


Abstract
Cannabis is widely used in the United States, with an estimated prevalence of 9.5%. Certain cannabinoids in Cannabis sativa, Δ9-tetrahydrocannabinol (THC) in particular, possess immune-modulating and anti-inflammatory activity. Depending on the context, the anti-inflammatory activity of cannabinoids may be beneficial (e.g., in treating inflammatory diseases) or detrimental to normal immune defense against pathogens. The potential beneficial effect of cannabinoids on chronic neuroinflammation has gained recent attention. Monocyte migration to the brain has been implicated as a key event in chronic neuroinflammation and in the etiology of central nervous system diseases including viral infection (e.g., human immunodeficiency virus–associated neurocognitive disorder). In the brain, monocytes can contribute to neuroinflammation through interactions with astrocytes, including inducing astrocyte secretion of cytokines and chemokines. In a human coculture system, monocyte-derived interleukin (IL)-1β due to Toll-like receptor 7 (TLR7) activation has been identified to promote astrocyte production of monocyte chemoattractant protein (MCP)-1 and IL-6. THC treatment of the TLR7-stimulated coculture suppressed monocyte secretion of IL-1β, resulting in decreased astrocyte production of MCP-1 and IL-6. Furthermore, THC displayed direct inhibition of monocytes, as TLR7-stimulated monocyte monocultures treated with THC also showed suppressed IL-1β production. The cannabinoid receptor 2 (CB2) agonist, JWH-015, impaired monocyte IL-1β production similar to that of THC, suggesting that THC acts, in part, through CB2. THC also suppressed key elements of the IL-1β production pathway, including IL1B mRNA levels and caspase-1 activity. Collectively, this study demonstrates that the anti-inflammatory properties of THC suppress TLR7-induced monocyte secretion of IL-1β through CB2, which results in decreased astrocyte secretion of MCP-1 and IL-6.
SIGNIFICANCE STATEMENT Because cannabis use is highly prevalent in the United States and has putative anti-inflammatory properties, it is important to investigate the effect of cannabinoids on immune cell function. Furthermore, cannabinoids have garnered particular interest due to their potential beneficial effects on attenuating viral-induced chronic neuroinflammation. This study utilized a primary human coculture system to demonstrate that the major psychotropic cannabinoid in cannabis, Δ9-tetrahydrocannabinol, and a cannabinoid receptor-2 selective agonist suppress specific monocyte-mediated astrocyte inflammatory responses.
Introduction
Cannabis use is common in the United States, with a prevalence of 9.5% in a 2012–2013 report (Hasin et al., 2015), and is expected to increase as more states legalize recreational and/or medical use. Cannabis (Cannabis sativa) is made up of more than 500 chemicals, 104 of which are defined as phytocannabinoids (ElSohly and Gul, 2014). One of the major cannabinoids within cannabis is Δ9-tetrahydrocannabinol (THC); as of 2014, THC constitutes, on average, 12% of the plant material (ElSohly et al., 2016). THC is the main psychotropic component of cannabis because it crosses the blood–brain barrier to alter cognition, including short-term memory (Atakan, 2012). Furthermore, THC is immunomodulatory, with the majority of the literature demonstrating immune-suppressive and anti-inflammatory activity in vivo and in vitro (Croxford and Yamamura, 2005; Karmaus et al., 2011, 2013; Katchan et al., 2016). THC modulates the activity of almost every immune cell type, including T cells, B cells, monocytes/macrophages, natural killer cells, and dendritic cells, and suppresses a range of inflammatory functions, including proliferation, chemotaxis, phagocytosis, and cytokine production (Croxford and Yamamura, 2005; Turcotte et al., 2015; Katchan et al., 2016). Immune cell modulation by THC occurs primarily through the binding of specific G protein–coupled receptors, termed cannabinoid receptors 1 and 2 (CB1 and CB2, respectively) (Pertwee, 1997); however, CB-independent effects have been observed (Brown, 2007; Karmaus et al., 2012).
A hallmark of central nervous system (CNS) diseases, including CNS viral infections, is chronic neuroinflammation, which is characterized by the following: viral entry into the brain; activation of cells, including astrocytes, microglia, and blood-derived immune cells (e.g., monocytes); and release of proinflammatory and toxic factors. If left unresolved, this inflammation ultimately leads to neuronal dysfunction and cognitive decline (González-Scarano and Martín-García, 2005; Chen et al., 2016; Colombo and Farina, 2016; Chitnis and Weiner, 2017; Klein et al., 2017; Soung and Klein, 2018). Activated monocytes are thought to be an important peripherally derived contributor to viral-mediated neuroinflammation [e.g., human immunodeficiency virus (HIV)], because they migrate into the brain, release viral products, and secrete cytokines [e.g., tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6] (Fischer-Smith et al., 2001; Williams et al., 2001, 2014a; Terry et al., 2012; Campbell et al., 2014; Scutari et al., 2017). The inflammatory cytokines produced by monocytes may have a profound effect on neuronal injury by promoting astrocyte dysfunction, including chronic astrocyte secretion of proinflammatory cytokines/chemokines (Andjelkovic et al., 2000; Sofroniew and Vinters, 2010; Rizzo et al., 2019). Activated astrocytes produce an array of cytokines and chemokines, including monocyte chemoattractant protein (MCP)-1 and IL-6, and these factors contribute to ongoing leukocyte infiltration and cytokine secretion when they are chronically elevated (Sofroniew, 2015). Specifically, IL-6 and MCP-1 are increased in the plasma and/or cerebrospinal fluid during specific chronic viral infections (e.g., HIV) and are associated with neuronal injury and/or cognitive impairment (Kamat et al., 2012; Anderson et al., 2015; Lake et al., 2015; Yuan et al., 2015).
Reports investigating monocyte–astrocyte interactions have identified that human monocytes/macrophages promote astrocyte release of various inflammatory factors (Genis et al., 1992; Andjelkovic et al., 2000; Muratori et al., 2010). We recently developed and characterized a human coculture system containing primary monocytes and astrocytes to study inflammatory responses induced by the Toll-like receptor 7 (TLR7) agonist, imiquimod (R837) (Rizzo et al., 2019). TLR7 activation was used to mimic a major mechanism by which viral-derived single-stranded RNA stimulates monocytes and astrocytes (Kawai and Akira, 2006; Furr and Marriott, 2012; Chattergoon et al., 2014; Cohen et al., 2015). TLR7-stimulated monocytes promoted astrocyte production of MCP-1 and IL-6. Furthermore, monocyte secretion of IL-1β was found to play a critical role in promoting astrocyte production of MCP-1 and IL-6, as an IL-1β neutralization antibody inhibited the astrocyte response. Furthermore, the direct addition of recombinant IL-1β to astrocyte monocultures mimicked the effects observed with monocytes (Rizzo et al., 2019).
