


Abstract
An optimized assay for the binding of [3H]dimethyl-W84 to its allosteric site on M2 muscarinic receptors has been used to directly measure the affinities of allosteric ligands. Their potencies agree with those deduced indirectly by their modulation of the equilibrium binding and kinetics of [3H]N-methylscopolamine ([3H]NMS) binding to the orthosteric site. The affinities and cooperativities of orthosteric antagonists with [3H]dimethyl-W84 have also been quantitated. These affinities agree with those measured directly in a competition assay using [3H]NMS. All these data are compatible with the predictions of the allosteric ternary complex model. The association and dissociation kinetics of [3H]dimethyl-W84 are rapid but the estimate of its association rate constant is nevertheless comparable with that found for the orthosteric radioligand, [3H]NMS. This is unexpected, given that the allosteric site to which [3H]dimethyl-W84 binds is thought to be located on the external face of the receptor and above the [3H]NMS binding site that is buried within the transmembrane helices. The atypical allosteric ligands tacrine and 4,4′-bis-[(2,6-dichloro-benzyloxy-imino)-methyl]-1,1′-propane-1,3-diyl-bis-pyridinium dibromide (Duo3) inhibit [3H]dimethyl-W84 binding with the same potencies and comparably steep slope factors as found for inhibition of [3H]NMS binding. Tacrine and Duo3 decrease [3H]dimethyl-W84 affinity, not the number of binding sites. It is suggested that these atypical ligands either bind to the two known spatially separated allosteric sites on muscarinic receptors with positive cooperativity or their binding to the common allosteric site modulates receptor-receptor interactions such that homotropic positive cooperativity within a dimer or higher oligomer is generated.
For a growing number of G protein-coupled receptors (GPCRs), ligand binding and function have been shown to be susceptible to allosteric modulation (see, for example, Soudijn et al., 2001; Christopoulos and Kenakin, 2002). This phenomenon has been intensively studied on muscarinic acetylcholine receptors (Tuček and Proška, 1995; Birdsall et al., 1996; Ellis, 1997; Holzgrabe and Mohr, 1998). Many features of allosteric modulation have been demonstrated: enhancement of agonist binding (Birdsall et al., 1997; Jakubík et al., 1997; Lazareno et al., 1998; 1999) and action (Birdsall et al., 1999), absolute subtype selectivity of allosteric action (Birdsall et al., 1997; Lazareno et al., 1998, 1999), mapping of ligand receptor interactions (Matsui et al., 1996; Tränkle et al., 1998a), defined maximum effects on function (Lüllmann et al., 1969; Clark and Mitchelson, 1976), and modulation of receptor signaling efficacy (Zahn et al., 2002). All five muscarinic receptor subtypes are sensitive to allosteric modulation (Ellis et al., 1991) with many of the typical quaternary ammonium modulators such as gallamine, alcuronium, W84, and dimethyl-W84, exhibiting an M2 subtype selectivity. These ligands are thought to interact with a “common allosteric site” on the extracellular face of muscarinic receptors (Ellis and Seidenberg, 1992; Matsui et al., 1996; Tränkle and Mohr, 1997; Tränkle et al., 1998b), although recent mutagenesis studies have revealed that these modulators may use different amino acids for their receptor interaction (e.g., Ellis and Seidenberg, 2000; Buller et al., 2002). A second allosteric site on muscarinic receptors, which exhibits a different pharmacology, has also been described previously (Lazareno et al., 2000; 2002).
Until recently, it was necessary to use radiolabeled ligands for the orthosteric neurotransmitter binding site to investigate the binding of allosteric agents. However, the discovery and synthesis of the potent radiolabeled allosteric agent [3H]dimethyl-W84 has now allowed binding events at the common allosteric site to be monitored directly (Tränkle et al., 1998b; 1999). Using a centrifugation assay, the specific binding of [3H]dimethyl-W84 to both the unoccupied and N-methylscopolamine (NMS)-occupied M2 receptors could be measured and equilibrium competition assays used to quantify the interactions of allosteric ligands, as monitored from the viewpoint of the allosteric site. The equilibrium interactions of gallamine and NMS at M2 receptors were measured using radioligands for both the allosteric and orthosteric sites (Tränkle et al., 1999). The binding data were shown to be fully compatible with the predictions of the allosteric ternary complex model (Stockton et al., 1983; Ehlert, 1988; Lazareno and Birdsall, 1995), often used to described allosteric actions in GPCRs.
The centrifugation assay suffers from the limitation of a poor ratio of specific to nonspecific binding caused, in part, by the presence of a saturable nonspecific [3H]dimethyl-W84 component with an apparent Kd of ∼30 μM (Tränkle et al., 1998b). This binding component interferes with measurements of the affinity of allosteric agents of lower potency. Furthermore, the time needed for separating the radiolabeled membranes by centrifugation precludes attempts to measure the [3H]dimethyl-W84 binding kinetics.
The aim of the present study was to develop a more robust [3H]dimethyl-W84 binding assay and to use it to gain insight into allosteric phenomena that could not be studied previously. We investigated the interaction of a number of orthosteric and allosteric ligands with [3H]dimethyl-W84. These included ligands such as gallamine and strychnine, but also AF-DX 384, which has been considered to bind to both the orthosteric and allosteric sites (Tränkle et al., 1998a).
There is another category of muscarinic allosteric agents that have been termed `atypical' allosteric modulators. These modulators may be distinguished from other allosteric modulators by their different interactions with the binding of radioligands to the orthosteric binding site (Potter et al., 1989; Tränkle and Mohr, 1997; Schröter et al., 2000). In particular, their modulation of the binding of orthosteric ligands is characterized by slope factors greater than 1, which, at least in principle, is compatible with positive homotropic cooperativity. The effects of the `atypical' allosteric agents, Duo3 and tacrine (Fig. 1) on the equilibrium binding and dissociation of [3H]dimethyl-W84 from M2 receptors have been investigated. Preliminary reports of some of the findings are described elsewhere (Tränkle et al., 2000a,b; Weyand et al., 2001).

Structures of Duo3 and tacrine.
Materials and Methods
Cell Culture and Preparation of Membranes. Chinese hamster ovary cells (CHO cells) stably transfected with the human M2-receptor gene (generously provided by Dr. N. J. Buckley, University of Leeds, UK) were grown in a medium consisting of nutrient mixture/Ham's F12 medium, 10% fetal calf serum, 100 units/ml penicillin G, 0.1 mg/ml streptomycin, 1 mM l-glutamine, and 0.2 mg/ml of G418. The cells were incubated in a humidified atmosphere at 37°C and 5% CO2. Sixteen hours before cell harvesting (cell confluence of ∼80%), the growth medium was removed and replaced with fresh medium enriched with 5 mM sodium butyrate. The cells were harvested by scraping in a cold homogenisation buffer (20 mM HEPES and 10 mM Na4EDTA, pH 7.4, 4°C). The cells were homogenized twice at ”level 6” for 6 s using a Polytron homogenizer (PT 10–35; Kinematica AG, Basel, Switzerland) and the membranes were pelleted at 40,000g for 10 min at 4°C in an Avanti J25 centrifuge with a JA 25.50 rotor (Beckman Coulter Fullerton, CA). The pellets were washed twice in 20 mM HEPES and 0.1 mM Na4EDTA, pH 7.4, 4°C ('storage buffer'), and the final pellets were stored as a membrane suspension in storage buffer (ca. 1.4 mg/ml) at -80°C. Porcine heart membranes were prepared as described previously (Tränkle and Mohr, 1997) and stored under the same conditions as the CHO membranes. Protein content was determined by the Lowry method.
Binding Assays. To prevent nonspecific binding of the radioligand and nonradioactive ligands to negatively charged surfaces, all handling of [3H]dimethyl-W84 (specific activity, 154–168 Ci/mmol) was carried out in 20 mM NaCl/0.01% bovine serum albumin. The centrifugation assay was carried out as described previously (Tränkle et al., 1998b). To develop the filtration assay, we systematically optimized the magnitude of specific [3H]dimethyl-W84 binding and the specific/nonspecific binding ratio with respect to radioligand concentration, receptor density, buffer composition, assay volume, filter surface, filter pretreatment, composition, a volume of filter washing solution, and the harvester used for filtration. Major efforts were made to reduce nonspecific binding to the filters and adsorption losses of the radioligand onto plastic and glass. The optimized assay uses [3H]dimethyl-W84 (1.5 to 2.0 nM) and M2-containing membranes (100 μg of protein/ml) in a buffer consisting of 10 mM HEPES, 20 mM NaCl, and 0.01% bovine serum albumin, pH 7.4, at 23°C (HEPES buffer). [In the [3H]dimethyl-W84 binding centrifugation assay (Tränkle et al., 1998b), the buffer was 4 mM Na2HPO4, 1 mM KH2PO4, pH 7.4, at 23°C (Na/K/Pi buffer)]. Nonspecific [3H]dimethyl-W84 binding was determined in the presence of 10 μM gallamine.
