


Abstract
Previous research has demonstrated that activity of glycogen synthase kinase-3 (GSK3) is necessary for the rewarding effects of cocaine. In the present study, a conditional GSK3β gene knockdown model was used to determine if GSK3β activity specifically in the nucleus accumbens is important for cocaine conditioned reward. The roles of accumbal GSK3β in morphine conditioned reward, trans-(±)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide methanesulfonate salt (U50,488H)-induced conditioned place aversion, and cognitive function were also studied. Adult male and female GSK3β-floxed or wild-type mice were injected with adeno-associated virus/Cre into the nucleus accumbens to reduce expression of GSK3β and underwent behavioral testing 4 weeks later. The development of cocaine-induced conditioned place preference was significantly attenuated in mice with reduced levels of GSK3β in the nucleus accumbens, whereas the development of morphine-induced place preference remained intact. Conditional knockdown of GSK3β in the accumbens prevented the development of conditioned aversion produced by U50,488H, a κ-opioid receptor agonist. Cognitive memory tests revealed deficits in object location memory, but not novel object recognition in mice with accumbal GSK3β knockdown. These data demonstrate that GSK3β in the nucleus accumbens is required for cocaine conditioned place preference and U50,488H conditioned place aversion, as well as spatial memory in object location task, indicating differential roles of GSK3β in the psychostimulant and opiate reward process, as well as in memory for spatial locations and object identity.
SIGNIFICANCE STATEMENT Knockdown of GSK3β in the nucleus accumbens attenuated the development of cocaine-induced place preference, as well as conditioned place aversion to U50,488H, a κ-opioid receptor agonist. In contrast, the development of morphine place preference was not altered by GSK3β knockdown. GSK3β knockdown in nucleus accumbens impaired performance in the object location task, but not the novel object recognition task. These results elucidate different physiological roles of accumbal GSKβ in conditioned reward, aversion, and memory.
Introduction
The importance of glycogen synthase kinase-3 (GSK3) function in the pathology of neuropsychiatric disorders including schizophrenia, bipolar disorder, and drug addiction has been recognized (Grimes and Jope, 2001; Gould et al., 2004; Lovestone et al., 2007; Perrine et al., 2008). Two isoforms of GSK3, GSK3α and GSK3β, exist. GSK3β is the major isoform in the brain and is highly abundant in the hippocampus, cortex, and striatum (Yao et al., 2002; Jope and Johnson, 2004). GSK3β activity is tightly regulated, with the major regulatory mechanism being the phosphorylation of an N-terminal serine, Ser9-GSK3β. GSK3β is a constitutively active kinase, and kinase activity is inhibited by phosphorylation of Ser9-GSK3β (Jope and Johnson, 2004; Kockeritz et al., 2006).
GSK3β is an important mediator of dopaminergic neurotransmission and the expression of dopamine-associated behaviors (Beaulieu et al., 2005, 2011). For example, elevated synaptic dopamine results in activation of GSK3β in the striatum of mice with genetic deletion of the dopamine transporter, and GSK3 inhibition reduces the hyperactive phenotype of these mice (Beaulieu et al., 2004, 2005). Similarly, previous reports from our laboratory have demonstrated that GSK3β activity in the nucleus accumbens is enhanced after acute cocaine exposure (Perrine et al., 2008; Miller et al., 2014), and dopaminergic D1 and D2 receptors as well as glutamatergic N-methyl-D-aspartate receptors contribute to the activation of GSK3β by cocaine (Miller et al., 2014). Activation of GSK3 is required for the rewarding effects of cocaine. Systemic administration of GSK3 inhibitors prior to cocaine or amphetamine conditioning blocks the development of place preference (Miller et al., 2014; Wickens et al., 2017). Although the requirement of GSK3 activity for the rewarding effects of psychostimulants has been established, the specific brain region(s) and GSK3 isoform contributing to this mechanism has yet to be determined. Furthermore, there are no reports indicating whether GSK3β is necessary for the conditioned rewarding or aversive effects of opioids.
Current pharmacological approaches cannot differentiate between GSK3α and β isoforms because the inhibitors, while selective for GSK3, are not selective for the individual isoforms (Coghlan et al., 2000). GSK3β knockout mice are embryonically lethal, and global heterozygous GSK3β knockout induces long-term spatial memory impairment (Kimura et al., 2008). Because of the limitations to these other approaches, the present study used a conditional knockdown approach to specifically study GSK3β in the nucleus accumbens of adult female and male mice. Using this genetic conditional knockdown model, the role of accumbal GSK3β in cocaine reward was investigated. This study also determined the importance of GSK3β in morphine conditioned reward, conditioned aversion produced by trans-(±)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide methanesulfonate salt (U50,488H, a κ-opioid receptor agonist), and memory for novel objects and object location. The results presented herein indicate that GSK3β in the nucleus accumbens had different roles in cocaine and morphine conditioned reward. Unexpectedly, GSK3β in the nucleus accumbens also contributed to conditioned place aversion produced by a κ-opioid receptor agonist and to object spatial memory.
Materials and Methods
Floxed GSK3β Mice
Floxed (flx) GSK3β flx/flx mice were generously provided by Dr. James Woodgett (Lunenfeld-Tanenbaum Research Institute, Mount Sinai Hospital, Toronto, Canada) (Kockeritz et al., 2006; Patel et al., 2008). Breeding and genotyping were carried out in core resources supported by our Center on Intersystem Regulation by Drugs of Abuse (under Grant P30 DA013429, to Ellen M. Unterwald, principal investigator). Wild-type (WT) littermates were used as controls. After weaning, mice were housed in single sex groups of four or five per cage without additional enrichment objects. Food and water were available ad libitum and mice were maintained on a 12-hour light/dark cycle (7:00 AM/7:00 PM). Mice were handled and weighed daily for 5 days before beginning behavioral testing. Behavioral testing occurred between 9:00 AM and 5:00 PM. Animal use was conducted with an approved protocol from the Temple University Institutional Animal Care and Use Committee and was consistent with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals.
Delivery of Adeno-Associated Virus into the Nucleus Accumbens.
