


Abstract
Adenosine A2B receptors are known as low-affinity receptors due to their modest-to-negligible affinity for adenosine and prototypic agonists. Despite numerous synthetic efforts, 5′-N-ethylcarboxamidoadenosine (NECA) still is the reference agonist, albeit nonselective for this receptor. In our search for higher affinity agonists, we developed decision schemes to select amino acids for mutation to the corresponding residues in the most homologous, higher affinity, human A2A receptor. One scheme exploited knowledge on sequence alignments and modeling data and yielded three residues, V11, L58, and F59, mutation of which did not affect agonist affinity. The second scheme combined knowledge on sequence alignments and mutation data and pointed to Ala12 and Asn273. Mutation of Ala12 to threonine did not affect the affinity for NECA, (R)-N 6-(phenylisopropyl)adenosine (R-PIA), and 2Cl Ado. The affinity of the N273Y mutant for NECA and R-PIA and for the antagonists xanthine amine congener (XAC), ZM241385, and SCH58261 was also unaltered. However, this mutant had a slightly increased affinity for a 2-substituted adenosine derivative, CGS21680. This prompted us to investigate other 2-substituted adenosines, with selectivity and high affinity for A2A receptors. All four compounds tested had improved affinity for the N273Y receptor. Of these, 2-(1-hexynyl)adenosine had submicromolar affinity for the N273Y receptor, 0.18 ± 0.10 μM, with a 61-fold affinity gain over the wt receptor. In addition, the non-NECA analog (S)-PHP adenosine had an affinity of 1.7 ± 0.5 μM for the wt receptor. The high affinity of (S)-PHP adenosine for the wt receptor suggests that further modifications at the 2-position may yield agonists with even higher affinity for A2B receptors.
Extracellular adenosine is best described as a local hormone, which exerts its effects via G protein-coupled adenosine receptors. Four different subtypes have been identified, the A1, A2A, A2B, and A3 receptors. Of these, the adenosine A2B receptors have been implied in atherosclerosis of smooth muscle cells, in mast cell activation resulting in bronchoconstriction in asthma, in intestinal (dys)function, and in dilation of certain vascular beds (for a recent review, see Feoktistov et al., 1998). Activation of adenosine A2B receptors has also been linked to the inhibition of tumor necrosis factor-α production (Le Vraux et al., 1993; Munro et al., 1998), although adenosine A2A(Munro et al., 1998) and A3 receptors (Sajjadi et al., 1996) may also play a role in the production of this cytokine.
The adenosine A2B receptors are known as low-affinity receptors (Bruns et al., 1986) due to their modest-to-negligible affinity for prototypic agonists such as NECA, R-PIA, and CGS21680. Despite the synthesis and biological testing of a wide range of agonists, NECA still is the prototypic agonist for the adenosine A2B receptor (De Zwart et al., 1998,1999a). As a consequence, the current repertoire of agonist ligands for the adenosine A2B receptors is very limited. Moreover, no selective agonists are available for this receptor subtype (Fredholm et al., 1998; Receptor and Ion Channel Nomenclature Supplement, 1999).
In contrast to the agonists, prototypic antagonists such as DPCPX, ZM241385, CGS15943, and SCH58261 share a good-to-reasonable affinity for the adenosine A2B receptor (Klotz et al., 1998; Ongini et al., 1999). The affinity of [3H]DPCPX enabled Robeva et al. (1996) to perform radioligand binding experiments on adenosine A2B receptors. Moreover, CGS15943, DPCPX, and ZM241385 have been used as lead compounds to synthesize antagonists with high affinity (Kim et al., 1998; De Zwart et al., 1999b). From a series of anilide derivatives of 8-phenyl-1,3-dialkylxanthines,Kim et al. (2000) identified several selective A2B receptor antagonists with affinities in the range of 1 to 3 nM. The most selective of these was MRS1754 with an affinity of 1.97 nM and ≥210-fold selectivity over the other human adenosine receptor subtypes.
Because the synthetic approach toward the development of high-affinity agonists has not yet yielded strong improvements in agonist affinity, we have initiated an alternative, molecular biological, approach to investigate the molecular phenomena behind the low affinity of adenosine A2B receptors for agonists. We developed two decision schemes that exploit combinations of current knowledge on mutation data that are available for the other adenosine receptors with ligand binding data, sequence alignments and three-dimensional receptor models. With these schemes we selected amino acids in the A2B receptor that might be involved in agonist binding. The selected residues were mutated into the corresponding amino acids of the most homologous human adenosine A2A receptor to aim for “gain-of-affinity” mutants.
