


Abstract
GABAA receptors containing α5 subunits (GABAA α5) are highly expressed in the hippocampus and negatively involved in memory processing, as shown by the fact that GABAA α5–deficient mice show higher hippocampus-dependent performance than wild-type mice. Accordingly, small-molecule GABAA α5 negative allosteric modulators (NAMs) are known to enhance spatial learning and memory in rodents. Here we introduce a new, orally available GABAA α5 NAM that improves hippocampal functions. ONO-8590580 [1-(cyclopropylmethyl)-5-fluoro-4-methyl-N-[5-(1-methyl-1H-imidazol-4-yl)-2-pyridinyl]-1H-benzimidazol-6-amine] binds to the benzodiazepine binding sites on recombinant human α5–containing GABAA receptors with a Ki of 7.9 nM, and showed functionally selective GABAA α5 NAM activity for GABA-induced Cl− channel activity with a maximum 44.4% inhibition and an EC50 of 1.1 nM. In rat hippocampal slices, tetanus-induced long-term potentiation of CA1 synapse response was significantly augmented in the presence of 300 nM ONO-8590580. Orally administered ONO-8590580 (1–20 mg/kg) dose-dependently occupied hippocampal GABAA α5 in a range of 40%–90% at 1 hour after intake. In the rat passive avoidance test, ONO-8590580 (3–20 mg/kg, by mouth) significantly prevented (+)-MK-801 hydrogen maleate (MK-801)–induced memory deficit. In addition, ONO-8590580 (20 mg/kg, p.o.) was also effective in improving the cognitive deficit induced by scopolamine and MK-801 in the rat eight-arm radial maze test with equal or greater activity than 0.5 mg/kg donepezil. No anxiogenic-like or proconvulsant effect was associated with ONO-8590580 at 20 mg/kg p.o. in the elevated plus maze test or pentylenetetrazole-induced seizure test, respectively. In sum, ONO-8590580 is a novel GABAA α5 NAM that enhances hippocampal memory function without an anxiogenic or proconvulsant risk.
Introduction
Because of the advent of this rapidly aging society, the number of adults with senile dementias, such as Alzheimer’s disease (AD), has increased (Akagi et al., 2015). Currently approved pharmacological treatments for AD are limited to cholinesterase inhibitors as well as the N-methyl-d-aspartate (NMDA) receptor antagonist, which acts on the glutamatergic pathway (Farlow et al., 2008). The limited efficacy of these drugs highlights the need for better treatment. Despite the tremendous efforts in search of disease-modifying agents focusing on the β-amyloid or tau pathways, none are clinically available. Therefore, a strong cognitive enhancer with a novel mode of action is still globally desirable.
GABA is a major inhibitory neurotransmitter in the adult brain. In patients with AD as well as in transgenic AD mouse models, altered GABAergic function has been reported. Levels of GABA in cerebrospinal fluid (CSF) (Samakashvili et al., 2011) and reactive astrocytes (Jo et al., 2014) are elevated in patients with AD. The GABAA receptor containing α5 subunits (GABAA α5) chloride channel, highly expressed in the hippocampus, which is involved in learning and memory (Quirk et al., 1996), is upregulated in the hippocampus in patients with AD (Kwakowsky et al., 2018) and in the 5xFAD mouse, a transgenic model for AD (Wu et al., 2014). GABAA α5–deficient mice show improved performance in the water maze test for studying spatial learning compared with wild-type mice (Collinson et al., 2002). Thus, the overactivation of the GABAA α5–mediated signaling pathway should contribute to the inhibition of neuronal activity and impairment of learning and memory in patients with AD. Therefore, agents with GABAA α5 inhibitory activity are expected to improve cognitive disorders like AD.
Studies in molecular genetic (mice with a point mutation in the GABAA α subunit) or pharmacological approaches suggest that GABAA α1 mediates the sedative effects of diazepam, whereas GABAA α2 and α3 accounts for anxiolytic and myorelaxant effects, respectively (Rudolph et al., 1999; McKernan et al., 2000; Rudolph and Möhler, 2004; Atack et al., 2005). The contribution of each subtype for the physiologic function has been elucidated by the genetically modified mice functionally lacking the diazepam binding site of the α1, α2, α3, or α5 subtype. These results suggest that α1 is responsible for the proconvulsant effect (Vergnes et al., 2001) and α2/3 for the anxiogenic effect (Horowski and Dorrow, 2002) of nonselective GABAA negative allosteric modulators (NAMs). Whereas GABAA positive allosteric modulators (PAMs) binding to the benzodiazepine (BZ) site, such as diazepam, increase the GABA response through GABAA α1, α2, α3, or α5 (α4 and α6 are diazepam insensitive), nonselective NAMs such as DMCM (methyl-6,7-dimethoxyl-4-ethyl-carboline-3-carboxylate) and N-methyl-β-carboline-3-carboxamide (FG-7142) decrease GABA response, resulting in a membrane depolarization and increased neuronal excitability (Haefely et al., 1993). Therefore, the opposing effects of PAMs and NAMs at the molecular level are reflected behaviorally in that NAMs are anxiogenic and proconvulsant effects (Haefely et al., 1993). The anxiogenic and proconvulsant liabilities of the nonselective NAMs prevent clinical application (Dorow et al., 1983). It was therefore hypothesized that a GABAA α5–selective NAM should improve cognitive impairment without anxiogenic and proconvulsant effects.