Mice between the ages of 8–10 weeks were placed within a stereotaxic apparatus (Stoelting Co., Wood Dale, IL) while under isoflurane anesthesia. The stereotaxic coordinates used for viral infusion into the nucleus accumbens were: anterior-posterior +1.4 mm anterior and ±0.6 mm lateral from bregma and −4.2 mm ventral (Paxinos and Franklin, 2001). A 26-gauge syringe (Hamilton Company, Reno, NV) was inserted through the incision site and delivered 0.5 µl of virus [adeno-associated virus (AAV) 1-Cre-Green Fluorescent Protein (GFP) or AAV-GFP, cytomegalovirus promotor, 1.48+13 VG/ml; Signagen Laboratories, Rockville, MD] at 0.1 µl/min with an infusion pump (Harvard Apparatus, Holliston, MA). After 5 minutes, the syringe needle was withdrawn and the infusion was repeated in the opposite hemisphere. Behavioral testing began 4 weeks following viral delivery. The GFP fluorescence was used to assess the accuracy of the bilateral nucleus accumbens infusions. The data from mice with one or both missed placements were removed from the analysis.
Drugs
Cocaine hydrochloride, morphine sulfate, and U50,488H were generously supplied by the National Institute on Drug Abuse’s drug supply program, dissolved in sterile saline, and injected in a volume of 3 ml/kg (cocaine, i.p.) or 5 ml/kg (morphine, s.c.) body weight. An equal volume of saline served as the vehicle control.
Experimental Procedures
Conditioned Place Preference and Aversion.
The procedures for conditioned place preference and conditioned place aversion were similar to those described in our prior publication (Miller et al., 2014) and by Liu et al. (2019). Place conditioning occurred in rectangular plastic chambers (45 × 20 × 20 cm) consisting of two unique compartments, one with white and black vertical striped walls and smooth flooring and the other with white walls with black circles and rough flooring. The two compartments were separated by a removable wall/door. Illumination in both compartments was equal and mice had no initial preference to either compartment. As such, a randomized, counterbalanced, unbiased conditioning procedure was used.
Cocaine-induced conditioned place preference.
Mice were injected with cocaine (10 mg/kg, i.p.) or saline, and immediately confined to one compartment of the conditioning chamber where they remained for 30 minutes. Conditioning occurred once per day for four consecutive days, resulting in two conditioning sessions with saline in one compartment and two sessions with cocaine in the opposite compartment of the conditioning chamber. The test for place preference occurred on day 5, when mice had access to both compartments for 30 minutes in a drug-free state. The time spent in each compartment was recorded. Preference scores (reported in seconds) were calculated as follows: (time spent in the cocaine-paired compartment) minus (time spent in the saline-paired compartment).
Morphine-induced conditioned place preference.
Mice were injected with saline or morphine (5 mg/kg, s.c.) and placed into one of the two compartments for 40 minutes. Conditioning occurred two times each day (9:00 AM and 3:00 PM) for 4 days, resulting in four place pairings with each morphine and saline. The test for place preference occurred on day 5, when mice were placed in the chamber for 20 minutes and allowed to explore both compartments. The amount of time spent in each compartment was measured, and preference scores were calculated as described previously.
U50,488H-induced conditioned place aversion.
The procedure was performed as reported by Liu et al. (2019) with slight modification. Mice remained in their home cage for 15 minutes after receiving an injection of saline or U50,488H (2.5 mg/kg, s.c.), after which they were placed into one of the two compartments of the conditioning chamber for 30 minutes. Mice were conditioned twice daily, once with U50,488H and once with saline (9:00 AM and 3:00 PM) for 4 days. Place aversion was tested in a drug-free state on day 5, when mice were allowed to freely move about both compartments for 15 minutes. The time spent in each compartment was recorded and a place aversion score was calculated as follows: (time in U50,488H-paired compartment) minus (time in saline-paired compartment).
Novel object recognition test.
This test was used to evaluate memory function and was performed as reported by Silverman et al. (2013). The experiment consisted of three phases: habituation, familiarization, and test phase. On the first day (day 1, habituation phase), each mouse was placed into an empty open-field chamber for 30 minutes without objects. The open-field apparatus is a plastic chamber (25 × 20.5 × 30 cm) with a smooth floor and four nontransparent red walls. On the second day (day 2, familiarization phase), each mouse was placed individually into the open-field chamber for 10 minutes, in which the chamber contained two identical sample objects (A + A). After the familiarization session, mice were placed in a clean version of their home cage with bedding only (no enrichment), one mouse per cage. The open-field arena and objects were cleaned thoroughly with 70% ethanol after each session. One familiar object and one novel object (A+ B) were placed in the arena in the same location where the two identical objects were located during the familiarization phase. After a 30-minute delay at the end of the familiarization session, the recognition test occurred. Individual mice were place back into the open-field arena with the familiar and novel objects for 10 minutes. The familiarization session and the recognition test were recorded by an overhead camera for subsequent analysis. Object investigation was defined as time spent sniffing the object with the nose pointed toward the object at a distance ≤1 cm. Exploration of the two objects (in seconds) during training and test sessions was measured by an experimenter blinded to experimental group. Recognition memory was defined as significantly more time spent investigating the novel versus the familiar object. A discrimination index (DI) was defined as follows: (time exploring the novel object − time exploring the familiar object)/[(time exploring both objects) × 100].
Object location memory test.
The procedures for the object location test were the same as the novel object recognition test described previously with the following exception: identical copies of the sample objects were presented during the test phase but one of the two objects was placed in a new location. A DI was calculated as follows: (time exploring the novel location of the object − time exploring the familiar location of the object)/[(time exploring both objects) × 100]. The same mice were used in both memory tasks with 1 week separating the two tests. Unique sets of objects were used in the two tasks.
Immunofluorescence.
