


Abstract
Mercuric ions are highly reactive and form a variety of organic complexes or conjugates in vivo. The renal proximal tubule is a primary target for mercury uptake and toxicity, and circumstantial evidence implicates organic anion transporters in these processes. To test this hypothesis directly, the transport and toxicity of mercuric-thiol conjugates were characterized in a Madin-Darby canine kidney cell line stably transfected with the human organic anion transporter 1 (hOAT1). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-terazolium bromide assays (for mitochondrial dehydrogenase) confirmed that mercuric conjugates of the thiols N-acetylcysteine (NAC), cysteine, or glutathione were more toxic in hOAT1-transfected cells than in the nontransfected cells. The NAC-Hg2+ conjugate was most cytotoxic, inducing greater than 50% cellular death over 18 h at a concentration of 100 μM. The cytotoxic effects were fully reversed by probenecid (an OAT1 inhibitor) and partially reversed byp-aminohippurate (an OAT1 substrate). Toxicity of this conjugate was reduced by the OAT1-exchangeable dicarboxylates α-ketoglutarate, glutarate, and adipate, but not by succinate, a nonexchangeable dicarboxylate. 203Hg-uptake studies showed probenecid-sensitive uptake of mercury-thiol conjugates in the hOAT1-transfected cells. The apparent K m for the NAC-Hg2+ conjugate was 44 ± 9 μM. Uptake of the NAC-Hg2+ conjugate was cis-inhibited by glutarate, but not by methylsuccinate, paralleling their effects on toxicity. Probenecid-sensitive transport of the NAC-Hg2+conjugate was also shown to occur in Xenopus laevisoocytes expressing the hOAT1 or the rOAT3 transporters, suggesting that OAT3 may also transport thiol-Hg2+ conjugates. Thus, renal accumulation and toxicity of thiol-Hg2+ conjugates may depend in part on the activity of the organic transport system.
The renal organic anion transport system is an elimination route for many xenobiotics, as well as for endogenous organic anions (Burckhardt et al., 2001). The basolateral uphill step for organic anion transport is mediated by transporters of the OAT family. OAT1 is a dicarboxylate/organic anion exchanger that takes up organic anions in exchange for intracellular α-ketoglutarate (Pritchard, 1995), and its gene was cloned recently for the rat (Sekine et al., 1997; Sweet et al., 1997), winter flounder (Wolff et al., 1997), and human (Reid et al., 1998; Cihlar et al., 1999; Hosoyamada et al., 1999; Lu et al., 1999). This transporter is multispecific and mediates the uptake of structurally dissimilar organic acids and some neutral compounds, including p-aminohippurate (PAH), nonsteroidal anti-inflammatory drugs (Apiwattanakul et al., 1999), β-lactam antibiotics (Jariyawat et al., 1999), cyclic nucleotides (Sekine et al., 1997), and nucleoside analogs (Cihlar et al., 1999). The other basolateral organic anion transporter, OAT3, also handles a diverse number organic anion substrates by a yet undescribed mechanism, and its gene has been cloned in the rat (Kusuhara et al., 1999), human (Cha et al., 2001), and the mouse (Brady et al., 1999), in which its role was characterized recently with the use of knockout mice (Sweet et al., 2002). Apical efflux of organic anions into the lumen occurs by ATP-binding cassette transport efflux pumps as well as by a yet uncharacterized potential-sensitive transporter (Krick et al., 2000;Burckhardt et al., 2001).
It is well established that inorganic mercury (Hg2+) is nephrotoxic (Zalups, 2000). Accumulation and uptake of Hg2+ is high in all three segments of the proximal tubule (Zalups and Barfuss, 1990;Zalups, 1991a,b). All of the epithelial cells in these segments contain the OAT1 and OAT3 transporters in their basolateral membrane (Hosoyamada et al., 1999; Sweet et al., 1999; Tojo et al., 1999; Cha et al., 2001; Hasegawa et al., 2002). Recent in vivo data from rats indicates that OAT1 and/or OAT3 provides a route for basolateral uptake of mercuric species. For example, intravenous coadministration of HgCl2 with the organic anion transporter substrate PAH inhibits Hg2+ uptake when Hg2+ was administered as HgCl2 or as thiol-Hg2+conjugates (Zalups, 1998a,b). In another study using the same technique, it was found that pretreatment with small aliphatic dicarboxylic acids (i.e., glutarate and adipate) before Hg2+ administration inhibited renal Hg2+ uptake in a dose-dependent manner (Zalups and Barfuss, 1998b). Because these acids are all exchangeable substrates of OAT1, it was concluded that there was a basolateral uptake pathway for mercuric ion species involving the organic anion transporter (Zalups, 2000; Zalups and Koropatnick, 2000). Recently, PAH and glutarate-sensitive basolateral uptake of thiol-Hg2+ conjugates was shown to occur in rabbit isolated perfused proximal (pars recta) tubules (Zalups and Barfuss, 2002), which further supports the potential role of basolateral organic anion transporters.
Because OAT1 and OAT3 are located in the basolateral membrane, they can act upon mercuric species present in the blood. More than 98% of the mercuric ions in the serum are bound to albumin. A smaller fraction is presumed to be bound to endogenous, low-molecular-weight thiols (Lau and Sarkar, 1979). However, the mercuric-albumin complex seems to be labile because the in vivo concentration of mercuric species in plasma decreases rapidly over time (Zalups, 1998b), suggesting that ligand exchange occurs between a mercuric-albumin complex and low-molecular-weight thiols (Zalups and Koropatnick, 2000). The low-molecular-weight thiol conjugates seem to be much less labile in an aqueous environment (Rabenstein, 1989; Farrell et al., 1990; Oram et al., 1996). In fact, administration of such thiols alters both the toxicity and disposition of Hg2+ in the kidney (Zalups and Barfuss, 1996, 1998a; Zalups, 1998b). For example, intravenous coadministration of endogenous low-molecular-weight thiols (including Cys, homocysteine, NAC) with Hg2+ to rats has been demonstrated to increase both the level of renal accumulation and intoxication, indicating that these low-molecular-weight thiol conjugates are taken up by specific molecular mechanisms (Zalups and Barfuss, 1996, 19998a; Zalups, 1998b), potentially including the organic anion transport system.