The BZ binding site, an allosteric site on the GABAA receptor, has the potential to obtain GABAA subtype–selective PAMs and NAMs (Atack, 2011). Approximately 124,000 compounds (Charles River Laboratories International, Inc., Essex, UK) were screened in both receptor binding and FLIPR Functional Assays (Molecular Devices, LLC, San Jose, CA) in human GABAA α5β3γ2 expressing the HEK293 cell line. As a result, a novel structural class of BZ site ligand with the potential to selectively modulate GABAA α5 function was identified. In this study, the efficacy of 1-(cyclopropylmethyl)-5-fluoro-4-methyl-N-[5-(1-methyl-1H-imidazol-4-yl)-2-pyridinyl]-1H-benzimidazol-6-amine (ONO-8590580), a GABAA α5 NAM with a novel chemotype, which was obtained after lead optimization from hit compounds, was evaluated.
Materials and Methods
Compounds.
ONO-8590580 (Fig. 1) (purity, ≥95%) was synthesized at ONO Pharmaceutical Co., Ltd. (Osaka, Japan). Donepezil, (−)-scopolamine hydrobromide trihydrate (scopolamine), (+)-MK-801 hydrogen maleate (MK-801), FG-7142, pentylenetetrazole (PTZ), and dimethylsulfoxide (DMSO) were purchased from Sigma-Aldrich (St. Louis, MO). L-655,708 (C18H19N3O4), a selective GABAA α5 ligand (Quirk et al., 1996), was purchased from Santa Cruz Biotechnology (Dallas, TX). ONO-8590580 and L-655,708 used for in vivo studies was first dissolved in DMSO and then added to WellSolve (Celeste Corporation, Tokyo, Japan) and water (final composition ratio of the solution: 2% DMSO, 20% WellSolve, 78% water). Scopolamine, MK-801, and PTZ was dissolved in saline. Donepezil was dissolved in water. FG-7142 was suspended in 0.1 vol% polyoxyethylene (20) sorbitan monooleate (Tween 80; Wako Pure Chemical Industries, Ltd., Osaka, Japan) in saline.

Chemical structure of ONO-8590580.
Animals.
All experimental procedures were approved by the institutional animal care and use committee of ONO Pharmaceutical Co., Ltd. All efforts were made to minimize the number of animals used and their suffering. Male Sprague Dawley rats (Crl:CD(SD), 6 weeks old; Charles River Laboratories Japan, Inc.) were used in the long-term potentiation (LTP) and in vivo receptor occupancy studies. Male Sprague Dawley rats (Crl:CD(SD), 7 weeks old; Charles River Laboratories Japan, Inc.) were used in the passive avoidance test and the elevated plus maze test. Male Wistar rats (Crlj:WI, 7–9 weeks old; Charles River Laboratories Japan, Inc.) were used for the eight-arm radial maze test. Male ICR mice (Crlj:CD1(ICR), 7–9 weeks old; Charles River Laboratories Japan, Inc.) were used for the PTZ proconvulsant test. Animals were housed in groups of less than five animals/cage in a temperature- and humidity-controlled animal room (temperature, 24 ± 2°C; relative humidity, 55% ± 15%) under a 12-hour light/dark cycle (light on from 8:00 AM to 8:00 PM). Food and water were available ad libitum.
Cell Cultures.
Human GABAA α1β3γ2 expressing the HEK293 cell line (catalog number CYL3053; Merck Millipore, Burlington, MA), human GABAA α2β3γ2 expressing the HEK293 cell line (catalog number CYL3072; Merck Millipore), human GABAAα3β3γ2 expressing the HEK293 cell line (catalog number CYL3068; Merck Millipore), and human GABAA α5β3γ2 expressing the HEK293 cell line (catalog number CYL3073; Merck Millipore) were cultured at 37°C (5% CO2, 95% air) until confluence in Dulbecco’s modified Eagle’s medium/Ham’s F-12 medium with l-glutamine (Invitrogen, Carlsbad, CA), supplemented with 10% fetal bovine serum (Invitrogen), 1% nonessential amino acids (Invitrogen), 400 μg/ml Geneticin (Invitrogen), 100 μg/ml Hygromycin B (Invitrogen), and 0.625 μg/ml Puromycin (Clontech, Mountain View, CA).
In Vitro Radioligand Binding Studies.
The GABAA α1β3γ2, α2β3γ2, α3β3γ2, and α5β3γ2 protein used for the scintillation proximity assay (SPA) was derived from membranes produced from the cell line expressing each complex. Membranes were prepared as previously published (Hadingham et al., 1993). Briefly, after routine culture in T175 flasks (CellBIND Surface; Corning Inc., Corning, NY), cells were washed with Dulbecco’s phosphate-buffered saline (PBS; Invitrogen), scraped from the flask into PBS, and pelleted by centrifugation (500g). The cells were resuspended in 10 mM potassium phosphate at pH 7.4 and homogenized (5000 rpm, 15 seconds) using a Precellys Tissue Homogenizer (Precellys 24-Dual; Bertin Technologies, Thiron-Gardais, France). The homogenates were centrifuged at 48,000g for 30 minutes, resuspended, and washed twice. The final resuspension of membranes took place in buffer containing 10 mM potassium phosphate and 100 mM KCl. The protein concentration in membrane preparations was confirmed using a BCA (bicinchoninic acid) Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA).
