


The concept that adenine and uridine nucleotides function as extracellular signaling molecules has been markedly expanded in the last decade. At least 15 nucleotide-activated cell surface receptors exist, and remarkably broad and varied physiological responses are known to occur downstream of nucleotide receptor activation. The significance of nucleotides as extracellular molecules also is underscored by ubiquitous distribution of several large classes of ectoenzymes that catalyze nucleotide breakdown and interconversion. Purinergic signaling was initially proposed on the basis of smooth muscle responses to autonomic nerve stimulation that were not blocked by adrenergic receptor antagonists (Burnstock, 1972; Burnstock, 1978; Burnstock and Kennedy, 1985). However, observation of responses to nucleotides in essentially all peripheral tissues including those not significantly innervated by the autonomic nervous system indicates that extracellular nucleotides arising from non-neuronal source underlie many important physiological processes (Ralevic and Burnstock, 1998). This Minireview focuses briefly on the receptors and ectoenzymes that mediate and terminate the action of extracellular nucleotides (Fig. 1) and summarizes the evidence regarding neuroendocrine secretion of ATP. We then consider in some detail current understanding of nucleotide release from nonexcitable tissues.

Interplay of released nucleotides, nucleotide metabolism, and activity of P2Y, P2X, and adenosine receptors. Released ATP, UTP, and UDP-glucose activate P2Y2, P2Y4, P2Y11, or P2Y14 receptors and ATP activates all P2X receptors. E-NTPDases generate ADP and UDP, transiently providing agonists for P2Y1 and P2Y6 receptor activation, respectively. Eventually all NTPs, NDPs, and UDP-sugars are degraded to NMPs by E-NTPDases and E-NPPs. Adenosine, the final product of adenine nucleotide hydrolysis, activates A1, A2a, A2b, and A3 adenosine receptors. Not depicted, 5′nucleotidase converts AMP to adenosine, NDPK and adenylyl kinase reversibly re-phosphorylate NDPs and AMP, respectively, and adenosine deaminase converts adenosine to inosine. ADO, adenosine; UDP-glu, UDP-glucose; URID, uridine; INOS, inosine.
Extracellular Nucleotides Activate Two Types of P2 Receptors
Nucleotide action is mediated by a large group of cell surface receptors included in the P2X and P2Y receptor families (Table 1). The P2X receptors are ligand-gated ion channels that gate extracellular cations in response to ATP. This family comprises seven receptors (P2X1 through P2X7) (North, 2002), and the possibility exists that these receptors form hetero-oligomers exhibiting pharmacological or regulatory properties distinct from those of the seven different homo-oligomeric forms. P2X receptors account for fast neurotransmission and largely, but not exclusively, are found on excitatory tissues. For example, the P2X7 receptor acts as an ATP-activated ion channel but also forms a pore, gating passage of molecules up to 1000 Da in response to ATP (Nuttle and Dubyak, 1994; Rassendren et al., 1997). This unique receptor plays an important role in cells involved in immunological and inflammatory responses. The P2Y receptors are G protein-coupled receptors that are categorized into a subfamily of receptors (P2Y1, P2Y2, P2Y4, P2Y6, and P2Y11) that predominantly couple to Gq, and therefore activate phospholipase C-β, and into a family of Gi-coupled receptors (P2Y12, P2Y13, and P2Y14) that inhibit adenylyl cyclase and regulate ion channels (Abbracchio et al., 2003).
The families of P2X and P2Y receptors
The IUPHAR recommendations for nomenclature of mammalian P2Y receptors are adopted here. Additional gene products tentatively named p2y5, p2y7, p2y9, and p2y10 receptors display no functional responses to nucleotides. The p2y3 receptor is the avian ortholog to the P2Y6 receptor. The Xenopus laevis p2y8 receptor shares high homology with the mammalian P2Y2 and P2Y4 receptors.
Ecto-Enzymes Metabolize Extracellular Nucleotides
Extracellular concentrations of nucleotides are precisely regulated by several groups of membrane-associated ectoenzymes (Zimmermann, 2000). Most prominent of these are the members of the ecto-nucleotidase 5′-triphosphate diphosphohydrolase (NTPDase) family. This family includes NTPDase1 (ecto-ATPDase, CD39, apyrase), which hydrolyzes both nucleoside triphosphates and nucleoside diphosphates; NTPDase2 (ecto-ATPase, CD39L1), which selectively hydrolyzes nucleoside triphosphates; and NTPDase3 (ectoATPDase, CD39L3), which hydrolyzes both nucleoside triphosphates and nucleoside diphosphates with a 3:1 preference for the triphosphates. These three isozymes are integral proteins of the plasma membrane containing short cytosolic N- and C-termini and large extracellular domains containing apyrase conserved regions. At least one additional member of this family, NTPDase5 (CD39L4), is apparently a secreted protein that may account at least in part for soluble nucleotidase activities found under various conditions (Mulero et al., 1999).
The ecto-phosphodiesterase/nucleotide pyrophosphatase (E-NPP) family comprises three different gene-encoded polypeptides with predicted short cytosolic N-termini, one transmembrane domain, and a long extracellular domain bearing the catalytic site (Zimmermann, 2000). This group of isoenzymes consists of E-NPP1 (PC-1), E-NPP2 (autotaxin, PD-Iα), and E-NPP3 (PD-1β, B10). E-NPP isoenzymes display broad substrate specificity hydrolyzing nucleoside 5′-triphosphate (NTP) to nucleoside monophosphate (NMP) plus pyrophosphate, nucleoside 5′-diphosphate (NDP) to NMP plus orthophosphate, dinucleoside polyphosphates to NPn-1 plus NMP, and cAMP to AMP. E-NPP isoenzymes also hydrolyze nucleic acids, NAD, and sugar nucleotides.
Ecto 5′-nucleotidase and alkaline phosphatases are glycosyl phosphatidyl inositol-linked enzymes that convert NMP to nucleoside plus orthophosphate. These enzymes are major determinants of the accumulation of extracellular adenosine from AMP generated during extracellular ATP (and ADP) hydrolysis. Alkaline prosphatase also hydrolyzes ATP and AMP. Extracellular nucleotide interconverting activities such as nucleoside mono- and di-kinases also have been described. The activity of extracellular nucleoside diphosphokinase (NDPK), which catalyzes transphosphorylation reactions such as the reversible phosphorylation of UDP or GDP by ATP (e.g., ATP + UDP ⇋ ADP + UTP), is high, surmounting that of nucleotidases under a number of conditions. For example, in 1321N1 astrocytoma and C6 glioma cells, NDPK rapidly generates ATP from endogenous ADP in the presence of an excess mass of UTP in the medium (Lazarowski et al., 1997b, 2000; Grobben et al., 1999). Adenylyl kinase (2ADP ⇌ATP + AMP) also exists as a potentially important transphosphorylating extracellular activity (Yegutkin et al., 2001, 2002; Donaldson et al., 2002; Picher and Boucher, 2003).