The goal of the present investigation was to determine whether the organic anion transporters play a mechanistic role in the uptake, accumulation, and toxicity of inorganic mercury in renal cells, thereby supporting prior in vivo studies. To this end, we used a cell-line transfected with hOAT1 to assess both uptake and cytotoxicity of Hg2+ and thiol- Hg2+complexes. In addition, we characterized the uptake of the NAC-Hg2+ conjugate in both hOAT1- and rOAT3-expressing oocytes.
Materials and Methods
Tissue Culture.
A mycoplasma-free type II MDCK subclone with low transepithelial resistance was supplied by Dr. Daniel Balkovetz (University of Alabama at Birmingham, Birmingham, AL). This line was originally developed in the laboratory of Dr. Kai Simmons (European Molecular Biology Laboratory, Heidelberg, Germany). These cells were grown in a humidified atmosphere of 5% CO2/95% O2 at 37°C in culture media composed of EMEM (Invitrogen, Carlsbad, CA). supplemented with 1 mM sodium pyruvate and 10% fetal bovine serum (Invitrogen). This EMEM formulation will be referred to as “supplemented EMEM” throughout this article. Cells were split every 3 to 7 days, and 5 to 10% of the culture was inoculated into new flasks.
Cell Transfection.
MDCK cells were transfected with hOAT1 cDNA ligated to pcDNA3.1 (Invitrogen) using SuperFect Reagent (QIAGEN, Valencia, CA) according to the manufacturer's protocol (5 μl of SuperFect/μg of DNA). Surviving cell clones were maintained in culture media with 200 μg/ml G418 (Invitrogen) and screened for organic anion transport activity by assaying the uptake of [3H]PAH, as described below. The clone displaying the greatest level of [3H]PAH uptake (greater than 20 times the level in nontransfected MDCK cells) was used in the present study.
Production of Isotopic Hg2+.
Mercuric oxide (3 mg) containing the stable isotope200Hg2+ and enriched202Hg2+ (target) were weighed and doubly sealed in quartz tubing (actual mercuric oxide isotopic composition, <0.05% 196Hg, 1.5%198Hg, 2.82% 199Hg, 4.24%200Hg, 3.11% 201, 86.99%202Hg, and 1.34% 204Hg). The double-encapsulated target was sent to the Missouri University Research Reactor facility to be irradiated (by neutron activation) for 4 weeks. The irradiated target was placed in protected storage for 10 days to allow for the isotopic decay of the newly formed197Hg2+. The target was removed from the quartz tubing with four 50-μl rinses of 1N HCl. All four rinses were placed and sealed in a single 1.7-ml polypropylene vial. A sample of the solution was then used to determine the precise solid content of Hg using plasma-coupled elemental mass spectrometry. The radioactivity of the solution was determined by use of a PerkinElmer Wallac (Gaithersburg, MD) Wizard 3“ 1480 Automatic Gamma Counter (203Hg counting efficiency,∼50%). The specific activities of the203Hg2+ used in the present study ranged between 8 and 12 mCi/mg Hg.
Hg2+ Tracer Uptake in hOAT1 Transfected and Nontransfected MDCK Cells.
In the MDCK cell clone selected for these experiments, hOAT1 is expressed in both apical and basolateral faces (J.B. Pritchard et al., unpublished results). Apical expression of hOAT1 allows for the uptake to be studied in cells grown on solid support. Thus, for these experiments, cells were plated in 24-well (2.0 cm2) cell-culture cluster plates (Corning Glassworks, Corning, NY) in supplemented EMEM at a density of 0.5 × 106 cells/well (added as 2 ml). They were then grown in a humidified atmosphere of 5% CO2/95% air at 37°C for 2 days. Media were changed after the first 24 h.
For transport assays, media were aspirated from wells, and cells were rinsed three times with three volumes of 3 ml each of Hank's buffered saline solution (HBSS) supplemented with 10 mM HEPES, pH 7.4. Transport buffer (333 μl; specific to each experiment) containing radioactive203Hg2+ was added to each well. Thiols were always added in a 4:1 M ratio to the mercuric cation concentration to ensure the formation of linear II coordinate complexes (Rabenstein, 1989). At defined times, wells were rinsed with cold (4°C) stop buffer [HBSS supplemented with 10 mM HEPES, pH 7.4, with 1 mM 2,3-dimercapto-1-propane-sulfonic acid (DMPS) and 200 μM probenecid]. Because DMPS oxidizes rapidly in aqueous solutions, it was added less than 1 h before use. Cellular Hg2+ uptake was determined after adding 1 ml of 1 N NaOH to each well and shaking (in an orbital shaker at 500 rpm) for 24 h. Cell lysate (700 μl) from each well was neutralized with 700 μl of 1 N HCl and added to 15 ml of EcoLume (ICN Biomedicals Inc., Costa Mesa, CA) scintillation fluid. The radioactivity of each sample was determined using a Tri-Carb 2900TR Liquid Scintillation Analyzer (PerkinElmer Life Sciences, Boston, MA;203Hg counting efficiency,∼80–90%). Of the remaining cell lysate in each well, 50 μl was processed for Bradford protein assay (Bradford, 1976). Data for each well were normalized to the corresponding protein concentration.
Cell Viability/Toxicity Testing.