Experimental Procedures
Materials
Adenosine deaminase was obtained from Boehringer Mannheim (Mannheim, Germany); cAMP was from Serva, Boehringer Ingelheim (Heidelberg, Germany). [3H]cAMP with a specific activity of 31.4 Ci/mmol and [3H]DPCPX with a specific activity of 120 Ci/mmol were obtained from NEN Life Science Products, DuPont Nemours (‘s-Hertogenbosch, the Netherlands). CGS15943 was a gift from Dr. J. Watthey and CGS21680 was a gift from Dr. R. A. Lovell, both from Ciba-Geigy (Summit, NJ). 2Cl Ado and R-PIA were from Sigma (St Louis, MO); Cilostamide was obtained from Tocris Cookson (Bristol, UK); Dulbecco’s modified Eagle's medium (DMEM) and F-12 powder media as well as trypsin (2.5% solution) were from Life Technologies (Breda, the Netherlands); and fetal calf serum and newborn calf serum were from Greiner (Alphen a/d Rijn, the Netherlands) and Hyclone (Logan, UT), respectively. MPE Ado (4) and NECA were gifts from Prof. R. A. Olsson of the University of South Florida (Tampa, FL). The synthesis of MPE Ado (4) has been described by Ueeda et al. (1991). Penicillin and streptomycin were from Duchefa (Haarlem, the Netherlands). Rolipram was a gift from Dr. N. Sprzagala, Schering AG (Berlin, Germany); SCH58261 was a gift from Dr. S. Dionisotti, Schering-Plough (Milan, Italy). 2-Thioadenosine was a gift from Dr. Kikukawa from the Tokyo College of Pharmacy (Tokyo, Japan). XAC was from Research Biochemicals International (Natick, MA). ZM241385 was a gift from Dr. S. M. Poucher, Astra-Zeneca Pharmaceuticals (Macclesfield, UK).
General Synthesis Methods and Materials
Chromatography.
Thin-layer chromatography was carried out using aluminum sheets (20 × 20 cm) with silica gel F254 from Merck (Darmstadt, Germany). Spots were visualized under UV (254 nm). Preparative column chromatography was performed on silica gel (230–400 mesh, American Society for Testing and Materials).
Instruments and Analyses.
Elemental analyses were performed for C, H, N (Leiden University, the Netherlands).13C NMR spectra were measured at 50.1 MHz with a JEOL JNM-FX 200 spectrometer equipped with a PG 200 computer operating in the Fourier-transform mode. 1H NMR spectra were measured at 200 MHz, using the above-mentioned spectrometer, or at 300 MHz, using a Bruker WM-300 spectrometer equipped with an ASPECT-2000 computer operating in the Fourier-transform mode. Chemical shifts for 1H and 13C NMR are given in ppm (δ) relative to tetramethylsilane as internal standard.
All high-resolution mass spectra were measured on a Finnigan MAT900 mass spectrometer equipped with a direct insertion probe for electron ionization experiments (70 eV with resolution 1000) or on a Finnigan MAT TSQ-70 spectrometer equipped with an electrospray interface for electrospray ionization experiments. Spectra were collected by constant infusion of the analyte dissolved in 80:20 methanol/H2O. Electrospray ionization is a soft ionization technique resulting in protonated, sodiated species in positive ionization mode and deprotonated species in the negative ionization mode. Melting points (not corrected) were determined in a Büchi capillary melting point apparatus.
2-(1-Hexynyl)adenosine (1).
This compound was synthesized as described by Abiru et al. (1991).
2-((E)-1-Hexenyl)adenosine (2).
This compound was synthesized as described by Vittori et al. (1996).
2-((N′-3-Methyl-1-butylidene)hydrazino)adenosine (3).
This compound was synthesized essentially as described byNiiya et al. (1992) except that 2-iodoadenosine was the starting material for the synthesis of 2-hydrazino-adenosine instead of 2-chloroadenosine.
2-((3R)-3-Hydroxy-3-phenyl-1-propynyl)adenosine (5) (R)-PHP Ado LUF5599.
One hundred and fifty milligrams (0.38 mmol) of 2-iodoadenosine was dissolved in 3 ml of dry acetonitrile and 3 ml of triethylamine. Then 5.4 mg of CuI (28.4 μmol), 3.5 mg of PdCl2 (19.7 μmol), and 11.5 mg of Ph3P (43.8 μmol) were added. Finally, 1.86 mmol (230 μl) of (S)-1-phenyl-2-propyn-1-ol ([α]20 D +28 ± 2°; er [S]:[R] > 98:2) was added, and the brownish solution was stirred overnight at room temperature under a nitrogen atmosphere. The mixture was concentrated in vacuo and purified by column chromatography (5% MeOH in ethyl acetate). The product was crystallized from ethanol. Yield 60 mg (0.15 mmol, 40%), mp 170–172°C; Rf 0.10 (10% MeOH in ethyl acetate); 1H NMR (DMSO-d6 ) δ 8.42 (s, 1H, H-8), 7.54-7.34 (m, 5H, phenyl), 6.26 (d, 1H, J = 5.84 Hz, H-1′), 5.85 (d, 1H, J = 5.84 Hz, OH-2′), 5.59 (d, 1H,J = 5.49 Hz, OH-5′), 5.47 (d, 1H, J = 5.84 Hz, OH-3′), 5.19-5.13 (m, 2H, CHOH, CHOH), 4.49-4.46 (m, 1H, H-2′), 4.12-4.07 (m, 1H, H-3′), 3.93-3.90 (m, 1H, H-4′), 3.65-3.54 (m, 2H, H-5′); MS m/z 398 (M+H)+; Anal. (C19H19N5O5) C, H, N.
2-((3S)-3-Hydroxy-3-phenyl-1-propynyl)adenosine (6) (S)-PHP Ado LUF5600.