Assessing Extracellular Nucleotides
The mechanism(s) and pathway(s) involved in the release of ATP and other nucleotides from the broad range of cell types known to be involved in physiologically important nucleotide-promoted signaling responses have proven difficult to define. Quantification of ATP release is complicated by a number of technical problems. For example, mechanical perturbations, including those occurring during tissue isolation, cell centrifugation, and routine cell culture procedures, result in uncontrolled release of large amounts of ATP from most cell types. Although mechanical release of ATP is a physiologically regulated process that clearly does not involve cell lysis, damaged cells provide an additional confounding factor in the interpretation of measurement of extracellular ATP. Pharmacological approaches to activate the process(es) involved in mechanically stimulated nucleotide release have not yet been identified. Therefore, no simple means have been elucidated to circumvent the potential problem of cell lysis attendant in studies of mechanically stimulated nucleotide release. Additional difficulty derives from the fact that nucleotide concentrations measured in the bulk phase of extracellular solutions often do not coincide with the local nucleotide concentration predicted near the cell surface by means of specific bio-sensors (e.g., cell-attached luciferase or P2Y receptors; see below). Local hydrolysis further complicates the task of accurately quantifying nucleotide concentrations at the cell surface upon release. Transphosphorylating reactions (NDPK- and adenyl kinase-mediated) contribute additional confusion in assessment of the primary source of intracellular nucleotides. Finally, the recent realization that uridine nucleotides and nucleotide sugars are released from cells either in parallel to or independent from adenine nucleotides (Lazarowski and Harden, 1999; Lazarowski et al., 2003) adds another level of complexity.
Several methods have been used to quantify extracellular ATP. The firefly luciferase assay is applied widely as a very sensitive means for quantitation of ATP. HPLC analysis of extracellular solutions provides an alternative method that also allows assessment of the integrated pattern of purines arising from ATP release and hydrolysis. HPLC analysis of radioactive species released from radiolabel cells is a semi-quantitative approach because the specific activities of the radiolabeled species are unknown, but the mass of ATP and adenyl purines can be quantified at subnanomolar levels after conversion to fluorescent N6-etheno-adenyl purine derivatives (Levitt et al., 1984; Yang et al., 1994; Huang et al., 2001). Quantitation in bulk solutions from various tissues and conditions has revealed the presence of nanomolar concentrations of extracellular nucleotides (Table 2).
Extracellular ATP levels on various tissues and cell lines
An incomplete list of published levels of extracellular ATP is presented as an illustrative example. In most cases, the data represent range of values obtained under various experimental (e.g., basal and stimulated) conditions. The concentration values in the bulk phase were not normalized for the volume of the extracellular solution, tissue surface area, or cell number.
Several approaches have been applied to infer the nucleotide concentrations at the cell surface while minimally disrupting the physiological status of the extracellular environment. The classic firefly luciferase assay was recently adapted to allow quantitation of ATP release in real-time in the presence of excess soluble luciferase and luciferin (Taylor et al., 1998; Beigi et al., 1999; Beigi and Dubyak, 2000). The premise of this approach is that luciferase molecules reaching the cell surface will favorably compete with endogenous ecto-ATPases for detection of ATP. An elegant variation of this method allows assessment of local ATP concentrations at the cell surface by anchoring the luciferase molecule to cells using a chimeric protein made of the IgG-binding domain of Staphylococcus aureus protein A fused with the luciferase enzyme (Beigi et al., 1999; Joseph et al., 2003). A less quantitative approach for assessing nucleotide levels at the cell surface takes advantages of measurement of changes in second messenger production (e.g., ΔCa2+) in cells expressing nucleotide receptors used as biosensor (Homolya et al., 2000; Sauer et al., 2000; Koyama et al., 2001; Coco et al., 2003). Atomic force microscopy in combination with myosin-functionalized sensor tips was introduced as a sensitive technique for quantitation of ATP levels with high-resolution topography in the surface microenvironment of human bronchial epithelial S9 CFTR+ cells (Schneider et al., 1999). Changes in luciferin fluorescence detected by confocal laser scanning microscopy were used recently to monitor the concentration of ATP on the surface of pancreatic acini (Sorensen and Novak, 2001). In contrast to the nanomolar concentrations reported for bulk solution measurements, these procedures yielded “local” ATP concentration values in the 1 to 25 μM range.
Release of ATP by Excitatory/Secretory Tissues
Exocytotic release of ATP by specialized tissues resembles and sometimes overlaps neurotransmitter release. Neurons, chromaffin cells, platelets, and secretory cells, including mast cells and pancreatic acinar cells, control the release of ATP, neurotransmitters, and other extracellular mediators after packaging in specialized granules called synaptic vesicles, chromaffin granules, and dense core granules (Evans et al., 1992; Burnstock, 1997; Sorensen and Novak, 2001). For example, catecholamines, serotonin, and ATP are transported and costored in chromaffin granules using the electrochemical force created by the V-type H+-ATPase of the granule membrane; intravesicular ATP concentrations may reach 150 mM [see Bankston and Guidotti (1996) and references therein]. Stimulation of chromaffin cells by preganglionic sympathetic neurons results in granule transport along the filaments of the cytoskeleton network to the subplasma membrane compartment, fusion of the granule with the plasma membrane, and release of contents into the extracellular space, a process commonly referred to as regulated exocytosis (Dean et al., 1984; Gualix et al., 1999).