Immunohistochemistry with dual fluorescent labeling was used to measure knockdown of GSK3β and confirm injection site. After behavioral testing was completed, anesthetized mice were perfused with 4% paraformaldehyde and brains were postfixed for 18 hours. Vibratome sections (40 µm) spanning the entire nucleus accumbens were obtained and incubated in goat anti-GFP (1:500, ab6673; Abcam) and rabbit anti-GSK3β (1:1000, D5C5Z XP mAb #12456; Cell Signaling Technology) primary antibodies at 4°C for 24 hours. After rinsing the sections in 0.1 M PBS, they were incubated with donkey anti-goat (1:500, Alexa Fluor 488; Invitrogen) and donkey anti-rabbit (1:500, Alexa Fluor 594; Invitrogen) secondary antibodies at room temperature for 2 hours. Sections from WT and GSK3β floxed mice were processed at the same time. Determination of the spatial distribution of AAV-infected nucleus accumbens neurons was made by visualization of labeling. To measure the changes in GSK3β immunoreactivity, brain sections were imaged using a Nikon epifluorescent microscope with an analog camera and immunoreactive product measured using a bioquantification software system (TCW 98; Bioquant, Nashville, TN), as previously described by our group (Miller et al., 2014). Briefly, WT mice were used as control to define threshold of staining. Threshold values were determined based on mean levels of GSK3β immunostaining. The threshold values were used for consistent auto-measurement of all brain sections. The level of GSK3β-immunostaining was calculated by dividing the area containing pixels at or above the defined background threshold by the area of the total number of pixels in the chosen field, which was multiplied by 100 to yield the percentage of staining.
Time Line of Experimental Procedures.
Figure 1 outlines the timing of the experimental procedures and treatment groups.
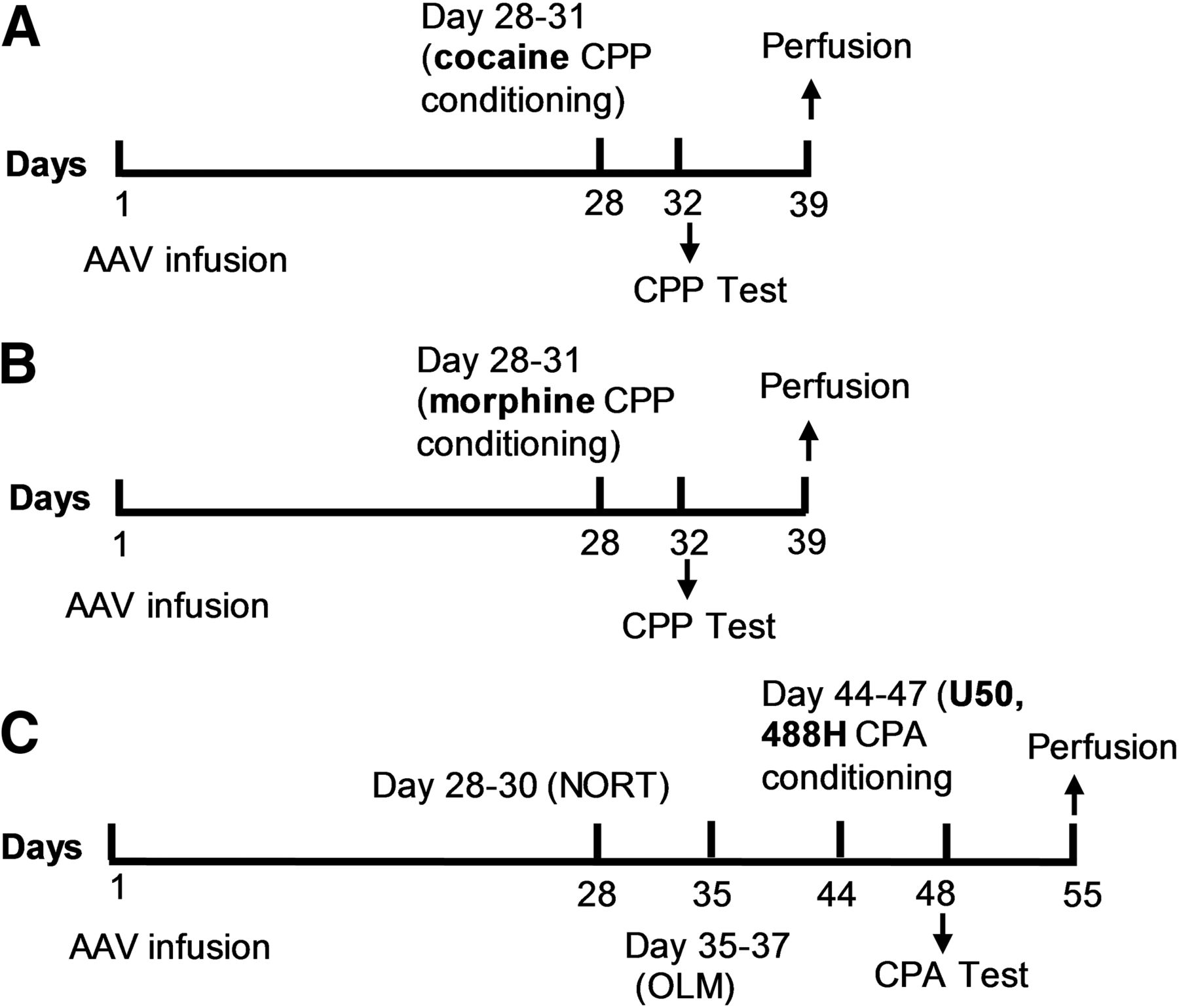
Time lines of experimental procedures. (A) WT and GSK3β flx/flx mice were injected with AAV-Cre-GFP, and GSK3β flx/flx mice were injected with AAV-GFP bilaterally into the nucleus accumbens on day 1. Mice underwent place conditioning with cocaine (10 mg/kg, i.p.) using a 4-day conditioning procedure. On day 5 (day 32), preference scores were determined during a 30-minute test session. (B) WT and GSK3β flx/flx mice were injected with AAV-Cre-GFP bilaterally into the nucleus accumbens on day 1. Mice underwent place conditioning with morphine (5 or 10 mg/kg, s.c.) using a 4-day conditioning procedure. On test day (day 32), preference scores were determined during a 20-minute test session. (C) WT and GSK3β flx/flx mice were injected with AAV-Cre-GFP bilaterally into the nucleus accumbens on day 1. Novel-object recognition (NORT) and object location memory (OLM) tests were carried out with a 1-week interval between each of the tests. Mice underwent place conditioning with U50,488H (2.5 mg/kg, s.c.) using a 4-day conditioning procedure. On test day (day 48), preference scores were determined during a 15-minute test session. Following behavioral tests, all mouse brains were analyzed for efficiency of GSK3β knockdown.