One hundred and fifty milligrams (0.38 mmol) of 2-iodoadenosine was dissolved in 3 ml of dry acetonitrile and 3 ml of triethylamine. Then 5.4 mg of CuI (28.4 μmol), 3.5 mg of PdCl2 (19.7 μmol), and 11.5 of mg Ph3P (43.8 μmol) were added. Finally, 1.86 mmol (230 μl) of (R)-1-phenyl-2-propyn-1-ol ([α]20 D −28 ± 2°; er [R]:[S] > 96:4) was added, and the brownish solution was stirred overnight at room temperature under a nitrogen atmosphere. The mixture was concentrated in vacuo and purified by column chromatography (5% MeOH in ethyl acetate). The product was crystallized from ethanol. Yield 80 mg (0.20 mmol, 53%), mp 130–132°C; Rf 0.11 (10% MeOH in ethyl acetate); 1H NMR (DMSO-d6 ) δ 8.42 (s, 1H, H-8), 7.55-7.34 (m, 5H, phenyl), 6.27 (d, 1H, J = 5.84 Hz, H-1′), 5.84 (d, 1H, J = 5.49 Hz, OH-2′), 5.59 (d, 1H,J = 6.18 Hz, OH-5′), 5.46 (d, 1H, J = 6.18 Hz, OH-3′), 5.19-5.13 (m, 2H, CHOH, CHOH), 4.49-4.46 (m, 1H, H-2′), 4.12-4.07 (m, 1H, H-3′), 3.93-3.90 (m, 1H, H-4′), 3.65-3.54 (m, 2H, H-5′); MS m/z 398 (M+H)+; Anal. (C19H19N5O5) C, H, N.
DNA Constructs
The human adenosine A2B receptor clone in pRc/CMV was obtained from Dr. A. Townsend-Nicholson (Garvan Institute, Sydney, Australia). Because in vitro polymerase reactions did not succeed with this construct, we transferred the gene to pUC19 using the restriction sites for HindIII and XbaI. Subsequently, we removed most of the upstream 5′ sequence using the available XmaI restriction sites. The removal of this palindromic sequence allowed the in vitro polymerase reactions to take place. Point mutations were introduced with the pGene-editor kit from Promega (Madison, WI) in such a way that either a restriction enzyme site was introduced or removed. These sites were SspBI (introduced to create the V11I and N273Y mutants), SapI (removed to create the L58V and F59L mutants), and HaeII (introduced to create A12T). The presence of the mutation was verified after transfer of the DNA construct to pcDNA3, with both restriction enzymes and DNA sequencing. DNA was introduced in competent XL10-blue bacteria via electroporation. DNA was isolated with the MaxiPrep kit from Qiagen (Chatsworth, CA).
Cell Culturing
COS-7 cells were obtained from Dr. R. Leurs (Vrije Universiteit, Amsterdam, the Netherlands) and grown in DMEM with 5% newborn calf serum, 2 mM glutamax (a stable glutamine analog from Life Technologies), 50 I.U./ml penicillin, and 50 μg/ml streptomycin at 37°C in a 7% CO2 atmosphere. CHO cells were grown in a 1:1 mixture of DMEM and Ham's F-12 medium, containing 10% newborn calf serum, 2 mM glutamax, 50 I.U./ml penicillin, and 50 μg/ml streptomycin at 37°C in a 5% CO2atmosphere.
Transfection
COS-7 cells and CHO cells were transfected in suspension with the modified DEAE/dextran method essentially as described in Brakenhoff et al. (1994). Briefly, 1 day before transfection the cells were subcultured to obtain 50% confluency at the day of transfection. On the day of transfection 100 μM chloroquin (freshly prepared as a 100× stock solution in PBS) was added to RPMI-1640 with 2% fetal calf serum (RSC). Separate DEAE and DNA solutions were made in RSC as follows. Per 106 cells 2 ml of RSC was added to 180 μl of DEAE (10 mg/ml in PBS). In another tube, 2 μg of DNA was added to 2 ml of RSC. In the meantime the cells were trypsinized, centrifuged, and resuspended in RSC (2 × 106 cells/ml). DEAE was added to the DNA solution and was homogenized by inverting the tube 10 times. After 2 min at room temperature the cells (0.5 ml containing 106cells) were added and the suspension was incubated at 37°C in 7% CO2 atmosphere. The cells were subsequently centrifuged (600g for 10 min), washed with medium, centrifuged again, and resuspended in medium. For [3H]DPCPX binding experiments, the cells were resuspended to obtain 1.5 × 106 cells/6 ml and plated in a 10-ml plate. For cAMP generation experiments the cells were plated in 24-wells plates in a volume of 400 μl at a density of 2 × 105 cells/well. The cells were used 48 h after transfection.