The molecular mechanisms that underlie the process of regulated exocytosis have been extensively investigated. Considerable evidence links regulated exocytosis to the SNARE [soluble N-ethyl maleimide-sensitive factor attachment protein (SNAP) receptor] hypothesis (Rothman, 1994). Accordingly, members of the large family of v-SNARE proteins located on vesicles pair with corresponding members of the t-SNARE family of proteins located on the target membrane to determine the specificity of vesicle targeting, docking, and fusion. Major components of the SNARE machinery include the t-SNAREs syntaxin and SNAP-25 and the v-SNARE synaptobrevin, also known as VAMP. The synaptic fusion complex is initiated by formation of a ternary (synaptobrevin, syntaxin, SNAP-25) complex, but additional proteins, including Rabs and Muncs, function at steps up-stream of the SNARE complex formation. During the late step of neurotransmitter release, syntaxin/SNAP 25 binds to the vesicle protein synaptotagmin, which probably acts as a Ca2+ sensor, triggering fusion of the presynaptic vesicle with the plasma membrane (Chapman et al., 1994; Li et al., 1996; Rettig and Neher, 2002; Zhang et al., 2002; Rickman and Davletov, 2003).
ATP Release by Nonexcitatory Cells
Extracellular ATP has been detected in most nonexcitatory tissues, including epithelial and endothelial cells, smooth muscles and fibroblasts, astrocytes, circulating lymphocytes, monocytes and red cells, hepatocytes, and chondrocytes, and in innumerable transformed cell lines. ATP-containing specialized granules have not been unambiguously identified in most of these tissues, and it was generally accepted until recently that vesicular secretion of ATP does not occur outside of excitable specialized tissues.
Nonlytic release of ATP and other nucleotides by nonexcitatory cells has been reported to occur in three different contexts: 1) enhanced release of nucleotides occurs after mechanical stimulation of endothelial, epithelial, glial, and other cells; 2) although less documented, pharmacological stimuli trigger ATP release; and 3) significant release of nucleotides occurs in the absence of an external stimulus (i.e., by resting cells). The properties of these three patterns of nucleotide release are discussed in the following sections.
Mechanical Stimulation. Mechanical release of ATP was first documented during sustained exercise of human forearm muscle (Forrester, 1972) almost simultaneously with Burnstock's proposal of the existence of purinergic receptors (Burnstock, 1972; Forrester, 1972). However, the widespread occurrence and significance of mechanical release of ATP was not realized until more recently. Pioneering work by Milner, Bodin, and coworkers (Forrester, 1972; Milner et al., 1990a,b; Bodin et al., 1991; Milner et al., 1992) illustrated that perfused endothelial cells nonlytically released ATP in response to increased flow rate. The potential autocrine/paracrine role of mechanically released ATP was suggested by studies showing that 1321N1 human astrocytoma cells over-expressing the P2Y1 receptor or the P2Y2 receptor (Filtz et al., 1994; Parr et al., 1994), but not empty vector-expressing cells, displayed increased inositol phosphate accumulation in response to mechanical forces. Moreover, a medium change resulted in elevation of inositol phosphate levels that was prevented by including apyrase in the extracellular solution. ATP concentrations in the medium of mechanically stimulated cells reached values (>100 nM) that were 50 to 100 times higher than resting ATP levels and that were comparable with EC50 values for P2Y2 receptor activation; increases in lactate dehydrogenase activity were not observed in the medium (Parr et al., 1994; Lazarowski et al., 1995). A number of studies followed illustrating that nonlytic release of pharmacologically relevant amounts of ATP were observed in a variety of cell types, including umbilical vein (Yegutkin et al., 2000) and other endothelial cells (reviewed in Burnstock, 1999), airway epithelial cells (Grygorczyk and Hanrahan, 1997; Watt et al., 1998; Homolya et al., 2000; Guyot and Hanrahan, 2002); fibroblasts (Grierson and Meldolesi, 1995), Madin-Darby canine kidney (MDCK), COS-7, and HEK-293 cells (Ostrom et al., 2000), astrocytes [Coco et al. (2003) and references therein], and chondrocytes (Graff et al., 2000). Mechanical release of ATP and activation of P2Y receptors seems to be important for the regulation of cell volume recovery in swollen hepatocytes and related cell lines (Wang et al., 1996; Feranchak et al., 2000). Mechanical stimuli employed to effect these responses included hypotonic cell swelling, cell surface pocking, magnetic field-oriented twisting, hydrostatic pressure, mechanical loading, stretching, and medium disturbance.
Epithelial cells are particularly sensitive to mechanical forces. Perhaps the most compelling evidence for a physiological role of mechanically induced ATP release follows from studies illustrating enhanced ATP release from the urothelium as a consequence of distention of the murine urinary bladder. ATP mediated-activation of P2X3 receptors on subepithelial sensory nerve terminals subsequently controls bladder volume reflexes of the autonomic nervous system (Ferguson et al., 1997; Cockayne et al., 2000; Vlaskovska et al., 2001). Based on these and other studies, it has been proposed that purinergic mechanosensory transduction effects neural reflexes in distended tubes and sacs (including ureter, vagina, salivary and bile ducts, gut, urinary and gall bladders, and lung) (Burnstock, 1999).
Mechanically stimulated release of ATP has been amply documented in airway epithelia (Grygorczyk and Hanrahan, 1997; Watt et al., 1998; Homolya et al., 2000; Guyot and Hanrahan, 2002). Airways continuously remove noxious materials through a mucociliary clearance process that requires epithelial liquid secretion. The cAMP-regulated cystic fibrosis transmembrane conductance regulator (CFTR) is a major Cl- channel promoting liquid secretion onto the lumen, and defective CFTR activity causes cystic fibrosis (CF), the most common genetic disease in the white population. An alternative Ca2+-regulated Cl- channel (CACC), which remains functionally active in CF cells, is also present on the mucosal surface of airway epithelial cells (Wine, 1999). Both cAMP-elevating adenosine A2b receptors and Ca2+ mobilizing P2Y2 receptors are highly expressed along the respiratory tree. The fact that the airway epithelium 1) is a poorly innervated tissue that receives little endocrine regulation and 2) is exposed to complex physical forces (e.g., surface tension, tidal breathing, and coughing) suggests that released nucleotides transduce intraluminal stress into enhanced mucociliary clearance via P2Y2 receptor-mediated CACC activation. Indeed, recent studies with well differentiated bronchial epithelial cells illustrated that luminal flow-induced shear caused a transient, apyrase sensitive, elevation of intracellular Ca2+ and enhanced Cl- secretion (Paradiso et al., 2001).