ONO-8590580 was diluted in 100% DMSO. For the SPA, 0.5 μl of ONO-8590580 per well was added to a white-walled, clear-bottomed Matrix 384-well plate (Thermo Fisher Scientific). The assay controls used to calculate the percentage of inhibition consisted of DMSO and 100 μM flumazenil (final concentration, 2 μM), both 0.5 μl per well, for full signal and full block, respectively. Protein (10 μl) was added to the plate at a concentration of 2.5 μg/well followed by 9.5 μl of polyvinyltoluene-wheat germ agglutinin beads at a concentration of 0.2 mg/well. Both protein and bead were diluted to the desired concentration with 10 mM potassium phosphate, pH 7.4, containing 100 mM KCl. The reagents were preincubated for 30 minutes at room temperature with shaking before initiation of the reaction by the addition of a radioligand. The ability of ONO-8590580 to inhibit the binding of either 30 nM [3H]-Ro15-1788 (PerkinElmer LAS GmbH, Rodgau, Germany) to GABAA α1, α2, or α3 or 2 nM [3H]-Ro15-4513 (PerkinElmer LAS GmbH) to GABAA α5 was measured. From the EC50, the Ki (inhibitory constant) was calculated from the Cheng-Prusoff equation (Cheng and Prusoff, 1973), using Kd (dissociation constant) for the binding of [3H]-Ro15-1788 to GABAA α1, α2, and α3 at concentrations of 13, 15, and 15 nM, respectively, and for the binding of [3H]-Ro15-4513 to GABAA α5 at a concentration of 2.0 nM. [3H]-Ro15-1788 5 μl was added at a concentration of 30 nM (final concentration, 6 nM), giving a final reaction volume of 25 μl and a DMSO concentration of 2% v/v. The plates were sealed, and the reaction mix was incubated at room temperature for a minimum of 2 hours with gentle agitation on a plate shaker. At the end of the incubation period, the plates were centrifuged for 2 minutes at 1000 rpm prior to reading on a PerkinElmer Microbeta. For each compound, the EC50 and Ki values were determined, the data were entered in Activity Base (IDBS), and the curves were fitted to the mean data. The Z′ factor values in the binding assays for GABAA α1, α2, α3, and α5 were 0.53, 0.53, 0.50, and 0.54, respectively.
In Vitro Efficacy.
Current recordings were performed using PatchXpress (Molecular Devices, LLC) with HEK293 cell lines expressing human GABAA α1, α2, α3, or α5 subunits associated with β3γ2 subunits. Cells were dissociated from poly-d-lysine–treated tissue culture flasks using a 1:1 mixture of TrypLE (Invitrogen) and Dulbecco’s PBS (Invitrogen). The dissociated cells were resuspended in Dulbecco’s modified Eagle’s medium/Ham’s F-12 medium (Invitrogen) supplemented with 10% fetal bovine serum (Invitrogen) and 1% nonessential amino acids (Invitrogen), and were allowed to recover for 1 hour at 37°C in a humidified incubator with 5% CO2. At the end of the recovery period, cells were centrifuged at 1000 rpm for 2 minutes, and the pellet was resuspended and diluted to 2.5 × 106 cells/ml in a solution (137 mM NaCl, 4 mM KCl, 10 mM HEPES, 1.8 mM CaCl2, 1 mM MgCl2, and 10 mM glucose, pH 7.35). Current responses were recorded in the presence of concentration-matched DMSO (0.3% v/v). Patch pipettes were filled with a solution composed 90 mM KCl, 50 mM KF, 11 mM EGTA, 10 mM HEPES, 1 mM MgCl2, and 2 mM Mg-ATP (pH 7.35). The holding potential was −60 mV. Control GABA responses were first determined at their concentration of GABA giving a current that was 20% of the maximum (EC20 concentration), and a test compound was preapplied for 1 minute prior to the addition of the EC20 concentration of GABA (GABAA: α1, 2.4 μM; α2, 1.5 μM; α3, 2.3 μM; α5, 0.34 μM). The test compound and/or GABA were applied for 5 seconds, with 2-minute wash periods between applications. To calculate EC50 values for ONO-8590580, GraphPad Prism software (version 5.01; GraphPad Software Inc., San Diego, CA) was used.
Long-Term Potentiation in Rat Brain Slice.
LTP was measured electrophysiologically using a multielectrode array system (MED64; Alpha MED Scientific Inc., Osaka, Japan), as described previously (Tsukamoto et al., 2003). Brain slices were prepared from rats in ice-cold artificial CSF, consisting of 124 mM NaCl, 3 mM KCl, 26 mM NaHCO3, 2 mM CaCl2, 1 mM MgSO4, 1.25 mM KH2PO4 and 10 mM glucose, pH 7.4. Parasagittal 350-μm-thick brain slices were cut on a Vibratome sectioning system, and the hippocampal regions were placed at the center of chamber with 64 embedded recording electrodes (MED64; Alpha MED Scientific Inc.) and perfused with 95% O2/5% CO2–saturated artificial CSF for at least 1 hour at 32°C. One of 64 planar microelectrodes was used to stimulate the Schaffer collaterals every 30 seconds. Field excitatory postsynaptic potentials (fEPSPs) were evoked by the stimulation of a Schaffer collateral-commissural pathway with a stimulus intensity set to produce a half-maximal slope of fEPSP. Synaptic strength was evaluated by measuring changes in the fEPSP slopes. To induce LTP, a θ-burst protocol (four pulses delivered with a frequency of 100 Hz, repeated 10 times with an interval of 200 milliseconds) was used.