We previously demonstrated that THC directly suppresses specific proinflammatory human monocyte processes, including transitioning into the CD16+ phenotype and interferon-γ–inducible protein 10 secretion (Rizzo et al., 2018). Because THC is known to cross the blood–brain barrier (Atakan, 2012; Cabral and Jamerson, 2014) and both monocytes and astrocytes express CB1 and CB2 (Sheng et al., 2005; Roth et al., 2015), THC has the potential to modulate monocyte and astrocyte inflammation. The objective of this study was to evaluate the effect of THC on monocyte-mediated astrocyte production of MCP-1 and IL-6 in TLR7-stimulated cocultures. In addition, because monocyte-derived IL-1β is a primary factor governing astrocyte production of MCP-1 and IL-6, the effect of THC on IL-1β secretion by monocytes was investigated.
Materials and Methods
Reagents.
Imiquimod (R837) was purchased from InvivoGen (San Diego, CA). The Golgi transport inhibitor (2 μM Monensin/3.0 μg/ml Brefeldin A in methanol) was purchased from Thermo Fisher Scientific (Waltham, MA). Trypsin-EDTA (0.25%) was also purchased from Thermo Fisher Scientific. The following antibodies were purchased from BioLegend (San Diego, CA; clone and catalog numbers are shown in parentheses, respectively): anti–CD45-Pacific Blue (HI30, 304029), anti–CD14-Pe-cyanine dye Cy7 (M5E2, 301814), anti–CD16-allophycocyanin (3G8, 302012), anti–CD56-peridinin chlorophyll protein complex (HCD56, 318342), anti–CD57-peridinin chlorophyll protein complex/cyanine dye Cy5.5 (HNK-1, 359622), anti–IL-6-allophycocyanin (MQ2-13A5, 501112), anti–IL-1β-fluorescein isothiocyanate (JK1B-1, 508206), and anti–MCP-1-r-phycoerythrin (5D3-F7, 502604). Recombinant human IL-1β was purchased from BioLegend.
Chemicals.
THC and 11-nor-9-carboxy-Δ9-tetrahydrocannabinol (THC-COOH) were obtained from the National Institutes of Health National Institute on Drug Abuse (Bethesda, MD) in 100% ethanol. The CB2 agonist, JWH-015, was purchased from Cayman Chemical (Ann Arbor, MI) and dissolved in 100% ethanol. For cell culture experiments, cannabinoids were serially diluted in supplemented astrocyte media. The vehicle concentration for each treatment was 0.03% ethanol.
Peripheral Blood Mononuclear Cell and Monocyte Isolation.
Peripheral blood mononuclear cells were isolated from human leukocyte packs (Gulf Coast Regional Blood Center, Houston, TX) by density gradient centrifugation using Ficoll-Paque PLUS (GE Healthcare Life Sciences, Pittsburgh, PA). The sex of the donors used is unknown. Pan monocytes were isolated by negative selection (Miltenyi Biotec, Bergisch Gladbach, Germany) per the manufacturer’s instructions. The mean ± S.D. monocyte purity for donors (N = 55) used in this report was 97.0% ± 1.0%.
Human Primary Astrocyte Cell Culture.
Fetal-derived normal human astrocytes (NHAs) isolated from the cerebral cortex were purchased from Lonza (Basel, Switzerland) and cultured according to the manufacturer’s instructions in astrocyte basal media (ABM) supplemented with the SingleQuots Kit (recombinant human epidermal growth factor, insulin, ascorbic acid, gentamicin sulfate-amphotericin, l-glutamine, and FBS). Media were changed every other day for passaging and plate seeding. NHAs were used up to passage 4. Primary astrocytes from two distinct donors were used (female and male).
Cannabinoid Treatment of Monocyte-Astrocyte Coculture.
NHAs were seeded in supplemented ABM 3–6 days prior to coculture at a cell density (based on doubling time) that would be an estimated 2 × 105 cells/ml on the day of coculture. On the day of monocyte isolation, astrocytes were treated with THC at concentrations noted in the respective figures. Monocytes were added immediately to the well at a final concentration of 1 × 104 cells/ml to establish a monocyte/astrocyte ratio of 1:20. Imiquimod/R837 (TLR7 agonist) was then added at 10 μg/ml and the cells were incubated at 37°C and 5% CO2 for 20 hours and a Golgi block was added to culture for an additional 4 hours. Prior to the addition of the Golgi block, an aliquot of the supernatant was removed and stored at −80°C. At 24 hours, wells were washed with nonsupplemented media and trypsinization was performed to remove astrocytes from the plate. The flow cytometry procedure below was performed to measure cell viability and cytokine production.
Cannabinoid Treatment of Astrocyte Monocultures Stimulated with Recombinant IL-1β and R837.
Primary astrocytes were seeded into 24-well plates as described in the coculture section above. Astrocyte monocultures were treated with cannabinoids and activated using recombinant IL-1β (noted in each experiment) plus R837 (10 μg/ml). Astrocytes were incubated for 20 hours and a Golgi block was added for 4 hours. At 24 hours, wells were washed with nonsupplemented media and trypsinization was performed to remove astrocytes from the plate. Cell viability and cytokine production was measured by flow cytometry as described below.
Cannabinoid Treatment of TLR7-Activated Monocytes.
Purified monocytes (4 × 105 cells/ml) cultured in supplemented ABM were treated with THC and stimulated with R837 (10 μg/ml). Cells were incubated at 37°C and 5% CO2 for the specific time points noted in each figure legend. Cells or supernatants were harvested and downstream measurements were performed.
Supernatant Measurements of Cytokines and Chemokines.
For particular experiments, supernatants were collected at 20 hours and stored at −80°C. ELISA Max (BioLegend) was used to quantify the levels of MCP-1, IL-6, or IL-1β in the supernatants.
Annexin-V and 7-Amino-Actinomycin D Staining of Human Monocytes.
Monocytes were removed from tissue culture plates using mini cell scrapers (Biotium, Fremont, CA) at time points stated in the figure legends and were transferred to 96-well U-bottom plates. The Pacific Blue Annexin V Apoptosis Detection Kit with 7-amino-actinomycin D (7-AAD; BioLegend) was used per the manufacturer’s instructions. Samples were analyzed by flow cytometry.
IL-1β Gene Expression Analysis.
Monocyte RNA was isolated using the RNeasy Mini Kit (Qiagen, Hilden, Germany). Reverse transcription polymerase chain reaction (PCR) was performed using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Real-time PCR using TaqMan probes targeting IL1B (Hs01555410_m1; Thermo Fisher Scientific) was used to quantify mRNA levels. The loading control was 18S ribosomal RNA (Thermo Fisher Scientific).
Caspase-1 Activity.
At the time points stated in the figure legends, monocytes were treated with the fluorescent caspase-1 inhibitor probe FAM-YVAD-FMK (FAM-FLICA Caspase-1 Assay Kit; ImmunoChemistry Technologies, Bloomington, MN) for 30 minutes. Cells were washed once and stained with the viability stain, Near-IR (Thermo Fisher Scientific), for 10 minutes. Cells were washed twice and fixed (BD Cytofix; BD Biosciences, San Jose, CA). Cell scraping or trypsinization was used to remove the adherent monocytes from the culture plates. Samples were analyzed via flow cytometry.