Agonist-Promoted ATP Release. Person and Gordon (1979) demonstrated that incubation of aortic endothelial and smooth muscle cells with thrombin resulted in the specific release of ATP, which was converted to ADP by vascular hydrolases. More recently, Yang et al. (1994) illustrated that endothelial cells isolated from guinea pig heart release nucleotides in response to bradykinin, acetylcholine, and serotonin. HPLC analysis of ethenylated derivatives indicated that ATP was the predominant purine released in response to hormone stimulation and that ATP dephosphorylated rapidly upon release. The authors also reported that ADP induced large increases of extracellular ATP in endothelial cell cultures, suggesting that an ADP-activated P2Y1 receptor was involved in the mechanism of ATP release. Buxton et al. (2001) reported the involvement of bradykinin receptors in the release of ATP in coronary endothelium and also illustrated that although application of ADP resulted in an increase in ATP levels, this occurred as a consequence of phosphorylation of ADP by NDPK rather than stimulation of P2Y1 receptors. Bradykinin and phenylephrine promoted ATP release in MDCK, COS-7, and HEK-293 cells (Ostrom et al., 2000). UTP also enhanced extracellular ATP accumulation in COS-7 and HEK-293 cells, but the effect of UTP probably reflected competitive inhibition of ATP hydrolysis and phosphorylation of endogenous ADP by NDPK rather than P2Y2 receptor-stimulated ATP release (Ostrom et al., 2000). In a study designed to investigate the potential role of connexin hemichannels in ATP release, Cotrina et al. (1998) reported that stimulation of purinergic receptors with 100 μM UTP in HeLa cells, C6 glioma cells, and U373 glioblastoma cells resulted in a Ca2+-dependent increase of extracellular ATP concentrations. However, other Ca2+-mobilizing agents such as bradykinin, endothelin, and the calcium ionophore A2317 had little or no effect on extracellular ATP levels. Because the likely contribution of NDPK in the phosphorylation of endogenous ADP was not examined, the significance of the ATP measurements in response to exogenous UTP remains unclear.
Release of ATP by Resting Cells. ATP has been detected on the surface of many resting nonsecretory tissues, and basal extracellular ATP levels probably reflect a dynamic steady state. In most tissues or cells, elevated extracellular ATP levels that follow from mechanically induced release (e.g., during tissue washes, addition of reagents, or simply transport of the tissue dish to the bench top) decay with apparent first-order kinetics as a consequence of the hydrolytic action of ecto-ATPases. ATP levels eventually reach and persist in the extracellular medium at low nanomolar (1-10 nM) concentrations for long incubation periods. Moreover, ATP levels on various types of resting cells, including primary cultures of polarized airway epithelial cells, epithelial cell lines, 1321N1 human astrocytoma cells, and C6 rat glioma cells reflect a steady state in which ATP metabolism is balanced by constitutive release. By measuring the extracellular ATP concentrations using the luciferase assay, and simultaneously monitoring the rate of hydrolysis of [γ-32P]ATP (added to the cells in high specific activity as a radiotracer), ATP levels were shown to persist in the low nanomolar range in the presence of active hydrolysis of [γ-32P]ATP (Donaldson et al., 2000; Lazarowski et al., 2000, 2001). Thus, resting cells release ATP at a rate that equals the rate of ATP hydrolysis at steady state. The calculated rates of constitutive ATP release ranged from ∼10 fmol/min/106 cells in C6 rat glioma cells to ∼500 fmol/min/106 cells in well differentiated cultures of primary bronchial epithelial cells. Basal ATP release was observed with various cells types by independent approaches. For example, Beigi and Dubyak (2000) showed that ATP levels on resting BAC-1.2F5 macrophages, determined in real time by the luciferase reaction, increased steadily by a factor of 3 over 10 min when the ecto-ATPase inhibitor α,β-metATP was added to the extracellular solution. Based on a similar concept, Schwiebert et al. (2002) reported that ATP concentrations on resting primary cultures of human vascular endothelial cells increased gradually when the cells were incubated at low temperature to reduce ecto-ATPase activity.
Ostrom et al. (2000) highlighted the physiological significance of basal ATP release by illustrating that release of ATP by resting MDCK cells establishes the set-point for autocrine activation of P2Y receptor-promoted signaling responses. Basal release of ATP has physiological implications for airway homeostasis. Studies with polarized resting epithelial cell lines have illustrated that released ATP confers tonic activation to the luminal P2Y2 receptor (Lazarowski et al., 2001) and, indirectly, to the A2b receptor (Huang et al., 2001), thus providing mechanisms to account for the basal activity of the calcium- (CACC) and cAMP- (CFTR) sensitive chloride channels.
Uridine Nucleotides and Nucleotide Sugars
Realization that several of the P2Y receptor subtypes recognize nucleotides other than adenine nucleotides has motivated initiatives to develop methods for quantifying physiologically relevant levels of these nucleotides. Interest in assaying extracellular levels of nonadenine nucleotides further derives from the realization that both uridine nucleotides and nucleotide sugars also autocrinely activate P2Y receptors. Functional assays with intact cells expressing recombinant P2Y4, P2Y6, or P2Y14 receptors that are selectively activated by UTP, UDP, and UDP-glucose, respectively, indicated that these nucleotide species are released from resting or mechanically stimulated cells in concentrations capable of stimulating their respective receptors (Lazarowski et al., 1997a; Homolya et al., 2000; Lazarowski et al., 2003).
UTP Measurements. Given the high substrate selectivity of the enzyme UDP-glucose pyrophosphorylase (UTP-glucose-1-phosphate uridylyltransferase, EC-2.7.7.9) for the reaction UTP + glucose-1P ⇌UDP-glucose + PPi, a sensitive assay for UTP was developed based on the use of [14C]glucose-1P as a substrate for the UTP-dependent conversion of [14C]glucose-1P to UDP-[14C]glucose (Lazarowski et al., 1997a, 1999). UTP levels were measured in a number of tissues, including primary astrocytes, primary airway epithelial cells, isolated human platelets, and several cell lines. UTP was detected in low nanomolar concentrations in the extracellular medium of most of the cells examined under resting conditions, and enhanced UTP levels were observed in all cases after mechanical stimulation or, as was the case with platelets, after the addition of thrombin. In all nonexcitable tissues, a ratio of 1:3-5 for extracellular UTP/ATP was observed both in resting and mechanically stimulated conditions, closely reflecting the relative intracellular abundance of these nucleotides. Thus, a common source and perhaps a common mechanism of release may exist for both ATP and UTP.