In Vivo Receptor Occupancy.
The occupancy of the BZ binding site of rat brain GABAA α5 was measured using an in vivo [3H]-Ro15-4513 binding assay (n = 3–4 in each group). ONO-8590580 was orally administered 1 hour before sacrifice. [3H]-Ro15-4513 (1.11 MBq/kg) was administered via the tail vein (1 ml/kg) 10 minutes before sacrifice. Rats were decapitated, and the brains were rapidly removed and homogenized in ice-cold assay buffer (10 mM KH2PO4, 10 mM K2HPO4, 100 mM KCl). Aliquots of homogenate were filtered and washed over GF/B Glass Fiber Filters (Brandel Inc., Gaithersburg, MD). To determine the amount of nonspecific [3H]-Ro15-4513 binding, a separate group of rats were pretreated for 30 minutes with L-655,708 (10 mg/kg, i.p.). The signal-to-background ratio was 26. The amount by which ONO-8590580 reduced the specific binding of [3H]-Ro15-4513 relative to the binding in nontreated rats was defined as the occupancy.
Passive Avoidance Test.
The test was performed as described previously (Mitsui et al., 2015). The step-through–type passive avoidance apparatus (Brain Science Idea Co., Ltd., Osaka, Japan) consisted of an illuminated compartment (height, 28 cm; length, 25 cm; width, 11 cm; 700–750 lux) connected to a darkened compartment (height, 30 cm; length, 32 cm; width, 33 cm; 0.03–0.05 lux) by a guillotine door. The experiment consisted of two trials: an acquisition trial and a retention trial (n = 10–15 in each group). In the acquisition trial, the rat was placed in the illuminated compartment 30 minutes after administration of MK-801 (0.1 mg/kg, s.c.) or saline. The guillotine door was then opened, and the rat was allowed to enter the dark compartment. The latency to enter the dark compartment was recorded (step-through latency). Once the rat entered the dark compartment, the door was closed and an electric foot shock was immediately applied (1 second, 2.2 mA). On the following day, the rat was placed in the illuminated compartment and again allowed to enter the dark compartment by opening the guillotine door. The step-through latency was once again recorded (retention trial). The maximum cutoff time for step-through latency was set at 300 seconds. During the retention trial, no foot shock was applied. Impairment of learning and memory was defined by a decrease in the time of step-through latency in the retention trial. ONO-8590580 or vehicle was administrated 1 hour before both an acquisition trial and a retention trial.
Eight-Arm Radial Maze Test.
The test was performed as described previously (Ohta et al., 1993). Briefly, rats (n = 10 in each group) were fed at a rate of approximately 13 g/day per rat (approximately 80% of normal daily feed intake). A daily training trial was carried out more than 10 times to allow the rats to learn how to perform the radial maze test. The trial was judged complete when the rat had chosen all eight baited arms or had spent 10 minutes in the maze. Entry into an arm that had not been previously visited was recorded as a correct response, and re-entry was counted as an error. The last training trial and the test trial to evaluate compounds were carried out in the same day. When a rat made no errors or only one error after the seventh choice in the last training trial, it was considered ready for compound testing. In the test trial, MK-801 and/or scopolamine were subcutaneously administered 0.5 hour before the test trial. ONO-8590580, donepezil, or vehicle was orally administered 1 hour before the trial. A rat was placed on the central hub of the maze and allowed to visit the wells at the end of each of eight arms. The number of errors (the number of repeat entries to arms of the maze already visited) and the latency until the bait of all eight wells had been consumed were counted.
Elevated Plus Maze.
Rats (n = 12 in each group) were given vehicle, ONO-8590580 (20 mg/kg, p.o.), or, as a positive control, the GABAA nonselective NAM FG-7142 (15 mg/kg, i.p.). After 1 hour, rats were placed in the elevated plus maze for 5 minutes. Light intensity in the open arms was set at 20 lux. A video camera fitted with a polarizing lens was mounted above the maze, connected to a tracking and analysis system (EthoVision XT; Noldus Information Technology Inc., Leesburg, VA). The open and closed arms (each 10 × 50 cm) and the central area (10 × 10 cm) of the plus maze were defined using the tracking system. The effect of ONO-8590580 on the time spent on the open arms was assessed.
PTZ Proconvulsant Test.
Mice (n = 8 in each group) were intraperitoneally administered vehicle, ONO-8590580 (10 mg/kg), or FG-7142 (10 mg/kg). After 30 minutes, the mice were infused with a 15 mg/ml PTZ solution (infusion rate, 0.2 ml/min) and the time taken to reveal clonic seizures was measured, and from this the dose administered was calculated.
Statistical Analysis.
Except where noted, data are expressed as the mean ± S.E.M. Statistical analyses were performed using SAS version 9.2 TS2M3 (SAS Institute Japan Inc., Tokyo, Japan) and its cooperative system EXSUS version 7.7.1 (CAC Corporation, Tokyo, Japan) with Student’s t test, Wilcoxon rank-sum test, Dunnett’s test, or Steel test, as appropriate. Tests were two sided and conducted at a 5% level of significance.