[3H]NMS competition assays used 0.2 nM [3H]NMS and 50 μg of protein/ml of membranes. Nonspecific [3H]NMS binding was determined in the presence of 1 μM atropine and did not exceed 5% of total binding. Specific binding of [3H]N-methylscopolamine under control conditions was characterized by a log affinity constant (M-1), logKd, of 9.54 ± 0.05 (Kd, 0.3 nM) and Bmax 1010 ± 50 fmol/mg of protein (mean ± S.E.M., n = 25). All homologous and heterologous inhibition experiments with [3H]dimethyl-W84 and various allosteric modulators were carried out in a 0.3-ml volume in 1.2-ml well plates (Abgene House, Epsom, UK) for 2 h. To measure the interaction of [3H]dimethyl-W84 with orthosteric agents, the incubation time was extended to 5 to 6 h. The incubation time necessary to equilibrate [3H]NMS binding in the presence of an allosteric modulator was calculated according to the following equation:  where t0.5obs is an estimate of [3H]NMS association half-life time in the presence of allosteric modulator X, t0.5off is the half-life time of [3H]NMS dissociation in the absence of allosteric modulator, and Kx,diss is the affinity of the modulator for the [3H]NMS-occupied receptor, calculated from the reciprocal of the concentration of modulator at which the half-life time of [3H]NMS dissociation is doubled. Equilibrium was assumed to be reached after 5 × t0.5obs. The incubation time in competition experiments with [3H]NMS and various orthosteric agents was 2 h.
where t0.5obs is an estimate of [3H]NMS association half-life time in the presence of allosteric modulator X, t0.5off is the half-life time of [3H]NMS dissociation in the absence of allosteric modulator, and Kx,diss is the affinity of the modulator for the [3H]NMS-occupied receptor, calculated from the reciprocal of the concentration of modulator at which the half-life time of [3H]NMS dissociation is doubled. Equilibrium was assumed to be reached after 5 × t0.5obs. The incubation time in competition experiments with [3H]NMS and various orthosteric agents was 2 h.
For dissociation experiments, membranes were incubated with the respective radioligand for 60 min. Thereafter, aliquots of the mixture were added to excess unlabeled ligand in buffer over a total period of 120 min, followed by simultaneous filtration of all samples. To determine the effect of Duo3 and tacrine on the dissociation of [3H]dimethyl-W84, dissociation was measured by addition of 10 μM gallamine in combination with the respective allosteric agents. In the case of [3H]NMS dissociation experiments, rebinding of the radioligand was prevented by 1 μM atropine in combination with the respective allosteric agent.
Receptor bound radioactivity was separated by filtration on a Tomtech 96-well Mach III Harvester (PerkinElmer Wallac, Turku, Finland) using glass fiber filtermats (Filtermat A; PerkinElmer Wallac) which had been pretreated with 0.2% polyethylenimine. Filtration was followed by a single rapid washing step (0.8 ml of 100 mM NaCl, 4°C, 1.7 s). Filtermats were dried for 3 min at 400 W in a microwave oven. Thereafter, scintillation wax (Meltilex A; PerkinElmer) was melted for 1 min at 90°C onto the filtermat using a Dri-Block DB-2A (Techne, Duxford, Cambridge, UK). The filters were placed in sample bags (PerkinElmer Wallac), and filter bound radioactivity was measured using a Microbeta Trilux-1450 scintillation counter (PerkinElmer Wallac).
Data Analysis. The binding data from individual experiments were analyzed by computer-aided, nonlinear regression analysis using Prism 3.02 (GraphPad Software, San Diego, CA). Homologous competition data obtained with [3H]dimethyl-W84 and [3H]NMS were analyzed using a four-parameter logistic function to give estimates of the bottom and top plateaus, the logIC50, and the slope factor, nH, of the curve. If the observed slope factors did not differ significantly from unity (partial F-test, P > 0.05), the logIC50 values were estimated with n constrained to 1. The affinity constants (expressed as logKl) and Bmax values for the radioligand were calculated according to DeBlasi et al. (1989). The affinities of the orthosteric agents for inhibiting the binding of [3H]NMS, expressed as logKi,orthoster, were calculated from the logIC50 values by applying the Cheng-Prusoff correction. Similarly, the affinities of the allosteric ligands for inhibiting the binding of [3H]dimethyl-W84 were expressed as logKi,alloster values.
Homologous [3H]dimethyl-W84 inhibition curves obtained with the centrifugation procedure of Tränkle et al. (1998b) were fitted using a two-site model. The affinity constant for the high-affinity component of this fit was calculated according to DeBlasi et al. (1989).
Analyses of the data for the effect of orthosteric agents on the specific binding of the allosteric ligand [3H]dimethyl-W84 and of the allosteric agents on the specific binding of the orthosteric radioligand [3H]NMS were based on the allosteric ternary complex model (Stockton et al., 1983; Ehlert, 1988). We used eq. 2 from Lazareno and Birdsall (1995), in which Rt was replaced by eq. 8 of the same study and a slope factor, nH, was included:  BLX denotes the specific binding of the respective radioligand L (either [3H]dimethyl-W84 or [3H]NMS) in the presence of the cooperatively interacting agent X (orthosteric agent or allosteric agent, respectively). B0 is the binding of L in the absence of X. Note that in eq. 2, Kl and Kx are the affinity constants for the binding of L and X, respectively, at the unliganded receptors. α denotes the cooperativity factor for the allosteric interaction between X and L (with α > 1, α < 1, α = 1 indicating positive, negative, and neutral cooperativity, respectively). The factor nH represents the slope factor of the curve. Plotting logX as the independent variable and BLX as the dependent variable, a fit of the data with L and Kl as known constants yielded estimates for B0, Kx, and α. In cases in which the slope factors of the curves were significantly different from unity (F-test, P < 0.05), the mechanistic implications are unclear, and the data were fitted to a model-independent four-parameter logistic equation yielding the slope factor, nH, and logKx,alloster, the negative log concentration of X giving a 50% reduction of radioligand binding by the test compound.
BLX denotes the specific binding of the respective radioligand L (either [3H]dimethyl-W84 or [3H]NMS) in the presence of the cooperatively interacting agent X (orthosteric agent or allosteric agent, respectively). B0 is the binding of L in the absence of X. Note that in eq. 2, Kl and Kx are the affinity constants for the binding of L and X, respectively, at the unliganded receptors. α denotes the cooperativity factor for the allosteric interaction between X and L (with α > 1, α < 1, α = 1 indicating positive, negative, and neutral cooperativity, respectively). The factor nH represents the slope factor of the curve. Plotting logX as the independent variable and BLX as the dependent variable, a fit of the data with L and Kl as known constants yielded estimates for B0, Kx, and α. In cases in which the slope factors of the curves were significantly different from unity (F-test, P < 0.05), the mechanistic implications are unclear, and the data were fitted to a model-independent four-parameter logistic equation yielding the slope factor, nH, and logKx,alloster, the negative log concentration of X giving a 50% reduction of radioligand binding by the test compound.
[3H]NMS and [3H]dimethyl-W84 dissociation data were analyzed assuming a monoexponential decay. The slowing actions of the allosteric agents on [3H]NMS dissociation were expressed as the percentage reduction of the apparent rate constant for [3H]NMS dissociation. The concentration-effect curves for reduction of the [3H]NMS dissociation rate constant by allosteric agents were fitted to a fourparameter logistic function. The parameter ”top” was the value of k-1 measured in the absence of test compound and was fixed at 100%, whereas the log affinity of the allosteric ligand, log Kx,diss, and slope factor, nH, were set as variables. The parameter ”bottom” was checked whether it, as a variable, yielded a significantly better fit compared with it being fixed at 0%. This was not the case in all experiments (P > 0.05, partial F-test). Finally, we tested whether the slope factors of the curves were different from unity (partial F-test, P < 0.05 was taken as the level of statistical significance). The [3H]dimethyl-W84 association data were analyzed using a simple monoexponential equation with the t = 0 data point constrained to the value of the nonspecific binding.