Quantitation of UDP-Glucose. A sensitive assay for UDP-glucose mass also was recently elucidated, applying the same principle of the above reaction with UDP-glucose pyrophosphorylase. This assay uses [32P]PPi as the substrate for the backward direction, and the UDP-glucose-dependent conversion of [32P]PPi to [32P]UTP is followed by HPLC (Lazarowski et al., 2003). Extracellular UDP-glucose was detected with primary and Calu-3 epithelial cells and with 1321N1 human astrocytoma, COS-7, CHO-K1, and C6 glioma cells. Unlike ATP and UTP, mechanical stimulation of cells resulted in little or no UDP-glucose release, but steady accumulation up to 20 nM UDP-glucose occurred over long incubations (1-3 h), suggesting that UDP-glucose is released constitutively, but not mechanically, and is metabolized slowly.
Potential Mechanisms for Nucleotide Release
Several mechanisms are probably involved in the release of nucleotides, given the diversity of conditions and cell types in which extracellular nucleotides have been detected. ATP binding cassette (ABC) transporters, connexin hemichannels, mitochondrial porins (VDAC), and stretch-activated channels all have been postulated as pathways, regulators, or sensors of nucleotide release, whereas other studies have suggested that nucleotides are released during cargo-vesicle trafficking and/or via exocytotic granule secretion. None of these mechanistic possibilities have been independently and unambiguously confirmed. Evidence favoring these and other proposed mechanisms for nucleotide release and their mechanistic implications are considered below.
ABC Transporters. ABC transporters perhaps have attracted the most attention among the various candidate mechanisms and pathways for ATP release. In particular, CFTR and the product of the MDR-1 gene, P-glycoprotein, have been implicated in the release of cytosolic ATP by epithelial and other cells. This hypothesis originated from studies suggesting a correlation between extracellular ATP concentrations and the level of expression of ABC transporters and by electrophysiological studies with whole-cell and single-channel activities showing that P-glycoprotein- and CFTR-containing patches displayed ATP conductance (Abraham et al., 1993; Reisin et al., 1994). The idea that ATP is channeled out of cells through CFTR gained additional support from comparative measurements of release of radioactivity from normal, CF, and CFTR-transfected airway epithelial cells prepermeabilized and loaded with [γ-32P]ATP (Schwiebert et al., 1995). However, subsequent electrophysiological studies showed that neither natively or heterologously expressed CFTR nor highly purified CFTR conducted ATP (Grygorczyk et al., 1996; Reddy et al., 1996; Braunstein et al., 2001). An alternative hypothesis proposes that CFTR and MDR-1 indirectly enhance the activity of a separate channel that itself conducts ATP. Basal and swelling-activated ATP release measured by the luciferin-luciferase assay was decreased in CF relative to non-CF epithelial cell lines and was also augmented in murine cell lines transfected with wild-type CFTR (Hazama et al., 2000; Braunstein et al., 2001; Roman et al., 2001). Elevation of cAMP levels in CFTR cRNA-injected Xenopus laevis oocytes resulted in increased ATP release that was also sensitive to changes in extracellular Cl- ion concentrations (Jiang et al., 1998). Moreover, cAMP-promoted CFTR Cl- channel activity was accompanied by enhanced extracellular ATP accumulation in swell-stimulated airway epithelial cells (Braunstein et al., 2001). In marked contrast, swelling-induced, CFTR-independent ATP release was reported to occur in intestinal epithelial cells (Hazama et al., 1999). Similarly, swelling-activated ATP release was augmented in murine C127 cells over-expressing CFTR but was not affected by cAMP levels, indicating that ATP release was independent of CFTR activation (Hazama et al., 2000). The role of CFTR in ATP release was further brought into question by studies with primary epithelial cell cultures and cell lines, and by in vivo studies of nasal secretions from normal subjects and CF patients. That is, similar levels of extracellular ATP and similar rates of ATP release were observed in normal and CF cells, as well as in null cells and cells expressing different levels of functionally active CFTR (Grygorczyk and Hanrahan, 1997; Watt et al., 1998; Hazama et al., 1999; Donaldson et al., 2000).
Fitz and coworkers (Roman et al., 1997, 2001) provided indirect evidence for involvement of MDR proteins in the control of extracellular ATP levels and have suggested that the P-glycoprotein regulates the activity of a structurally unrelated ATP channel. This hypothesis follows from studies of swell-activated HTC hepatoma cells illustrating that 1) MDR proteins confer enhanced cell volume recovery that was impaired in the presence of ATP scavengers, 2) extracellular ATP accumulation decreased in the presence of MDR-1 protein inhibitors, and 3) MDR-1-expressing HTC cells displayed a greater swell-activated ATP conductance relative to control cells. The idea that P-glycoprotein is involved in the release of nucleotides may not apply to other cell types. Recent studies with ureter epithelium indicated that stretch-induced ATP release was not affected by verapamil, a known inhibitor of MDR-1 protein function and other related ABC transporters (Knight et al., 2002). Swell-promoted ATP release in ocular ciliary epithelial cells also was unaffected by inhibitors of MDR-1 protein (Mitchell et al., 1998).
Stretch- and Voltage-Activated Channels. Although the role, if any, of CFTR and other structurally related ABC transporters in the release of ATP remains unclear, other mechanisms are almost certainly involved, as suggested by recent studies with epithelial, endothelial, and astroglial cells. In epithelial cells, a swell-induced ATP conductance clearly separate from any activity of CFTR was resolved in a vesicle preparation and shown to be inhibited by 50 μMGd3+ (Braunstein et al., 2001), a nonselective blocker of stretch-activated cation channels (SAC) (Sackin, 1995; Caldwell et al., 1998). These results suggest a close association between SACs and ATP conducting channels. However, nonspecific inhibition of luciferase and Ca2+ channels by >10 μM Gd3+ (Boudreault and Grygorczyk, 2002) casts doubt on the reliability of ATP measurements based on the luciferin-luciferase assay or on ATP-promoted Ca2+ responses in intact cells. Indeed, Boudreault and Grygorczyk (2002) found no evidence for the involvement of Gd3+-sensitive SAC channels in the release of ATP by 16HBE14o- bronchial epithelial cells, human lung carcinoma cells, and NIH 3T3 fibroblasts, using controlled incubation conditions to selectively inhibit SAC by Gd3+ without affecting the ATP assay. In ureter epithelium, 10 μM Gd3+ failed to inhibit ATP release in response to distention (Knight et al., 2002).
Observation that a volume-dependent anion channel conducted ATP in C127i mouse mammary cells (Sabirov et al., 2001; Hisadome et al., 2002) led to the suggestion that the plasmalemmal voltage-dependent anion channel (pl-VDAC), a murine splicing variant of the mitochondrial ATP porin VDAC-1 (Rostovtseva and Colombini, 1996; Buettner et al., 2000), may act in an ATP-releasing pathway. Although pl-VDAC transcripts were found in murine tracheal epithelial cells (Okada et al., 2002), no data are available demonstrating that pl-VDAC participates in ATP transport across the plasma membrane.