Results
In Vitro Binding of ONO-8590580 to GABAA Receptors.
Inhibition of [3H]-Ro15-4513 binding showed that ONO-8590580 binds with high affinity to GABAA α5 (Ki = 7.9 nM) (Fig. 2). The inhibition of [3H]-Ro15-1788 binding showed that ONO-8590580 also binds to GABAA α1, α2, and α3 with Ki values of 140, 32, and 24 nM, respectively (Fig. 2).

In vitro binding affinity of ONO-8590580 at human recombinant GABAA receptors. HEK293 cell lines stably expressing human GABAA α1, α2, α3, and α5β3γ2 were used in an SPA. Affinity at GABAA α1, α2, and α3 was measured using a [3H]-Ro15-1788 binding assay, whereas affinity at GABAA α5 was measured using [3H]-Ro15-4513.
Efficacy of ONO-8590580 on Each Subtype of GABAA Receptors.
ONO-8590580 concentration-dependently inhibited the current induced by the EC20 concentration of GABA in human GABAA α5β3γ2 expressing the HEK293 cell line (Fig. 3). The calculated maximum inhibition was −44.4%, and the EC50 and Hill slope values were −1.1 nM and −0.50, respectively. ONO-8590580 was not effective or only slightly effective in GABAA α1, α2, and α3 (Fig. 3).

Efficacy of ONO-8590580 on human recombinant GABAA receptors. HEK-293 cell lines stably expressing human GABAA α1, α2, α3, (n = 3), and α5β3γ2 (n = 5) were used in a patch-clamp study. The ability of ONO-8590580 to attenuate or potentiate the current induced by the EC20 concentration of GABA was assessed by whole-cell patch-clamp electrophysiology. Data are expressed as the mean and S.E.M.
Effect on LTP in Rat Hippocampal Slice.
LTP was induced using a standard paradigm that involves a θ-burst stimulus (Fig. 4). In control experiments, the fEPSP slopes were enhanced after the θ-burst compared with the baseline, then returned to baseline 1 hour after stimulation. In the presence of ONO-8590580 (300 nM), the slopes after the θ-burst were also enhanced compared with the baseline, but did not return to baseline values 1 hour after stimulation. Therefore, ONO-8590580 significantly induced LTP after a θ-burst in the CA1.

Effect of ONO-8590580 (300 nM) on LTP in rat hippocampal slice. The fEPSPs in response to stimulation of the Schaffer collateral-commissural pathway were recorded. To produce LTP, a θ-burst protocol (four pulses delivered with a frequency of 100 Hz, repeated 10 times with an interval of 200 milliseconds) was used. Data are expressed as the mean and S.E.M. (n = 7–8 slices). *P < 0.05 compared with the control group (Student’s t test). ONO, ONO-8590580.
In Vivo Receptor Occupancy in Rat Hippocampus.
ONO-8590580 competed the binding of [3H]-Ro15-4513, a GABAA α5 specific binder, in the rat hippocampus after oral administration (Fig. 5). The ratio of GABAA α5 occupancy values produced by ONO-8590580 at 1, 3, 10, and 20 mg/kg were 44%, 53%, 71%, and 89%, respectively. The ED50 value of ONO-8590580 with 95% confidence intervals was 1.9 mg/kg (1.0 2–3.72 mg/kg).

Receptor occupancy of ONO-8590580 on rat hippocampal GABAA α5. ONO-8590580 at 1, 3, 10, and 20 mg/kg was orally administered 1 hour before sacrifice. [3H]-Ro15-4513 was intravenously administered 10 minutes before sacrificing the rats. Data are expressed as the mean and S.E.M. (n = 3–4).
Effect on MK-801–Induced Cognitive Deficit in Passive Avoidance Test.
MK-801 (a NMDA antagonist)–treated rodents have been validated as an animal model of cognitive dysfunction associated with dementia (van der Staay et al., 2011). It is reported that negative modulation of GABAA α5 in rats partially prevents the memory impairment induced by MK-801 (Timić Stamenić et al., 2015). Therefore, we evaluated the effect of ONO-8590580 on MK-801–induced cognitive deficit in the passive avoidance test. The latencies in the acquisition trial did not significantly differ regardless of the treatment administered 1 hour before the trial (data not shown). ONO-8590580 (3, 10, and 20 mg/kg) significantly increased the escape latency compared with control in the retention trial (Fig. 6).