[3H]DPCPX Binding Experiments
COS-7 cells were scraped from the plates and counted. Radioligand binding experiments were performed essentially as described in De Zwart et al. (1999b). On average, 3 × 105 cells were incubated in a total volume of 100 μl of 140 mM NaCl, 5 mM KCl, 5 mM glucose, 20 mM Tris, pH 7.4, together with 6 nM [3H]DPCPX in the presence or absence of nonlabeled ligand at the concentrations indicated. Nonspecific binding was determined in the presence of 10 μM CGS15943. The cells were incubated during 3 h at 25°C in a shaking water bath. Subsequently, cells were filtered over GF/B filters on a Millipore manifold and were washed eight times with 2 ml of the above-mentioned buffer. The remaining radioactivity on the filters was determined in a liquid scintillation counter (LKB Rackbeta 1214) 2 h after the addition of 3.5 ml of scintillation cocktail (Emulsifier Safe; Packard, Meriden, CT).
cAMP Generation
CHO cells were washed two times with 200 μl of DMEM/HEPES (50 mM HEPES in DMEM, pH 7.4). Cells were subsequently incubated for 40 min at 37°C with 50 μM rolipram, 50 μM cilostamide, and 5 I.U./ml adenosine deaminase in DMEM/HEPES. Next, either DMEM/HEPES (basal cAMP production), forskolin 10 μM (final concentration), or ligand was added in a volume of 100 μl and the cells were incubated for 15 min at 37°C. The incubations were terminated by aspiration of the medium and subsequent addition of 200 μl of 0.1 N HCl. The 24-well plates were kept at −20°C until cAMP determination.
cAMP Determination
cAMP levels in the cell homogenates were determined essentially as described in De Zwart et al. (1998). Protein kinase A was isolated as described by Leurs et al. (1994). Samples, 50 μl, were incubated for 2.5 h on ice with protein kinase A and 20,000 dpm [3H]cAMP. A calibration curve (0.025–16 pmol) was made with nonlabeled cAMP diluted in 0.1 N HCl. Samples were filtered over GF/C filters and washed two times with 2 ml of ice-cold 50 mM Tris buffer, pH 7.4. Filters were counted 24 h after the addition of scintillation fluid (Emulsifier Safe). Nonspecific binding was determined in the absence of protein kinase A and was subtracted from total binding.
Results
To identify amino acids of the adenosine A2B receptor that might explain the low affinity of this receptor for agonists, the following decision scheme was developed (Fig. 1). In total three filters were applied. Because adenosine A2Breceptors have not yet been the subject of mutation studies, the point mutation data that are available for the other adenosine receptors were used as the first filter. These mutation data were gathered from the G protein-coupled receptor database, the GPCRDB (Horn et al., 1998; the GPCRDB can be accessed at http://www.gpcr.org/7tm/) via the tinyGRAP mutant subdatabase (Beukers et al., 1999). The white amino acids in the so-called snake-like diagram of the human adenosine A2B receptor passed only the first filter (Fig.2). The second filter selected the amino acids for which agonist binding was affected by the mutation. The residues that passed both the first and second filter are shown in black. The third filter consisted of the amino acid alignments that are present in the GPCRDB (Horn et al., 1998). The amino acids that are conserved among the adenosine A2B receptors from different species, but different in the other adenosine receptors were selected for mutation. The chicken A2B receptor was not included because it shares relatively little (<65%) amino acid identity with the other A2B receptors that are >85% homologous (see the phylogenetic tree in the GPCRDB as described by Horn et al., 1998). Computer models of the human A1 and A2A receptor (IJzerman et al., 1994; Kim et al., 1995; Rivkees et al., 1999) as well as the models present in the GPCRDB, which have been constructed from the α-carbon template as defined by Baldwin et al. (1997), confirmed that the selected residues were present in the transmembrane domains and were facing the central core, where adenosine ligand binding is assumed to occur.

Decision scheme to identify amino acids that might explain the low affinity of prototypic agonists for the adenosine A2B receptor. ∗, chicken A2B sequence was not included because it shares relatively little sequence similarity with the other A2B receptor sequences.
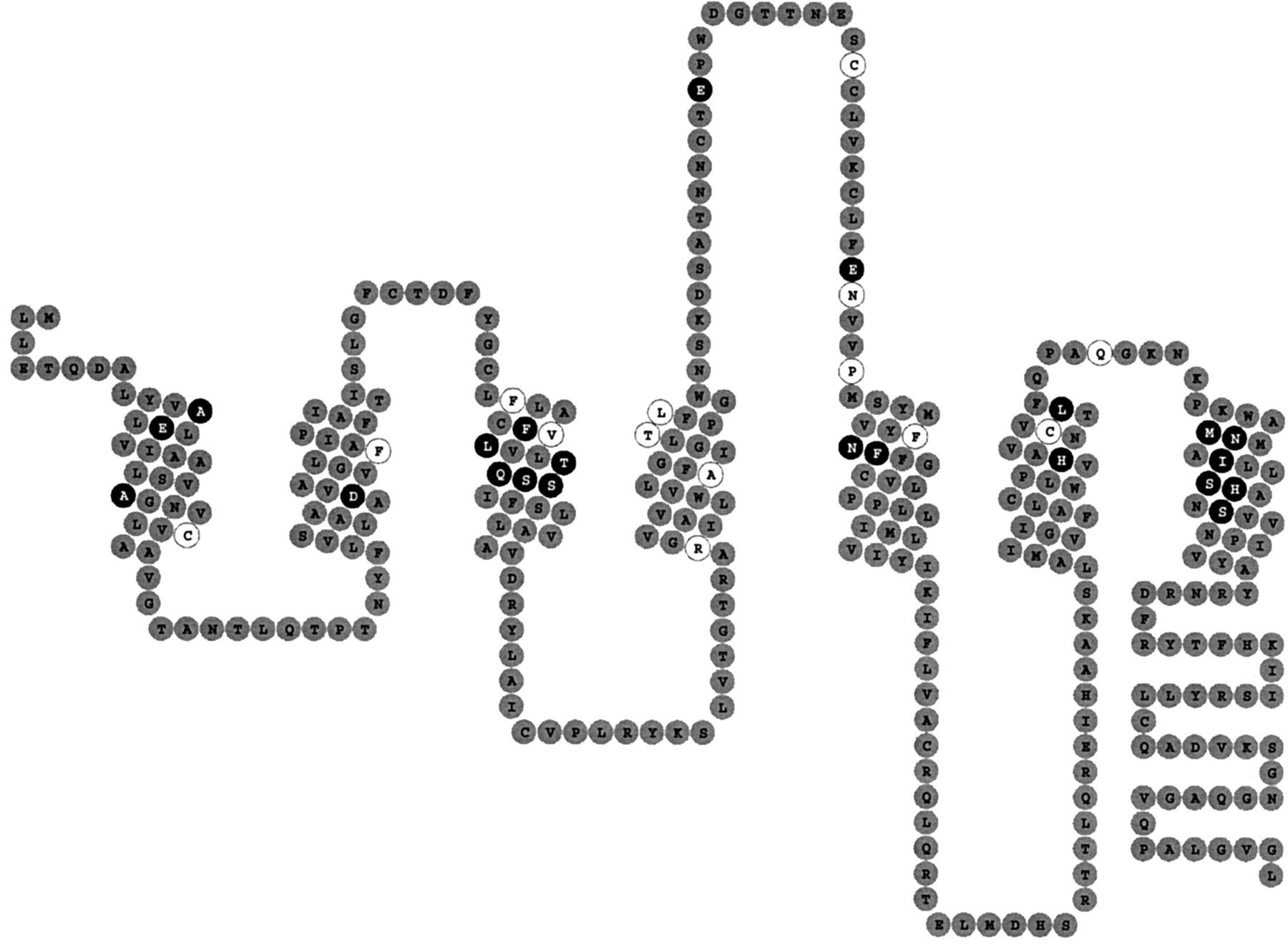
Two-dimensional representation or so-called “snakeplot” of the human adenosine A2B receptor. Amino acid residues that have passed only the first filter are shown in white, those that passed the first two filters are shown in black.
Based on this decision scheme two amino acids were selected. Their position in the transmembrane domains and the sequence alignment of the transmembrane domains are represented in Fig.3. Because we aimed for “gain-of-function” mutations we mutated these residues in the A2B receptor to their counterparts in the most closely related human adenosine A2A receptor. The selected mutations were A12T (TM1) and N273Y (TM7). Ala23 (TM1) was not considered as mutation of the corresponding Pro25 of the human A1 receptor to Leu, as present in the human adenosine A2A receptor, strongly decreased the affinity of agonists (Rivkees et al., 1999). Hence, this amino acid is not a likely candidate to explain the low affinity of A2B receptors versus all other adenosine receptor subtypes.

Sequence alignments of transmembrane domains 1 and 7 of the adenosine receptors. The amino acids that were selected for mutation based on the first two filters (mutation data and affected agonist binding) are depicted in bold face. Those residues that also passed the third filter based on the additional sequence alignment criterion are underlined. None of the amino acids in transmembrane domains 2 to 6 passed the third filter. ∗, Pro25 of the human adenosine A1 receptor has been mutated to Leu as present in the human adenosine A2A receptor (Rivkees et al., 1999). This mutant had a strongly decreased affinity for agonists, making this residue an unlikely candidate to mutate in the human A2Breceptor.
The mutants were made as described under Experimental Procedures and COS-7 cells were transiently transfected with the different DNA constructs. An overview of theK i values of NECA for the wt and mutant A2B receptors, as determined in radioligand binding experiments, is presented in Table1. The nontransfected and mock-transfected (pcDNA3) COS-7 cells did not show any specific binding, indicating that endogenous A2B receptors did not interfere with the assay. Transfection of the original DNA construct, containing the 5′ upstream nucleotide sequence, resulted in a receptor with a similar affinity for NECA of 2.6 μM (1.8–3.5,n = 2) versus 2.2 ± 0.6 μM for the wt receptor. Besides radioligand binding, we also studied cAMP production upon stimulation with NECA. NECA, at a concentration of 100 μM, did not induce cAMP production in either the nontransfected or the mock-transfected CHO cells. The EC50 values of NECA for the wt and the other mutant receptors were comparable and varied from 0.8 μM for the A12T receptor, to 1.5 μM for the wt and N273Y receptor. Again, removal of the 5′ upstream nucleotide sequence had no effect because the EC50 value of NECA for this construct was 3.7 μM.
Affinity of NECA for the wt and mutant human A2B receptors
Because neither the affinity nor the EC50 value of NECA was much affected by the mutations, we decided to explore several substitution positions of the adenosine molecule. Thus, next to NECA (5′-position) we tested R-PIA (N6-position) and 2Cl Ado (2-position). The results of these experiments are summarized in Table 2. Because the only significant change in affinity was observed for 2Cl Ado on the N273Y human A2B receptor, we decided to explore the C2 region of agonists in greater detail.
Affinities of R-PIA and 2Cl Ado for wt and mutant A2B receptors
A series of structurally diverse agonists with high affinity and selectivity for the adenosine A2A receptor were tested to explore the C2 region (Table3). CGS21680, the reference A2A compound, had a very low affinity for the wt receptor. However, the affinity of this compound for the N273Y receptor was slightly, but significantly increased. Two other ligands,3 and 4, with a flexible side chain at the 2-position had improved affinity for the N273Y receptor. The affinity of compound 2, with a less flexible side chain, for the wt receptor was comparable with the affinity of compounds 3 and4. On the other hand, the affinity of compound 2for the N273Y receptor was 21-fold higher than the affinity for the wt receptor. 2-(1-Hexynyl)adenosine (1), with a rigid triple carbon bond at the 2-position of adenosine, had a 5-fold higher affinity for the wt receptor, with respect to compound 2. The affinity of 2-(1-hexynyl)adenosine (1) for the N273Y receptor was improved over 10-fold, resulting in a 61-fold selectivity for the N273Y receptor versus the wt receptor. Its affinity for the A12T receptor was not increased, being 46 μM (51–41 μM,n = 2).
Affinity of N273Y and the wt human A2B receptors for 2-substituted agonists and representative antagonists
To investigate the requirements for the side chain at the 2-position of adenosine in even greater detail we synthesized (R)-PHP Ado (5, LUF5599) and (S)-PHP Ado (6, LUF5600). Both the wt and the N273Y receptor showed a 10-fold stereoselectivity. The affinities of the (R)- and (S)-enantiomers for the wt receptor were 10 ± 4.5 and 1.7 ± 0.5 μM, respectively (Table 3). The compounds were approximately 4- to 10-fold more potent on the N273Y receptor, 2.9 ± 1.1 and 0.19 ± 0.1 μM (Table 3).
In contrast to the affinity of the agonists, the affinities of the reference antagonists ZM241385, SCH58261, and XAC were not affected by the N273Y mutation (Table 3). In addition, the affinities of ZM241385, SCH58261, and XAC for the A12T receptor were also unaltered, and were 20 nM (9.5–31 nM, n = 2), 440 ± 80 nM (n = 3), and 12 nM (6.7–17 nM, n = 2), respectively.
2-(1-Hexynyl)adenosine (1), the agonist with the highest affinity and selectivity for the N273Y mutant versus the wt receptor, was tested for its ability to stimulate the production of cAMP via the wt and the N273Y receptors. The endogenous level of adenosine A2B receptors in COS-7 cells is too high to perform cAMP experiments (data not shown). CHO cells were used instead because 100 μM NECA did not induce cAMP production in either nontransfected or mock-transfected cells. Radioligand binding studies could not be carried out in the CHO cells because the expression levels were too low to obtain any specific binding (data not shown). Figure4 shows the production of cAMP upon stimulation with 2-(1-hexynyl)adenosine (1).

cAMP production induced with 2-(1-hexynyl)adenosine (1) in CHO cells transiently transfected with the N273Y human A2B receptor (▪) and the wt human A2B receptor (■).
The EC50 values of 2-(1-hexynyl)adenosine (1) were 0.56 ± 0.10 and 5.6 ± 1.1 μM for the N273Y and the wt receptor, respectively. The amount of cAMP produced at a concentration of 100 μM 2-(1-hexynyl)adenosine (1) varied from 62 ± 22 pmol of cAMP/106 cells for the N273Y receptor to 49 ± 11 pmol of cAMP/106 cells for the wt receptor. Basal levels were low, i.e., approx. 10 pmol of cAMP/106 cells and approx. 8 pmol of cAMP/106 cells for the N273Y and the wt receptor, respectively. Maximal levels of cAMP were obtained in the presence of 10 μM forskolin and were 475 and 560 pmol of cAMP/106 cells, for the mutant and the wt receptor, respectively.
To evaluate an alternative decision scheme to select amino acids for mutation we used a combination of sequence alignments and three-dimensional computer models of the four adenosine receptor subtypes. The receptor models, although speculative, were instrumental in predicting which amino acids face the inner core formed by the seven TM domains and yielded three candidates for mutation. These residues, Val11 and Phe59 as well as the not strictly conserved neighbor residue Leu58, were also mutated to the corresponding residues of the human A2A receptor. Evaluation of these mutants indicated that neither the affinities, nor the EC50 values of NECA, R-PIA, and 2Cl Ado were affected in the V11I and the L58V mutant receptors (data not shown). The F59L mutant did not show specific binding or stimulation of cAMP production with NECA (data not shown).
Discussion
This study is the first to describe mutants of adenosine A2B receptors. Two mutations were selected based on the decision scheme presented in Fig. 1. Mutation data available for other adenosine receptors and sequence alignments, both present in the GPCRDB (Horn et al., 1998), proved to be essential elements of the scheme. Some general strategies applied to select amino acids for mutation have been described by Kristiansen et al. (1996). These strategies can be subdivided into two categories, either by “educated guess” or “at random”. Examples of the former include investigations of residues that are either part of consensus sites, conserved among subtypes of receptors, or present as polymorphisms in the general population. Examples of more random approaches are besides random mutagenesis itself, the construction of chimeric receptors, the exchange of amino acids from one receptor subtype to another, “Ala scans”, and the mutation of amino acids that share a physical property (e.g., charge). Fewer reports exist where multiple parameters have been combined, such as ligand binding data and sequence alignments (Meng et al., 1996; Akeson et al., 1997).
Our approach (Fig. 1) combined knowledge-based (sequences and mutation data) and rule-based (alignments) information and helped us in a substantial further selection to yield only two likely candidates for mutation, A12 and N273. An alternative, mainly rule-based (alignments and receptor models) strategy did not seem very effective because it yielded three candidates (V11, L58, and F59) that did not show any gain in affinity for agonists.
The affinity of the wt adenosine A2B receptor for agonists and antagonists agreed very well with available radioligand binding and cAMP data (Klotz et al., 1998; Cristalli et al., 1998;Linden et al., 1999). Both the A12T and the N273Y mutant receptor showed specific [3H]DPCPX binding as well as dose-dependent cAMP production upon stimulation with NECA. The affinity and potency of NECA were the same for both the wt and the mutant receptors.
Sequence alignments show that the A12T mutant A2Breceptor is the equivalent of the G14T mutant human adenosine A1 receptor. This mutant A1receptor had a 7-fold increased affinity for NECA, a 93-fold increased affinity for 2Cl Ado, and a 137-fold increased affinity for R-PIA (Rivkees et al., 1999). The affinity of NECA, 2Cl Ado, and R-PIA for the A12T receptor human A2B receptor was not affected. Mutation of the neighboring residue, Val11 to Ile, in the human A2B receptor did not affect the binding of any of the agonists, NECA, 2Cl Ado, or R-PIA, either. Apparently, the threonine residue in the A12T mutant does not serve the same role in the adenosine A2B receptor as the corresponding threonine in the G14T adenosine A1 receptor. These differences cannot be explained by the experimental setup used because both radioligand binding assays were carried out with [3H]DPCPX on intact, transiently transfected COS-7 cells. In addition, also the affinity for the prototypic antagonists SCH58261, XAC, and ZM241385 was not affected by the A12T mutation.
The N273Y mutant showed a slight, but significant decrease in affinity for 2Cl Ado. Moreover, this mutant had an increased affinity for the reference A2A receptor agonist CGS21680. Other adenosine derivatives with small substituents at the 2-position, such as 2-hydrazino (intermediate of the synthesis of compound 3) and 2-thio adenosine did not displace [3H]DPCPX binding at a concentration of 10 μM.
The 3-fold reduction in affinity for 2Cl Ado and the 3-fold increase in affinity for CGS21680 prompted us to study several other large substituents at the 2-position. Thus, we selected four structurally diverse 2-substituted adenosine analogs with a high affinity and selectivity for the A2A receptor. All four compounds had a higher affinity for the N273Y receptor than for the wt receptor. The affinity of compounds 3 and 4, both with flexible side chains, was only slightly higher for the N273Y receptor versus the wt receptor. Compound 2, with a more rigid side chain, had a similar affinity for the wt receptor compared with compounds 3 and 4. The increase in affinity of compound 2 for the N273Y receptor resulted in a 21-fold selectivity for the mutant versus the wt receptor. Finally, the compound with the rigid triple carbon bond, 2-(1-hexynyl)adenosine (1), had a slightly improved affinity for the wt receptor and a submicromolar affinity for the N273Y receptor, leading to a 61-fold selectivity for the mutant receptor versus the wt receptor. This increase in affinity is specific for the N273Y mutant receptor because the affinity of 2-(1-hexynyl)adenosine (1) for the A12T mutant receptor was 46 μM (51–41 μM, n = 2). These data suggest a direct interaction of Tyr273 with the triple carbon bond as present in 2-(1-hexynyl)adenosine. The more lipophilic nature of the tyrosine versus the wt asparagine residue as well as the ability of tyrosine to form π-π interactions would indeed favor an interaction with a triple carbon bond over the -O-, -NH-, or double carbon bonds as present in the other 2-substituted analogs.
The increased affinity of 2-(1-hexynyl)adenosine (1) for the N273Y receptor was also reflected in its ability to activate the receptor. The EC50 value for the N273Y receptor, 0.56 ± 0.10 μM, was 10-fold higher than the EC50 value for the wt receptor, 5.6 ± 1.1 μM. In contrast to our findings, Vittori et al. (1999) reported that 2-(1-hexynyl)adenosine had an EC50 value >100 μM on wt human A2B receptors stably transfected in CHO cells (Vittori et al., 1999).
The residue equivalent to N273 has also been subjected to mutation in the human adenosine A2A receptor. That residue, Tyr271, was changed into Ala, resulting in the absence of specific binding of both [3H]XAC and [3H]CGS21680 (Kim et al., 1995). The authors indicated that mutation of this residue into Phe, His, or Arg did not affect the affinity of either agonists or antagonists. According to their model this Tyr residue points toward the interface of TM1 and TM7, suggesting that this amino acid is not involved in ligand binding.
The data obtained with 2-(1-hexynyl)adenosine (1) suggested a different role for the Tyr in the N273Y A2Breceptor mutant. Hence, we decided to investigate the alkynyl side chain at the 2-position of adenosine in greater detail. To that end we synthesized (R)- and (S)-PHP Ado (5,LUF5599 and 6, LUF5600). This alkynyl side chain was chosen because the NECA analog of PHP Ado had been tested as a racemate byCristalli et al. (1998) and proved to be the most active on the A2B receptor within a series of 2-alkynyl NECA derivatives.
The affinity of (S)-PHP Ado was higher than of (R)-PHP Ado for both the wt and the N273Y mutant receptor. The fact that the stereoselectivity of (R)-PHP Ado (5, LUF5599) and (S)-PHP Ado (6, LUF5600) was retained in the N273Y mutant receptor is further evidence that the overall structure of the receptor is not affected by the mutation. Interestingly, Klotz et al. (1999) have tested the (R)- and (S)-enantiomers of the NECA analog on human A1, A2A, and A3 receptors. They found stereoselectivity for the S- over the R-compound on the A2A and A3 receptors, but not on the A1 receptor. Apparently, the A2A, A2B, and A3 receptors share a common amino acid(s) that is responsible for the observed stereoselectivity.
The submicromolar affinity of (S)-PHP Ado (6, LUF5600) and 2-(1-hexynyl)adenosine (1) indicated that the N273Y mutant receptor is a gain-of-function mutant. Both compounds 2-(1-hexynyl)adenosine (1) and (S)-PHP Ado shared the same affinity for the N273Y mutant receptor. On the wt receptor, however, (S)-PHP Ado had an increased affinity, 1.7 ± 0.5 μM, in comparison to 2-(1-hexynyl)adenosine (1), 11 ± 4.3 μM. In addition to the fact that (S)-PHP Ado is the first non-NECA agonist with a low micromolar affinity, these data also demonstrate that selection of the proper substituent at the 2-position yields agonists with increased affinity for the wt receptor. Mutation of Asn273 to Tyr does not explain, however, why 2Cl-Ado and non-C2-substituted agonists such as NECA have low affinity for adenosine A2Breceptors, suggesting that additional residues on the receptor impede agonist binding.
Interestingly, the wt chicken adenosine A2Breceptor has a Tyr rather than Asn at the position equivalent to Asn273 of the wt human receptor (Worpenberg et al., 1997). From our data it would be worthwhile to study the 2-alkynyladenosines on a chicken A2B receptor preparation (Tasca et al. 1999) to investigate whether their affinities are higher than on wt human receptors.
In conclusion, our strategy to identify amino acids that may be responsible for the fact that adenosine A2Breceptors have a low affinity for agonists has yielded a mutant receptor, N273Y, that shows strong (up to 61-fold) gain of function for 2-alkynyladenosines. This finding suggests that N273 is located near the 2-position of agonists. The increased affinity of (S)-PHP Ado for the wt receptor encourages us to continue our search for agonists with a high affinity for A2B receptors.
Footnotes
- Received April 26, 2000.
- Accepted September 11, 2000.
-
Send reprint requests to: Margot Beukers, Division of Medicinal Chemistry, Leiden/Amsterdam Center for Drug Research, Gorlaeus Laboratories, Einsteinweg 55, P.O. Box 9502, 2300 RA Leiden, the Netherlands. E-mail:beukers{at}chem.leidenuniv.nl
Abbreviations
- NECA
- 5′-N-ethylcarboxamidoadenosine
- R-PIA
- (R)-N6-(phenylisopropyl)adenosine
- CGS21680
- 2-[4-(2-carboxyethyl)phenylethylamino]-5′-N-ethylcarboxamidoadenosine
- DPCPX
- 8-cyclopentyl-1,3-dipropylxanthine
- ZM241385
- 7-amino-2-(2-furyl)-5-[2-(4-hydroxyphenyl)ethyl]amino[1,2,4]-triazolo[1,5-a][1,3,5]triazine
- CGS15943
- 5-amino-9-chloro-2-(2-furyl)[1,2,4]triazolo[1,5-c]quinazoline
- SCH58261
- 5-amino-2-(2-furyl)-7-(2-phenylethyl)-pyrazolo[4,3-e][1,2,4]triazolo[1,5-c]pyrimidine
- MRS1754
- 8-[4-[((4-cyanophenyl)carbamoylmethyl)oxy]phenyl]-1,3-di(n-propyl)xanthine
- 2Cl Ado
- 2-chloroadenosine
- DMEM
- Dulbecco's modified Eagle's medium
- MPE Ado
- 2-[2-(4-methylphenyl)ethoxy]-adenosine
- XAC
- xanthine amine congener, 8-[4-[[[(2-aminoethyl)carbonyl]methyl]oxy]-phenyl]-1,3-di-n-propylxanthine
- PHP Ado
- 2-((3-hydroxy-3-phenyl)propyn-1-yl)adenosine
- CHO
- Chinese hamster ovary
- GPCRDB
- G protein-coupled receptor database
- TM
- transmembrane domain
- wt
- wild-type
- RSC
- RPMI-1640 with 2% fetal calf serum
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}