Data Analysis
Conditioned place preference and aversion data were analyzed by one-way ANOVA (for three groups) or two-tailed t test (for two groups). The relationships between the extent of GSK3β knockdown in nucleus accumbens and place preference score were analyzed using correlation analysis (Pearson’s test). Novel object recognition and object location tasks were analyzed by two-way ANOVA with object (or location) and genotype factors followed by Bonferroni’s test for multiple comparisons. Discrimination index data were analyzed by one-way ANOVA followed by Bonferroni post tests. Two-way ANOVAs with main factors of genotype and sex were used to assess potential sex differences. GraphPad Prism 4 (La Jolla, CA) was used for statistical analyses.
Results
Verification of GSK-3β Knockdown in the Nucleus Accumbens.
Brains were obtained 5–7 weeks after AAV injection, following behavioral testing, for immunohistochemical analysis of GSK3β levels and confirmation of viral injection location. Three animals were removed from the study due to missed placements. Figure 2 shows representative images of immunostaining against GSK3β in the accumbens from WT (Fig. 2A) and GSK3β flx/flx (Fig. 2B) mice. As can be seen in the magnified images (40×) in Fig. 2, cells in GSK3β flx/flx mice infected with AAV-Cre-GFP (Fig. 2B) had no GSK3β fluorescence in the cytoplasm compared with cells expressing GFP in WT mice (Fig. 2A). Quantification of GSK3β immunoreactivity revealed significantly lower levels of GSK3β in the nucleus accumbens of GSK3β flx/flx mice injected with AAV-Cre-GFP compared with WT controls injected with the same vector or GSK3β flx/flx mice injected with AAV-GFP (F2,85 = 166.3, P < 0.0001) (Fig. 2C). Immunofluorescence analysis of accumbens injected with AAV-Cre-GFP showed a mean 65.85% (51%–77%) reduction in GSK3β-IR in the GSK3β flx/flx mice compared with GSK3β WT littermates (Fig. 2C). These data demonstrate effective knockdown of GSK3β in the nucleus accumbens of GSK3β flx/flx mice by AAV-Cre recombinase delivery.

Representative images show immunostaining of GFP (green), GSK3β (red), and 4,6-diamidino-2-phenylindole dihydrochloride (DAPI) (blue) in the nucleus accumbens of WT (A) and GSK3β flx/flx (B) mice. Immunostaining demonstrates proper targeting of the nucleus accumbens and reductions in the levels of GSK3β protein in GSK3β flx/flx mice (B, inset). (A) GFP is expressed in cells of the WT accumbens, along with GSK3β (10×). Higher magnification (40×) shows overlap of GFP and DAPI in an individual cell with intense GSK3β staining in the surrounding cytoplasm. (B) GFP is expressed in cells of the GSK3β flx/flx accumbens (10×). Cells that express GFP do not express GSK3β as shown in higher magnification (40×). (C) Quantification of GSK3β fluorescence in the nucleus accumbens of WT and GSK3β flx/flx mice injected with AAV-Cre-GFP (flx/flx+) or AAV-GFP (flx/flx−) shows significant reduction of GSK3β following AAV-Cre-GFP injection into the accumbens of GSK3β flx/flx mice. Data are represented as mean ± S.E.M. N = 20–33/group; AC, anterior commissure.
GSK3β Knockdown in the Nucleus Accumbens Attenuated the Development of Cocaine-Induced Conditioned Place Preference.
The role of GSK3β in the nucleus accumbens in cocaine conditioned reward was evaluated in conditional GSK3β knockout mice and controls. Four weeks following AAV-Cre injection, when Cre recombination is maximal (Kaspar et al., 2002; Ahmed et al., 2004), cocaine-induced conditioned place preference was performed. One-way ANOVA of preference score revealed a significant difference among the three groups (F2,34 = 12.64, P < 0.0001) (Fig. 3A). Post-hoc analysis showed that GSK3β flx/flx mice injected with AAV-Cre-GFP into the accumbens had significantly lower preference scores compared with WT mice injected with AAV-Cre-GFP (P < 0.001) and GSK3β flx/flx mice injected with AAV-GFP (P < 0.01). Thus, knockdown of GSK3β in the nucleus accumbens significantly impaired the development of cocaine-induced place preference. In Fig. 3B, the correlation between preference score and accumbal GSK3β intensity for individual mice is shown. There was a significant positive correlation between preference scores and the intensity of GSK3β in the nucleus accumbens (r2 = 0.6353, P = 0.0011), showing that mice with the lowest levels of GSK3β had the lowest preference scores. This finding further supports the contention that activation of GSK3β in the accumbens contributes to cocaine reward. Analysis of the data for potential sex differences (two-way ANOVA with sex and genotype factors) did not reveal a significant difference between males and females, albeit the study may have been underpowered to detect sex differences.

GSK3β knockdown in the nucleus accumbens attenuates the rewarding effect of cocaine. (A) Conditioned place preference scores are shown for WT mice injected with AAV-Cre, GSK3β flx/flx with AAV-GFP, and GSK3β flx/flx with AAV-Cre conditioned with 10 mg/kg of cocaine. Knockdown of GSK3β in the GSK3β flx/flx + AAV-Cre mice resulted in significantly lower preference scores. Data are expressed as mean + S.E.M. (B) Cocaine preference scores and GSK3β intensity for individual GSK3β flx/flx+ mice are shown. Pearson correlation analysis showed a significant positive correlation of preference score and GSK3β intensity in nucleus accumbens of GSK3β flx/flx+ mice (r2 = 0.6353, P = 0.0011). ***P < 0.001, GSK3β flx/flx-AAV-Cre vs. WT mice; ##P < 0.01, GSK3β flx/flx-AAV-Cre vs. GSK3β flx/flx-AAV-GFP. N = 11–13/group as indicated in the figure; F, female; M, male.