Drugs. Dimethyl-W84 and the bis tertiary amine 2,2′-[1,6-hexanediylbis[(methylamino)-1,3-propanediyl]]bis[5-methyl-1H-isoindole-1,3-(2H)-dione], WDuo3, Duo3, and tacrine hydrochloride were synthesized and kindly provided by Dr. Holzgrabe and coworkers (Institute of Pharmacy and Food Chemistry, University of Würzburg, Würzburg, Germany). The synthesis of the radiolabeled compound [3H]dimethyl-W84 (specific activity, 154–168 Ci/mmol) was carried out by Amersham Biosciences (Braunschweig, Germany) using the method described by Tränkle et al., 1998b. [3H]NMS (specific activity, 70–82 Ci/mmol) was purchased from PerkinElmer Life Sciences (Homburg, Germany). Alcuronium dichloride and AF-DX 384 were generously provided by Hoffmann-La Roche AG (Grenzach-Wyhlen, Germany) and Dr. Karl Thomae GmbH (Biberach an der Riβ, Germany), respectively. Atropine sulfate, (-)-scopolamine hydrobromide, (-)-scopolamine N-methylbromide, (-)-scopolamine N-butylbromide, gallamine triethiodide, and strychnine hydrochloride were from Sigma Chemical (München, Germany). (+)-Dexetimide hydrochloride and (-)-levetimide hydrochloride were kindly provided by Janssen-Cilag AG (Neuss, Germany). W84 was provided by Dr. Joachim Pfeffer (University of Kiel, Germany). Ham's F12 medium, fetal calf serum, penicillin G, streptomycin, glutamine, G418 disulfate, trypsin-EDTA solution, and HEPES were purchased from Sigma-Aldrich Chemie (Steinheim, Germany). Sodium butyrate was from Acros Organics (Geel, Belgium).
Results
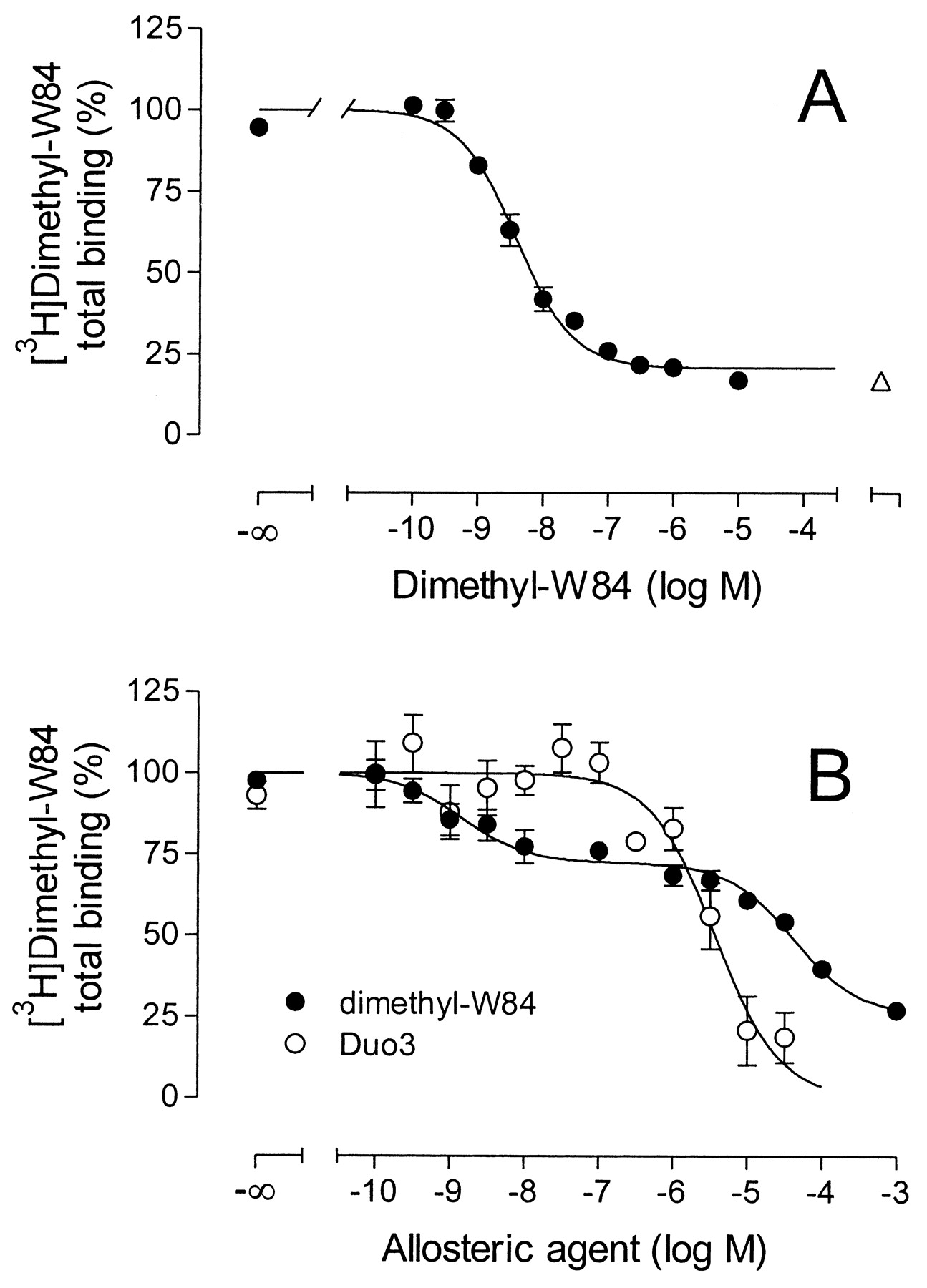
[3H]Dimethyl-W84 Binding Characteristics. Using the new filtration assay, the inhibition of [3H]dimethyl-W84 binding to M2 receptors, expressed on CHO cell membranes by unlabeled dimethyl-W84, followed a one-site inhibition curve (Fig. 2A). The ratio of saturable to nonsaturable binding was up to 4:1. High-affinity [3H]dimethyl-W84 binding was characterized by a logKl of 8.47 ± 0.04 (Kd, 3 nM; Table 1) and Bmax = 511 ± 45 fmol/mg of protein (means ± S.E.M, n = 22). For comparison, inhibition of [3H]dimethyl-W84 by unlabeled dimethyl-W84 at porcine heart M2 receptors using the centrifugation procedure is shown in Fig. 2B. This inhibition curve is biphasic as previously reported (Tränkle et al., 1998b; 1999), with the high- and low-affinity components of saturable [3H]dimethyl-W84 binding, each contributing 37% to the total binding; the remainder was nonsaturable binding. The parameters describing the high-affinity binding component, measured here in the absence of NMS (Fig. 2B), were logKl = 8.89 ± 0.18 (Kd, 1.3 nM) and Bmax = 105 ± 37 fmol/mg of protein (mean ± S.E.M, n = 4). Thus, the filtration assay gave nearly the same Kl value for the high-affinity component of dimethyl-W84 binding, but completely eliminated the saturable low-affinity binding component seen in the centrifugation assay. The low-affinity saturable component was also present in centrifugation assays using M2 CHO cell membranes (data not shown) and is therefore not a specific characteristic of porcine heart membranes.

Inhibition of [3H]dimethyl-W84 binding by increasing concentrations of the indicated test compounds using filtration (A) or centrifugation (B) to separate bound radioactivity. Binding was measured (A) in HEPES buffer using membranes from CHO cells stably expressing M2 receptors and (B) in Na,K,Pi-buffer with porcine cardiac membranes. Binding of [3H]dimethyl-W84 (A, 1.5 nM; B, 0.3 nM) in the absence of unlabeled modulator was set 100% (corresponding to 2100 cpm/0.3 ml and ∼13,500 dpm/1.5 ml, respectively). Error bars are shown when they exceed the size of the symbols. The open triangle in A indicates [3H]dimethyl-W84 binding in the presence of 10 μM gallamine. Illustrated are representative mean values ± S.E.M. from 22 experiments (A) with each data point measured in triplicate and (B) of six and three experiments carried out as quadruplicate determinations with dimethyl-W84 and Duo3, respectively.