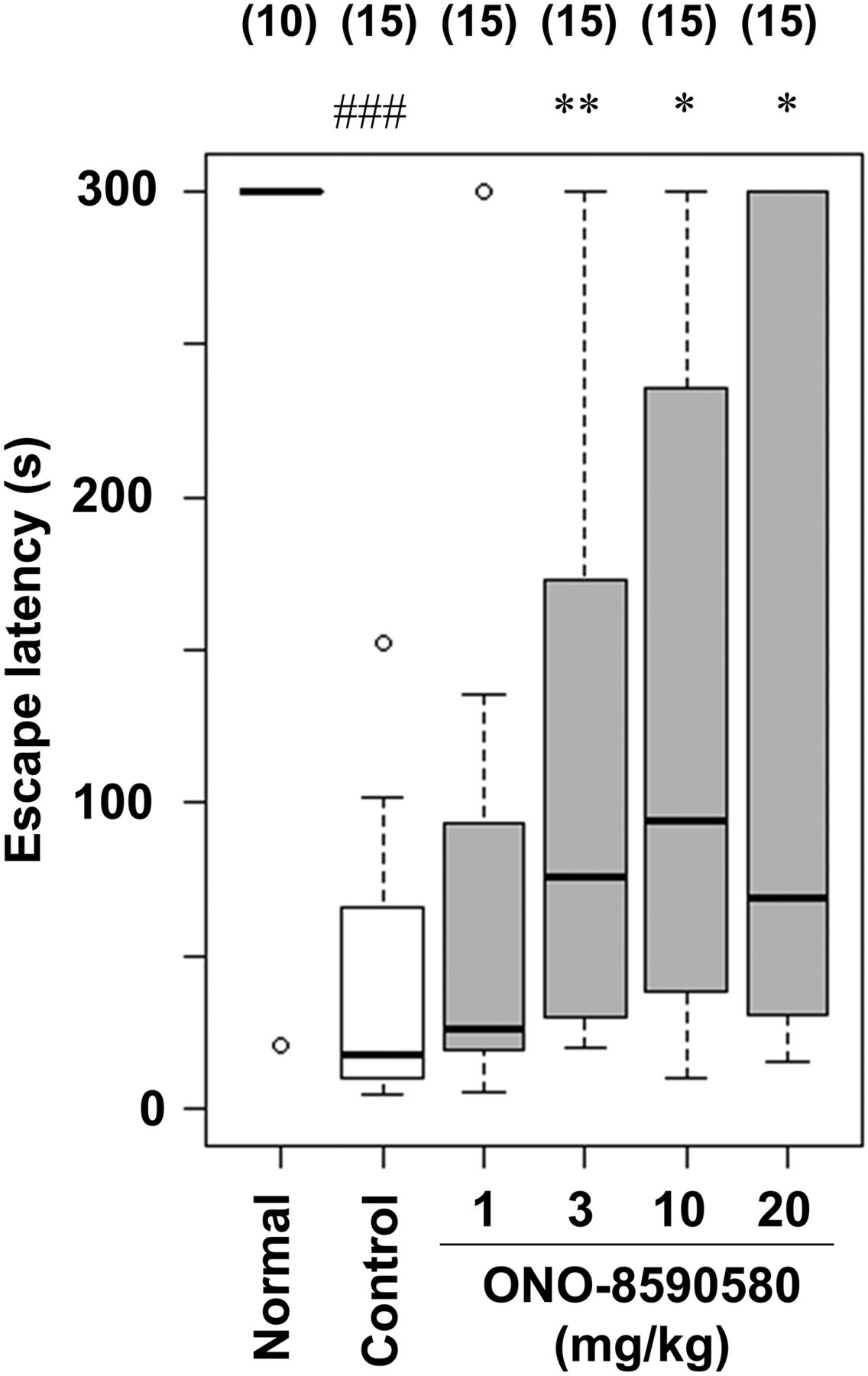
Effect on MK-801–induced cognitive deficit in rat passive avoidance test. On day 1 (acquisition trial), MK-801 (0.1 mg/kg, s.c.) was administered to rats 30 minutes before the trial. A retention trial was conducted 24 hours after the acquisition trial. Vehicle or ONO-8590580 was orally administered 1 hour before the acquisition and retention trial. Data are represented by box plot (n = 10–15 in each group). ###P < 0.001 compared with Normal group (Wilcoxon rank-sum test). *P < 0.05; **P < 0.01 compared with the control group (Steel test).
Effect on MK-801/Scopolamine-Induced Cognitive Deficit in Eight-Arm Radial Maze Test.
Patients with AD have both cholinergic neuron degeneration (Everitt and Robbins, 1997; Schliebs and Arendt, 2006) and hypofunction of glutamatergic neurotransmission (Hardy et al., 1987; Lin et al., 2014) in the brain. Therefore, the cognitive deficit in rats induced by the combination of scopolamine, a muscarinic antagonist, and MK-801 would be useful as a model of AD dementia. We investigated the effect of ONO-8590580 on the MK-801/scopolamine-induced cognitive deficit in the eight-arm radial maze test, as described by Li et al. (1996). Doses of MK-801 and scopolamine for this test were 0.075 (i.p.) and 0.2 mg/kg (i.p.), respectively, because we confirmed that each single administration at these doses increased the number of errors respectively, and that in combination there was a stronger effect than for each single administration (data not shown). As a positive control and for comparison purposes, a 0.5 mg/kg dose of donepezil (p.o.), which decreases both the number of errors and the running time in the scopolamine-induced cognitive deficit in the eight-arm radial maze test (Sugimoto et al., 2002), was also evaluated. ONO-8590580 (20 mg/kg, p.o.) significantly decreased the number of errors and the total latency compared with the control (Fig. 7). Donepezil (0.5 mg/kg, p.o.) also decreased total latency, but not the number of errors compared with the control.

Effect on MK-801/scopolamine-induced cognitive deficit in the rat eight-arm radial maze test. Vehicle, ONO-8590580, or donepezil (DNP) was administered orally 1 hour before the trial. MK-801 (0.05 mg/kg) and scopolamine (0.15 mg/kg) were administered subcutaneously 0.5 hour before the test trial. The number of errors and the latency until the bait of all eight wells had been consumed were counted. Data are expressed as the mean and S.E.M. (n = 10 in each group). **P < 0.01 compared with the control group (Dunnett test). $P < 0.05 compared with the control group (Student’s t test).