Flow Cytometry.
Staining buffer (PBS, 1% bovine serum albumin, and 0.1% NaN3) was used to wash cells in between staining and fixing steps. Cells were first incubated with LIVE/DEAD Fixable Near-IR Dead Cell Stain (Thermo Fisher Scientific) and then with staining buffer containing 20% human AB serum to block Fc receptors. BD Cytofix was used to fix cells. For intracellular staining, cells were stained with antibody in BD Perm/Wash (BD Biosciences). Data analysis was performed using FlowJo version 10 software. For analysis of coculture experiments, astrocytes were gated apart from monocytes based on forward-scatter area and height and optimization experiments verified that the astrocytes were positive for glial fibrillary acidic protein (Rizzo et al., 2019). The percentage of MCP-1+ or IL-6+ cells within the astrocyte population, as well as the mean fluorescence intensity (MFI) of MCP-1 or IL-6 within the respective positive population, is reported in the figures. The gates set for each protein were based on isotype controls. For experiments examining intracellular expression of IL-1β and caspase-1 activity, viable monocytes were first gated prior to examination of the respective intracellular endpoint.
Monocyte surface staining was performed to determine purity immediately after isolation using anti-CD14, CD16, CD45, CD56, and CD57. Single cells were first gated based on expression of the common leukocyte antigen, CD45, to differentiate leukocyte from nonleukocyte events. Within the CD45+ population, the total monocyte population was determined by surface expression of CD14 and CD16. Specifically, total monocytes were identified as CD14++CD16− (classic), CD14++CD16+ (intermediate), or CD14+/−CD16+ (nonclassic) (Wong et al., 2012). The natural killer cell surface markers, CD56 and CD57, were used to identify and remove natural killer cells from the purity determination. Purity was defined as the percentage of monocytes (classic, intermediate, or nonclassic) lacking CD56/CD57 within the CD45+ population.
Statistical Analysis.
Statistical analysis was performed using Prism 7 or 8 software (GraphPad, San Diego, CA). The experimental data were graphed as the mean ± S.E.M. The statistical tests performed for each experiment are indicated in the figure legends.
Results
THC Treatment of the TLR7-Stimulated Coculture Resulted in Decreased Astrocyte Production of MCP-1 and IL-6.
To determine the effect of THC on monocyte-induced astrocyte secretion of MCP-1 and IL-6 in response to R837, monocytes and astrocytes were cocultured at a 1:20 ratio, treated with THC (1, 5, and 10 μM), and stimulated with R837 (10 μg/ml) for 20 hours. A Golgi block was added for 4 hours and astrocytes were measured for intracellular MCP-1 and IL-6 via flow cytometry. Prior to the addition of the Golgi block, an aliquot of supernatant was collected for MCP-1 and IL-6 quantification. Flow cytometry plots from a representative donor demonstrating the effect of THC (10 μM) on astrocyte production of MCP-1 and IL-6 in the R837-treated coculture are shown in Figs. 1 and 2, A–C, respectively. THC at the highest concentration examined (10 μM) modestly decreased the percentage of astrocytes expressing MCP-1 (Fig. 1D). Furthermore, THC decreased MCP-1 levels (MFI) within the MCP-1+ astrocytes as well as supernatant MCP-1 at 5 and 10 μM (Fig. 1, E and F). When astrocyte production of IL-6 was examined, THC decreased the percentage of IL-6+ astrocytes and IL-6 levels (MFI) in a concentration-dependent manner, with significant suppression of the percentage of IL-6+ astrocytes observed at 5 and 10 μM and levels (MFI) at 10 μM (Fig. 2, D and E). As observed with MCP-1, the supernatant level of IL-6 was also decreased with THC in the R837-stimulated coculture at 5 and 10 μM (Fig. 2F). THC treatment had no observable effect on astrocyte viability (data not shown).

THC treatment of the TLR7-stimulated coculture resulted in decreased astrocyte production of MCP-1. Human primary monocytes (n = 9) were cocultured with primary astrocytes at a ratio of 1:20 (monocytes/astrocytes), treated with vehicle or THC (1, 5, and 10 μM) and stimulated with R837 (10 μg/ml). Monocyte and astrocyte cell concentrations were 1 × 104 and 2 × 105 cells/ml, respectively. Cells were cocultured for 24 hours, with a Golgi block added 4 hours before cell harvest. An aliquot of the supernatant was collected prior to the addition of the Golgi block. Intracellular staining and flow cytometry were performed to determine astrocyte production of MCP-1 (percent positive and MFI). ELISA Max technology from BioLegend was used to determine supernatant levels of MCP-1. (A–C) Flow cytometry plots from one representative donor. (D-F) A repeated-measures ANOVA with a Dunnett post hoc test was performed. (F) The data set was log transformed prior to statistical testing. Asterisks denote a statistical difference from the vehicle plus R837 control (*P < 0.05; ***P < 0.001). All graphs are means ± S.E.M. PE, r-phycoerythrin; VH, vehicle.

THC treatment of the TLR7-stimulated coculture resulted in decreased astrocyte production of IL-6. Human primary monocytes (n = 9) were cocultured with primary astrocytes at a ratio of 1:20 (monocytes/astrocytes), treated with vehicle or THC (1, 5, and 10 μM), and stimulated with R837 (10 μg/ml). Monocyte and astrocyte cell concentrations were 1 × 104 and 2 × 105 cells/ml, respectively. Cells were cocultured for 24 hours, with a Golgi block added 4 hours before cell harvest. An aliquot of the supernatant was collected prior to the addition of the Golgi block. Intracellular staining and flow cytometry were performed to determine astrocyte production of IL-6 (percent positive and MFI). ELISA Max was used to determine supernatant levels of IL-6. (A–C) Flow cytometry plots from one representative donor. (D) The data set was log transformed prior to statistical testing. (E and F) The data sets were normalized to the vehicle plus R837 control, which served as 100%. The supernatant level of IL-6 of the vehicle plus R837 control in (C) was 59.7 ng/ml. (D-F) A repeated-measures ANOVA with a Dunnett post hoc test was performed. Asterisks denote a statistical difference from the vehicle plus R837 control ( **P < 0.01; ***P < 0.001). All graphs are means ± S.E.M. APC, allophycocyanin; VH, vehicle.
THC Treatment of the TLR7 Plus IL-1β–Stimulated Astrocyte Monoculture Promoted a Decrease in Astrocytes Expressing MCP-1 and IL-6.
To examine whether THC directly acts on astrocytes, astrocyte monocultures from two separate donors were treated with THC and stimulated with IL-1β plus R837 for 20 hours. THC treatment decreased the percentage of astrocytes expressing MCP-1 and IL-6, which was consistent between both donors (Fig. 3, A, C, E, and G). THC had no effect on the levels of MCP-1 (MFI) in both donors and there was a minor decrease in astrocyte levels of IL-6 (MFI) (Fig. 3, B, D, F, and H). As in the coculture, THC treatment had no observable effect on astrocyte viability (data not shown).