GSK3β Knockdown in the Nucleus Accumbens Had No Effect on the Development of Morphine-Induced Conditioned Place Preference.
The requirement of accumbal GSK3β in morphine conditioned reward was tested to investigate the generalizability of the aforementioned findings to another class of drugs. Three weeks after injection of AAV-Cre-GFP into the nucleus accumbens, mice underwent 4-day morphine (5 mg/kg) conditioning followed by a test for place preference. As shown in Fig. 4A, no significant difference was found in preference scores between GSK3β flx/flx and WT mice with AAV-Cre injections into the accumbens (t = 1.01, df = 38, P > 0.05). No significant correlation was found between morphine place preference scores and accumbal GSK3β intensity (r2 = 0.0039, P = 0.80). To further investigate a potential role of accumbal GSK3β in morphine conditioned reward, another dose of morphine was tested, 10 mg/kg. As shown in Fig. 4B, no significant difference was found in preference scores between GSK3β flx/flx and WT mice with AAV-Cre injections into the accumbens. No significant differences in preference scores were found between males and females of either genotype at any dose of morphine. These data suggest that GSK3β activation in the nucleus accumbens is not required for morphine conditioned reward.

Morphine conditioned place preference was not changed by GSK3β knockdown in the nucleus accumbens. Conditioned place preference was established with 5 mg/kg (A) or 10 mg/kg (B) of morphine in WT mice and GSK3β flx/flx. No significant differences in preference scores were found between the two groups at either dose. Data are expressed as mean + S.E.M. N = 18–22/group.
GSK3β Knockdown in the Nucleus Accumbens Attenuated the Development of U50,488H-Induced Conditioned Place Aversion.
Place conditioning can assess not only the rewarding but also the aversive effects of drugs. Conditioned place aversion to U50,488H, a κ-opioid receptor agonist, was performed in GSK3β flx/flx and WT mice 4 weeks after the injection of AAV-Cre-GFP in the nucleus accumbens. As shown in Fig. 5, one-way ANOVA of preferences scores revealed a significant difference between groups (F2,27 = 6.06, P < 0.001). Post-hoc analysis demonstrated that GSK3β flx/flx mice with intra-accumbens injection of AAV-Cre showed significantly less place aversion compared with the WT mice with AAV-Cre (P < 0.05) or GSK3β flx/flx mice with control vector injection (P < 0.05). No significant differences were observed between males and females in preference scores. There was a significant negative correlation between preference scores and the intensity of GSK3β in the nucleus accumbens (r2 = 0.48, P = 0.03), indicating that mice with the lowest levels of GSK3β had the least place aversion. These data indicate that GSK3β in nucleus accumbens is required for the development of U50,488H-induced place aversion.
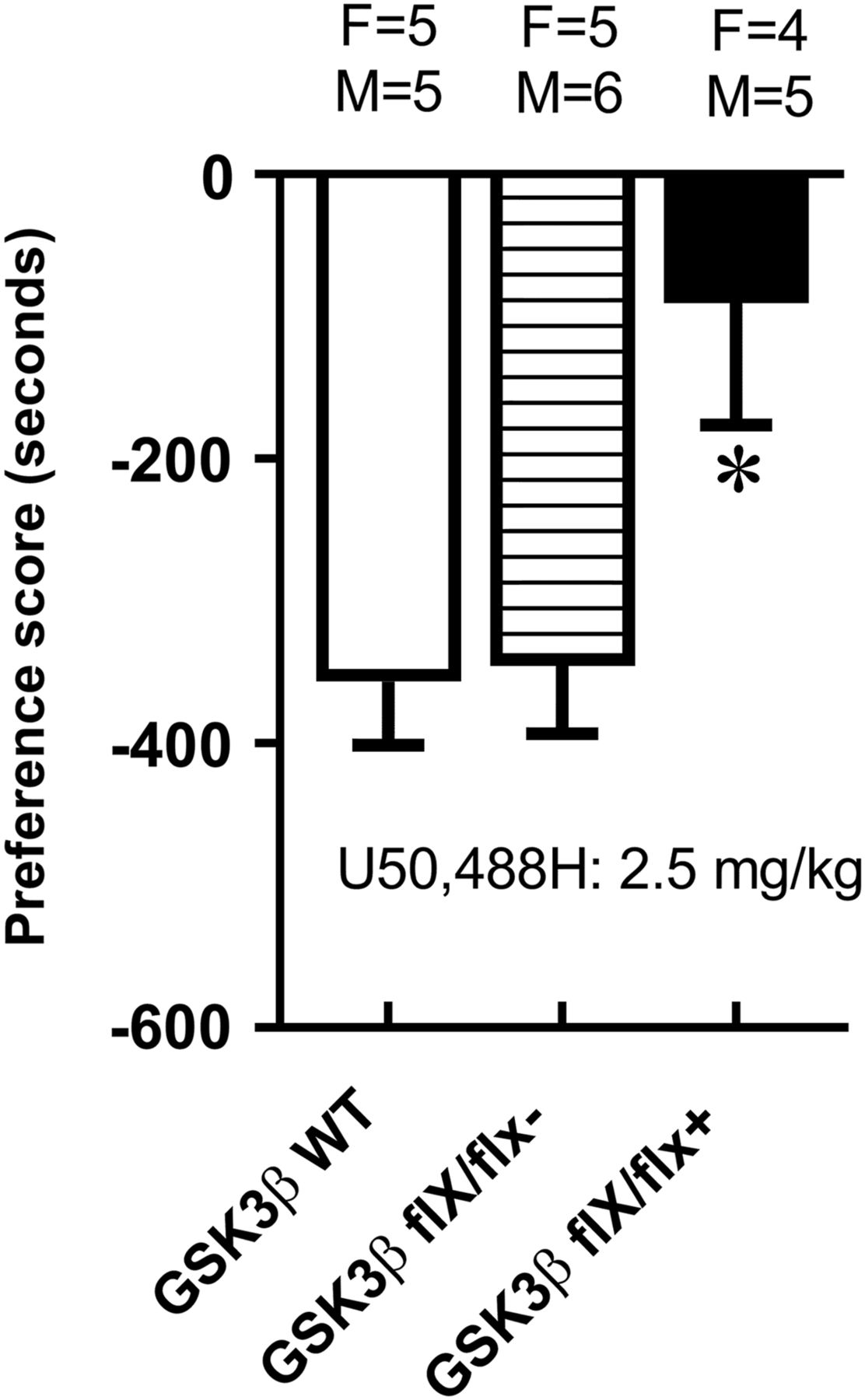
GSK3β knockdown in nucleus accumbens attenuated the aversive effect of U50,488H. Conditioned place aversion was established with 2.5 mg/kg of U50,488H. Place aversion scores were significantly reduced in GSK3β flx/flx with AAV-Cre vs. WT mice with AAV-Cre. Data are expressed as mean + S.E.M. *P < 0.05, GSK3β flx/flx-AAV-Cre vs. WT mice. N = 9–11/group.