Parameters characterizing the interactions of allosteric agents with the equilibrium binding of [3H]dimethyl-W84 and with the equilibrium binding and dissociation of [3H]NMS from M2 receptors
The data are mean values ± S.E.M. of 3 to 22 experiments carried out in triplicate. For details, see text.
Figure 2B also shows a [3H]dimethyl-W84-Duo3 inhibition curve using the centrifugation assay. According to a logistic curve fit, the foot of the curve was not statistically different from 0% and the slope factor not different from unity (F-test, P > 0.05). This illustrates how the saturable low-affinity component of [3H]dimethyl-W84 binding may confound the characterization of the effect of a nonradioactive allosteric ligand on the high-affinity component. Duo3 apparently interfered with both components of saturable [3H]dimethyl-W84 binding at similar concentrations, thus preventing characterization of the receptor interaction of this allosteric agent. It was also not possible to determine how Duo3 inhibited nonsaturable binding.
Inhibition of [3H]Dimethyl-W84 Binding by Various Allosteric Modulators. Specific [3H]dimethyl-W84 binding was inhibited by the allosteric agents gallamine, W84, WDuo3, alcuronium, and strychnine (Fig. 3). The slope factors were not different from unity (F-test, P > 0.05), and the maximum inhibition of total [3H]dimethyl-W84 binding at high concentrations of these ligands did not differ (data not shown). These data are compatible with a competitive interaction of the allosteric ligands with [3H]dimethyl-W84 at M2 receptors. The measured affinities are listed in Table 1 as logKi,alloster values.

Inhibition of specific [3H]dimethyl-W84 binding by increasing concentrations of typical allosteric modulators in membranes from CHO cells containing M2 receptors. Gallamine (10 μM) was included in each experiment to determine nonspecific [3H]dimethyl-W84 binding. Specific binding of [3H]dimethyl-W84 (1.5 nM) in the absence of inhibitor was set at 100%. Error bars are shown when they exceed the symbols. Curve fitting was based on a one-site model. Illustrated are mean values ± S.E.M. of three or four experiments carried out in triplicate.
Modulation of [3H]NMS Binding by Various Allosteric Ligands. If the interaction between [3H]dimethyl-W84 and the other agents takes place at the allosteric site of the M2 receptor, the ligands should have the allosteric effects on the binding of an orthosteric radioligand, such as [3H]NMS, that are predicted by the allosteric ternary complex model. This required testing in equilibrium and kinetic binding assays under the new assay conditions, because ionic strength and ionic composition of the medium are well known to have profound effects on the binding of allosteric ligands with multiple positive charges (Pedder et al., 1991; Tränkle et al., 1996). The results of the equilibrium and kinetic assays are shown in Fig. 4, A and B, respectively. Qualitatively, the equilibrium data were in line with the known actions of the allosteric agents under the different conditions: gallamine, dimethyl-W84, and W84 displayed a negative cooperativity with NMS, whereas WDuo3 and alcuronium had positively cooperative effects. The slopes of the curves did not deviate from unity (F-test, P > 0.05) and the parameters associated with the quantitative analysis of the cooperative interactions, logKx,alloster and α, are compiled in Table 1. According to the allosteric ternary complex model, the log affinity of a modulator for NMS-occupied M2 receptors is given by log(α × Kx,alloster). This value, obtained from the analysis of the equilibrium assay, can be compared with the potency of the allosteric ligands to slow down [3H]NMS dissociation, logKx,diss. If the allosteric ternary complex model is valid in this system, these two values should be the same.

Effects of allosteric ligands on the binding of [3H]NMS to M2 receptors in membranes from CHO cells. In the equilibrium studies (A), the binding of [3H]NMS (0.2 nM) is set to 100%. The curves show the fits of the data to the allosteric ternary complex model (for details, see Data Analysis), with dimethyl-W84 and alcuronium exhibiting positive cooperativity and the other ligands exhibiting negative cooperativity. In the dissociation experiments (B), the dissociation rate constant for [3H]NMS in the absence of allosteric ligands is set to 100%. All allosteric ligands fully inhibited [3H]NMS dissociation at high concentrations. The curves show the fits of the data to a simple inhibition curve. In both figures, the error bars are shown when they exceed the size of the symbols. Data are the mean values ± S.E.M. of three to six independent experiments performed in triplicate (equilibrium experiments) or duplicate (dissociation experiments).
In both the absence and the presence of the modulators, the [3H]NMS dissociation curves were monophasic. Under control conditions, the rate constant for [3H]NMS dissociation, k-1, was 0.21 ± 0.02 min-1 (mean ± S.E.M., n = 22; half-time, 3.3 min). A plot of the apparent rate constant of [3H]NMS dissociation versus increasing concentrations of the modulators yielded concentration-effect curves for all compounds that extrapolated to 0 (Fig. 4B). The affinities of the allosteric ligands for inhibiting the [3H]NMS dissociation rate constant, logKx,diss, are shown in Table 1 and, as expected, agree with the log(α × Kx,alloster) values. In addition, the affinities of the allosteric agents for the unliganded receptor, derived from their effects on [3H]NMS binding (logKx,alloster), are in good agreement with those values for competition with [3H]dimethyl-W84 binding (logKi,alloster).
Allosteric Interaction of Orthosteric Agents with the Binding of [3H]Dimethyl-W84. If [3H]dimethyl-W84 labels the allosteric site, it should undergo cooperative interactions with orthosteric antagonists and the derived affinity constants for the unliganded receptor should match those seen in competition experiments with [3H]NMS. The interaction of a series of antagonists with the binding of [3H]dimethyl-W84 (1.5 nM) is depicted in Fig. 5. All compounds concentration dependently decreased specific [3H]dimethyl-W84 binding. However, high concentrations of some orthosteric ligands did not inhibit 100% of the specific [3H]dimethyl-W84 binding. These plateau levels were a manifestation of a negative cooperative interaction of the antagonists with the radioligand. Whereas (+)-dexetimide and (-)-levetimide reduced specific [3H]dimethyl-W84 binding to zero at high concentrations, the other orthosteric agents showed a submaximum inhibitory action; the most pronounced effect was seen with atropine and NMS (Fig. 5). Curve-fitting using equations derived from the allosteric ternary complex model (Eq. 2) yielded estimates of logKx,orthoster, α, and the slope factor nH. Analysis of the steepness of the inhibition curves revealed only the slope for (-)-levetimide to be significantly different from unity. This may be because of the high concentrations of this compound used in the assay. Therefore, the data for (-)-levetimide were fitted to a four-parameter logistic equation with n = 1.30 ± 0.13 (Table 2). The logKx,orthoster values of (+)-dexetimide, NMS, scopolamine, atropine, AF-DX 384, and N-butylscopolamine were derived from curve fits with the slope factor constrained to 1 (Table 2).

Inhibition of specific [3H]dimethyl-W84 binding by increasing concentrations of orthosteric agents in membranes from CHO cells containing M2 receptors. Nonspecific [3H]dimethyl-W84 binding was defined in the presence of 10 μM gallamine. Specific binding of [3H]dimethyl-W84 (1.5 nM) in the absence of orthosteric agent was set 100%. Error bars are shown when they exceed the symbols. The curve fits were based on the ternary complex model of allosteric interactions applying Eq. 2 (see Materials and Methods); for (-)-levetimide, only logistic curve fitting was carried out; for details, see Data Analysis. The data are mean values ± S.E.M. of three to seven experiments carried out in triplicate.
Parameters characterizing the interaction of orthosteric ligands with the equilibrium binding of [3H]dimethyl-W84 and [3H]NMS at M2 receptors
The data are mean values ± S.E.M. of three to seven independent experiments. For details see text.
The binding constants of the orthosteric agents derived from competition experiments with [3H]NMS are also compiled in Table 2. As expected, the inhibition curves revealed slopes not significantly different from unity (data not shown). The logKi,orthoster values obtained with [3H]NMS (Table 2) matched the logKx,orthoster values obtained with [3H]dimethyl-W84, providing further evidence that the site of saturable [3H]dimethyl-W84 binding in the filtration assay is the allosteric site of the M2 receptor.