THC treatment of the TLR7 plus IL-1β–stimulated astrocyte monoculture decreased the percentage of astrocytes expressing MCP-1 and IL-6. Primary astrocytes were treated with vehicle or THC (1, 5, and 10 µM) and stimulated with both IL-1β (20 or 100 pg/ml) and R837 (10 µg/ml). Cells were cultured for 24 hours, with a Golgi block added 4 hours before cell harvest. Intracellular staining and flow cytometry were performed to determine astrocyte production of MCP-1 and IL-6 (percent positive). (A–H) Experiments performed on astrocyte donor 1 (A–D) and astrocyte donor 2 (E–H). For each astrocyte donor, experimental wells were performed in triplicate. A repeated-measures ANOVA with a Dunnett post hoc test was performed. Asterisks denote a statistical difference from the corresponding vehicle control (*P < 0.05; **P < 0.01; ***P < 0.001). All graphs are means ± S.E.M. VH, vehicle.
THC Treatment of Coculture and Monocyte Monoculture Stimulated with R837 Resulted in Decreased Monocyte Production of IL-1β.
Given that monocyte-derived IL-1β is a critical factor governing astrocyte secretion of MCP-1 and IL-6 in the R837-stimulated coculture (Rizzo et al., 2019), the effect of THC on TLR7-mediated monocyte secretion of IL-1β was examined. First, we determined the effect of THC on supernatant IL-1β levels in the R837-stimulated coculture supernatants (the same cocultures as displayed in Figs. 1 and 2). The left graph in Fig. 4A shows a significant increase in the amount of supernatant IL-1β in the R837-stimulated coculture compared with the nonstimulated coculture. THC treatment decreased the level of supernatant IL-1β in a concentration-dependent manner, with suppression starting at the lowest concentration measured (1 μM) (Fig. 4A, right).

THC treatment of coculture and monocyte monoculture stimulated with R837 resulted in decreased monocyte production of IL-1β, and the CB2 agonist, JWH-015, suppressed IL-1β in a similar manner as THC. (A) The same THC-treated R837-stimulated coculture experiments performed in Figs. 1 and 2 were measured for supernatant IL-1β via the enzyme-linked immunosorbent assay (ELISA). (B) Human monocyte monocultures (n = 21; 4 × 105 cells/ml) were treated with vehicle or THC (1, 5, and 10 µM) and stimulated with R837. (C and D) Human monocyte monocultures (n = 8; 4 × 105 cells/ml) were treated with vehicle or THC (0.1, 0.5, and 1 µM) and stimulated with R837. (E and F) Human monocytes (n = 8–12) were treated with THC, JWH-015, or THC-COOH and stimulated with R837. (A–F) Supernatants were harvested at 20 hours postactivation for IL-1β and/or IL-6 (D) quantification (ELISA). Graphs on the left in (A) and (B) present the raw levels of IL-1β produced with R837 activation. Graphs on the right in (A) through (F) were normalized to the R837-stimulated vehicle control, which serves as 100%. For the graphs on the left in (A) and (B), data sets were log transformed and a paired t test was performed (*P < 0.05; ***P < 0.001). For the graphs on the right in (A) through (E), a repeated-measures ANOVA with a Dunnett post hoc test was performed. For (F), a paired t test was performed between the activated (R837) vehicle control and activated treatment group (THC or THC-COOH). For normalized data sets in (A) through (F), asterisks denote a statistical difference from the vehicle plus R837 control (*P < 0.05; ***P < 0.001). All graphs are means ± S.E.M. VH, vehicle.
To determine whether THC directly suppresses monocyte secretion of IL-1β, monocyte monocultures were treated with THC and stimulated with R837 for 20 hours. R837 stimulation of monocytes promoted a marked increase in IL-1β levels (Fig. 4B, left). The differences in IL-1β levels between the coculture and monocyte monoculture were due to an increase in the number of monocytes in the monocultures (4 × 105 vs. 1 × 104 cells/ml) (Fig. 4, A and B, left). THC displayed similar effects as in the coculture, with a concentration-dependent decrease in secreted IL-1β with 1 μM THC suppressing IL-1β by approximately 30% compared with the vehicle control (Fig. 4B, right). These experiments were extended to determine whether lower concentrations of THC (0.1 and 0.5 μM) would impair monocyte-derived IL-1β. As shown in Fig. 4C, THC at concentrations as low as 0.5 μM significantly decreased IL-1β levels, which was within the plasma concentration range (0.25–0.85 μM) after smoking cannabis (Huestis, 2007). To determine whether the THC-mediated suppression of TLR7-activated monocytes was specific to IL-1β secretion, we determined the effect of THC on monocyte secretion of IL-6 from the same donors in Fig. 4C. Interestingly, THC had no effect on supernatant IL-6 levels (Fig. 4D), suggesting that THC specifically suppresses monocyte production of IL-1β. In addition, we did not observe a significant effect on monocyte secretion of IL-6 by 5 μM THC but did observe an inhibitory effect at 10 μM (data not shown).
The CB2 Agonist, JWH-015, Suppresses Monocyte Secretion of IL-1β Similar to That of THC.
Because THC is known to modulate immune cell activity, in part, through CB2 (Cabral and Griffin-Thomas, 2009; Tanasescu and Constantinescu, 2010), we investigated whether the CB2 agonist, JWH-015, displayed a similar effect on R837-mediated monocyte secretion of IL-1β as THC. Monocyte monocultures were treated with either THC (1, 5, and 10 μM) or JWH-015 (1, 5, and 10 μM) and stimulated with R837 for 20 hours. JWH-015 decreased monocyte-derived IL-1β in a concentration-dependent manner with significant suppression observed at 5 and 10 μM (Fig. 4D). The suppression with JWH-015 was similar to that of THC, suggesting that THC acts, in part, through CB2 (Fig. 4D). However, some differences were observed, including variation in the sensitivity between the two compounds, as 1 μM THC, but not JWH-015, promoted a significant decrease in IL-1β (Fig. 4D). In addition, 10 μM THC displayed a greater level of suppression on IL-1β secretion (88%) compared with JWH-015 (57%) (Fig. 4D). To examine the possibility of CB-independent mechanisms, R837-stimulated monocytes were treated with the inactive THC metabolite, THC-COOH (Sharma et al., 2012). THC-COOH treatment (10 μM) had no effect on monocyte secretion of IL-1β compared with the potent suppression observed with the equivalent concentration of THC (Fig. 4E).
THC at Higher In Vitro Concentrations (10 μM) Promotes Monocyte Apoptosis in Response to TLR7 Activation.