Ca2+ wave propagation was examined as an approach to assess ATP release by mechanically stimulated DU-145 human prostate cancer cells. Ca2+ waves were inhibited by the nonselective channel blockers 4,4′-diisothiocyanato-stilbene-2,2′-disulfonic acid, niflumic acid, tamoxifen, and Gd3+, but not by the P-glycoprotein-associated anion channel blockers verapamil and quinidine, the CFTR antagonist glibenclamide, or the gap junction blocker 1-heptanol (Sauer et al., 2000). The molecular targets of the effective Ca2+ wave inhibitors remain unknown.
Connexin Hemichannels. Cotrina et al. (1998) reported that UTP promoted Ca2+-dependent ATP release in C6 rat glioma cell lines selected for the expression of recombinant connexin 43 or connexin 32. Conditions known to promote the elevation of intracellular Ca2+ in these cells (e.g., Ca2+ionophores, bradykinin, endothelin) had no effect on ATP release. Although an endogenous P2Y2 receptor was implicated, potential contribution of NDPK-catalyzed phosphorylation of endogenous ADP by UTP was not examined. In a recent study, connexin 43-expressing C6 cells were reported to display increased levels of extracellular ATP in response to mechanical stimulation. This effect was potentiated by low extracellular calcium ([Ca2+]e), a condition reported to favor the opening of connexin hemichannels, and was impaired by maneuvers that inhibit connexin 43 permeability [e.g., Gd3+ and flufenamic acid (Stout et al., 2002)]. Similarly, extracellular ATP levels in resting hippocampal astrocytes and glial cells were enhanced by low [Ca2+]e and were partially reduced (30%) by the gap-junction inhibitor anandamide (Coco et al., 2003). However, ATP release by mechanically stimulated astrocytes was not affected by anandamide or by flufenamic acid, suggesting that connexin hemichannels may be involved in ATP release under basal conditions in a low-[Ca2+]e environment, but other mechanisms mediate ATP release by stimulated astrocytes (Coco et al., 2003). No evidence for the involvement of connexin hemichannels in ATP release was observed in human airway epithelial cells (Guyot and Hanrahan, 2002) or murine mammary cells (Sabirov et al., 2001). Observation that calcium wave propagation in spinal cord astrocytes from connexin 43 (-/-) mice are primarily mediated by extracellular diffusion of ATP further argue against a role of connexin hemichannels in ATP release (Scemeset al., 2000).
In summary, the ATP conductances described in cultured hepatoma cells and in bovine tracheal epithelia remain unidentified, and prevalent evidence does not support the idea that SACs or connexins fulfill a role in ATP release. Whether pl-VDAC is a cell surface ATP channel also waits for conclusive results.
Second Messengers and Signaling Cascades Involved in Nucleotide Release
Few studies have examined the signaling events that exist upstream of ATP release mechanisms. Small but significantly enhanced ATP accumulation was reported in forskolin-treated 3T3-fibroblasts and COS-7 cells transfected with CFTR (Braunstein et al., 2001). However, no effect on ATP release was observed by elevating cAMP levels in mammary carcinoma cells (Hazama et al., 1999), primary human nasal epithelial cells (Watt at al., 1998), and several epithelial cell lines (Grygorczyk and Hanrahan, 1997) regardless of the level of CFTR expression. In MDCK, COS-7, and HEK cells, bradykinin and phenylephrine, but not forskolin or PGE1, promoted ATP release, suggesting involvement of effectors downstream of Gq and phospholipase C, but not of Gs and adenylyl cyclase (Ostrom et al., 2000). Calcium-activated ATP release was reported in primary human vascular endothelial cells (Schwiebert et al., 2002) and in ocular ciliary epithelial cells, in which ionomycin-triggered ATP release was sensitive to the calmodulin kinase inhibitor KN-62 (Mitchell et al., 1998). In light of these observations, it is conceivable that the enhanced ATP accumulation reported previously with bradykinin-, acetylcholine- and serotonin-stimulated endothelium (Yang et al., 1994; Buxton et al., 2001) also reflected Ca2+-promoted ATP release.
Joseph et al. (2003) recently absorbed chimeric protein A-luciferase to antibodies against glycosyl phosphatidyl inositol-linked plasma membrane proteins to assess ATP levels at the surface of 1321N1 human astrocytoma cells. Activation of endogenous receptors by thrombin or carbachol promoted ATP release through a mechanism that in the case of muscarinic receptor activation was largely accounted for by elevation of cytosolic Ca2+ but in the case of thrombin receptor activation was only partially Ca2+ dependent. In contrast, the basal release of ATP from 1321N1 cells was not dependent on elevation of Ca2+ levels.
Other lines of evidence suggest that ATP release is controlled by multiple kinases. PI 3-kinase is an important modulator of bile flow in cholangiocytes. Studies with hepatocytes and cholangiocytes also have implicated PI 3-kinase in the regulation of ATP release (Feranchak et al., 1998, 1999), and the PI 3-kinase inhibitors wortmannin and LY294002 decreased basal and swell-stimulated ATP release in hepatoma and Mz-Cha-1 human cholangiocarcinoma cells. Tyrosine kinase inhibitors (herbimycin A and tyrphostin 46) reduced the release of ATP by swell-stimulated aortic endothelium. Most notably, marked decreases in ATP release by swollen aortic endothelial cells were observed by inhibiting Rho or Rho kinase with the clostridium botulinum C3 exoenzyme or Y-26732, respectively; conversely, enhanced ATP release resulted in the presence of lysophosphatidic acid, which activates Rho (Koyama et al., 2001). Based on the current notion that the thrombin receptor PAR-1 signals in part via Gα12 → guanine nucleotide exchange factor → Rho (Seasholtz et al., 1999), the thrombin-activated ATP release (discussed above) in endothelial cells (Pearson and Gordon, 1979) or in 1321N1 human astrocytoma cells (Joseph et al., 2003) may involve Rho activation. Because both PI 3-kinase and Rho have been implicated in the regulation of vesicle trafficking and cytoskeleton organization, one interpretation of the above results (Feranchak et al., 1998, 1999, 2000) is that ATP release in part involves vesicle transport.