Cell viability was measured with an MTT-based in vitro toxicology assay (TOX1; Sigma, St. Louis, MO). This assay measures mitochondrial dehydrogenase activity by the conversion of the yellow tetrazolium dye MTT to purple formazan crystals. Cells were plated in supplemented EMEM at a density of 5.0 × 104 cells/well (added as 200 μl/well) in sterile 96-well microtiter plates (Corning Glassworks) and allowed to grow for 48 h in a humidified atmosphere of 5% CO2/95% air at 37°C. Supplemented EMEM was changed after the first 24 h by inversion. Excess media adhering to the plate was blotted off with sterile gauze (Johnson & Johnson Medical, Arlington, TX). After 48 h, wells were again washed with two washes of 200 μl/well of HBSS. After washing, test compounds were added to individual wells (200 μl/well) in unsupplemented EMEM, and cells were grown for 18 h (unless otherwise specified) in a humidified atmosphere of 5% CO2/95% O2 at 37°C. At the conclusion of the exposure period, media were removed by inversion and blotting, wells were washed with 200 μl of HBSS, and 100 μl of 0.5 mg/ml (1.2 mM) MTT in HBSS was added to each well. Cells were incubated for 2 h, and 100 μl of solubilization buffer (10% Triton X-100, 0.1 N HCl in isopropyl alcohol) were added to each well. This buffer both lysed the cells (releasing the formazan) and dissolved the water-insoluble formazan crystals. After overnight incubation at room temperature, full solubilization had occurred, and plates were read at 570 nm with a SpectraMax 340 microtiter plate reader (Molecular Devices Corporation, Sunnyvale, CA) running SoftMax Pro.
Oocyte Preparation and Hg2+ Tracer Uptake in hOAT1- and rOAT3-Expressing Oocytes.
Female Xenopus laevisliver-fed frogs were obtained from Xenopus I (Ann Arbor, MI). The ovaries were removed from tricaine-anesthetized frogs, and oocytes were isolated and defolliculated as described previously (Sweet et al., 1997). Briefly, this procedure uses collagenase A digestion followed by an incubation in a K2HPO4buffer. Oocytes were then stored in an incubator at 18°C and allowed to recover overnight in oocyte Ringer's 2 (OR-2; 82.5 mM NaCl, 2.5 mM KCl, 1 mM Na2HPO4, 3 mM NaOH, 1 mM CaCl2, 1 mM MgCl2, 1 mM sodium pyruvate, and 5 mM HEPES, pH 7.6) supplemented with 5% horse serum and 50 μg/ml gentamicin. After the recovery period, stage IV and V oocytes were microinjected with 16.1 nl of either high-performance liquid chromatography water or capped RNA (hOAT1 or rOAT3 clones at 1.93 μg/μl).
Transport assays were conducted 3 days after injection. The oocytes were separated into groups of 10 in 24-well plates and washed three times with 1 ml of unsupplemented OR-2. Then, as with cell-transport assays, 333 μl of OR-2 containing 5 μM Hg2+(with203Hg2+) and 20 μM NAC, with or without 250 μM probenecid, was added to each well and incubated for 1 h at room temperature. After incubation, oocytes were rinsed three times with 1 ml of ice-cold stop buffer composed of OR-2 supplemented with 200 μM probenecid and 1 mM DMPS (added no more than 1 h before use). Oocytes were then individually lysed in 200 μl of 10% SDS. After complete lysis, 4 ml of EcoLume (ICN Biomedicals) scintillation fluid was added to each vial, and samples were counted for radioactivity with use of a Tri-Carb 2900TR Liquid Scintillation Analyzer (PerkinElmer Life Sciences).
Statistical Analysis.
Results are presented as representative data from at least two experiments. Data are expressed as the mean ± standard error. For cytotoxicity studies, a sample size of n = 4 was used, and for uptake studies, a sample size of n = 3 was used. Assuming that each sample was mutually independent, statistical analysis was performed using two-way analysis of variance on log-transformed data followed by either Tukey's or Dunnett's post hoc test. Data were analyzed with the use of SAS version 8.0 (SAS Institute, Cary, NC). Differences among means were considered statistically significant at P < 0.05.
Results
Cellular Viability.
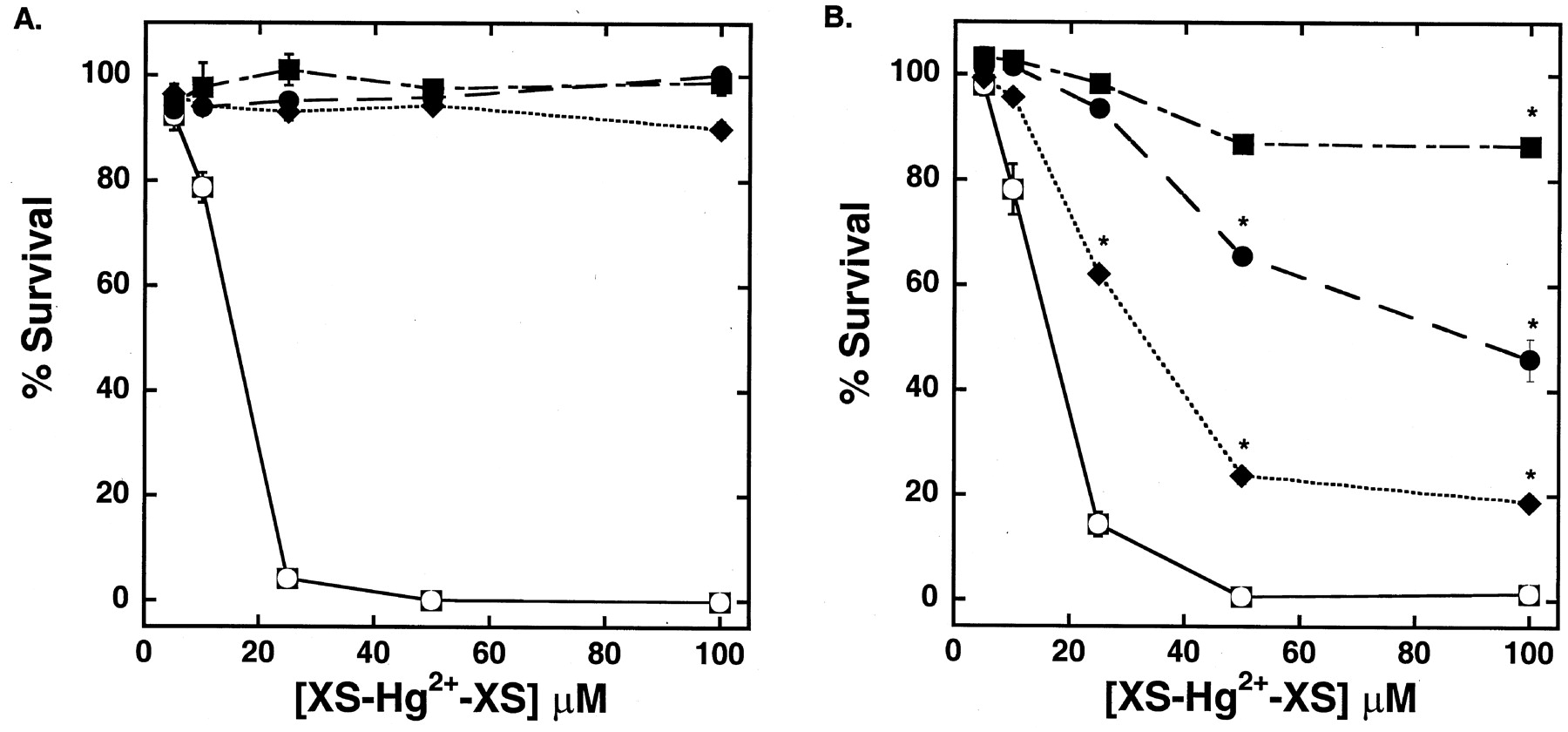