Because cannabinoid-mediated apoptosis has been observed in vitro with ≥10 μM THC (Rieder et al., 2010), we determined whether THC induced monocyte apoptosis and/or cell death when stimulated with R837. Monocyte monocultures were treated with THC, JWH-015, or THC-COOH and stimulated with R837 for 20 hours. Cells were harvested and stained for Annexin-V (marker of apoptosis) and 7-AAD (marker of cell death). The gating strategy in Fig. 5, A and B, was used to identify cells that were 1) viable but apoptotic (7-AAD−Annexin-V+) or 2) dead and apoptotic (7-AAD+Annexin-V+). The IL-1β secretion profile for the donors used in this viability analysis is depicted in Fig. 5C. THC at the highest concentration used (10 μM) induced monocyte apoptosis, as evidenced by an increase in the percentage of dead/apoptotic and viable/apoptotic monocytes compared with the vehicle control (Fig. 5, D and E). However, THC at concentrations ranging from 0.1 to 5 μM had no significant effect on the percentage of dead/apoptotic monocytes and viable/apoptotic monocytes (Fig. 5, D and E). A second experiment was performed using concentrations of 1, 5, and 10 μM THC, and the same apoptotic effect was observed but only at 10 μM THC (data not shown). In addition, JWH-015 and THC-COOH displayed minimal to no effect on viability or apoptosis (Fig. 5, D and E), suggesting that 10 μM THC promotes apoptosis through a mechanism that is unrelated to CB2, which appears to be independent of nonspecific lipophilic action.
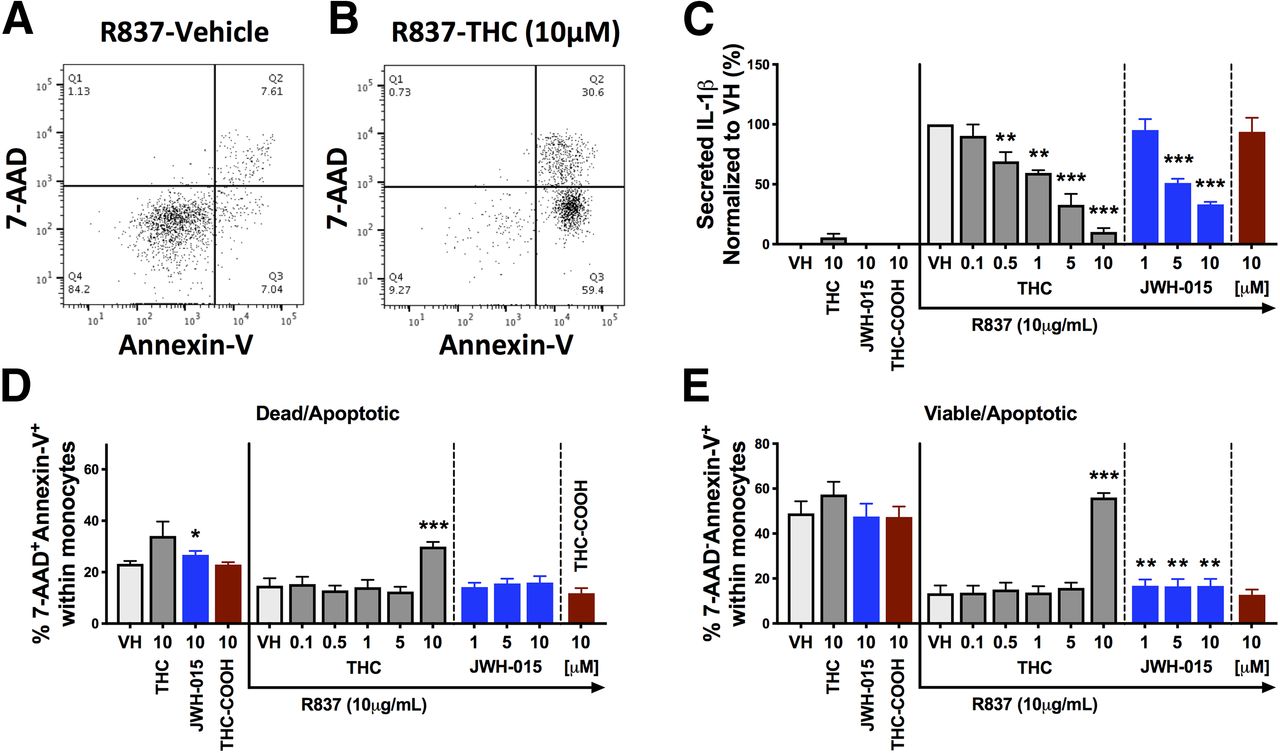
THC at 10 μM induced monocyte apoptosis in response to TLR7 activation. Human monocytes (n = 4) were treated with vehicle, THC (0.1–10 μM), JWH-015 (1, 5, and 10 μM), or THC-COOH (10 μM) and stimulated with R837 (10 μg/ml). Monocytes and supernatants were harvested at 20 hours postactivation and an Annexin-V/7-AAD staining kit was used to measure apoptosis and viability. Supernatants were collected for IL-1β quantification. (A and B) Flow cytometry plots from one representative donor comparing the vehicle control and THC (10 μM) of R837-stimulated monocytes. Dead and apoptotic cells are defined as 7-AAD+ and Annexin-V+, respectively. (C–E) A repeated-measures ANOVA with a Dunnett post hoc test was performed. Asterisks denote a statistical difference from the vehicle plus R837 control (*P < 0.05; **P < 0.01; ***P < 0.001). All graphs are means ± S.E.M. VH, vehicle.
THC Decreases IL1B mRNA Levels within R837-Stimulated Monocytes.
To evaluate the mode of action by which THC impairs monocyte secretion of IL-1β, we first determined the kinetics of IL-1β suppression by THC. Isolated monocytes were treated with vehicle or THC (5 μM) and stimulated with R837 (10 μg/ml). Supernatants were harvested at 2.5, 5, 10, 15, 20, and 25 hours and quantified for IL-1β. As observed in Fig. 6A, significant suppression of IL-1β by THC was observed as early as 5 hours and suppression was sustained over the duration of the time course (25 hours), demonstrating that THC suppressed early cellular events. Because transcriptional induction of IL1B mRNA is an early event that is critical for IL-1β production and secretion, we examined the effect of THC on IL1B mRNA levels. Monocyte monocultures were treated with THC and stimulated with R837 for 3 and 5 hours. RNA was isolated and reverse transcription quantitative PCR (TaqMan) was used to quantify monocyte expression of IL1B. Monocyte expression of IL1B was significantly lower with THC treatment, with significance detected at 5 and 10 μM at both 3 and 5 hours after R837 activation (Fig. 6, B and C). However, there was not a significant effect on IL1B levels at 1 μM THC despite the 30% suppression observed on supernatant IL-1β at the same THC concentration (Fig. 4B), suggesting that THC also modulates other aspects of the IL-1β pathway.
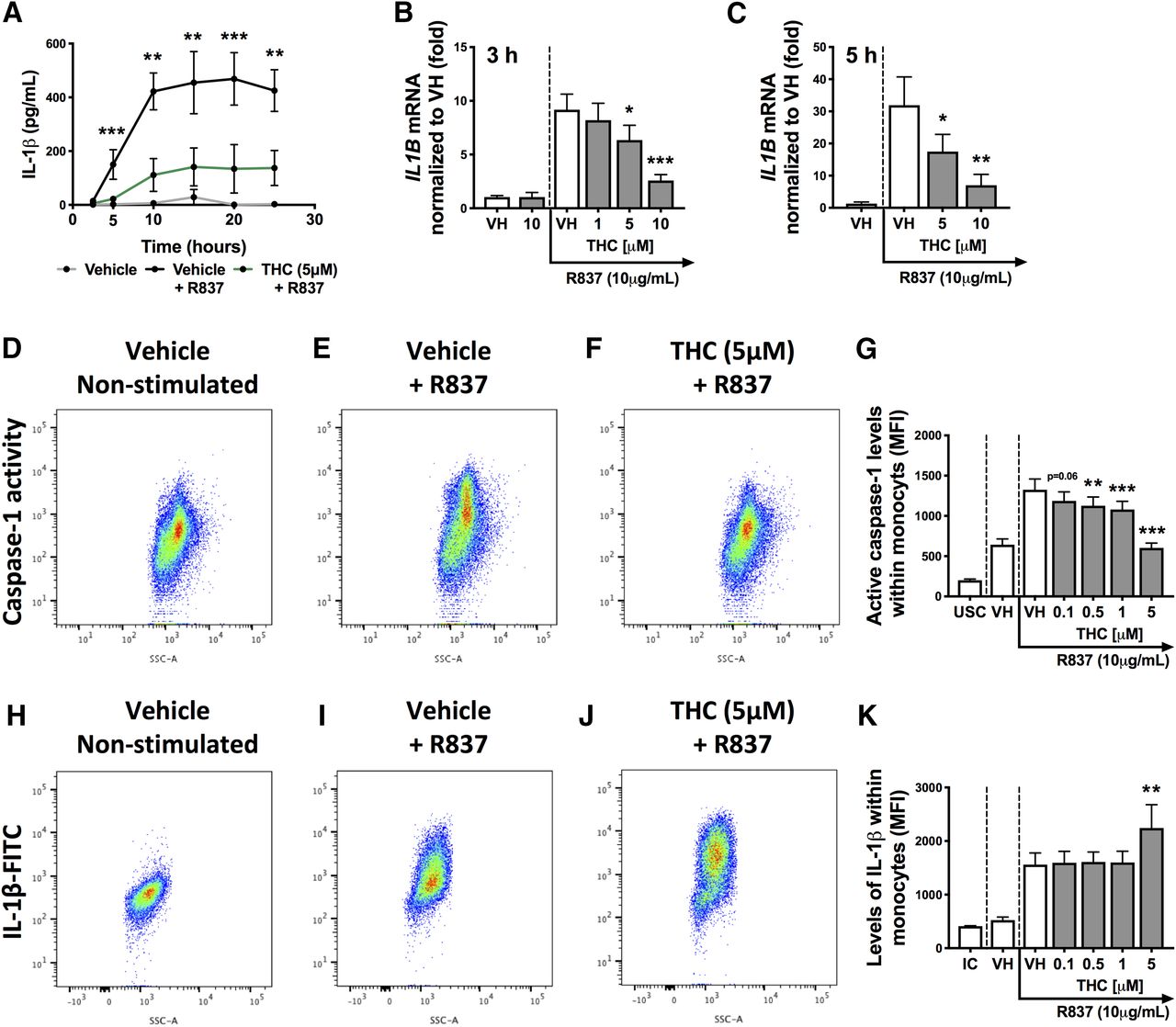
THC treatment decreased IL1B mRNA levels and caspase-1 activity within R837-stimulated monocytes, resulting in impaired IL-1β secretion. (A) Human monocytes (n = 4) were treated with vehicle or THC (5 μM) and stimulated with R837 (10 μg/ml) for 2.5, 5, 10, 15, 20, and 25 hours. Supernatants were collected at each time point and IL-1β levels were measured. (B) Human monocytes (n = 6) were treated with vehicle or THC (1, 5, and 10 μM) and stimulated with R837 (10 μg/ml) for 3 hours. (C) Human monocytes (n = 4) were treated with vehicle or THC (5 and 10 μM) and stimulated with R837 (10 μg/ml) for 5 hours. (B and C) Monocytes were harvested and IL1B mRNA was quantified. IL1B mRNA levels were normalized to the nonstimulated vehicle control. (D–G) Human monocytes (n = 7) were treated with vehicle or THC (0.1, 0.5, 1, and 5 μM) and stimulated with R837. Cells were harvested at 15 hours postactivation and quantified for caspase-1 activity (MFI) via flow cytometry. (H–K) Human monocytes (n = 7) were treated with vehicle or THC (0.1, 0.5, 1, and 5 μM) and stimulated with R837. Cells were harvested at 20 hours and quantified for intracellular IL-1β expression (MFI) via flow cytometry. Flow cytometry plots in (D) through (K) are from the same representative donor. (A) The data set was log transformed and a two-way repeated-measures ANOVA with a Sidak post hoc test was performed. Asterisks denote a statistical difference between the vehicle plus R837 and THC plus R837 treatment groups (**P < 0.01; ***P < 0.001). (B–K) A repeated-measures ANOVA with a Dunnett post hoc test was performed. Asterisks denote a statistical difference from the vehicle plus R837 control (*P < 0.05; **P < 0.01; ***P < 0.001). All graphs are means ± S.E.M. IC, isotype control; UC, unstained control; VH, vehicle.
THC Decreases Caspase-1 Activity within R837-Stimulated Monocytes, Resulting in Increased Intracellular Accumulation of IL-1β.
We next examined another major pathway involved in IL-1β production and maturation potentially affected by THC. IL-1β secretion requires both transcription/translation of the pro form (pro–IL-1β) and inflammasome-mediated caspase cleavage of pro–IL-1β into the mature form (Lopez-Castejon and Brough, 2011; Afonina et al., 2015). The major caspase involved in the cleavage of pro–IL-1β in monocytes is caspase-1, which becomes activated with inflammasome formation and activation (Afonina et al., 2015; Vanaja et al., 2015). To determine whether THC modulates caspase-1 activity, we first determined the time at which monocyte caspase-1 activity was significantly increased with R837 compared with nonstimulated cells. A fluorescence caspase-1 inhibitor probe (FAM-YVAD-FMK), which labels active caspase-1 molecules, was used to measure caspase-1 activity within monocytes. First, isolated monocytes were stimulated with R837 (10 μg/ml) and monocytes were harvested at 5, 15, and 20 hours to define the time point at which caspase-1 activity was induced. Caspase-1 activity (MFI) was significantly induced by R837 treatment at the 5- and 15-hour time points (Supplemental Fig. 1). Based on the aforementioned kinetics, the effect of THC on caspase-1 activity was evaluated at 15 hours, which not surprisingly also paralleled the peak levels of IL-1β (Fig. 6A). Representative flow cytometry plots from one donor demonstrating the effect of THC on caspase-1 activity at 15 hours in response to R837 activation are shown in Fig. 6, D–F. R837 activation elicited an approximately 2-fold increase in caspase-1 activity levels (MFI) within the monocyte population, which was decreased by THC at concentrations as low as 0.5 μM (Fig. 6G).