Nucleotide Release via Vesicle Trafficking
Recent studies have suggested the existence of nucleotide release via secretory pathways in astrocytes, epithelial, endothelial, and other cells. Indeed, SNAREs and other elements of the regulated exocytosis machinery are essential components of vesicular traffic not only in excitatory tissues but in all eukaryotic cells. The v-SNARE protein synaptobrevin II and the t-SNARE proteins syntaxin I and SNAP-25 have been recently identified in astrocytes, suggesting that astrocytes are competent for undergoing vesicular exocytosis [reviewed in Volknandt (2002)]. Astrocytes synthesize and release a number of amino acid-related signaling molecules, including glutamate, which is released in large amounts by receptor-promoted Ca2+-dependent exocytotic mechanisms as well as by the synaptic venom α-latrotoxin (Parpura et al., 1995a,b; Bezzi et al., 1998). Release of ATP by astrocytes also has been amply documented, and recent experiments illustrating that elevation of intracellular Ca2+ in astrocytes results in glutamate (Innocenti et al., 2000) and ATP release (Guthrie et al., 1999; Queiroz et al., 1999) suggest that ATP release may be exocytotic. There are recent clues supporting this hypothesis. First, subcellular fractionation of astrocytes yielded a major ATP-containing fraction that copurified with secretogranin II, a vesicle protein that is released by stimulated astrocytes in a Ca2+-dependent manner (Coco et al., 2003). Second, release of ATP by mechanically stimulated astrocytes was sensitive to the v-ATPase inhibitor bafilomacyn A1, which also caused depletion of quinacrine-stainable (probably ATP-containing) granules. However, release of ATP by mechanically stimulated astrocytes was only partially sensitive to tetanus toxin, whereas glutamate release was essentially completely inhibited (Coco et al., 2003), suggesting that astrocytes, as is the case in neurons, express dual secretory pathways involving an ATP/secretogranin II-containing dense-core granules and clear electron lucent synaptic vesicles (containing glutamate). Whether astrocytes constitute another example of specialized secretory cells or provide emerging evidence for regulated secretion as a more general phenomenon pertinent to most tissues is an important question under very active investigation.
The exocytotic pathway is essential for protein sorting and for the integrity of membrane polarization in epithelial and endothelial cells, and recent evidence suggests that vesicle trafficking also is involved in ATP release from these cells. Shear-induced ATP release in distended urothelium was sensitive to brefeldin A, a reagent that blocks traffic between the endoplasmic reticulum (ER) and the cys Golgi (Knight et al., 2002). Brefeldin A was previously shown to impair ATP release in oocytes (Maroto and Hamill, 2001). In vascular endothelium, shear-induced ATP release was sensitive to monensin and N-ethylmaleimide (Bodin and Burnstock, 2001), and low temperatures inhibited ATP release across the apical membrane of human umbilical vein endothelial cell monolayers stimulated by ionomycin or by hypotonic shock (Schwiebert et al., 2002). These results are consistent with exocytotic release of ATP, although they do not rule out the possibility that trafficking of an ATP channel or its upstream regulators accounts for ATP release.
Separate evidence for constitutive release of nucleotides via vesicular trafficking has been provided recently by illustrating that a number of cells release UDP-glucose to the extracellular medium (Lazarowski et al., 2003). UDP-glucose plays a central role in the quality control of glycoproteins and is concentrated up to 20 times in the lumen of the ER and probably in pre-Golgi structures, where it serves as a glucose donor substrate for glucosyl-transferases that glucosyl-tag denatured domains of newly synthesized glycoproteins for subsequent chaperone recognition (Parodi, 2000). Luminal UDP-glucose is converted during this process to UDP, which in turn is hydrolyzed to UMP. A specific UDP-glucose/UMP antiporter translocates cytosolic UDP-glucose to the lumen and UMP to the cytosol. Because UDP-glucose/UMP antiporter activity has been detected primarily in the ER and less prominently in Golgi structures (Hirschberg et al., 1998), it seems unlikely (but not impossible) that this transporter effects efflux of cytosolic UDP-glucose to the extracellular space through the plasma membrane. Vesicular UDP-glucose is more likely to be delivered as a cargo molecule during transport of ecto-glycoproteins and vesicle fusion with the plasma membrane (Fig. 2). By analogy, vesicular trafficking of membrane-targeted phosphoproteins (phosphorylated in luminal domains) may provide a constitutive pathway for the release of ATP. In this regard, it is also noteworthy that ADP, GDP, and UDP are detected at the surface of resting cells in amounts comparable with or greater than their respective nucleoside triphosphates (Lazarowski et al., 2000, 2001). Without underplaying the role of ecto-NTPases in the conversion of extracellular NTPs to ADP, UDP, or GDP, vesicular secretion also may contribute to direct release of these residual products of luminal protein kinases and glycosyltransferases to the extracellular space.