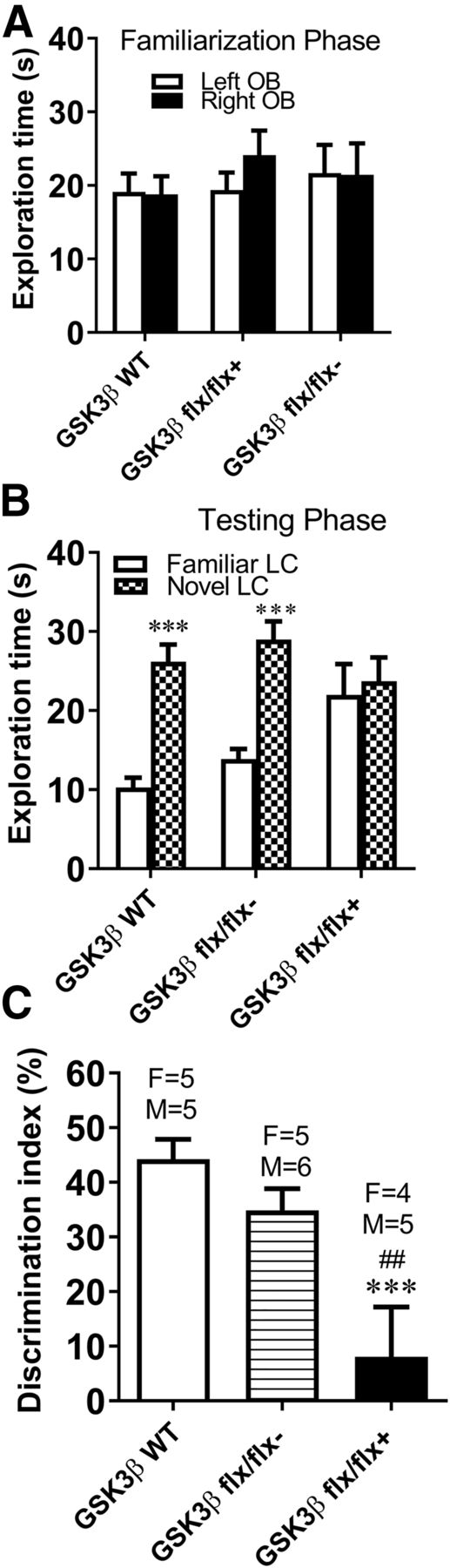
GSK3β Knockdown in the Nucleus Accumbens Did Not Impair Performance in the Novel Object Recognition Task.
The novel object recognition task was used to investigate the role of GSK3β in cognitive function. As shown in Fig. 6A, no preference for either object was found for any of the experimental groups during the 10-minute familiarization session (P > 0.05). Thirty minutes after the familiarization training, mice underwent a 10-minute recognition test. Two-way ANOVA of exploration time during the test session revealed a significant main effect of object (F1,54 = 69.4, P < 0.0001) and no significant main effect of genotype (F2,54 = 0.3, P > 0.05) (Fig. 6B). All three experimental groups showed a significant preference for exploring the novel object as shown by the discrimination index (P < 0.001) (Fig. 6C). Consistent with the strong preference for novel relative to familiar objects, the DI of the three groups was 45.97 ± 3.72 for WT with AAV-Cre, 36.30 ± 2.91 for GSK3β flx/flx with AAV-GFP, and 38.65 ± 4.44 for GSK3β flx/flx with AAV-Cre. No group differences in total exploration times during training or testing were found (data not shown). Likewise, there were no significant differences between males and females in their performance on the novel object recognition task. No significant correlation was observed between DI and accumbal GSK3β intensity (r2 = 0.0082, P = 0.45). These data show that novel object recognition remains intact in mice with knockdown of GSK3β in the nucleus accumbens.

Novel object recognition task remains intact in mice with GSK-3β knockdown in nucleus accumbens. WT, GSK3β flx/flx with AAV-Cre, and GSK3β flx/flx with AAV-GFP mice underwent novel object recognition testing. (A) There was no innate preference for either object during the familiarization phase. (B) All groups explored the novel object more than the familiar object during the testing phase. (C) No significant differences in the DI were found. Data are expressed as mean + S.E.M. ***P < 0.001 vs. familiar object (OB). N = 9–11.
GSK3β Knockdown in the Nucleus Accumbens Impaired Performance in the Novel Location Task.
