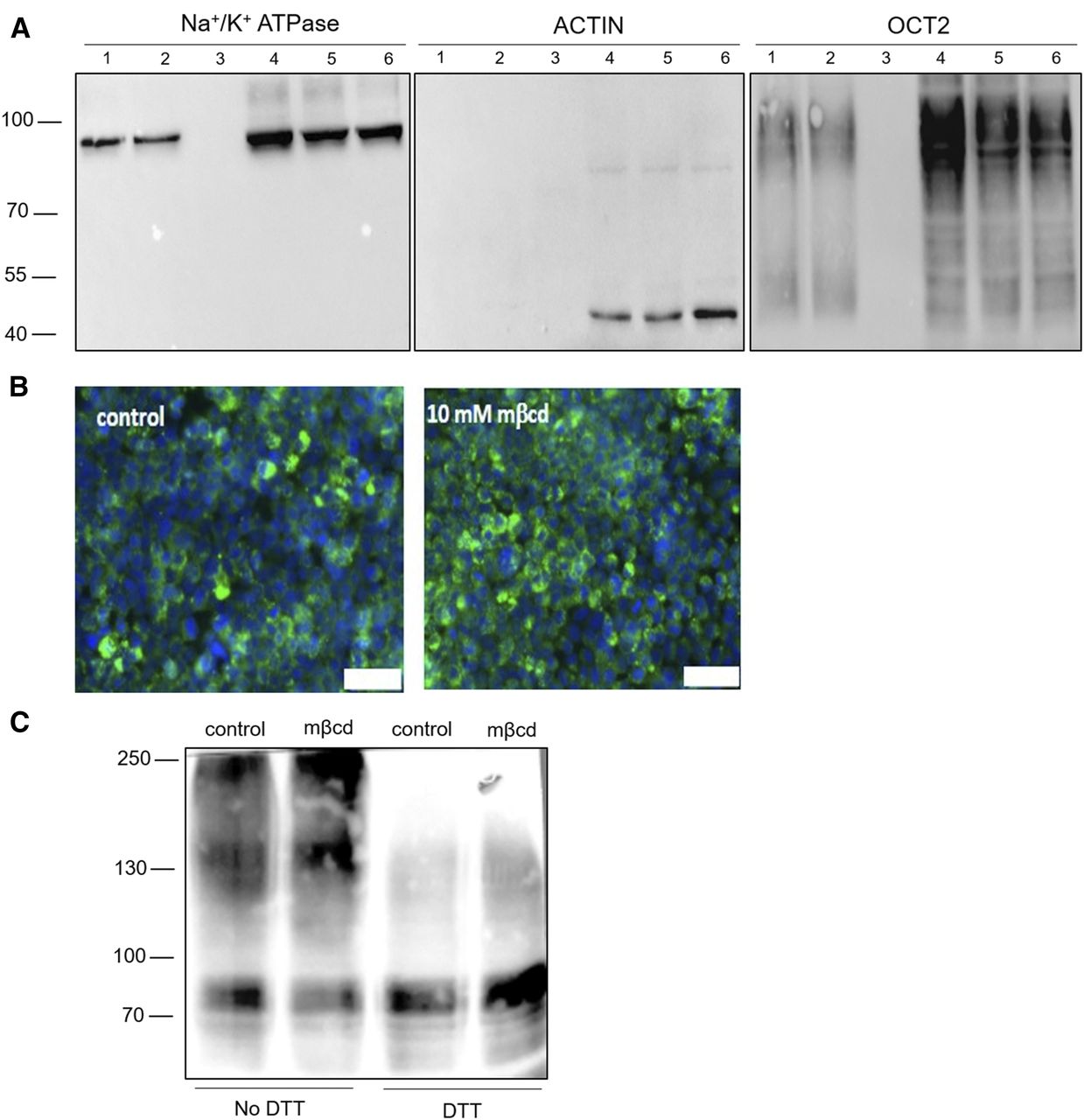
Western blotting and immunofluorescence of OCT2. Representative Western blots from three independent experiments of OCT2 expression biotinylated at the cell surface (1, 2, and 3) and in the total lysate (4, 5, and 6); actin and Na+/K+ controls are shown. Lanes 1 and 4 are loaded with samples from control OCT2-HEK293 cells, lanes 2 and 5 with samples from OCT2-HEK293 cells exposed to 10 mM mβcd for 20 minutes. (A) Samples from OCT2-HEK293 cells incubated without biotinlyation reagent are shown in lanes 3 and 6. (B) Representative immunostaining from three independent experiments of OCT2 after 20-minute incubation with mβcd. Scale bar, 100 µm. (C) Representative Western blot of cell lysates from OCT2-HEK293 treated for 20 minutes with or without mβcd and then denaturated in the presence or absence of DTT. The experiment was performed 3 times.
Mβcd Exposure Did Not Affect OCT2 Oligomerization State.
It has previously been shown that OCT2 exists in monomeric and oligomeric states and that disruption of oligomerization by DTT exposure or mutagenesis of specific cysteine residues reduced the OCT2 transport rate (Brast et al., 2012). The oligomerization state of OCT2 after mβcd treatment was assessed by Western blotting under nonreducing conditions. In Fig. 5C it can be seen that most OCT2 appears to be in the monomer state and a weaker band is observed at the size compatible with a tetramer, although the presence of heterocomplex cannot be excluded. No obvious change that could explain the dramatic effect of mβcd on OCT2-mediated MPP+ influx kinetics could be discerned.
Incubation with mβcd Shifted OCT2-Mediated MPP+ Influx Kinetics from Sigmoidal to Hyperbolic.
The kinetic basis underlying the reduced MPP+ influx upon cholesterol removal was assessed by measuring the OCT2-mediated influx (5 seconds) of MPP+ as a function of the extracellular concentration of substrate after a 20-minute preincubation with mβcd at an extracellular concentration of 10 mM. Figure 6, A–C, shows a sigmoidal and a hyperbolic curve of MPP+ OCT2-mediated influx in control and mβcd-treated cells, respectively, suggesting that the removal of cholesterol abolished the allosteric binding of MPP+ to OCT2. The switch from homotropic positive cooperativity to one-binding site kinetics was confirmed by the Eadie-Hofstee and Hill linear transformations (Fig. 6, D–F; Table 1).
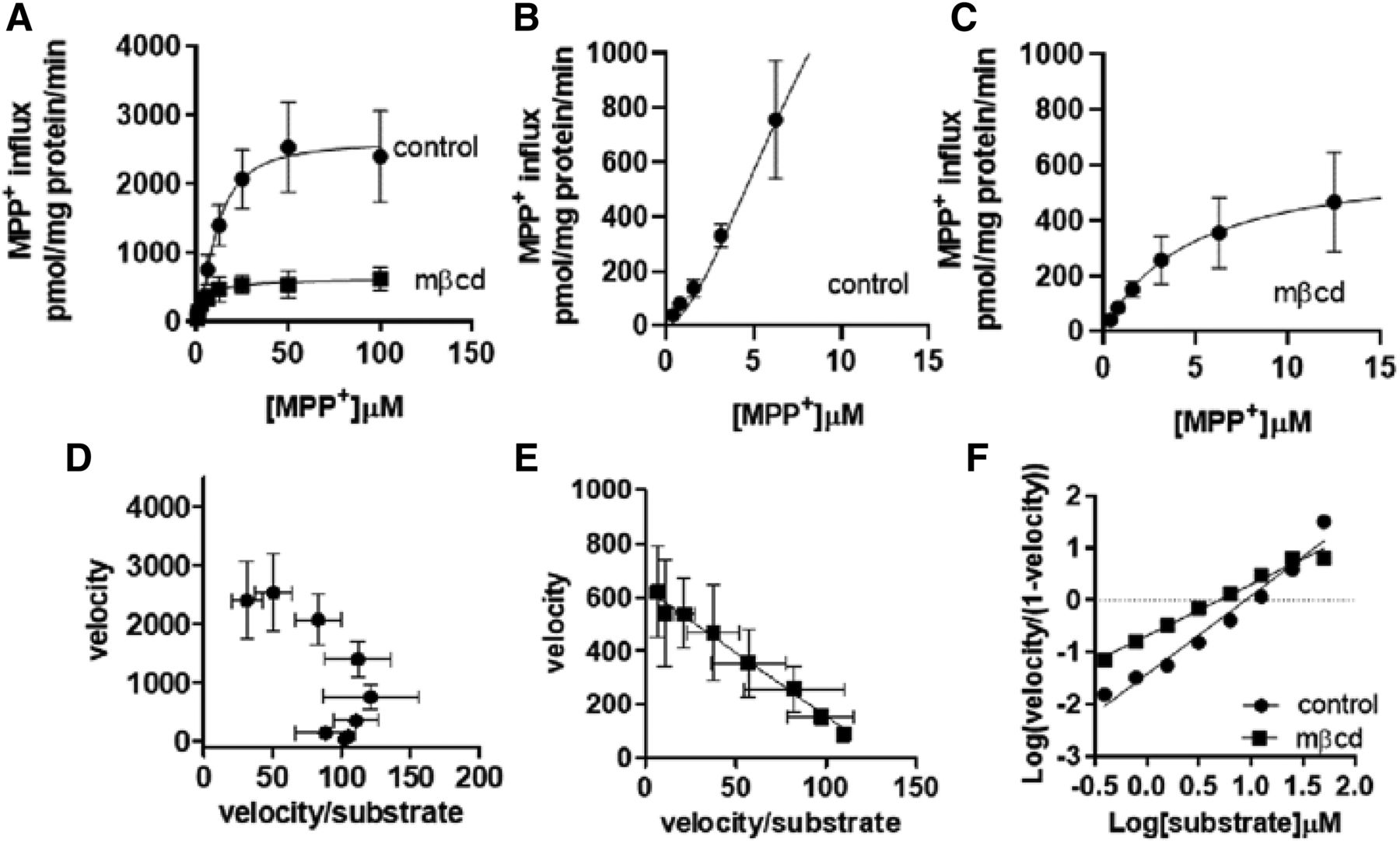
Kinetic analysis of MPP+ influx. Initial uptake of increasing extracellular concentrations of MPP+ was assessed over 5 seconds in OCT2-HEK293 cells after 20-minute incubation with mβcd. All data were corrected for uptake in WT-HEK293 cells. Line was best fit to an allosteric sigmoidal equation Y = Vmax * Xh/(Kprime + Xh), where Kprime is an estimation of Km, Vmax is the maximum capacity, and h is the Hill slope (A). Enlargement of the first part of OCT2 influx kinetics in (B) control-treated and (C) mβcd-treated cells. Eadie-Hofstee transformation in (D) control and (E) mβcd-treated cells. (F) Hill transformation. Results represent the mean ± S.D. from four independent experiments.
Kinetic parameters of MPP+ influx mediated by OCT2 after mβcd treatment
Values are mean ± S.D.
Discussion
OCT2-mediated influx kinetics that deviate from the classic hyperbole (one binding site kinetics) to the sigmoid with two cooperative binding sites are not surprising and were previously shown for gentamicin and choline by our group (Gai et al., 2016; Visentin et al., 2018). Moreover, the presence of multiple interacting binding sites has been described studying rat OCT1, whose protein sequence is 69% identical to that of human OCT2 (Gorbunov et al., 2008; Minuesa et al., 2009, 2017). Nonetheless, for the very same substrates, one binding site kinetics for the human OCT2 have been reported as well. These discordant results may stem from the varying in vitro models used in these studies: OCT2-mediated influx has been described as the classic hyperbola consistent with the Michaelis-Menten model in Xenopus laevis oocytes and in Chinese hamster ovary (CHO) cells, and as sigmoid in HEK293 cells (Gorboulev et al., 1997; Budiman et al., 2000; Gai et al., 2016; Severance et al., 2017; Visentin et al., 2018).
Here we show that after cholesterol depletion by exposure to the oligosaccharide mβcd, MPP+ uptake in HEK293 cells overexpressing OCT2 loses the allosterism and best fits the Michaelis-Menten equation. Of note, the MPP+ influx kinetics in OCT2-HEK293 cells depleted of cholesterol reconcile with previously published data using alternative models, with a calculated OCT2-mediated MPP+ influx Km (Table 1) comparable to those computed from uptake values measured in Xenopus laevis oocytes (1.2 µM) or in CHO cells overexpressing OCT2 (3.9 µM) (Gorboulev et al., 1997; Severance et al., 2017).
To clarify the contribution of cholesterol content on the discrepancies observed among experimental models, it will be interesting to perform head-to-head assessments of the lipid content and influx kinetics in different models. Cholesterol-dependent allosterism has previously been reported for other transmembrane proteins, including Mg++- and Na+/K+-ATPase, acetylcholinesterase, and the transport of estradiol-17-β-glucuronide by multidrug resistance-associated protein 2 (MRP2) (Bloj et al., 1973a,b; Guyot et al., 2014). Notably, cholesterol can either promote or inhibit allosterism, and the effect seems to be enzyme dependent. Although cholesterol did induce allosterism of OCT2 as well as of Mg++- and Na+/K+-ATPase, MRP2 and acetylcholinesterase allosteric binding was observed only in the absence of cholesterol (Bloj et al., 1973a; Guyot et al., 2014).
OCT2 has been shown to exist at the plasma membrane in monomeric and oligomeric form (Brast et al., 2012). Oligomerization is an important determinant of correct localization, function, and regulation of different transporter proteins, and membrane lipid composition can affect the oligomerization state of the transmembrane proteins embedded therein (Mörs et al., 2013; Alguel et al., 2016; Gupta et al., 2017). Because the removal of cholesterol from OCT2-HEK293 cells did not seem to alter the equilibria between the different states of oligomerization observed for this transporter, it is plausible to hypothesize that the allosterism is a feature of OCT2 binding pockets regardless of the oligomerization state.
In line with this, Keller et al. (2019) recently showed that cell-free-expressed rat OCT1 reconstituted in nanodiscs or proteoliposomes displayed one high-affinity binding site and two low-affinity binding sites for MPP+ per transporter monomer, with the latter two being responsible for the transport and the first one serving exclusively as an allosteric site. Interestingly, when the binding of the rat OCT1 monomer was assessed in nanodiscs formed with 1,2-dimyristoyl-sn-glycero-3-phospho-(1-rac-glycerol) (DMPG) or 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) and 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC), only two low-affinity binding sites with overlapping KD were detected, with the high-affinity binding site not being accessible. However, when the binding was assessed in the presence of 1,2-diacyl-sn-glycerol—phospho-l-serine, 1,2-diacyl-sn-glycero-3-phosphocholine and cholesterol, three binding sites were detected with three calculated KD values: 0.1, 10, and 36 µM, indicating that the lipid composition regulates rat OCT1 allosteric binding (Keller et al., 2019). It is possible that the human OCT2 is equipped with a high-affinity binding site that regulates the allosterism between two low-affinity binding sites as well. However, this needs to be addressed using similar synthetic approaches.
Multispecificity and allosterism are pillars of efficient detoxifying proteins, including transporters (Bodo et al., 2003; Zelcer et al., 2003; Hirono et al., 2005). While multispecificity allows recognizing a broad range of molecules, allosterism confers the ability to adapt to increasing concentrations of substrates. This is evident in Fig. 5: the loss of allosterism upon cholesterol depletion led to a rapid saturation of the OCT2 transport molecules and, in turn, to a limited transport capacity. Because OCT2 plays a key role in the clearance of many drugs and toxins, it is conceivable that pathophysiologic changes in renal membrane cholesterol content may shift the OCT2 kinetics from hyperbolic to sigmoidal, or vice versa.
It has been previously shown that OCT2 expression varies in several pathophysiologic conditions. Rats with acute kidney injury showed lower Oct2 levels (Ji et al., 2002; Matsuzaki et al., 2008; Shim et al., 2009; Maeng et al., 2012). Obesity, both in human and in mice, induces OCT2 expression (Gai et al., 2016). Renal cell carcinoma cells have low OCT2 levels as compared with the surrounding normal tissue (Liu et al., 2016; Visentin et al., 2018). This work adds another level of complexity in the study of OCT2-mediated tubular secretion in renal pathology.
Acknowledgments
The authors would like to thank Hermann Koepsell, University of Wurzburg, for providing OCT2-HEK293 cells.
Authorship Contributions
Participated in research design: Hörmann, Kullak-Ublick, Visentin.
Conducted experiments: Hörmann, Gai, Visentin.
Performed data analysis: Hörmann, Visentin.
Wrote or contributed to the writing of the manuscript: Hörmann, Visentin.
Footnotes
- Received June 20, 2019.
- Accepted October 11, 2019.
This work was supported by the Swiss National Science Foundation [Grant 310030_175639] (to G.A.K.-U.).
Abbreviations
- CHO
- Chinese hamster ovary
- DMEM
- Dulbecco’s modified Eagle’s medium
- DMPG
- 1,2-dimyristoyl-sn-glycero-3-phospho-(1-rac-glycerol)
- DPPC
- 1,2-dipalmitoyl-sn-glycero-3-phosphocholine
- DTT
- dithiothreitol
- HEK293
- human embryonic kidney 293 cells
- mβcd
- methyl-β-cyclodextrin
- MPP+
- N-methyl-4-phenylpyridinium acetate
- MRP2
- multidrug resistance-associated protein 2
- OCT
- organic cation transporter
- POPC
- 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine
- RAMEB
- cholesterol-presaturated mβcd
- TEA
- tetraethanolamine bromide
- TLC
- thin layer chromatography
- WT
- wild type
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}