Effect in Elevated Plus Maze Test and PTZ Test.
In the rat elevated plus maze test, the nonselective GABAA NAM FG-7142 significantly decreased the time spent on the open arms, whereas ONO-8590580 had no effect compared with vehicle-treated animals (Fig. 8A).

No anxiogenic-like effect in the rat elevated plus maze test (A) and no proconvulsant effect in the mouse PTZ test (B). (A) Vehicle (p.o.), ONO-8590580 (20 mg/kg, p.o.), or FG-7142 (15 mg/kg, i.p.) was administered 1 hour before the test. One hour after administration, rats were given a 5-minute trial on the elevated plus maze, and the time spent on the open arm was calculated. (B) Vehicle (i.p.), ONO-8590580 (10 mg/kg, i.p.), or FG-7142 (10 mg/kg, i.p.) was administered 30 minutes before the test. Mice were infused intravenously with PTZ via the tail vein. Data are expressed as the mean and S.E.M. (n = 12 in each group in elevated plus maze test, n = 8 in each group in PTZ test). $P < 0.05 compared with the control group (Student’s t test).
In the mouse PTZ test, FG-7142 decreased the dose of PTZ required to induce clonic seizure, whereas ONO-8590580 had no effect on the threshold for PTZ-induced clonic seizure (Fig. 8B).
Discussion
The present study demonstrated that ONO-8590580, a functionally selective GABAA α5 NAM, significantly enhanced LTP in rat hippocampal slices and improved cognitive deficits in rats without anxiogenic-like or proconvulsant effects.
ONO-8590580 has only 3.3–17.7 times higher binding selectivity for GABAA α5 compared with GABAA α1, α2, and α3, but it shows very high functional selectivity for GABAA α5. This in vitro profile is similar to those for α5IA (Dawson et al., 2006) and MRK-016 (Atack et al., 2009), but differs from that of RO4938581, which has both binding selectivity and function selectivity for GABAA α5 (Ballard et al., 2009). It is not known whether compounds display different phenotypes in vivo depending on the presence or absence of binding selectivity to GABAA α5. It is reported that there are endogenous ligands to the BZ binding site: endozepines (Farzampour et al., 2015). This finding indicates that different phenotypes may be shown by each compound with and without binding selectivity. Among 82 off-target agents, ONO-8590580 bound only to the adenosine A3 receptor with an inhibition of 68% at a drug concentration of 10 μM; thus, ONO-8590580 is highly selective for GABAA α5 (Eurofins Panlabs Inc., Taipei, Taiwan; data not shown).
The physiologic properties of hippocampal-related cognitive process may involve long-term changes in synaptic efficacy, such as in LTP (Bliss and Collingridge, 1993). Nonselective GABAA NAMs increase LTP (Seabrook et al., 1997), whereas nonselective GABAA PAMs impair LTP (del Cerro et al., 1992). In this study, the selective GABAA α5 NAM enhanced LTP after a θ-burst in a rat hippocampal slice. This result is consistent with other reports showing that other GABAA α5 NAMs enhance LTP (Dawson et al., 2006; Atack et al., 2009; Ballard et al., 2009). Etomidate, which has memory-blocking properties, inhibited LTP, and this effect was reversed by L-655,708 (Martin et al., 2009). These findings indicate that GABAA α5 holds an important role for the enhanced LTP observed with nonselective GABAA NAMs. GABAA α5 is predominantly localized to extrasynaptic space in the hippocampus (Farrant and Nusser, 2005; Brickley and Mody, 2012). It has been reported that tonic inhibition mediated by GABAA α5 affects the induction and maintenance of LTP (Ge et al., 2008; Martin et al., 2010; Li et al., 2012). Since ONO-8590580 inhibits the function of GABAA α5, tonic inhibition may be suppressed and LTP may be consequently enhanced.
ONO-8590580 significantly improved MK-801–induced cognitive deficit in the rat passive avoidance test. MK-801 impairs contextual specificity of hippocampal immediate-early gene expression, which is critical for the maintenance of synaptic plasticity and memory consolidation (Kubik et al., 2014). GABAA α5 is expressed in the bases of the spines and the adjacent shafts of the dendrites (Fritschy and Brünig, 2003), and modulates the excitatory input arising at the spines via NMDA receptor (Glykys et al., 2008; Brickley and Mody, 2012). Therefore, it is suggested that ONO-8590580 improved MK-801–induced cognitive impairment indirectly via enhancing the excitatory input in hippocampal CA1 spines.