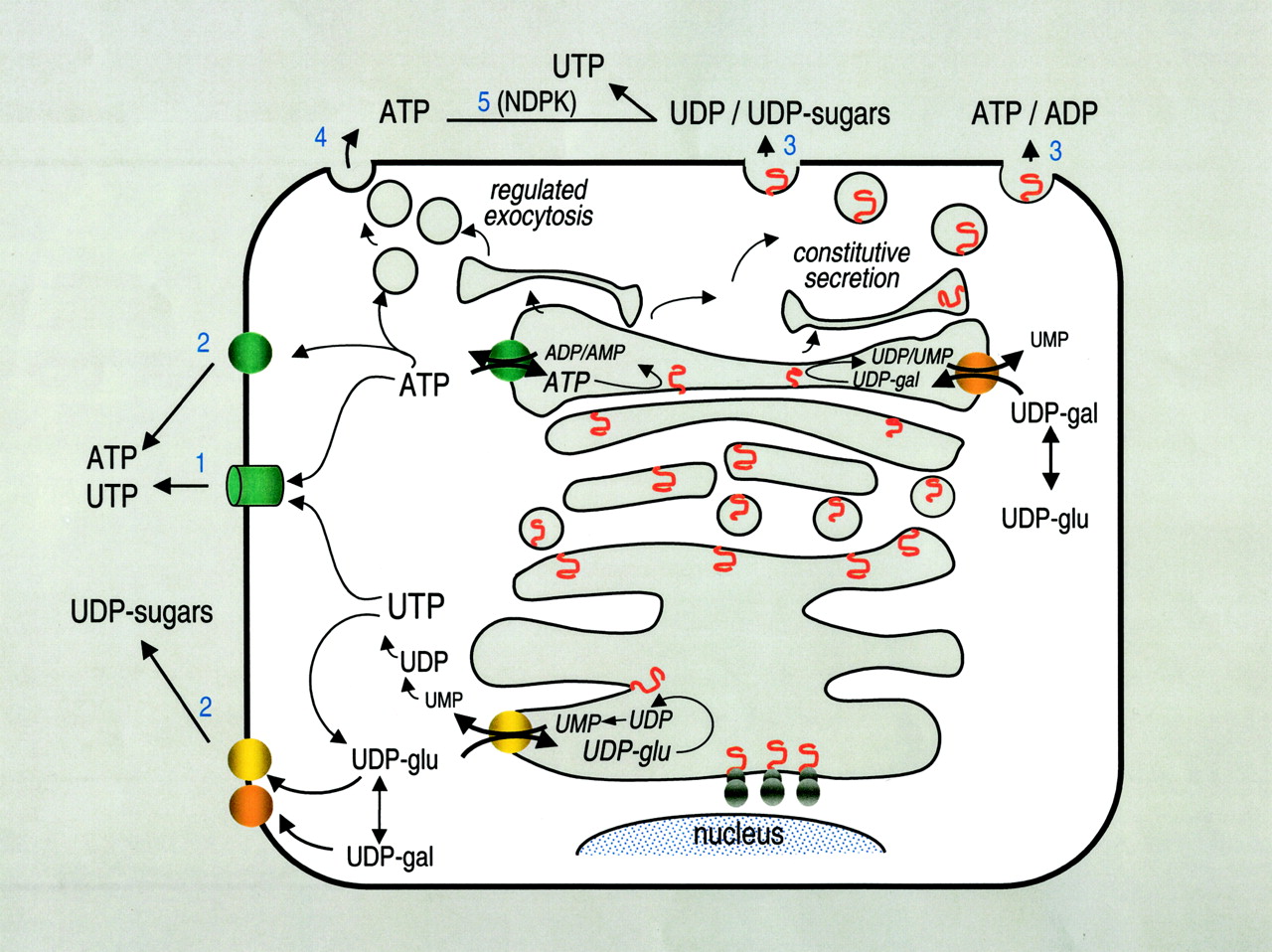
Pathways for nucleotide and nucleotide sugar release. Several scenarios possibly account for the basal and stimulated release of nucleotides by nonexcitatory cells: 1, a not yet identified ATP conductance effluxes cytosolic ATP and perhaps UTP out of the cells; 2, cytosolic ATP and UDP-sugars hypothetically can be exported via specific transporters similar to those existing in ER and Golgi; 3, vesicular luminal ATP, ADP, and AMP, as well as UDP-sugars, UDP, and UMP potentially may be released as residual cargo products of the constitutive secretory pathway; and 4, dense-core-granule-like post Golgi vesicles storing ATP may be competent for regulated exocytosis. 5, note that the constitutive secretory pathway also might provide UDP substrate for NDPK-catalyzed reactions and therefore constitutes an alternative source of extracellular UTP.
Concluding Remarks
The diversity of conditions in which ATP, UTP, and UDP-glucose release is observed suggests that several (perhaps cell-specific) mechanisms are involved. The two most likely possibilities include involvement of a plasma membrane channel or transporter and exocytosis (Fig. 2). Current evidence does not support a direct role of CFTR, MDR-1, connexin 43, or stretch-activated channels as pathways for the efflux of cytosolic ATP under physiological conditions; whether these elements modulate the activity of separate nucleotide-releasing pathways is under debate. Release of UDP-glucose by resting epithelial and other nonexcitatory cells provides an attractive explanation for the basal release of nucleotides via the constitutive secretory pathway. The decreased ATP release observed in mechanically stimulated vascular endothelial cells and urothelium pretreated with vesicle trafficking inhibitors further suggests that discrete subpopulations of nucleotide-containing vesicles may escape the constitutive pathway to be positioned for fusion with the plasma membrane under certain conditions (e.g., after mechanical stress). Whether epithelial, endothelial, and other “nonsecretory” tissues are competent for undergoing regulated exocytosis of nucleotides, whether cytosolic nucleotides make their way to the extracellular space via facilitated diffusion or transport, and whether these two potential releasing pathways coexist and by what mechanism(s) they are activated are under intense investigation. The position of extracellular nucleotides and P2X- and P2Y-receptor mediated signaling at the heart of many physiological and patho-physiological processes make the nucleotide release apparatus(es) a potentially valuable therapeutic target.
Footnotes
-
ABBREVIATIONS: NTPDase, ecto-nucleotidase 5′-triphosphate diphosphohydrolase; E-NPP, ecto-phosphodiesterase/nucleotide pyrophosphatase; NTP, nucleoside 5′-triphosphate; NMP, nucleoside 5′-monophosphate; NDP, nucleoside 5′-diphosphate; NDPK, nucleoside diphosphokinase; HPLC, high-performance liquid chromatography; SNARE, soluble N-ethyl maleimide-sensitive factor; SNAP, soluble N-ethyl maleimide-sensitive factor attachment protein; MDCK, Madin-Darby canine kidney; HEK, human embryonic kidney; CFTR, cystic fibrosis transmembrane conductance regulator; CF, cystic fibrosis; CACC, calcium-regulated chloride channel; ABC, ATP binding cassette; VDAC, voltage-dependent anion channel; MDR, multidrug resistance; SAC, stretch-activated channels; PI, phosphoinositide; LY294002, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one; Y-26732, (R)-(+)-trans-N-(4-pyridyl)-4-(1-aminomethyl)-cyclohexanecarboxamide; ER, endoplasmic reticulum.
- Received April 7, 2003.
- Accepted June 12, 2003.
- The American Society for Pharmacology and Experimental Therapeutics
References
In this issue





{kind=link}
{kind=link}
Jump to section
- Article
- Extracellular Nucleotides Activate Two Types of P2 Receptors
- Ecto-Enzymes Metabolize Extracellular Nucleotides
- Assessing Extracellular Nucleotides
- Release of ATP by Excitatory/Secretory Tissues
- ATP Release by Nonexcitatory Cells
- Uridine Nucleotides and Nucleotide Sugars
- Potential Mechanisms for Nucleotide Release
- Second Messengers and Signaling Cascades Involved in Nucleotide Release
- Nucleotide Release via Vesicle Trafficking
- Concluding Remarks
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters