


Abstract
The specific involvement of NAD(P)H:quinone oxidoreductase 1 (NQO1) in the bioactivation of quinone prodrugs has been shown through the use of the inhibitor of NQO1, dicoumarol. Disadvantages of using dicoumarol to inhibit NQO1 include its lack of specificity and its competitive mechanism of inhibition. The concentration of dicoumarol required for inhibition of NQO1 varies according to the substrate under evaluation, which may lead to either false conclusions of the involvement of NQO1 or the alteration of other cellular processes. We have reported previously on the chemical and biochemical properties of ES936, a mechanism-based inhibitor of NQO1 in cell-free systems. In this study, we investigated the effects of ES936 in cellular systems. ES936 (100 nM) inhibits more than 95% of NQO1 activity within 30 min and is stable in complete media at this concentration for a minimum of 2 h. The duration of inhibition is cell line-specific because a new protein must be generated for resumption of activity. ES936 abrogates the toxicity of streptonigrin, with greater effects seen in cell lines expressing higher levels of NQO1. ES936 does not inhibit other cellular reductases, nor does it alter cellular levels of acid-soluble thiols. Some evidence of DNA strand breaks was observed at the concentrations of ES936 required for the inhibition of NQO1 activity. From our studies, we propose the use of ES936 (100 nM) as a mechanism-based inhibitor of NQO1 in cellular systems and for use as a component of the routine activity assay for NQO1.
NAD(P)H:quinone oxidoreductase 1 (NQO1; DT-diaphorase, EC 1.6.99.2) is an obligate two-electron reductase that is present at high levels throughout many human solid tumors (Cresteil and Jaiswal, 1991; Malkinson et al., 1992; Mikami et al., 1998). NQO1 is generally considered to be a detoxification enzyme (Ernster, 1967; Lind et al., 1982; Thor et al., 1982) that reduces quinones directly to hydroquinones, eliminating the formation of reactive oxygen species generated by redox cycling. NQO1 may also act as an antioxidant enzyme by regenerating antioxidant forms of ubiquinone and vitamin E quinone (Beyer et al., 1996; Siegel et al., 1997). Alternatively, NQO1 may also bioactivate quinones to reactive DNA-alkylating agents. It is in this capacity that NQO1 may be useful in the treatment of cancer, and there has been considerable interest in the development of chemotherapeutic prodrugs that are specifically bioactivated by NQO1.
The involvement of NQO1 in biochemical reactions has historically been determined by inhibiting its enzymatic activity with dicoumarol, a nonspecific competitive inhibitor that binds to the pyridine nucleotide binding site in NQO1 (Hosoda et al., 1974; Hollander and Ernster, 1975). Dicoumarol has been shown to inhibit a variety of enzymes (Wang et al., 1982; Segura-Aguilar et al., 1986; Ross et al., 1993), including glutathione (GSH) transferases and GSH peroxidase II (Mays and Benson, 1992; Karczewski et al., 1999), as well as NADH-ubiquinone reductase (Tampo and Yonaha, 1996). It may also affect mitochondrial oxidative phosphorylation (Karczewski et al., 1999), potentiate tumor necrosis factor-α–induced apoptosis in HeLa cells, and inhibit stress-activated protein kinase and nuclear factor-κB activation (Cross et al., 1999). Many of the ancillary effects of dicoumarol in cells are probably caused by the relatively high micromolar concentrations necessary to inhibit NQO1 activity. Even at micromolar concentrations of dicoumarol, only partial inhibition of NQO1 may occur, depending on the quinone substrate (Preusch et al., 1991; Nakamura and Hayashi, 1994). Therefore, the data obtained with the use of dicoumarol cannot solely be interpreted in terms of NQO1 inhibition.
We reported recently the chemical and biochemical properties of ES936, a mechanism-based inhibitor of NQO1, in cell-free systems (Winski et al., 2001a). ES936 undergoes reduction by NQO1 followed by the loss of para-nitrophenol to generate a reactive iminium species that alkylates one of two tyrosine residues in the active site, resulting in irreversible inhibition of the enzyme. Our previous work used purified recombinant human wild-type NQO1 and allowed for the investigation of the specific mechanism of ES936 interaction with the human wild-type NQO1*1 protein. However, the biological consequences of the use of ES936 in cells are not known and need to be defined before beginning routine use of this agent to inhibit NQO1 in cellular systems.
ES936, although a potent and specific NQO1 inhibitor, is a quinone and therefore may have deleterious effects in cellular systems. This study examines the biochemical, cytotoxic, and genotoxic effects of ES936 in cells that express a range of NQO1 activities. To measure the effectiveness of ES936 as an inhibitor of NQO1, we evaluated the ability of ES936 to abrogate the toxicity of an antitumor quinone bioactivated by NQO1. ES936, as a mechanism-based inhibitor of NQO1, provides a potentially useful tool to clarify the precise in vivo role of NQO1 without the ancillary effects of the NQO1 inhibitor, dicoumarol.
Materials and Methods
Materials. 2,6-Dichlorophenol-indophenol (DCPIP), MTT, NADH, dicoumarol, streptonigrin GSH, and 5,5′-dithio-bis(2-nitrobenzoid acid) were purchased from Sigma Chemical Co. (St. Louis, MO). ES936 was synthesized as described previously (Beall et al., 1998). Human and rat liver microsomes were obtained from BD Gentest (Woburn, MA).
Cell Culture. HCT116 and HT-29, human colon carcinoma cell lines, and the human breast cancer cell line MDA-MB-468 (MDA468) were obtained from the American Type Culture Collection and were grown as monolayers in either minimal essential medium or RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM l-glutamine, 100 U/ml of penicillin, and 100 μg/ml streptomycin (complete media) at 37°C in a humidified atmosphere with 5% CO2. The MDA468 NQ16 cell line is a NQO1 stable transfectant cell line generated from the human breast cancer cell line, MDA468. The parental MDA468 cell line is homozygous for a polymorphism in NQO1 (NQO1*2) that results in extremely low NQO1 activity (<5 nmol DCPIP/min/mg protein). The cytomegalovirus-driven mammalian expression vector pcDNA3.0 containing human wild-type NQO1 cDNA (construction described by Gustafson et al., 1996) was used to transfect MD468 cells as described previously (Winski et al., 1998). The stable expression of NQO1 has been monitored by repeated measurement of NQO1 activity as well as by NQO1 immunoblot analysis, as described previously (Gustafson et al., 1996).
Enzyme Activity Assay. NQO1 activity was measured as the rate of dicoumarol-inhibitable DCPIP reduction in cell cytosolic samples as described previously (Ernster, 1967) with modifications (Benson et al., 1980) and normalized to total cytosolic protein (Lowry et al., 1951). Briefly, cells were lifted with trypsin/EDTA, neutralized with complete media, and centrifuged at 1500 rpm for 5 min (4°C); the supernatant was then aspirated, and the pellet was resuspended in 200 to 500 μl of buffer A (25 mM Tris-HCl, 250 μM sucrose, 5 μM FAD). The cell suspension was probe-sonicated on ice for 5 s and then centrifuged at 13,000 rpm for 15 min with the supernatant removed to a clean tube and kept on ice for analysis. NQO1 activity was measured in 1-ml reactions (27°C) containing 25 mM Tris-HCl, pH 7.4, 0.7 mg/ml bovine serum albumin, 200 μM NADH, and 40 μM DCPIP. Reactions were performed in the presence and absence of 20 μM dicoumarol. Cytochrome P450 reductase was measured in cellular samples as the NAD(P)H-dependent reduction of cytochrome c according to the method used by Vermilion and Coon (1978). The activity of cytochrome b5 reductase was measured as the NADH-dependent reduction of cytochrome c according to the method used by Mihara and Sato (1978).
NQO1 Inhibition by ES936. The inhibition of NQO1 activity by ES936 was monitored over a time and concentration range to determine the lowest concentration of ES936 that would inhibit >95% of NQO1 activity over a given duration of exposure. Cells were treated with varying concentrations of ES936 in complete media for 30, 60, or 120 min, at which time cell cytosols were prepared and assayed for NQO1 activity.
ES936 Biostability. To determine the biostability of ES936 in tissue culture media, the inhibitor was added to complete media at a final concentration of 100 nM and maintained at 37°C in a humidified atmosphere with 5% CO2. At various times afterward, cells that were 60 to 70% confluent were dosed with an aliquot of the media containing ES936 for 30 min, at which time the NQO1 activity was measured as described above.
Growth Inhibition Assays. Toxicity was determined by growth inhibition (MTT) assays. HCT116 cells and MDA468 NQ16 cells were seeded at 1000 to 3000 cells/well, respectively, in 96-well plates and were allowed to attach for 16 to 20 h. Plates (in triplicate) were treated with ES936 in complete media or with streptonigrin in serum-free media for 2 h. Additional plates were pretreated with ES936 (100 nM) in complete media for 30 min before exposure to streptonigrin in serum-free media for 2 h. After drug treatment, the media were removed and replaced with complete media. Cells were then allowed to grow for 3 to 5 days. Cell viability was assessed by measuring the NADH-dependent reduction of MTT to a formazan product that was extracted from cells with dimethyl sulfoxide. The optical density of the extract was determined at 550 nm with a microplate reader. IC50 values, defined as the concentration that results in a 50% reduction in cell density (from untreated controls), were calculated from a minimum of three separate experiments.
Soluble Thiols. Cells were seeded 24 h before treatment, and all cells used were approximately 60 to 70% confluent. Soluble thiols were determined according to a modified method used by Sedlak and Lindsay (1968) after treatment with 100 nM ES936 in complete media for the indicated times. After dosing, media were aspirated, and the cells were rinsed with phosphate-buffered saline, lifted with trypsin/EDTA, and neutralized with complete media. The cell suspension was vortexed, and 1-ml aliquots were distributed into two tubes on ice. Cells were centrifuged at 1,500 rpm for 5 min (4°C), and the supernatant was aspirated. The cell pellet in one tube was rinsed with phosphate-buffered saline, vortexed lightly, recentrifuged, and aspirated, and the pellet was brought up in buffer A for determination of protein concentration according to the methods used by Lowry et al. (1951). The cell pellet of the second tube was lysed in 5% trichloroacetic acid, vortexed immediately for 10 s, and then centrifuged at 5,000 rpm for 10 min to pellet cellular protein. The supernatant was removed to a glass tube containing 2 ml of 0.4 M Tris-HCl, pH 8.9, and 5,5′-dithio-bis(2-nitrobenzoid acid) was added to a final concentration of 100 μM. Samples were vortexed and incubated for 5 min at room temperature, and the absorbance at 412 nm was determined. Results were expressed as μM acid-soluble thiols/mg protein calculated from a reduced glutathione calibration curve.
NAD(P)H Oxidation. Quinone-stimulated microsomal-dependent NAD(P)H oxidation was measured spectrophotometrically at 340 nm in 1-ml reactions at 32°C. Reactions were performed in 100 mM potassium phosphate buffer, pH 7.4, containing 200 μM NAD(P)H, 330 mg rat microsomal protein, and 50 μM quinone (ES936, menadione, and β-lapachone). Reactions were started with the addition of quinone, and the decrease in absorbance at 340 nm was monitored for 1 min.
Comet Assay. DNA damage was evaluated by the single-cell gel electrophoresis method, commonly known as the alkaline comet assay. The comet assay is a sensitive method used to evaluate DNA strand breaks in single cells. Briefly, cells are treated with drug before embedding in a thin coating of agarose on a microscope slide. The slides are submerged in an alkaline buffer and subjected to horizontal electrophoresis. Single- or double-strand breaks cause a relaxation of supercoiled DNA, allowing migration toward the anode, forming a “comet”. The percentage of DNA in the comet tail provides an indication of the degree of DNA damage. For this study, cells were incubated in complete media with 100, 250, or 500 nM ES936. After a 2-h incubation period, the cells were lifted, immediately placed on ice, and processed as described previously (Winski et al., 2001b). Comet slides were examined on a Nikon Eclipse TE300 (Nikon, Tokyo, Japan) equipped with epifluorescence capabilities. An excitation filter of 530 nm and a magnification of 400× was used to visually score a minimum of 200 comets on each slide. Each comet was visually identified as belonging to one of four classes according to the degree of DNA damage observed, in a fashion similar to that of Collins et al. (1997). Slides were scored blinded. Comet classes are as follows and are shown in Fig. 1: class 0, intact nuclei, no DNA damage present; class 1, well-defined nucleus with light tail formation, <20% DNA in the tail; class 2, defined nucleus with 20 and 75% DNA in the tail; class 3: nucleus is no longer well-defined and tail consists of more than 75% of the DNA.

Representative images of comets in class 0, no DNA damage present; class 1, well-defined nucleus with light tail formation, <20% DNA in the tail; class 2, defined nucleus with 20 and 75% DNA in the tail; and class 3, nucleus is no longer well-defined and tail consists of more than 75% of the DNA.
To verify the visual classification system, random fields of comet images from all treatment groups were captured with a Nikon Coolpix 990 digital camera mounted to the microscope. All single-comet images (no image overlap) in each photo were copied into individual files with the use of Adobe Photoshop 6.0 (Adobe Systems, Mountain View, CA). The single images were visually scored and placed into one of the four classes. The percentage of DNA in the tail was then determined for each image using Euclid Comet Analysis software (St. Louis, MO). The mean percentage of DNA in the tail was calculated and assigned to each comet class.
A score for each slide was derived by multiplying the comet class (0–3) by the percentage of the comets in that class and then summing all scores to obtain a total for the slide. Scores for each slide could range between 0 (100% of comets in class 0) and 300 (100% of comets in class 3) (units are arbitrary) and represent the overall DNA damage of the cell population for a specific treatment.
Statistical Analysis. Statistical analysis was performed using NCSS software (Kaysville, UT). The two-sample t test was used to evaluate differences between two independent groups, and P values <0.05 were considered significant. Differences between more than two independent groups were evaluated with analysis of variance followed by Scheffé F test for multiple comparisons. Data are presented as means ± S.E.M.
Results
Inhibition of NQO1 by ES936. ES936 efficiently inhibited NQO1 activity in all cell lines tested; studies demonstrate efficient inactivation of NQO1 at concentrations up to 100 nM (Fig. 2). The minimum concentration that inhibits more than 95% of NQO1 activity varied between cell lines, presumably because of the variability in NQO1 levels. The MDA NQ16 cell line had the highest NQO1 activity (2400 ± 144 nmol DCPIP reduced/min/mg protein) of all lines examined in this study and required the highest concentration of ES936 (100 nM) to achieve more than 95% inhibition, regardless of the duration of exposure. A concentration of 25 nM ES936 inhibits more than 95% of the NQO1 activity in HCT116 cells (average NQO1 activity = 300 ± 25 nmol DCPIP reduced/min/mg protein). More than 95% of NQO1 activity is inhibited in HT29 cells (average NQO1 activity = 1500 ± 43 nmol DCPIP reduced/min/mg protein) by 50 nM ES936 for 60 min or by 100 nM ES936 for 30 min (data not shown).

Inhibition of NQO1 by ES936 in the MDA468 NQ16 cell line (A) and the HCT116 cell line (B). NQO1 activity was measured after 30 min (○), 60 min (□), or 120 min (▵) of exposure at each ES936 concentration. Values given are the means ± S.E.M. of a minimum of three independent experiments. Error bars are not visible in most cases.
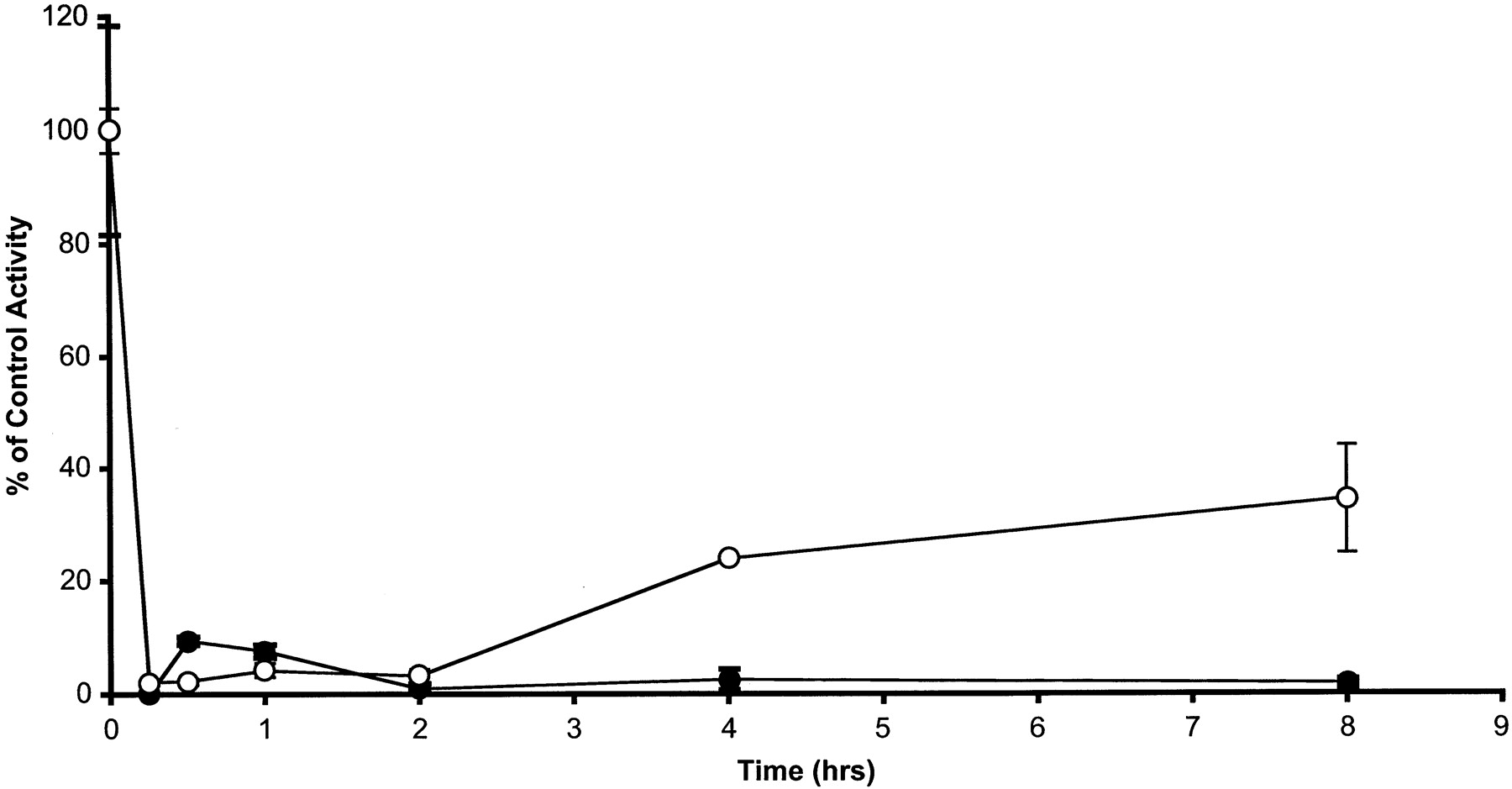
Biostability of ES936. The biostability of 100 nM ES936 in complete media was measured over time (Fig. 3). In these experiments, ES936 was preincubated in complete media (minus cells) for the indicated times, after which the ES936-containing media was added to cells, and NQO1 activity was measured after 30 min. In experiments with the HCT116 cell line, more than 95% inhibition of NQO1 activity was observed at all time points. In experiments with the very high NQO1 activity cell line MDA468 NQ16, a reduction in the ability of ES936 to inhibit NQO1 was observed between 2 and 4 h of incubation in media.

Inhibition of NQO1 activity in MDA468 NQ16 cells (○) and HCT 116 cells (•) by ES936 after incubation of 100 nM ES936 in media at 37°C for the durations indicated. NQO1 activity was measured in cell sonicates after a 30-min dosing period. Results are the mean ± S.E.M. of a minimum of three independent experiments.
Growth-Inhibitory Activity of ES936. The growth-inhibitory activity of ES936 was measured by use of the MTT assay in the MDA468 NQ16 and HCT116 cell lines because these two lines represent the upper and lower limits of NQO1 activity, respectively, examined in this study. IC50 values are shown in Table 1. The IC50 values for the cell lines used in this study are 8 to 20 times higher than the concentration of ES936 needed for effective inhibition of NQO1. The ability of ES936 to affect the toxicity of compounds that are metabolized by NQO1 was also investigated. ES936 pretreatment significantly increased the IC50 value for streptonigrin by approximately 6-fold for the MDA468 NQ16 cell line and 2-fold for the HCT116 cell line, which is consistent with a bioactivation role of NQO1 in streptonigrin metabolism, as described previously (Beall et al., 1996).
Calculated IC50 values for MDA468 NQ-16 and HCT116 cell lines
IC50 values are the average of a minimum of three separate measurements ± S.E.M.
Other Reductases. We examined the effects of ES936 on cytochrome P450 reductase and cytochrome b5 reductase in cell lines and purified microsomal preparations to ascertain whether other cellular reductases in addition to NQO1 were inhibited by this compound. As shown in Table 2, treatment with 100 nM ES936 for 30 min did not inhibit the activity of cytochrome P450 reductase or cytochrome b5 reductase in the cell lines tested, although more than 95% of the NQO1 activity was inhibited. Although the activities of these two reductases were low in these cell lines, there was no significant difference between treated and untreated cells, regardless of initial NQO1 activity. Furthermore, we evaluated the effects of ES936 at 100 nM in purified human and rat liver microsomes supplemented with NADH/NAD(P)H (data also shown in Table 2). As seen in the cell lines tested, the activities of cytochrome P450 reductase or cytochrome b5 reductase were not significantly different between treated and control human or rat microsomes. In these experiments, we also observed no effect of ES936 at concentrations up to 10 μM on the activities of either cytochrome P450 reductase or cytochrome b5 reductase (data not shown).
Average reductase activity ± ES936 pretreatment at 100 nM concentration for 30 min
Results are means of a minimum of three independent experiments ± S.E.M.
Cellular Nonprotein Thiols. To determine the effect of ES936 on cellular redox homeostasis, total cellular thiols were measured in vehicle-treated (control) cells and cells treated with 100 nM ES936 for various times (Fig. 4). Statistical analysis revealed no difference between treated and control cells for either cell line at any time point.

Levels of acid-soluble thiols in MDA468 NQ16 cells (○) and HCT116 cells (•) compared with cells treated with 100 nM ES936 for 15, 30, 60, or 120 min (□ and ▪, respectively). Values represent the means ± S.E.M. of a minimum of three independent experiments. There was no statistical difference (α = 0.05) in cellular thiols between control and ES936-treated cells at each time point (two-sample t test), or between all time points as determined by analysis of variance followed by Scheffé test for multiple comparisons.
Microsomal-Dependent NAD(P)H Oxidation. The ability of ES936 to stimulate microsomal NAD(P)H oxidation was measured in purified rat liver microsomes. ES936 did stimulate low levels of NAD(P)H oxidation, but compared with other quinones (menadione, β-lapachone) under identical conditions, the rate of NAD(P)H oxidation with ES936 was very low (Fig. 5).
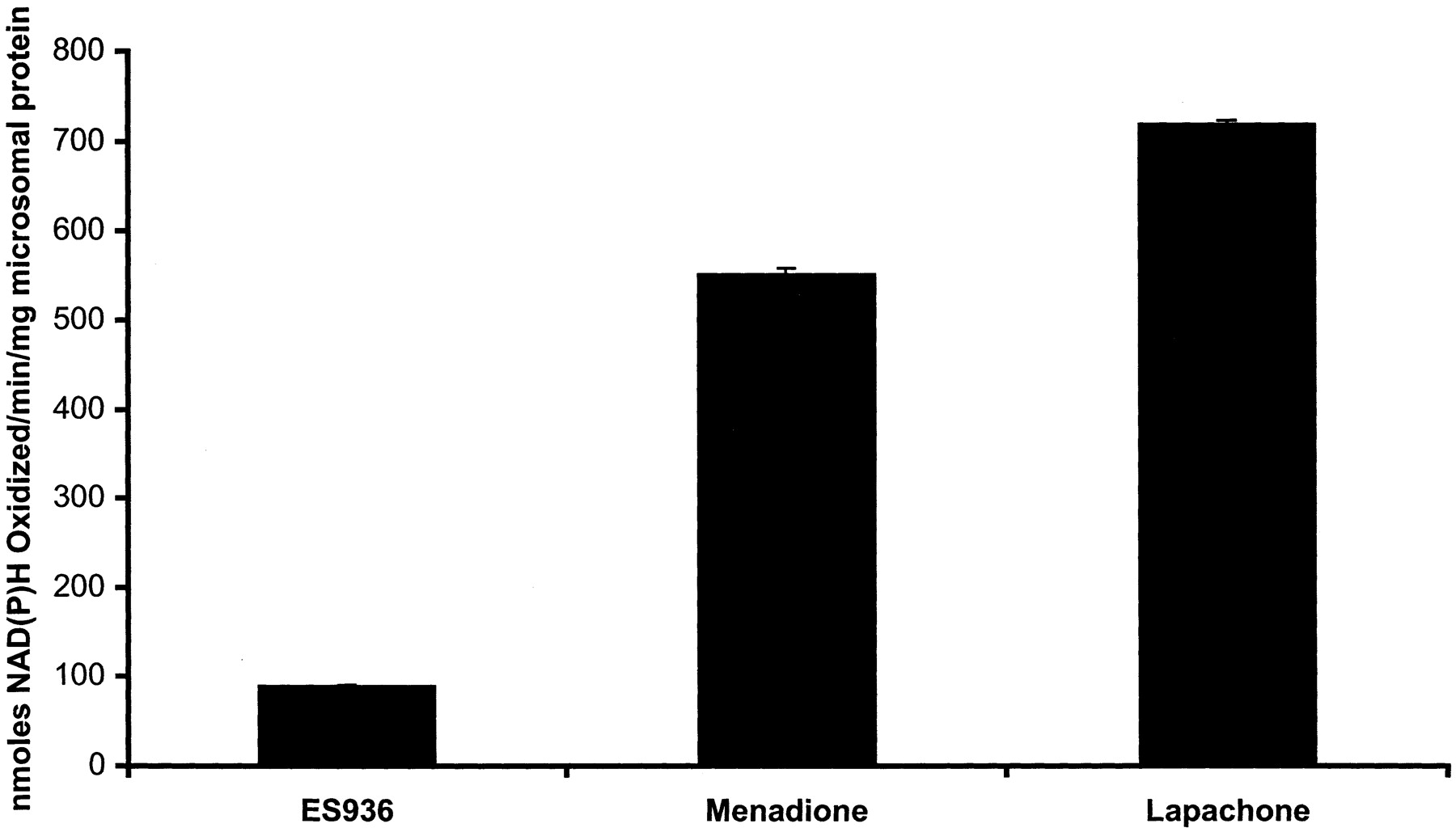
Quinones (ES936, menadione, and β-lapachone) were examined for their ability to stimulate rat liver microsomal-dependent NAD(P)H oxidation. Reaction conditions are described under Materials and Methods. Values represent the means ± S.E.M. of three independent experiments.
DNA Damage. We used the comet assay to investigate the potential for the induction of DNA strand breaks after ES936 exposure. A dose-dependent increase in the degree of DNA damage with increasing concentration of ES936 can be seen in a comparison of the overall comet scores (see Materials and Methods) for each treatment (Fig. 6A). For each ES936 concentration evaluated, the contribution of each degree of DNA damage is presented in Fig. 6B. In both cell lines studied (open and closed symbols in Fig. 6B), ES936 at a concentration of 100 nM (triangles) induced a similar number of comets in each class, and the level of damage was up to twice that seen in the controls. At doses of ES936 greater than 100 nM in the MDA468 NQ16 cells, more severely damaged nuclei are seen, as indicated by increased numbers of class 3 comets. In HCT116 cells, doses greater than 100 nM ES936 resulted in more comets formed in classes 1 and 2 and fewer in class 3, indicating less severe damage to the nuclei. This is reflected in the lower total comet score (Fig. 6A) for HCT116 cells compared with scores for MDA468 NQ16 cells at each increasing dose of ES936.

DNA strand breaks assessed by the comet assay in MDA468 NQ16 cells (open symbols) and HCT116 cells (filled symbols) after exposure to ES936 for 2 h. Total DNA damage (A) was determined after treating cells with varying concentrations of ES936 (as shown), visually sorting comets into one of four classes (0 through 3) depending on the percentage of DNA in the tail, determining the percentage of comets in each class, multiplying the percentage in a class by the number assigned to that class, and then summing the products to obtain a total score per treatment. The fold difference between treatment and control cells in each comet class for each concentration of ES936 (triangles represent 100 nM, squares represent 250 nM, and circles represent 500 nM) is shown in B. Results are the average of two independent experiments.
Discussion
ES936 is a mechanism-based inhibitor of NQO1 that rapidly inactivates more than 95% of NQO1 protein at concentrations as low as 25 nM. Concentrations of ES936 required for inhibition are roughly 1000-fold lower than concentrations of dicoumarol, the classic NQO1 inhibitor. ES936 inhibits NQO1 at concentrations well below those that induce growth-inhibitory effects, as indicated by IC50 values determined by the MTT assay. For consistency between cell lines used in this study, 100 nM ES936, the minimum concentration required to inhibit NQO1 in MDA468 NQ16 cells within 30 min of exposure, was used for all cell lines. After dilution of ES936 to a 100-nM concentration in complete media, effective inhibition of NQO1 activity (> 95% of control activity) could be observed in HCT116 cells up to 8 h after a 30-min incubation of cells in the treated media. However, complete inhibition of NQO1 activity in MDA468 NQ16 cells, which express high levels of NQO1, was only seen up to 2 h after the same 30-min incubation of cells in the treated media. Because HCT116 cells are inhibited by lower concentrations of ES936 relative to MDA468 NQ16 cells, this indicates some degradation of ES936 in complete media. However, unless cells have extremely high levels of NQO1 (such as stable transfectants), it is probable that 100 nM ES936 will inhibit cellular NQO1 efficiently for incubation periods of up to 8 h. Clearly, the duration of NQO1 inhibition by ES936 depends on the initial cellular content of NQO1, and the effectiveness of ES936 should be examined in each cell line in which it is to be used as an inhibitor of NQO1.
To measure of the effectiveness of ES936, we evaluated its ability to abrogate the toxicity of streptonigrin, an aminoquinone antitumor antibiotic efficiently bioactivated by NQO1 (Beall et al., 1996). Streptonigrin was chosen as a test compound because it showed the best correlation between NQO1 activity and toxicity of the more than 31,000 compounds tested in the National Cancer Institute's 60-tumor cell line panel (Paull et al., 1994). Streptonigrin is 6-fold less toxic to MDA468 NQ16 cells and approximately 2-fold less toxic to HCT116 cells when pretreated with 100 nM ES936. Although NQO1 is clearly inhibited by ES936, it does not inhibit other cellular reductases such as NAD(P)H cytochrome P450 reductase or NADH cytochrome b5 reductase. We have reported previously a similar result in the BE cell line (Winski et al., 1998). The activities of xanthine oxidase and xanthine dehydrogenase were not measured because they are consistently below the levels of detection in these cell lines (Beall et al., 1996; Winski et al., 1998).
Electrophilic and redox cycling reactions of quinones may lead to decreases in cellular thiol content. Levels of glutathione, the most abundant intracellular thiol (>99.5% of total glutathione is GSH) (Anderson, 1985) and an important detoxicant for free radicals, would be expected to decrease if ES936 were generating reactive oxygen species through one-electron reduction and redox cycling. The lack of change in acid-soluble thiol levels, regardless of the duration of exposure, indicates that ES936 is not generating substantial levels of reactive oxygen species. These data correlate with the relatively low degree of NAD(P)H oxidation detected in rat liver microsomes.
Some DNA damage may occur with the concentrations of ES936 (≤100 nM) used to inhibit NQO1 activity. A small increase in the DNA content of the comet tail after treatment with ES936 is consistent with the induction of single-strand breaks. To place the extent of DNA damage resulting from 100 nM ES936 exposure in perspective, exposure to streptonigrin at a dose 1000-fold lower results in a higher overall comet score (data not shown). The low amount of rat liver microsome-dependent NAD(P)H oxidation seen with ES936 treatment could relate to the DNA damage observed using the comet assay. No data are available to determine whether the degree of DNA damage observed at 100 nM ES936 might result in cellular toxicity, because DNA repair was not evaluated in this study. ES936, at all concentrations used in this study, had no effect on p53 status in HCT116 cells (wild-type p53), indicating that downstream markers of DNA damage were not activated (Anwar et al., 2003).
In summary, ES936 is a potent and specific inhibitor of NQO1 in cellular systems at concentrations of more than 1000-fold lower than the nonspecific inhibitor dicoumarol. The resumption of NQO1 catalytic activity depends on the synthesis of new protein, which is cell line-dependent, and therefore the duration of inactivation induced by ES936 varies between cell lines. The inhibition of other reductases by ES936 was not observed in two different cell lines. Using streptonigrin, a compound that is bioactivated by NQO1, we validated the use of ES936 as an inhibitor of NQO1 in cells. One caveat in the use of ES936 is the induction of a relatively small amount of DNA damage in the form of strand breaks. The degree of damage was relatively minor compared with that observed with other genotoxic quinones, such as streptonigrin, and was in agreement with a limited amount of redox cycling observed in rat liver microsomal systems. However, care should be taken if ES936 is used as an inhibitor of NQO1 in studies in which DNA damage is used as an endpoint. From the findings in our studies, we propose the use of ES936 (100 nM) as a mechanism-based inhibitor of NQO1 in cellular systems and for use as a component of the routine activity assay for NQO1.
Footnotes
-
This work was supported by National Institutes of Health grant R01-CA51210.
-
ABBREVIATIONS: NQO1, NAD(P)H:quinone oxidoreductase 1; ES936, 5-methoxy-1,2-dimethyl-3-[(4-nitrophenoxy)methyl]indole-4,7-dione; GSH, glutathione; DCPIP, 2,6-dichlorophenol-indophenol; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium.
- Received February 5, 2003.
- Accepted May 22, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}