In patients with AD, the presence of not only cholinergic neuron degeneration in the nasal forebrain but also hypofunction of NMDA receptor–mediated neurotransmission in cortical and hippocampal regions in postmortem and imaging studies of the brains (Hardy et al., 1987; Bi and Sze, 2002; Lin et al., 2014). Blockage of NMDA receptor–mediated neurotransmission has been reported to prevent the induction of LTP in the hippocampus (Bashir et al., 1991; Seabrook et al., 1997). MK-801 is known to induce memory deficit in animals (Suryavanshi et al., 2014). In addition, the decrease in NMDA receptors in the hippocampus and entorhinal cortex of patients with AD has been reported to correlate with the disease neuropathological progression, as assessed by postmortem examination (Kravitz et al., 2013). Thus, we used a rat model with cognitive deficit induced by a cotreatment model of muscarinic receptor antagonist scopolamine (0.2 mg/kg) and the NMDA receptor antagonist MK-801 (0.075 mg/kg) as a model reflecting a part of AD dementia in the eight-arm radial maze test. The dose of donepezil that was effective for total latency was 0.5 mg/kg in this model, but that dose did not improve the number of errors. Donepezil at this dose significantly improves the cognitive deficit induced by scopolamine in 8-arm radial maze test (Sugimoto et al., 2002), and it also improves the cognitive deficit induced by lesions of the medial septum in the water maze test more effectively at a dose of 0.5 mg/kg than at 2 mg/kg (Ogura et al., 2000). Therefore, 0.5 mg/kg donepezil is an appropriate dose to investigate the efficacy of donepezil. The present study in which ONO-8590580, but not donepezil, significantly decreased the number of errors may suggest that ONO-8590580 could be more potent for the treatment of patients with AD. The data showing that ONO-8590580 has no anxiogenic-like or proconvulsant effects are in agreement with the behavioral phenotype of α5−/− mice (Collinson et al., 2002).
The receptor occupancy was 44%, 53%, 71%, and 89% at 1, 3, 10, and 20 mg/kg (p.o.), respectively. The effective doses of ONO-8590580 were 3–20 mg/kg (p.o.) in the passive avoidance test and 20 mg/kg (p.o.) in the eight-arm radial maze test. Axiogenic-like and proconvulsant effects were not observed at 20 mg/kg (p.o.) and 10 mg/kg (i.p.), respectively. From these results, the target level of occupancy for clinical studies might be 53%–89%.
In the brains of patients with AD, the number of reactive astrocytes increase (Jo et al., 2014) and phenotypically switch from GABA negative to GABA producing (Oh and Lee, 2017). Released GABA from reactive astrocytes to the extracellular space activates GABAA α5 and induces tonic inhibition (Farrant and Nuzzer, 2005; Kim et al., 2017). Tonic inhibition may suppress the activity of excitatory neurons in the hippocampus, decrease LTP, and impair learning and memory (Ge et al., 2008; Martin et al., 2010; Li et al., 2012). In accordance with previous findings (Dawson et al., 2006; Atack et al., 2009; Ballard et al., 2009), this study confirmed that a GABAA α5 NAM increases LTP and improves learning and memory. In addition, the GABAA α5 expression level in hippocampus increases in patients with AD (Kwakowsky et al., 2018). These results suggest that astrocytic GABA and extrasynaptic GABAA α5 are important players in the pathogenesis of AD, and that ONO-8590580, a GABAA α5 NAM, is an attractive agent with which to treat AD. Since reactive astrocytes have also been described in patients with Parkinson disease, stroke, epilepsy, brain trauma, and other neurodegenerative diseases (Luchetti et al., 2011; Brichta et al., 2013), ONO-8590580 could also be considered for the treatment of such neurodegenerative diseases as well as AD. Future work is needed to explore these exciting possibilities.
The development of another chemotype of GABAA α5 NAM, α5IA, was discontinued because of preclinical renal toxicity (Atack, 2010), and MRK-016 was poorly tolerated in the elderly volunteers (Atack, 2011). A clinical trial of RG1662 was conducted in subjects with Down syndrome (Costa and Scott-McKean, 2013), but there is no information in patients with AD.
We have shown in this study that ONO-8590580, a novel GABAA α5 NAM, will improve cognitive impairment without anxiogenic or proconvulsant side effects. These findings indicate that ONO-8590580 could be beneficial for preclinically evaluating cognitive disorders like AD.
Authorship Contributions
Participated in research design: Kawaharada, Yasuhiro, Clark, Maidment, Katsumata, and Kaneko.
Conducted experiments: Kawaharada, M. Nakanishi, N. Nakanishi, Hazama, Clark, and Maidment.
Contributed new reagents or analytic tools: Higashino, Lewis, and Chambers.
Performed data analysis: Kawaharada, M. Nakanishi, N. Nakanishi, Hazama, Clark, and Maidment.
Wrote or contributed to the writing of the manuscript: Kawaharada, Katsumata, and Kaneko.
Footnotes
- Received December 29, 2017.
- Accepted April 17, 2018.
This research was funded by ONO Pharmaceutical Co., Ltd. (Osaka, Japan).
Abbreviations
- AD
- Alzheimer’s disease
- BZ
- benzodiazepine
- CSF
- cerebrospinal fluid
- DMSO
- dimethylsulfoxide
- fEPSP
- field excitatory postsynaptic potential
- FG-7142
- N-methyl-β-carboline-3-carboxamide
- GABAAα5
- GABAA receptors containing α5 subunits
- HEK293
- human embryonic kidney 293
- L-655,708
- C18H19N3O4
- LTP
- long-term potentiation
- MK-801
- (+)-MK-801 hydrogen maleate
- NAM
- negative allosteric modulator
- NMDA
- N-methyl-d-aspartate
- ONO-8590580
- 1-(cyclopropylmethyl)-5-fluoro-4-methyl-N-[5-(1-methyl-1H-imidazol-4-yl)-2-pyridinyl]-1H-benzimidazol-6-amine
- PAM
- positive allosteric modulator
- PBS
- phosphate-buffered saline
- PTZ
- pentylenetetrazole
- SPA
- scintillation proximity assay
- Copyright © 2018 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}