Mice underwent the object location task, whereby they were trained with two identical objects for in the familiarization session. Thirty minutes following the familiarization session, mice were placed in the same arena with one object located in a new position. As shown in Fig. 7A, during the familiarization session, no preference for the object position was found in WT mice (P > 0.05), GSK3β flx/flx mice with intra-accumbens injection of AAV-Cre (P > 0.05), or GSK3β flx/flx mice with AAV-GFP infusion (P > 0.05). During the test phase (Fig. 7B), two-way ANOVA of exploration time revealed a significant main effect of location (F1,54 = 31.2, P < 0.001) and significant location × genotype interaction (F2, 54 = 5.3, P < 0.01), but no significant main effect of genotype (F1,54 = 1.9, P > 0.05). Post-hoc analysis showed that WT mice spent more time with the object in the novel location (P < 0.001), thus exhibiting memory for the familiar object location, as did GSK3β flx/flx mice with control vector infusion (P < 0.001). In contrast, time spent in exploring the objects did not differ between the familiar and novel locations in GSK3β flx/flx mice with knockdown of GSK3β in the accumbens. The DI was significantly lower in mice with accumbal GSK3β knockdown compared with control WT mice (P < 0.001) or control GSK3β flx/flx mice (P < 0.01), as shown in Fig. 7C. No differences in total exploration times between groups during training or testing were found. Analysis of males and females separately by two-way ANOVA with sex and genotype factors revealed no significant sex differences. There was no significant correlation between the DI and accumbal GSK3β intensity (r2 = 0.12, P = 0.35). These data demonstrate that GSK3β knockdown in nucleus accumbens produced a cognitive deficit in location memory in male and female mice.
Performance in the novel location task was impaired in mice with conditional knockdown of GSK3β in the nucleus accumbens. (A) No innate preference for object position was found in the three groups of mice during the familiarization phase. (B) WT mice with AAV-Cre and GSK3β flx/flx mice with AAV-GFP, but not the mice with GSK3β knockdown in the nucleus accumbens, spent more time exploring the object in a new location than the prior location. ***P < 0.001 vs. familiar object (OB). (C) GSK3β knockdown significantly reduced the DI compared with controls, indicating impairment in special memory. ***P < 0.001 vs. WT mice with AAV-Cre; ##P < 0.01 vs. GSK3β flx/flx mice with AAV-GFP. Data are expressed as mean + S.E.M. N = 9–11/group.
Discussion
This study elucidated the role of GSK3β specifically within the nucleus accumbens in drug reward and aversion and object and spatial memory tasks. Prior work using systemically administered GSK3 inhibitors has demonstrated the importance of GSK3 in drug reward and memory (Wu et al., 2011; Miller et al., 2014; Shi et al., 2014; Wickens et al., 2017). GSK3β is widely expressed throughout the brain, but the region(s) where GSK3β influences these functions is largely unknown. Knockdown of accumbal GSK3β in floxed mice impaired their ability to develop a cocaine-induced place preference. Further analysis revealed a significant positive correlation between nucleus accumbens GSK3β levels and cocaine preference scores. Mice with low GSK3β (<40% of control) showed no preference for the cocaine-paired environment, supporting the importance of GSK3β specifically in the nucleus accumbens in mediating cocaine-induced place preference. In contrast, mice with reduced accumbens GSK3β showed a significant preference for the morphine-paired environment similar to WT mice. The ability of the GSK3β knockdown mice to develop and express a morphine place preference supports the contention that cocaine reward processes were affected by the knockdown rather than an inability of the mice to learn or remember the place conditioning task. Thus, GSK3β signaling in the accumbens is a critical component of positive rewarding actions of cocaine, but is not necessary for morphine conditioned reward.
Dopaminergic neurons of the ventral tegmental area (VTA) projecting to the nucleus accumbens comprise the primary pathway mediating the reinforcing effects of drugs of abuse and natural rewards (Wise, 1996; Koob and Le Moal, 1997). Despite its central role in reward processes, actions of opiates versus stimulants on this circuit differ (Koo et al., 2012) . For example, opiates and stimulants regulate structural plasticity in opposing manners. Opiates decrease the number and complexity of dendritic spines on accumbens medium spiny neurons (MSNs), as well as the size of VTA dopamine neurons (Sklair-Tavron et al., 1996; Robinson and Kolb, 2004; Russo et al., 2007), whereas stimulants increase MSN dendritic complexity and spine density (Robinson et al., 2001; Norrholm et al., 2003; Lee et al., 2006; Sarti et al., 2007) and increase VTA dopaminergic neuronal size (Sarti et al., 2007). Other evidence suggests that morphine and cocaine may induce conditioned place preference through different physiologic mechanisms. German and Fields (2007) recorded neurons in the nucleus accumbens of rats during the expression of morphine-induced place preference and found that MSNs slowed firing rates when the rat was in the morphine-paired zone. In contrast, Sjulson et al. (2018) found that cocaine-induced place preference was associated with recruitment of D2-positive MSNs to fire in the cocaine-paired location. Thus, cocaine and morphine interact with the VTA-accumbens pathway in unique ways. The present results show the different reliances of cocaine versus morphine on GSK3β signaling in the nucleus accumbens in the development of drug-associated place preference. Although the accumbens is clearly a site of importance, further studies are needed to identify neuron-specific contributions of GSK3β, and GSK3β in other brain regions may work independently or in concert with the accumbens to mediate cocaine reward.
GSK3β in cocaine self-administration has been reported (Crofton et al., 2017). GSK3β was reduced by 35% in the medial shell of the accumbens by delivery of AAV-shGSK3β in the rat. GSK3β knockdown did not affect cocaine acquisition, progressive ratio responding, extinction, or cocaine-primed reinstatement behaviors. Rats with reduced GSK3β in the accumbens medial shell showed a small but significant increase in responding at high unit doses of cocaine, but no differences at lower doses (Crofton et al., 2017). It is difficult to compare self-administration and place conditioning models given the different processes involved in operant responding versus Pavlovian condition, as well as the level of drug exposure (high drug exposure in self-administration vs. low drug exposure in place conditioning). Furthermore, the larger reduction in GSK3β in both the accumbens core and shell regions that was achieved in the current study could have a greater influence on cocaine reward than the limited reductions in GSK3β in the medial shell in the aforementioned study, given the known differences in the role of core versus shell in reward processes (e.g., West and Carelli, 2016). These procedural differences (e.g., extent and location of GSK3β knockdown, cocaine dose, and species) need to be rigorously tested to make firm conclusions about GSK3β in cocaine conditioned reward versus self-administration.
Like conditioned place preference, conditioned place aversion involves similar Pavlovian processes, but with a negative component such that animals avoid a context previously associated with an aversive state. The activation of κ-opioid receptors produces a negative state associated with dysphoria and anxiety (Lalanne et al., 2014), and thus subjects given a κ-opioid receptor agonist in a specific environment show a conditioned aversion to that environment (Shippenberg and Herz, 1986). Motivational systems in the nucleus accumbens are necessary not only for obtaining rewards but also for the avoidance of harm (Kohls et al., 2013; Wise and Koob, 2014; Wenzel et al., 2015). Previous work has indicated that dopaminergic transmission in the accumbens is required for the κ-opioid receptor agonist U50,488H to produce conditioned place aversion (Bals-Kubik et al., 1993). Furthermore, intra-accumbens infusion of the D1-receptor antagonist R(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride (SCH 23390) blocks the acquisition of κ-receptor agonist–induced place aversion (Shippenberg et al., 1993). The present results demonstrate that knockdown of GSK3β interfered with the development of U50,488H-induced place aversion, indicating that accumbal GSK3β signaling plays an important role in the negative state produced by κ-receptor agonists. A recent report demonstrated that subpopulations of mesolimbic dopamine neurons respond differently to appetitive and aversive stimuli (de Jong et al., 2019). Excitation to reward-predicting cues dominates in accumbens lateral shell dopamine terminals, whereas aversive prediction signals in ventral accumbens medial shell (de Jong et al., 2019). Other results support the conclusion that the nucleus accumbens is involved in both reward and aversive learning but indicates that reward-based behaviors are dependent on transmission of the direct striatonigral pathway, whereas aversive learning behaviors are dependent on indirect striatopallidal transmission (Hikida et al., 2010). Further studies are needed to establish the roles of GSK3β in different subregions and cell populations of the accumbers in both cocaine reward and U50,488H-induced conditioned aversion.
The novel object recognition test is used to detect both impairment and enhancement of memory. A short delay (0.5–3 hours) between training and testing phases is used for studying memory impairment, and a longer delay (24 hours) is used for testing memory improvement (Papp et al., 2017). The current study used a 30-minute delay interval after object training (familiarization) to test for memory deficits in mice with accumbal GSK3β knockdown. The results demonstrated intact novel object memory; however, performance on the object location task was impaired by GSK3β knockdown. Thus, differential roles of accumbal GSK3β exist between memory for object identity and the respective spatial location. Different brain regions contribute to performance of the novel object recognition and object location tasks. The insular, perirhineal, and medial prefrontal cortices contribute to the learning and memory formation of object recognition (Mendez et al., 2015; Tanimizu et al., 2018). Object location recognition is dependent on spatial learning and memory. The role of hippocampus in novel object recognition and/or location memory formation is controversial. Several studies have shown that the hippocampus is required for object location memory but not novel object recognition memory (Mumby et al., 2002; Balderas et al., 2008; Roozendaal et al., 2010), especially in relation to one-trial object recognition tasks (Forwood et al., 2005) such as those used in the current study. The nucleus accumbens contributes to the processing of hippocampal-dependent spatial information (Ferretti et al., 2005), and performance on spatial tasks depends on functional interaction between the hippocampus and accumbens (Ito et al., 2008). Neuroanatomical evidence has demonstrated that the accumbens medial shell receives input from the ventral hippocampus, whereas the core and rostrolateral shell are innervated largely by the dorsal hippocampus (Groenewegen et al., 1999). Other work has shown that the core and shell of the nucleus accumbens and their dopamine innervation are involved in different forms of recognition memory (Nelson et al., 2010). For example, dopamine depletion in the core or the core plus the shell, but not the shell alone, disrupts novel object memory after a 24-hour delay, but not after a short (5-minute) delay. In contrast, dopamine depletion in the shell or a larger shell plus core lesions impaired short-term location memory (10-minute delay) in the object location test (Nelson et al., 2010). These evidences support our present finding that knockdown of GSK3β in the nucleus accumbens (core plus shell) affects short-term spatial location memory, but not nonspatial object memory.
In summary, these data demonstrate the importance of GSK3β in the nucleus accumbens for the development of both cocaine conditioned place preference and U50,488H conditioned place aversion, but not for morphine place preference. Moreover, GSK3β signaling in the nucleus accumbens engages during the object location task, but not in the novel object recognition task. Thus, GSK3β in the nucleus accumbens plays differential roles in cocaine and opiate reward process, as well as in memory for object identity and the respective spatial location.
Acknowledgments
We acknowledge Dr. Jim R. Woodgett from the Lunenfeld-Tanenbaum Research Institute in Toronto, Canada, for generously providing the GSK3β floxed mice used to establish our breeding colony. We also thank Ahmed Mirza and Michael Zaykaner for contributing to the successful completion of this study. We are grateful to the National Institute on Drug Abuse drug supply program for the generous contribution of the drugs used in this study.
Authorship Contributions
Participated in research design: Shi, Barr, Unterwald.
Conducted experiments: Shi, Barr, von Weltin, Wolsh.
Contributed new reagents or analytic tools: Shi, Barr.
Performed data analysis: Shi, Barr.
Wrote or contributed to the writing of the manuscript: Shi, Barr, Unterwald, von Weltin, Wolsh.
Footnotes
- Received April 19, 2019.
- Accepted July 31, 2019.
↵1 X.S. and J.L.B. contributed equally to this work.
This work was supported by the National Institutes of Health [Grants R01 DA043988 (to E.M.U.) and P30 DA013429 (to E.M.U.)]. The authors declare that they have no conflicts of interest.
Abbreviations
- AAV
- adeno-associated virus
- DI
- discrimination index
- GSK3
- glycogen synthase kinase-3
- flx
- floxed
- MSN
- medium spiny neuron
- U50, 488H, trans-(±)-3,4-dichloro-N-methyl-N-[2-(1-pyrrolidinyl)cyclohexyl]benzeneacetamide methanesulfonate salt; VTA
- ventral tegmental area
- WT
- wild type
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}