As shown in Fig.1, sensitivity of control and hOAT1-transfected MDCK cells to 18 h of exposure to HgCl2 was similar through the range of concentrations tested. The LD50 for mercuric chloride was approximately 16 μM. Mercuric conjugates of Cys, NAC, or GSH were less toxic than HgCl2 in both cell types. However, sensitivity of the nontransfected cells was markedly lower than that in the cells expressing hOAT1. As seen in Fig. 1A, there was no significant toxicity of the thiol-Hg2+ conjugates in the control cells, even at Hg2+ concentrations four times greater than the concentration of HgCl2 that yielded complete loss of viability. In contrast, exposure to the thiol-Hg2+ conjugates caused marked toxic effects in hOAT1-transfected cells over the same concentration range (Fig. 1B). Cellular viability in the cells transfected with hOAT1 was reduced by 81% during the 18 h of exposure to 100 μM of the NAC-Hg2+ conjugate. The Cys-Hg2+ conjugate (100 μM) was intermediate in toxicity, with a 54% decrease in survival. The GSH-Hg2+ conjugate was the least toxic, with only 13% mortality at the same concentration and time of exposure.

Viability of nontransfected (A) and hOAT1-transfected (B) MDCK cells after an 18-h exposure to increasing concentrations of HgCl2 (circle in filled square) (no conjugate), and thiol-Hg2+ conjugates (with a 4-fold excess of thiol) including GSH (▪), NAC (♦), and Cys (●). Points reflect means of replicates of four ± S.E. Two-way ANOVAs (Tukey's test) of logarithmically transformed data were used to compare the presence of hOAT1 at each of the different chemical doses. *, P< 0.05.
In hOAT1-expressing cells, the time profile of cell death induced by the NAC-Hg2+ conjugate (the thiol conjugate most toxic to hOAT1-expressing cells) was compared with that induced by HgCl2 (which was equally toxic to both hOAT1-transfected and nontransfected cells). As seen in Fig.2, there was a time lag in the toxic effects induced by the NAC-Hg2+ conjugate, compared with that induced by HgCl2 in hOAT1-expressing cells. Moreover, measurable toxic effects were not apparent until after 12 h of exposure. Toxic effects induced by HgCl2 occurred more rapidly and were clearly apparent by 4 h.

Time-dependence of toxicity of 20 μM HgCl2 (circle in filled square) and the NAC-Hg2+ conjugate (400:100 μM) (▪) on hOAT1-transfected MDCK cells. Points reflect means of replicates of four ± S.E. Two-way ANOVAs (Dunnett's test) of logarithmically transformed data were used to compare the effect of the treatments to untreated control cells. *, P < 0.05.
If the toxic effects of the mercuric conjugates in the hOAT1-transfected cells were directly related to the cellular uptake of the conjugates, then inhibition of hOAT1-mediated transport should result in a decreased level of toxic injury. As expected, when 200 μM probenecid or PAH (OAT1 inhibitors) was coincubated with the NAC-Hg2+ conjugate, the level of toxicity in the hOAT1-transfected MDCK cells was markedly reduced (Fig.3). Treatment with probenecid increased the level of survival of the hOAT1-transfected cells from 34 to 93%, similar to that seen in the nontransfected cells treated in the same manner. PAH also prevented an increase in cell death, although it was not as effective as probenecid, increasing the fraction of surviving hOAT1-transfected cells from 34 to 54%. The palliative effects of probenecid and PAH in the cells exposed to mercuric conjugates of Cys were less pronounced than those in the cells exposed to the NAC-Hg2+ conjugate. Thus, the level of protection provided by probenecid was only partial (from 50 to 75% survival), and PAH did not significantly reduce the toxic effects of the Cys-Hg2+ conjugate.

Effect of coincubation of 200 μM probenecid (Prob) or 200 μM PAH with mercuric conjugates of NAC and cysteine on the viability of hOAT1-transfected MDCK cells after an 18-h exposure. Points reflect means of replicates of four ± S.E. Two-way ANOVAs (Tukey's test) of logarithmically transformed data were used to compare the presence of hOAT1 at each of the treatments. *,P < 0.05.
A second class of specific OAT1 inhibitors includes dicarboxylates that exchange for organic anions at this transporter. These include glutarate, α-ketoglutarate, adipate, and suberate (Pritchard and Miller, 1993). Other dicarboxylates, such as succinate, fumarate, and malate, are not effective inhibitors of OAT1. As shown in Fig.4, the level of cellular death induced by mercuric conjugates of NAC was significantly reduced by the exchangeable dicarboxylates glutarate, α-ketoglutarate, and adipate. In contrast, succinate, a nonexchangeable dicarboxylate (Chatsudthipong and Dantzler, 1992), did not provide significant protection.

Effect of coincubation of various dicarboxylate acids (at 500 μM), including glutarate (GA), α-ketoglutarate (α-KG), Succinate (Suc), and Adipate (Adip), with 100 μM of the NAC-Hg2+ conjugate (concentration refers to mercuric species) on viability of hOAT1-transfected MDCK cells after an 18-h exposure. Points reflect means of replicates of four ± S.E. Two-way ANOVA (Dunnett's test) was used to compare the effect of the treatments to untreated control cells. *, P < 0.05.
Isotopic Hg2+ Uptake.
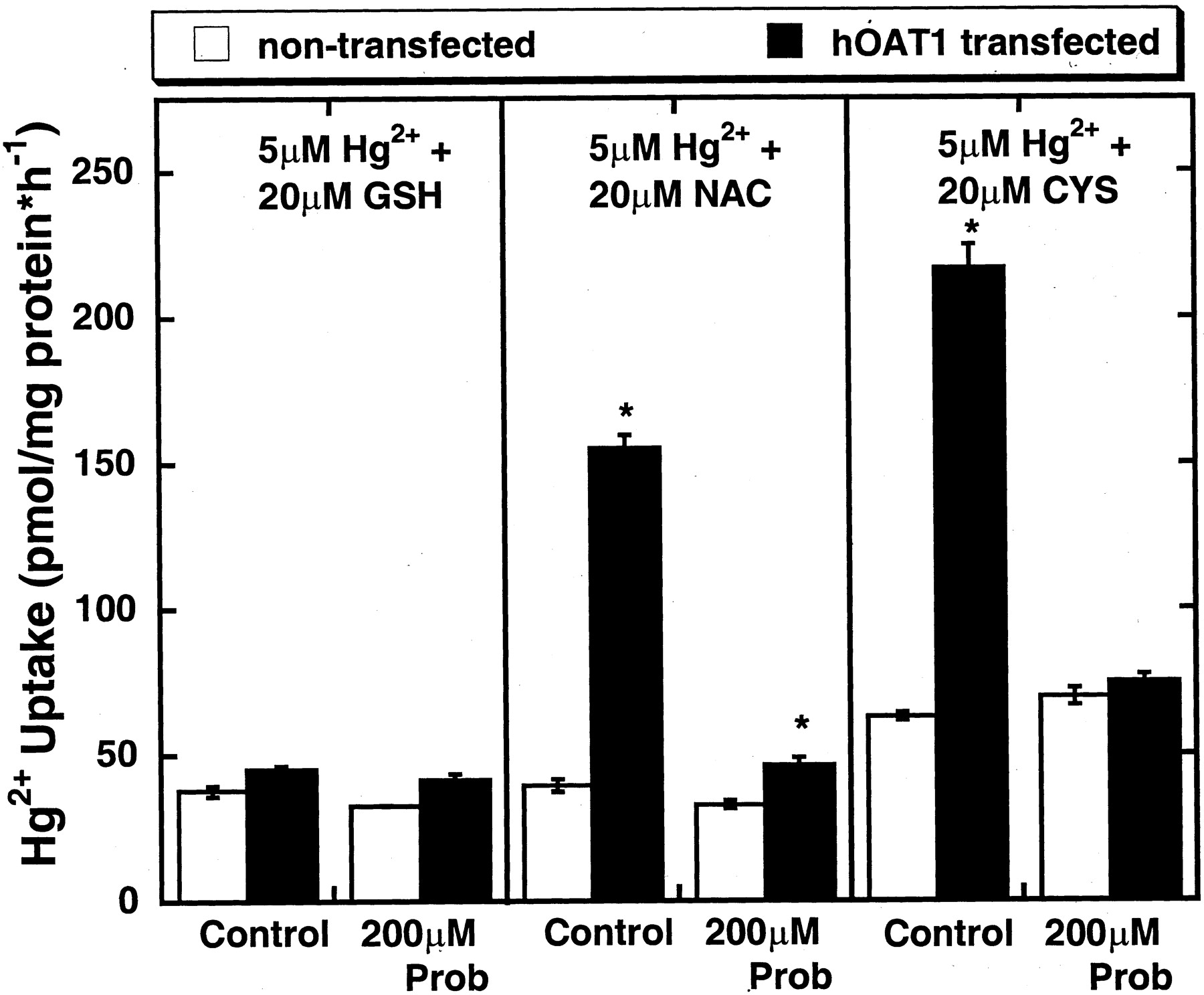
The data presented above suggest that the presence of hOAT1 provides a specific pathway for thiol-Hg2+ conjugates into cells and that increased cellular uptake leads to a corresponding increase in the level of toxicity. To test this directly, we measured the uptake of Hg2+ in control and hOAT1-expressing cells. As seen in Fig. 5, there was 3- to 4-fold greater uptake of the NAC-Hg2+ and Cys-Hg2+ conjugates in hOAT1-expressing cells than in control cells. However, uptake of the GSH-Hg2+ conjugate (the least toxic mercuric thiol species) was similar in both cell types. The hOAT1-dependent uptake (difference between uptake in hOAT1 transfected and nontransfected cells) of the NAC-Hg2+ and Cys-Hg2+ conjugates was almost completely abolished by probenecid (200 μM).

Uptake of thiol-Hg2+ conjugates over a 1-h time interval by hOAT1-transfected and nontransfected MDCK cells. Points reflect means of replicates of three ± S.E. Two-way ANOVAs (Tukey's test) of logarithmically transformed data were used to compare the presence of hOAT1 at each of the treatments. Prob, probenecid; *, P < 0.05.
As shown in Fig. 6, the uptake of the NAC-Hg2+ conjugate increased steadily over 2 h in both hOAT1-transfected and -nontransfected cells. The initial rate of uptake was only 0.6 pmol·mg · protein−1·min−1 for the nontransfected cells and 4.2 pmol·mg · protein−1·min−1 for the hOAT1-transfected cells. By 2 h, uptake in the hOAT1-transfected cells had not reached steady state and was 6-fold greater than that in nontransfected cells. Because uptake was linear at 40 min (Fig. 6), this time was used for kinetic studies. As shown in Fig.7, the apparentK m for hOAT1-dependent uptake of mercuric conjugates of NAC was 44 ± 9 μM, and theV max was 9 ± 1 pmol·(mg · protein)−1·min −1.

Time course of the NAC-Hg2+ conjugate (20:5 μM) uptake by nontransfected (circle in filled square) and hOAT1-transfected (▪) MDCK cells. Points reflect means of replicates of three ± S.E. Two-way ANOVA (Tukey's test) of logarithmically transformed data were used to compare the presence of hOAT1 at each of the treatments; *, P < 0.05.

Kinetics of the NAC-Hg2+ conjugate uptake (40 min) by hOAT1-transfected (▪) and nontransfected MDCK cells (circle in filled square). The hOAT1 component (▾) was fitted to a hyperbolic equation with the use of KaleidaGraph 3.0 (Synergy Software, Reading, PA). Points reflect means of replicates of three ± S.E. Two-way ANOVAs (Tukey's test) of logarithmically transformed data were used to compare the presence of hOAT1 at each of the treatments. *,P < 0.05.
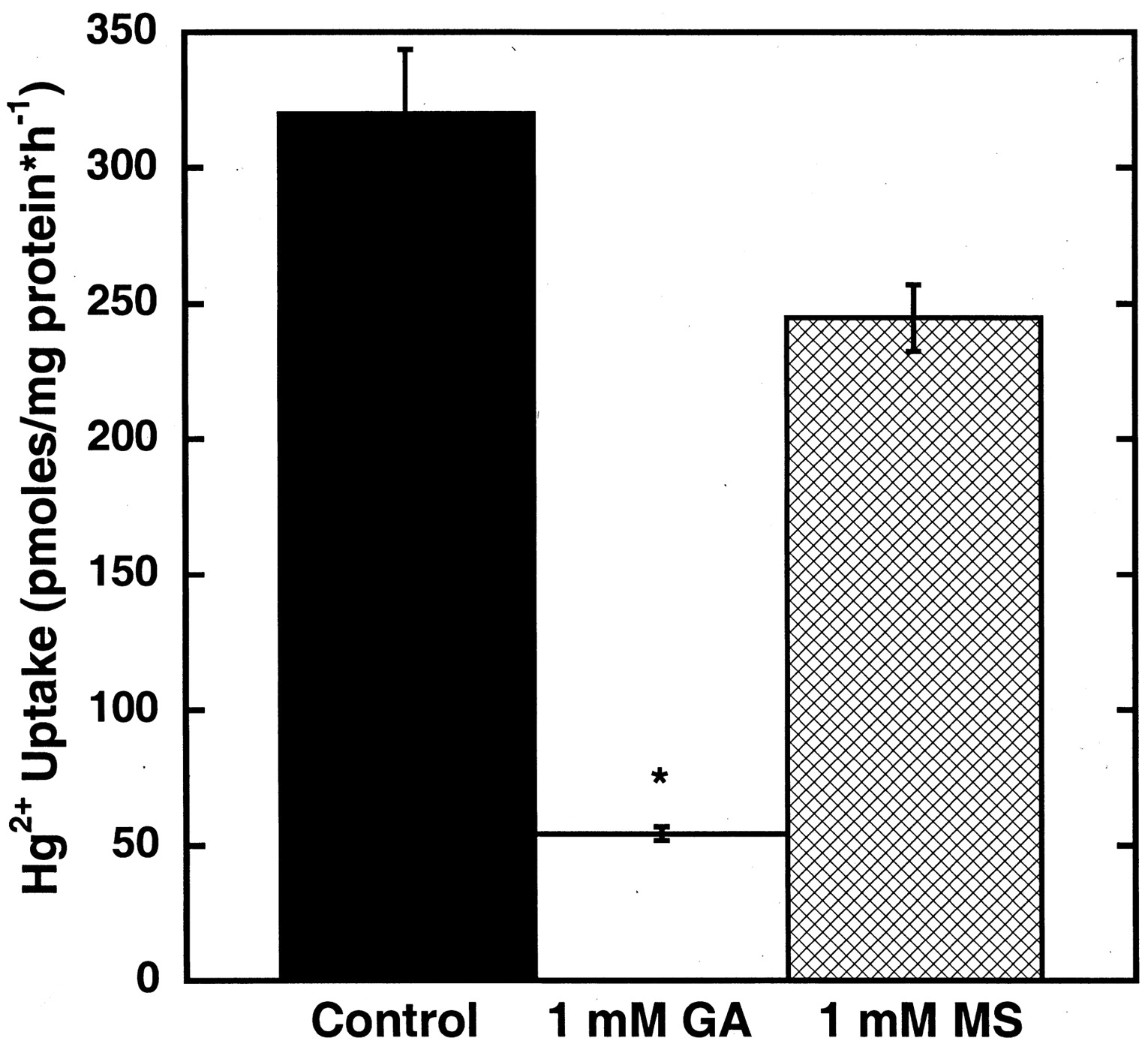
To confirm the participation of hOAT1 in the uptake of the NAC-Hg2+conjugate, the effects of exchangeable (glutarate) and nonexchangeable (methylsuccinate) dicarboxylates on uptake were assessed. As shown in Fig. 8, 1 mM glutarate almost completely cis-inhibited the hOAT1-mediated component of the inward transport of the NAC-Hg2+ conjugate. However, 1 mM methylsuccinate did not significantly alter uptake compared with the level of uptake in hOAT1-transfected cells exposed only to the NAC-Hg2+ conjugate.

Effect of coincubation of glutarate (GA) or methyl succinate (MS) with the NAC-Hg2+ conjugate (20:5 μM) on that species' uptake by hOAT1-transfected MDCK cells over a 1-h time duration. Points reflect means of replicates of three ± S.E. Two-way ANOVAs (Dunnett's test) of logarithmically transformed data were used to compare the effect of the treatments with those of untreated control cells; *, P < 0.05.
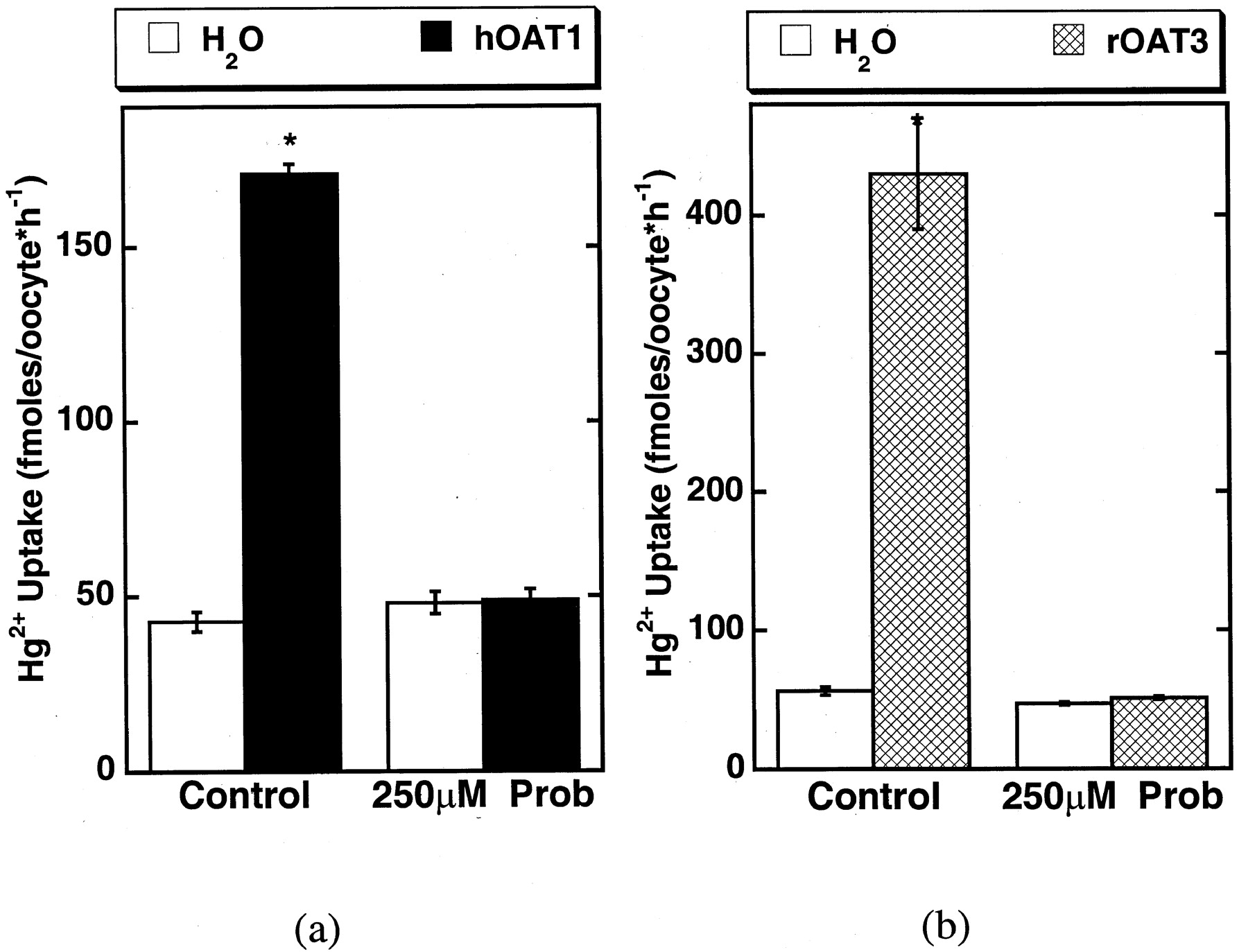
As a final test of the ability of hOAT1 to transport the mercuric conjugates and to determine whether the second basolateral organic anion transporter, OAT3, could also contribute to Hg2+ conjugate uptake, hOAT1 and rOAT3 clones were expressed in X. laevis oocytes (Fig.9). Uptake of 5 μM NAC-Hg2+ conjugate in hOAT1-expressing oocytes was much greater than that in water-injected control cells. hOAT1-mediated uptake was reduced to control levels by 250 μM probenecid (Fig. 9a). rOAT3-expressing oocytes also showed marked probenecid-sensitive uptake of Hg2+-NAC (Fig.9b). Thus, the expression of either clone bestowed enhanced ability to accumulate the NAC-Hg2+ conjugate in this expression system, suggesting that both OATs may contribute in vivo to basolateral accumulation of thiol-Hg2+ conjugate.

Uptake (60-min) of 5 μM NAC-Hg2+conjugate uptake by hOAT1 (a) and rOAT3 (b) expressing X. laevis oocytes in the presence and absence of 250 μM probenecid (Prob). Control experiments were done in water-injected oocytes. Points reflect means of replicates of 9 to 10 ± S.E. Two-way ANOVAs (Dunnett's test) of logarithmically transformed data were used to compare the response in clones with that of water-injected controls; *, P < 0.05.
Discussion
The kidney is the primary target organ for inorganic mercury toxicity (Zalups, 2000). Within the kidney, the primary sites of mercury accumulation and toxicity are the S2 and S3 segments of the proximal tubule (Zalups and Barfuss, 1990; Zalups, 1991a,b). These are also the primary sites of OAT1 and OAT3 expression (Hosoyamada et al., 1999; Sweet et al., 1999; Tojo et al., 1999; Cha et al., 2001; Hasegawa et al., 2002). Moreover, in vivo studies suggest that OAT1 and/or OAT3 may participate in uptake of toxic mercuric species, because a variety of OAT inhibitors decrease the severity of the nephropathy induced by the mercuric species (Zalups, 1998a,b; Zalups and Barfuss, 1998b). In agreement with in vivo studies, the data reported here provide molecular evidence that hOAT1 and rOAT3 can mediate the uptake of thiol-Hg2+ conjugates (particularly the NAC-Hg2+ and Cys-Hg2+conjugates). Both uptake and toxicity of the thiol-Hg2+ conjugates were much greater in MDCK cells expressing hOAT1 than in control cells lacking this transporter (Figs. 1 and 5). Furthermore, both uptake and toxicity were reduced by inhibitors of OAT-mediated transport, indicating that thiol-Hg2+ conjugates are substrates of hOAT1 (Figs. 3-5). This is also the case for uptake of the NAC-Hg2+ conjugate by rOAT3 in the X. laevis oocyte expression system (Fig. 9). In contrast, uptake of inorganic mercuric ions seems to occur by an hOAT1-independent mechanism, because this form of mercury was equally toxic to hOAT1-transfected and -nontransfected cells. It is important to note that inorganic mercury was far more toxic than any of the conjugated species. Indeed, not only was the overall toxicity greater, but toxicity was observed at much lower concentrations. Thus, formation of mercury complexes or conjugates was generally protective. However, as shown in the present study, some of these conjugates are OAT substrates. Therefore, OAT1 and OAT3 seem to provide specific paths for the entry of certain thiol-Hg2+ conjugates into renal cells, potentially leading to proximal tubular damage. Likewise, competition for this transport leads to the prevention of toxic effects of Hg2+ upon administration of OAT inhibitors such as PAH and probenecid in vivo (Zalups, 1998a,b) and in vitro (Fig.3).
The basis for transport of mercuric-thiol conjugates by hOAT1 and rOAT3 seems to rest in the structure of the conjugates (Burckhardt et al., 2001). For instance, the NAC-Hg2+ conjugate is similar in structure to many mercapturic acids (N-acetylcysteine S conjugates of various organic molecules). In fact, mercapturic acids have been shown previously to be secreted by the renal proximal tubule (Stevens and Jones, 1989). In addition, it was recently shown that several mercapturic acids [i.e.,S-(2,4-dinitrophenyl)-N-cysteine] are specific hOAT1 substrates (Pombrio et al., 2001).
Among the thiol conjugates studied, both the hOAT1-dependent uptake and toxicity of the NAC-Hg2+ conjugate were particularly striking. This conjugate was also taken up by the rOAT3-expressing oocytes. These results are entirely consistent with the in vivo demonstration by Zalups and Barfuss (1998a) that uptake of this conjugate is localized entirely to the basolateral membrane, the site of OAT1 and OAT3 expression (Hosoyamada et al., 1999; Sweet et al., 1999; Tojo et al., 1999; Cha et al., 2001; Hasegawa et al., 2002). However, the relative contributions of the various thiol-Hg2+ conjugates to mercury toxicity in vivo remain an open question. Certainly, the plasma concentrations of cysteine- and GSH-mercuric conjugates far exceed that of NAC in vivo (Mansoor et al., 1992; Chassaing et al., 1999). On the other hand, clinical administration of thiols, such as NAC for various medical conditions (i.e., acetaminophen poisoning, etc.) (Schiodt et al., 2002), may greatly alter the potential for formation and renal accumulation of specific thiol-Hg2+ conjugates.
Finally, although this study clearly links the uptake and toxicity of mercuric compounds with OAT1 and potentially OAT3, this does not mean that organic anion transporters are the only potential paths for proximal tubular uptake of mercuric conjugates. Evidence already exists for the involvement of certain luminal amino acid transporters as well (Cannon et al., 2001). Thus, the story is not yet complete, but the ability of OAT1 and OAT3 to transport the mercuric conjugates, coupled with in vivo evidence that OAT inhibitors reduce renal toxicity of mercury, indicates that this transport plays an important role in the development of renal damage induced by Hg2+.
Acknowledgments
We thank Dr. Safaraz Ahmed, Ramsey Walden, and Porché D. Kirkland for their invaluable assistance in the radioisotopic uptake portion of this project. We also thank Laura Hall for her assistance with the X. laevis oocyte preparation and injection. Furthermore, we thank Dr. Shyamal D. Peddeda for his assistance in the statistical analysis. We also thank Dr. Delon Barfuss at Georgia State University for his assistance in supplying radioactive mercury for the experiments discussed in this manuscript.
Footnotes
- Received May 9, 2002.
- Accepted November 18, 2002.
-
R.K.Z. is supported, in part, by National Institute of Environmental Health Science grants ES05157, ES05980, and ES11288.
Abbreviations
- OAT
- organic anion transporter
- DMPS
- 2,3-dimercapto-1-propane-sulfonic acid
- EMEM
- Eagle's modified essential medium
- Cys
- cysteine
- GSH
- glutathione (reduced)
- HBSS
- Hanks' buffered saline solution
- hOAT1
- human organic anion transporter 1
- MDCK
- Madin-Darby canine kidney cells
- MTT
- 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylterazolium bromide
- NAC
- N-acetylcysteine
- PAH
- p-aminohippurate
- ANOVA
- analysis of variance
- rOAT3
- rat organic anion transporter 3
- OR-2
- oocyte Ringer's 2
- U.S. Government





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}