We also found very similar α values (0.19, 0.27) for the interaction of dimethyl-W84 and NMS, independent of whether [3H]dimethyl-W84 (Table 2) or [3H]NMS (Table 1) was the probe. Furthermore, the binding constants of dimethyl-W84 and NMS binding at free M2 receptors in Tables 1 and 2, derived from heterologous interaction (logKx,dimethyl-W84 = 8.55; logKx,nms = 9.40), corresponded to those derived from homologous competition (pKl,dimethyl-W84 = 8.47; pKl,nms = 9.37). These results support the ternary complex model of allosteric interactions.
Kinetics of [3H]Dimethyl-W84 Binding. Association of [3H]dimethyl-W84 (1.5 nM) to the M2 receptors was so rapid that the rising phase of the association time course could not be recorded with precision (Fig. 6A). Lowering the incubation temperature from 23° to 4°C did not retard the [3H]dimethyl-W84 association reaction to a measurable extent (data not shown). Both at 23° and 4°C, the scatter in the data resulted in there being no statistically significant difference between the [3H]dimethyl-W84 binding level at the first time point measurement (after 15 s) and the binding level of the remaining time points (t test, P > 0.05). Nevertheless, it was possible to fit the individual association curves at 23°C to an exponential curve with kobs of 0.14 ± 0.02 s-1 (half-time, 5 s, n = 2).

Kinetics of [3H]dimethyl-W84 binding to M2 receptors in membranes from CHO cells. A, association of [3H]dimethyl-W84. Ordinate, [3H]dimethyl-W84 binding at equilibrium as a percentage of nonspecific [3H]dimethyl-W84 binding determined in the presence of 10 μM gallamine (bottom dashed line). The top dashed line illustrates the mean level of [3H]dimethyl-W84 (1.5 nM) binding of all measurements. Data are mean values ± S.E.M. of two experiments carried out in duplicate. B, dissociation of [3H]dimethyl-W84. Ordinate, binding of [3H]dimethyl-W84 (1.5 nM) as a percentage of the binding level before the start of the dissociation reaction. Abscissa, time in minutes after addition of gallamine (10 μM) to measure the dissociation of [3H]dimethyl-W84. C, dissociation of [3H]dimethyl-W84 in the absence and presence of Duo3 and tacrine. The curves in B and C are the best-fit monoexponential curves. Data are mean values ± S.E.M. of two (B) and three to seven (C) experiments performed in duplicate.
Dissociation of [3H]dimethyl-W84 is illustrated in Fig. 6B. After incubation of the membranes with [3H]dimethyl-W84 for 30 min, dissociation was monitored by addition of 10 μM gallamine. [3H]Dimethyl-W84 binding rapidly decreased to the level of nonspecific binding. When a monoexponential curve was fitted to the data, a dissociation rate of 0.07 ± 0.01 s-1 (mean ± S.E.M., n = 3, t1/2,diss ≈ 9 s) was obtained. However, because the first accessible time point was measured at t = 15 s, this estimate of k-1,dimethyl-W84 is only an approximation. The use of 20 μM alcuronium in the dissociation experiment instead of 10 μM gallamine yielded essentially identical results (k-1,dimethyl-W84 = 0.09 ± 0.02 sec-1, mean ± S.E.M. n = 3, t1/2,diss ≈ 8 s). Lowering the incubation temperature from 23° to 4°C did not retard the [3H]dimethyl-W84 dissociation reaction to a measurable extent (data not shown). In addition [3H]dimethyl-W84 dissociation was not measurably affected in the presence of both 1 μM atropine and 10 μM gallamine (data not shown). Taken together, these data demonstrate that [3H]dimethyl-W84 has very fast binding kinetics.
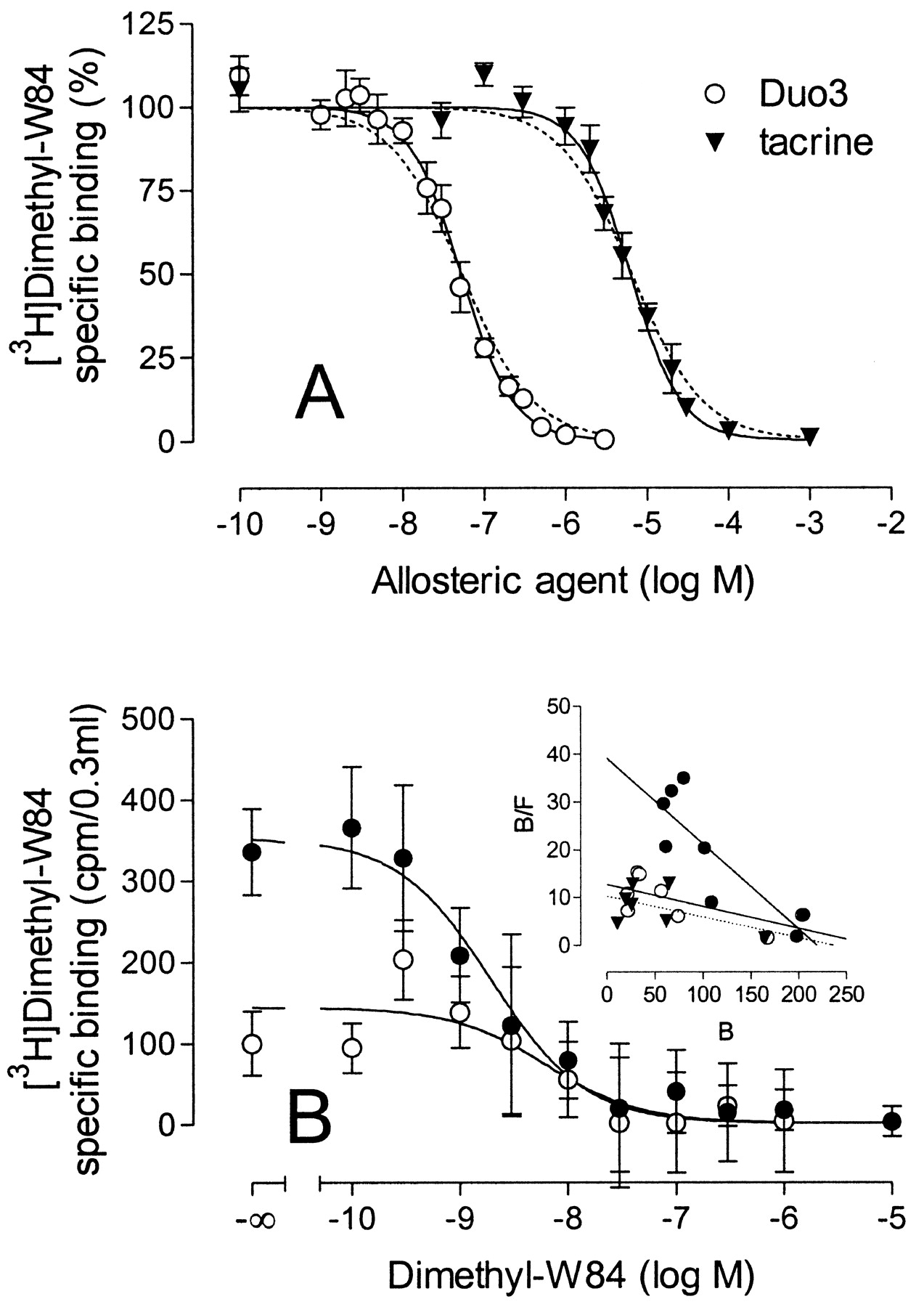
Effect of the Atypical Allosteric Modulators Duo3 and Tacrine on [3H]NMS Binding and Dissociation. The allosteric actions of Duo3 and tacrine on orthosteric ligand binding were present under the incubation conditions of the filtration assay. The inhibition of [3H]NMS binding by Duo3 and tacrine is illustrated in Fig. 7, open symbols. The incubation times in these experiments were extended to 15 h to avoid kinetic artifacts causing the extent of inhibition of the binding of [3H]NMS at high concentrations of the allosteric ligands to be overestimated. The inhibition curves reached a plateau at levels of binding above nonspecific binding. This plateau effect, one of the characteristic features of low negatively cooperative allosteric actions, was accentuated when the binding assay was carried out in the Na/K/Pi buffer (data not shown). It was possible to fit the inhibition curves to an allosteric model with the introduction of a slope factor (Table 3). The slope factors for Duo3 and tacrine were significantly larger than unity (F-test, P < 0.05), and the estimates of the cooperativity of Duo3 and tacrine with NMS were both ∼0.1.
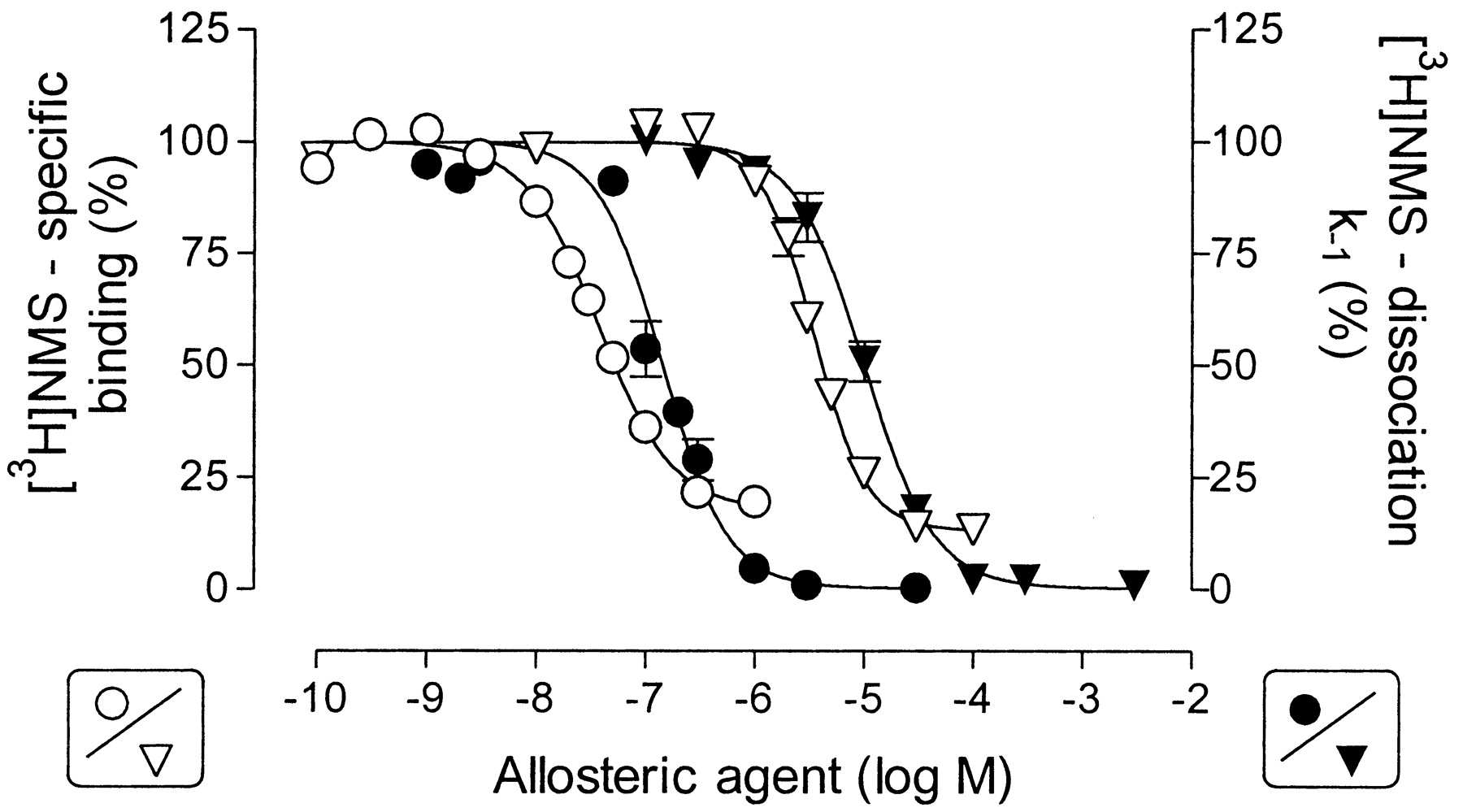
Effect of the atypical modulators, Duo3 (circles) and tacrine (triangles), on [3H]NMS equilibrium binding (open symbols) and on [3H]NMS dissociation (filled symbols) from M2 receptors in membranes from CHO cells. Left ordinate, specific binding of [3H]NMS (0.2 nM) as a percentage of the control value in the absence of allosteric agent. Right ordinate, apparent rate constant of [3H]NMS dissociation as a percentage of the control value in the absence of allosteric agent. The lines were obtained from curve fitting, applying a four-parameter logistic equation (for details see Data Analysis). Error bars are shown when they exceed the symbols. Data are mean values ± S.E.M. of five to seven inhibition experiments carried out in triplicate and of three to four dissociation experiments performed in duplicate.
Parameters characterizing the interactions of Duo3 and tacrine with the equilibrium binding of [3H]dimethyl-W84 and with the equilibrium binding and dissociation of [3H]NMS from M2 receptors
The data are mean values ± S.E.M. of four to seven experiments. Equilibrium and dissociation binding experiments were carried out with triplicate and duplicate replicates, respectively.
Duo3 and tacrine also reduced the rate constant for [3H]NMS dissociation from M2 receptors (Fig. 7, closed symbols). This occurred at 4- to 6-fold higher concentrations than those at which these ligands inhibit [3H]NMS equilibrium binding. The curves for the inhibition of the [3H]NMS dissociation rate constant, obtained by logistic curve fitting, extrapolate to 0% at high concentrations of the allosteric ligands and the slope factors of the curves were significantly greater than unity (F-test, P < 0.05). The estimates of potency of tacrine and Duo3 at the NMS-occupied receptor, obtained from the equilibrium studies, agreed reasonably well with the values estimated from the [3H]NMS dissociation assays (Table 3).
Effects of Duo3 and Tacrine on [3H]Dimethyl-W84 Binding. We examined how Duo3 and tacrine modulated the binding of [3H]dimethyl-W84. As depicted in Fig. 8A, both Duo3 and tacrine concentration-dependently inhibited the binding of [3H]dimethyl-W84 down to levels not significantly different from those of nonspecific binding. The slope factors of the inhibition curves for both Duo3 and tacrine were significantly greater than unity (F-test, P < 0.05) (Table 3). Increasing the concentration of [3H]dimethyl-W84 from 1.5 to 10 nM did not change the steep slopes and also had no detectable effect on the ability of these ligands to inhibit 100% of the specific binding (data not shown).
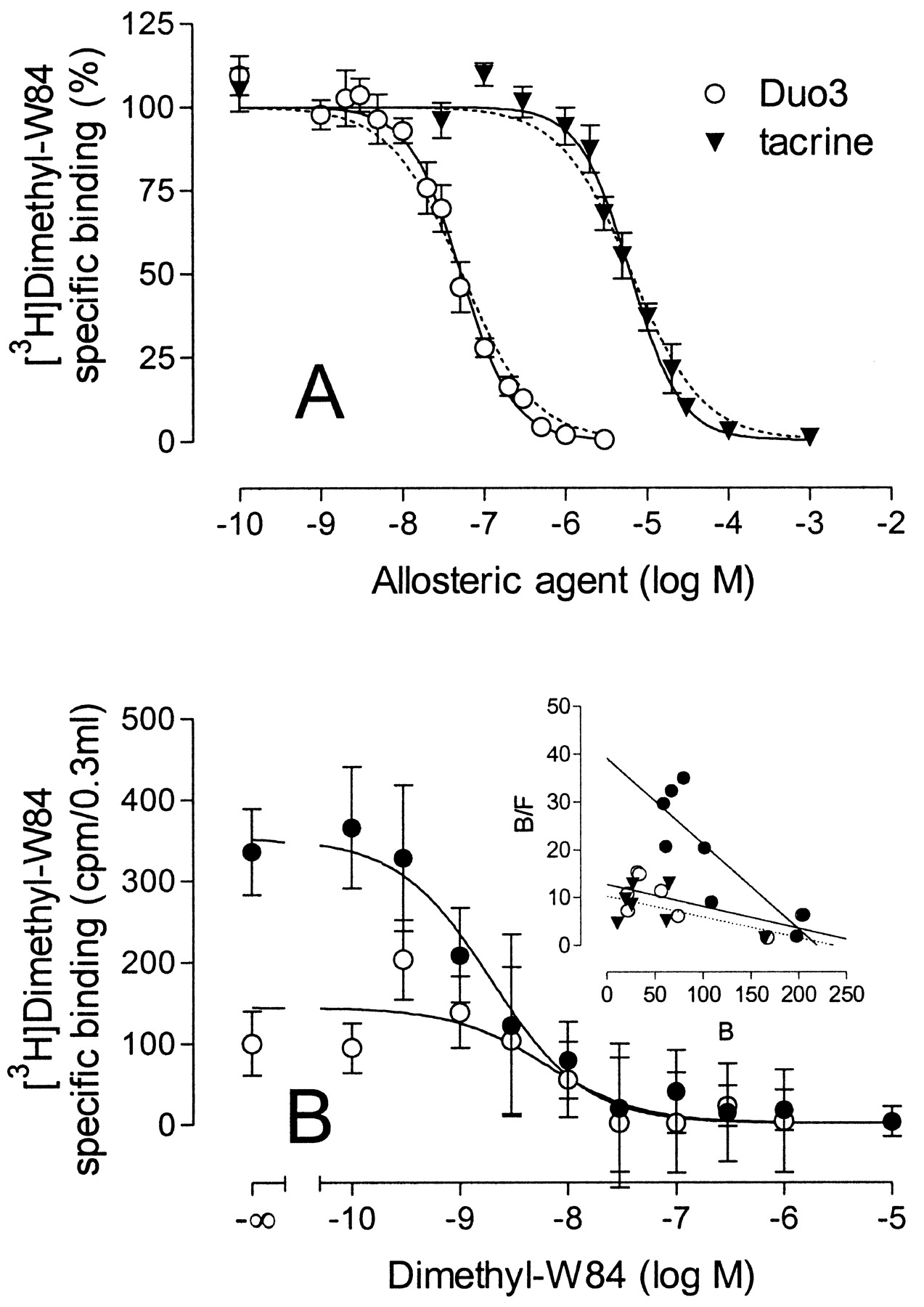
Effect of the atypical modulators Duo3 (○) and tacrine (▾) on [3H]dimethyl-W84 binding to M2 receptors in membranes of CHO cells. Nonspecific [3H]dimethyl-W84 binding was defined by the binding in the presence of 10 μM gallamine. A, inhibition of specific [3H]dimethyl-W84 (1.5 nM) binding by Duo3 and tacrine. Ordinate, specific binding of [3H]dimethyl-W84 as a percentage of the binding level in the absence of allosteric agent. 100% represents means ± S.E.M. and are 1091 ± 100 cpm (Duo3 experiments) and 1084 ± 143 cpm (tacrine experiments). Curve fitting was based on a four-parameter logistic equation with a variable slope factor (solid line) or a slope factor fixed to unity (dashed line); for details, see Data Analysis. Illustrated are mean values ± S.E.M. of seven experiments carried out in triplicate. B, homologous competition of specific [3H]dimethyl-W84 binding in the absence (•) and presence (○) of 0.1 μM Duo3. Ordinate, [3H]dimethyl-W84 specific binding (cpm/0.3 ml). The curves are the best fits to a logistic function. Data are means ± S.E.M. of one representative experiment of three performed in quadruplicate. Inset, Scatchard plots of these data and data for an analogous experiment using tacrine. Ordinate, ratio of specifically bound ligand over free ligand (femtomoles per milligram of protein per nanomolar). Abscissa, specifically bound ligand (femtomoles per milligram of protein). Both Duo3 (○, 0.1 μM) and tacrine (▾, 10 μM) reduced the affinity of [3H]dimethyl-W84 for M2 receptors relative to the control (•), but the numbers of binding sites (Bmax) were not significantly changed.
To gain more insight into the mechanism of action of Duo3 and tacrine, we attempted to discover whether the reduction of [3H]dimethyl-W84 binding results from a decrease in the affinity or the number of binding sites. Figure 8B illustrates the effect of an IC75 concentration of Duo3 on the homologous dimethyl-W84/[3H]dimethyl-W84 inhibition curve. The Scatchard plot, shown in the inset of Fig. 8B, illustrates that 0.1 μM Duo3 reduced the affinity of [3H]dimethyl-W84 by approximately 4-fold without significantly changing the number of binding sites (t test, two-tailed, P > 0.05). The inset also includes the similar result of an analogous experiment with tacrine (10 μM).
Because an alteration of radioligand dissociation by a test ligand unambiguously demonstrates allosteric behavior, we investigated whether Duo3 and tacrine at a single high concentration would decrease the dissociation of [3H]dimethyl-W84. Figure 6C illustrates that neither 30 μM Duo3 nor 300 μM tacrine induced a measurable change in the dissociation of [3H]dimethyl-W84.
Discussion
The allosteric ternary complex model is the simplest model that can be used to describe the binding of two ligands to separate but conformationally linked sites on a receptor. It underpins much of the analysis of the binding properties of receptors when allosteric ligands or allosteric interactions are involved. In general, radioligands are only available for the orthosteric site on G protein-coupled receptors; hence, the binding and actions of ligands at the allosteric site can only be inferred from their effects on the kinetics and equilibrium binding of the orthosteric radioligand. In particular, it is possible to check quantitatively whether the concentration dependence of the effect of the allosteric ligand on the dissociation kinetics of the radioligand are in accord with the equilibrium effects of the allosteric ligand on the binding of the radioligand (Lazareno and Birdsall, 1995). In addition, it has been possible to investigate interactions between allosteric ligands by their effects on the binding and kinetics of an orthosteric radioligand (Ellis and Seidenberg, 1992; 2000; Waelbroeck, 1994; Tränkle and Mohr, 1997; Lazareno et al., 2000, 2002). For most studies to date, the ternary complex model has provided a quantitative explanation of the allosteric interactions as monitored from the orthosteric site.
At present, [3H]dimethyl-W84 is the only radioligand available that binds to an allosteric site on muscarinic receptors. A centrifugation-based assay was used to demonstrate that there was a 1:1 stoichiometry of [3H]dimethyl-W84 and [3H]NMS binding sites on M2 muscarinic receptors, and the binding affinity of [3H]dimethyl-W84, in the presence of NMS, was the same as that measured from the effects of dimethyl-W84 on the equilibrium binding and kinetics of [3H]NMS (Tränkle et al., 1998b). It was also possible to demonstrate that the affinity of gallamine and its cooperativity with NMS was the same whether the binding was measured using [3H]NMS or [3H]dimethyl-W84, all predictions of the allosteric ternary complex model (Tränkle et al., 1999).
However, this assay had a poor ratio of specific to nonspecific binding (∼0.3:1), and there was also a low-affinity, high-capacity saturable nonreceptor binding component (Fig. 2B). This meant that it was not possible to obtain detailed information regarding the measurement of cooperativities between [3H]dimethyl-W84 and orthosteric ligands, the affinities of low affinity allosteric ligands, whether the allosteric ligands might be interacting cooperatively with [3H]dimethyl-W84, the kinetics of [3H]dimethyl-W84 binding, and how they might be affected by orthosteric and other allosteric ligands.
A [3H]dimethyl-W84 filtration assay, optimized for a high ratio of specific/nonspecific binding to M2 receptors was developed. The low-affinity binding component was absent (Fig. 2A), possibly because it was washed away during the brief ice-cold wash of the filters. Although the buffer conditions were changed, the measured log affinity constant for [3H]dimethyl-W84 (8.47) was near that found in the centrifugation assay (8.89; Tränkle et al., 1999).
The measured kinetics of [3H]dimethyl-W84 were fast (Fig. 6, A and B) and, remarkably, were still fast even at 4°C. The estimate of the dissociation half-time was 9 s (k-1 = 0.08 s-1) and is the first direct estimate of the kinetics of an allosteric muscarinic ligand. Previous indirect measurements had only suggested that, whether the orthosteric site was occupied or not, the allosteric ligand equilibrated very rapidly (Tränkle et al., 1997). Therefore, it is unsurprising that the association was essentially complete at the first time point (15 s; Fig. 6A). The association rate constant (k+1) of [3H]dimethyl-W84, predicted from its affinity constant (3 × 108 M-1) and dissociation rate constant (0.08 sec-1), assuming simple association kinetics, is 2 × 107 M-1s-1. At the concentration of [3H]dimethyl-W84 used in the assays, 1.5 nM, the predicted half-time for association is ∼6 s, which is near the value of 5 s estimated from the data in Fig. 6A. The data are therefore compatible with a simple bimolecular association of [3H]dimethyl-W84 with the M2 receptor.
An analogous calculation for [3H]NMS gives a k+1 of 1.2 × 107 M-1s-1. Interestingly, the association rate constants for the two radioligands are comparable and 'typical' for antagonist-receptor interactions in which the ligand is not severely conformationally constrained. This similarity might not be expected, given that the 'common allosteric site', to which ligands such as dimethyl-W84 and gallamine bind, is considered to be on the extracellular face of the receptor (Ellis et al., 1993; Leppik et al., 1994; Tuček and Proška, 1995; Matsui et al., 1996; Ellis and Seidenberg, 2000; Buller et al., 2002), whereas the NMS binding site is viewed as less accessible, being buried within the transmembrane helices of the receptor (Lu et al., 2002).
The Bmax estimates for [3H]dimethyl-W84 were approximately 50% of the Bmax for [3H]NMS, which contrasts with the anticipated 1:1 ratio observed in the centrifugation assay (Tränkle et al., 1998b). This may be a result, in part, of loss of specific [3H]dimethyl-W84 binding during the washing step, but there are also difficulties in precisely estimating Bmax values from homologous inhibition competition assays. There is, however, no evidence for the ratio of the number of common allosteric sites per orthosteric site being greater than 1.
It was possible to demonstrate that several allosteric ligands could inhibit completely the specific binding of [3H]dimethyl-W84 with slope factors not significantly different from 1 (Table 1). Their log affinities for the unliganded receptor agreed with those estimated from [3H]NMS-allosteric ligand equilibrium experiments (mean difference, 0.05 ± 0.17, n = 5), as predicted from the allosteric ternary complex model. Similarly, the estimates of the log affinities of the allosteric ligands for the NMS-occupied receptor obtained from the equilibrium and kinetic studies using [3H]NMS agreed with each other (mean difference 0.15 ± 0.13, n = 5, Table 1).
A number of orthosteric ligands inhibited the binding of [3H]dimethyl-W84 submaximally (Fig. 5, Table 2), providing estimates of the affinity of the orthosteric ligands for the unliganded receptor and the cooperativity between [3H]dimethyl-W84 and these ligands. The well known very large (10,000-fold) stereoselectivity of dexetimide and levetimide was evident, as was the relatively low negative cooperativity between dimethyl-W84 and both NMS and atropine. The mean difference between the estimates of the log affinities of the orthosteric ligands for the unliganded receptor (0.03 ± 0.06, n = 6, Table 2) was not significantly different from 0, again as predicted by the allosteric ternary complex model. The two estimates of the cooperativity between NMS and dimethyl-W84 using the two radioligands, 0.19 and 0.27, were also similar. Negative cooperativities of this magnitude mean that it should be possible to discover an orthosteric ligand that enhances the binding of [3H]dimethyl-W84. This positive cooperativity should increase the precision of the [3H]dimethyl-W84 binding assay and possibly enable the dissociation kinetics of the orthosteric ligand-[3H]dimethyl-W84-receptor complex to be measured.
AF-DX 384 is thought not to be a 'pure' orthosteric ligand because it interferes with binding to both the orthosteric and allosteric sites of the M2 receptor (Tränkle et al., 1998a). In the current study, we found that AF-DX 384 did not fully inhibit the binding of [3H]dimethyl-W84 (Fig. 5), suggesting an allosteric mechanism in which both ligands can bind simultaneously to the M2 receptor and interact with each other allosterically. If AF-DX 384 were purely allosteric, as proposed for the analog AF-DX 116 (Lanzafame et al., 2001), the interaction between [3H]dimethyl-W84 and AF-DX 384 should have been competitive.
Thus far, all the evidence discussed has been in agreement with the predictions of the allosteric ternary complex model. However, tacrine and Duo3 have been reported to inhibit the specific binding of [3H]NMS with slope factors greater than 1 (Table 3, Fig. 7) (Potter et al., 1989; Schröter et al., 2000). This might suggest simultaneous and positively cooperative binding of these ligands to both the orthosteric and allosteric sites. However, tacrine and Duo3 also slow down the dissociation rate of [3H]NMS with slope factors greater than 1 (Table 3, Fig. 7) (Potter et al., 1989; Tränkle and Mohr, 1997; Schröter et al., 2000). In this experiment, the orthosteric site is occupied by [3H]NMS; therefore, the effects causing the slope factors to be greater than 1 are allosteric in nature and independent of occupancy of the orthosteric site. The potencies of tacrine or Duo3 to inhibit the dissociation of [3H]NMS, relative to their inhibitory potencies in equilibrium binding assays (Fig. 7), predicts a 3- to 4-fold negative cooperativity with NMS at M2 receptors and that high concentrations of tacrine and Duo3 should not produce the 100% inhibition of [3H]NMS binding. This is observed for the first time for these compounds because of the incubation times were extended sufficiently to avoid kinetic artifacts (Fig. 7). These results are in agreement with data for a series of allosteric pentacyclic carbazolones that have positive, neutral, and negative cooperativity with NMS at M1-M4 receptors (Gharagozloo et al., 2002). Like tacrine and Duo3, they have slope factors greater than 1 in equilibrium and kinetic assays and also behave as predicted according to the equations for the allosteric ternary complex model with a slope factor inserted (Gharagozloo et al., 2002).
Using [3H]dimethyl-W84 to label the allosteric site, we also show that the Duo3 and tacrine inhibition curves have slope factors greater than unity, with potencies very similar to those found for the [3H]NMS inhibition curves (Table 3, Fig. 8). The inhibitory effect of Duo3 and tacrine is to decrease the affinity of [3H]dimethyl-W84 without affecting the Bmax. Therefore [3H]dimethyl-W84, binding to the common allosteric site, is sensing the same effects of Duo3 and tacrine as [3H]NMS binding to, or dissociating from, the orthosteric site. We could not detect an effect of tacrine or Duo3 on the dissociation rate of [3H]dimethyl-W84, which would be compatible with their having an action at another allosteric site.
We are left with the question of what molecular event generates the positive homotropic cooperativity. Given the fact that two allosteric sites on muscarinic receptors have now been described (Lazareno et al., 2000; 2002) with allosteric ligands, such as gallamine and KT5720, interacting with neutral cooperativity, it is possible that Duo3 (or tacrine) can bind to both these sites but with positive cooperativity. A further possibility, as suggested previously for tacrine (Potter et al., 1989) and the pentacyclic carbazolone analogs (Gharagozloo et al., 2002), is that muscarinic receptors exist as dimers or as a monomer-dimer equilibrium. Duo3 and tacrine would bind to only a single allosteric site on the receptor, but its binding would change the conformation of putative receptor-receptor interaction surfaces such that a positively cooperative interaction would be generated between the tacrine/Duo3 sites on a dimer or higher oligomer; communication between the receptors is being mediated via ligand binding. The receptor residues that are important for the binding of these ligands and the generation of their steep slope factors are being investigated.
Acknowledgments
We thank Iris Jusen and Angela Popham for their expert technical assistance. [3H]Dimethyl-W84 was developed in collaboration with Dr. Ulrike Holzgrabe, University of Würzburg, Germany.
Footnotes
-
This work was supported by the Deutsche Forschungsgemeinschaft and the Medical Research Council. U.V. is a member of the Deutsche Forschungsgemeinschaft Graduiertenkolleg 677.
-
ABBREVIATIONS: GPCR, G protein-coupled receptor; M2 receptor, M2 subtype of muscarinic acetylcholine receptor; W84, hexane-1,6-bis(dimethyl-3′-phthalimidopropyl-ammonium bromide); dimethyl-W84, N,N′-bis[3-(1,3-dihydro-1,3-dioxo-5-methyl-2H-isoindol-2-yl)propyl]-N,N,N′,N′-tetramethyl-1,6-hexanediaminium diiodide; Duo3, 4,4′-bis-[(2,6-dichloro-benzyloxy-imino)-methyl]-1,1′-propane-1,3-diyl-bis-pyridinium dibromide; NMS, (-)-N-methylscopolamine; CHO, Chinese hamster ovary; WDuo3, 1,3-bis[4-(phthalimidomethoxyimino-methyl)-pyridinium-1-yl] propane dibromide; AF-DX 384, 5,11-dihydro-11-{[(2-{2-[(dipropylamino)methyl]-1-piperidinyl}ethyl)amino]-carbonyl}-6H-pyrido(2,3-b)(1,4)benzodiazepine-6-one; AF-DX 116, 11-(2-((diethyl-amino)methyl)-1-piperidinylacetyl)-5,11-dihydro-6H-pyrido(2,3-b)-(1,4)-benzodiazepine-6-one.
- Received January 24, 2003.
- Accepted April 8, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}