Because caspase-1 activity is important for the cleavage of pro–IL-1β into its mature form (Afonina et al., 2015; Vanaja et al., 2015), we speculated that suppression of caspase-1 activity observed with THC might result in intracellular accumulation of noncleaved pro–IL-1β. To investigate this possibility, monocytes were treated with THC (0.1–5 μM) and stimulated with R837 for 20 hours. Monocytes were harvested and intracellular IL-1β was examined via intracellular staining using flow cytometry. The anti–IL-1β clone used detects total IL-1β and does not distinguish between the pro and mature forms. Flow cytometry plots from the same representative donor as in the caspase-1 activity analysis are shown in Fig. 6, H–J. Interestingly, 5 μM THC promoted an increase in intracellular IL-1β expression (Fig. 6K), suggesting that THC-mediated impairment of caspase-1 activity resulted in the accumulation of intracellular pro–IL-1β.
Discussion
As cannabis use continues to become more prevalent in the United States due to legalization of medicinal and recreational use, it is important to evaluate the effect of cannabinoids within cannabis on aspects of human health, including immune function. In this study, we evaluated the effect of the major psychotropic and anti-inflammatory cannabinoid, THC, on monocyte-mediated astrocyte inflammatory responses in the context of viral (TLR7)–mediated neuroinflammation. With the use of a human primary monocyte and astrocyte coculture system stimulated with the TLR7 agonist, imiquimod/R837, we demonstrated that THC suppresses monocyte secretion of the proinflammatory cytokine, IL-1β, and astrocyte production of cytokines and chemokines (MCP-1 and IL-6) involved in leukocyte trafficking into the brain. Interestingly, the anti-inflammatory actions of THC on monocyte-astrocyte inflammation may be beneficial in the context of chronic viral infection in the CNS by putatively decelerating persistent leukocyte migration into the brain as well as by suppressing low-level neuroinflammation that exists due to the presence of activated (CD16+) monocytes, as is the case during HIV infection (Campbell et al., 2014; Williams et al., 2014a). The results shown here accompany already published reports showing that HIV-infected cannabis users display a lower level of circulating activated (CD16+) monocytes compared with noncannabis-using HIV-infected donors (Manuzak et al., 2018; Rizzo et al., 2018; Castro et al., 2019). In addition to HIV infection, THC and CB1/CB2 agonists have been shown to display beneficial anti-inflammatory actions in animal models that induce neuroinflammation (e.g., experimental autoimmune encephalomyelitis) (Maresz et al., 2007; Rom and Persidsky, 2013; Turcotte et al., 2015). In this context, cannabis use and/or cannabinoid-based therapies (e.g., synthetic THC such as dronabinol, or Marinol; AbbVie Inc., North Chicago, IL) may provide beneficial anti-inflammatory effects on specific mechanisms contributing to chronic neuroinflammation.
Conversely, THC may have detrimental effects on normal host immune defense, because impairing important monocyte-astrocyte inflammatory responses necessary for an effective antiviral immune response in the brain could result in delayed pathogen clearance and the spread of infection to normal adjacent tissue. The THC-mediated impairment in TLR7-mediated monocyte secretion of IL-1β and astrocyte secretion of MCP-1 and IL-6 could lead to decreased peripheral leukocyte migration into the brain as well as dysregulation of key immune cell effector responses (Furr and Marriott, 2012; Sofroniew, 2015). For example, the THC-mediated suppression of IL-1β observed here may be relevant to individuals with West Nile or herpes simplex virus infection, because IL-1β has been identified as an important cytokine in promoting an effective antiviral immune response in the CNS, including restricting viral replication and initiating effector responses (e.g., CD8+ T-cell responses) (Sergerie et al., 2007; Ramos et al., 2012). Furthermore, because influenza has been identified in the brain and induces neuroinflammation (Jurgens et al., 2012; Hosseini et al., 2018), the immune-suppressive effects of THC observed here might also be detrimental to host defense against this virus. Albeit in a different inflammatory location, in vivo THC treatment has been shown to suppress specific immune responses necessary for viral clearance from the lung, including leukocyte migration, in an animal model of influenza infection (Buchweitz et al., 2007; Karmaus et al., 2013).
When the coculture was separated into astrocyte and monocyte monocultures, THC modulated both cell types, albeit to varying degrees. Specifically, THC suppressed both MCP-1 and IL-6 production in IL-1β/R837–treated astrocytes; however, the magnitude of suppression was modest compared with the inhibitory effects observed in the coculture. These findings suggest that THC affects the monocyte population as well. The direct suppressive effects of THC on astrocyte production of MCP-1 and IL-6 are in agreement with findings by Sheng et al. (2005) in which the synthetic CB1/CB2 agonist, WIN55,212-2, suppressed astrocyte production of several inflammatory factors (e.g., MCP-1) in response to IL-1β activation. Because nuclear factor-κB (NFκB) is one of the major transcription factors involved in the proinflammatory response by astrocytes and is a known target of cannabinoid receptors (Cabral and Griffin-Thomas, 2009; Sofroniew, 2015; Colombo and Farina, 2016), it is tempting to speculate that THC impairs IL-1β–mediated astrocyte production of MCP-1 and IL-6 by suppressing the activation and/or transcriptional activity of NFκB. In addition to astrocytes, THC has been identified to suppress inflammatory processes in other glial populations, including IL-6 and IL-1β production by lipopolysaccharide-activated microglia (BV-2 cell line) (Kozela et al., 2010). Future studies are needed to clarify the specific molecular mechanisms responsible for THC-mediated suppression of inflammatory cytokine production.
When monocyte-derived IL-1β was examined in either the TLR7-stimulated coculture or monocyte monoculture, THC suppressed IL-1β, which is a critical factor in mediating astrocyte production of MCP-1 and IL-6 in the coculture system (Rizzo et al., 2019). Interestingly, THC at concentrations as low as 0.5 μM suppressed the R837-mediated monocyte secretion of IL-1β. This is significant because 0.5 μM is within the range of peak plasma concentration of THC (0.24–0.87 μM) after inhalation of a cannabis cigarette containing 3.55% THC (Huestis, 2007). In addition, present-day cannabis material contains roughly 12% THC (ElSohly et al., 2016), which is almost 4-fold higher than the report above, suggesting that concentrations of 1 μM used in this study are also within the observable ranges in vivo. The THC-mediated suppression of IL-1β observed here is similar to a report that demonstrated the suppressive effect of THC on lipopolysaccharide-induced monocyte secretion of IL-1β (Zurier et al., 2003), suggesting a common mode of action by THC on TLR-mediated pathways of IL-1β secretion. Furthermore, studies utilizing monocyte/macrophage and microglia cell lines have also shown immunomodulatory action by THC on IL-1β secretion (Shivers et al., 1994; Kozela et al., 2010), suggesting that THC modulation of IL-1β secretion can be generalized to other cell types.
Given that TLR7-activated monocyte-derived IL-1β is a critical factor in neuroinflammation, we explored the mode of action by which THC suppresses monocyte secretion of IL-1β. First, we identified that the CB2 agonist, JWH-015, decreased monocyte-derived IL-1β in response to TLR7 activation, suggesting that THC acts through CB2, which is consistent with other studies on THC-mediated modulation of monocyte function (Williams et al., 2014b; Roth et al., 2015). The nonactive CB metabolite, THC-COOH, had no effect on monocyte secretion of IL-1β, which further supports a CB-dependent and not a nonspecific mechanism. In addition, the anti-inflammatory effects observed with JWH-015 on monocyte secretion of IL-1β provide supportive evidence for CB2 as a therapeutic target for the treatment of inflammatory conditions, which has garnered substantial interest in the context of neuroinflammation (Cabral and Griffin-Thomas, 2009; Dhopeshwarkar and Mackie, 2014; Cassano et al., 2017).
The current body of literature shows that cannabinoids at high concentrations (>10 μM) can induce apoptosis in various immune cell populations as a pathway to immunosuppression (Zhu et al., 1998; McKallip et al., 2002; Lombard et al., 2007; Wu et al., 2008, 2018; Rieder et al., 2010). In our study, only 10 μM THC induced monocyte apoptosis when monocytes were activated through TLR7. This observation is consistent with prior reports showing cannabinoid-induced apoptosis at elevated in vitro concentrations in human monocytes as well as different species and/or cell types, and involves the activation of specific caspases (e.g., caspases-1, 3, 8, and/or 9) (Zhu et al., 1998; McKallip et al., 2002; Herrera et al., 2006; Jia et al., 2006; Wu et al., 2008, 2018; Rieder et al., 2010). Because THC treatment decreased caspase-1 activity within TLR7-activated monocytes, caspase-1–mediated apoptosis is unlikely. Additional studies are needed to elucidate the mechanism of THC-mediated monocyte apoptosis when stimulated through TLR7, including the contribution of specific caspases. Conversely, the CB2-selective agonist, JWH-015, displayed a minimal effect on monocyte apoptosis, suggesting that THC-mediated monocyte apoptosis at 10 μM is likely CB2 independent, which is in contrast with reports identifying THC and CB2 agonists promoting apoptosis through CB2 (McKallip et al., 2002; Herrera et al., 2006; Jia et al., 2006; Lombard et al., 2007). No apoptotic effects were observed at THC concentrations less than 5 μM or with JWH-015, suggesting that THC and JWH-015 suppress IL-1β secretion through a nonapoptotic, CB2-dependent mechanism.
When the mode of action was evaluated, THC was identified to suppress both major signaling pathways for IL-1β secretion (i.e., transcription/translation of pro–IL-1β and inflammasome-mediated caspase-1 activation) (Lopez-Castejon and Brough, 2011; Afonina et al., 2015). Specifically, we identified that THC treatment resulted in decreased IL1B mRNA levels in TLR7-stimulated monocytes. Based on the CB1/CB2 body of literature, it could be speculated that THC, through ligation of CB2, alters IL1B mRNA levels through decreasing the activation and/or activity of key transcription factors that regulate IL1B in response to TLR activation, including NFκB and cAMP response element binding protein (Gray et al., 1993; Cogswell et al., 1994; Cabral and Griffin-Thomas, 2009; Bosier et al., 2010; McCoy, 2016). Interestingly, the THC-mediated decrease in IL1B mRNA levels was only observed starting at 5 μM THC compared with the THC-mediated suppression of IL-1β secretion at 0.5 μM, suggesting that THC modulates additional components of the IL-1β production pathway. Indeed, THC was found to suppress caspase-1 activity (the major marker of inflammasome activation), which was observed at concentrations as low as 0.5 μM THC. Furthermore, the THC-mediated inhibition of caspase-1 activity was paralleled with an accumulation of intracellular IL-1β, suggesting impaired cleavage of pro–IL-1β by caspase-1 and further implicating a role of impaired caspase-1 activity and inflammasome activation as a primary mechanism for the THC-mediated impairment of monocyte secretion of IL-1β. Similar findings were reported in experimental autoimmune encephalomyelitis and models of dextran sulfate sodium–induced colitis. Specifically, these studies showed that CB2 activation suppressed inflammasome-mediated caspase-1 activation and the mechanism was partially due to the induction of autophagy (Shao et al., 2014; Ke et al., 2016), which has been identified as a key negative regulator of inflammasome activation (Sun et al., 2017; Takahama et al., 2018). A similar mechanism may be responsible for the effects observed here, because both THC and JWH-015 suppressed TLR7-mediated monocyte secretion of IL-1β.
A limitation in this investigation is whether the concentrations of THC used in this study are relevant to those present within the brain of cannabis users. Because it is not feasible to directly quantify THC concentrations in the brain immediately after cannabis use by humans, we based the in vitro concentrations in this report on the peak plasma concentrations, which range from 0.24 to 0.87 μM after inhalation of a cannabis cigarette containing 3.55% THC (Huestis, 2007). As discussed above, this concentration range has the potential to be elevated due to the increased THC content in present-day cannabis preparations (ElSohly et al., 2016). Second, the focus of this study was on the in vitro effects of THC; however, the in vivo effects of THC when inhaled through the use of cannabis may differ from the results observed here. This could be due to cannabis containing more than 100 structurally related cannabinoids as well as other plant-derived constituents (e.g., terpenes) that may alter the actions of THC or possess their own immunomodulatory effects (National Academies of Sciences, Engineering, and Medicine, 2017). Finally, the coculture used in this study only examines the role of THC on monocyte-mediated astrocyte inflammation; however, multiple other cell types (e.g., microglia, pericytes, and endothelial cells) are present in the brain and have inflammatory functions (Jansson et al., 2014; O’Carroll et al., 2015). Thus, future studies will be needed to address the effect of THC on the inflammatory crosstalk between other cell types.
Authorship Contributions
Participated in research design: Rizzo, Crawford, Bach, Sermet, Amalfitano, Kaminski.
Conducted experiments: Rizzo, Bach, Sermet.
Performed data analysis: Rizzo, Crawford, Kaminski.
Wrote or contributed to the writing of the manuscript: Rizzo, Amalfitano, Kaminski.
Footnotes
- Received June 7, 2019.
- Accepted August 1, 2019.
This work was supported by the National Institutes of Health National Institute on Drug Abuse [Grant R01-DA047180] and the National Institutes of Health National Institute of Environmental Health Sciences [Grant T32-ES07255].
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- 7-AAD
- 7-aminoactinomycin D
- ABM
- astrocyte basal media
- CB
- cannabinoid receptor
- CNS
- central nervous system
- HIV
- human immunodeficiency virus
- IL
- interleukin
- MCP
- monocyte chemoattractant protein
- MFI
- mean fluorescence intensity
- NFκB
- nuclear factor-κB
- NHA
- normal human astrocyte
- PCR
- polymerase chain reaction
- R837
- imiquimod
- THC, Δ9-tetrahydrocannabinol
- THC-COOH
- 11-nor-9-carboxy-Δ
- 9-tetrahydrocannabinol
- TLR
- Toll-like receptor
- TNF
- tumor necrosis factor
- Copyright © 2019 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}