


Abstract
OCTN2 is an Na+-dependent transporter for carnitine, which is essential for fatty acid metabolism, and its functional defect leads to fatal systemic carnitine deficiency (SCD). It also transports the organic cation tetraethylammonium (TEA) in an Na+-independent manner. Here, we studied the multifunctionality of OCTN2, by examining the transport characteristics in cells transfected with mouse OCTN2 and in juvenile visceral steatosis (jvs) mice that exhibit a SCD phenotype owing to mutation of the OCTN2 gene. The physiological significance of OCTN2 as an organic cation transporter was confirmed by usingjvs mice. The embryonic fibroblasts fromjvs mice exhibited significantly decreased transport of [14C]TEA. Pharmacokinetic analysis of [14C]TEA disposition demonstrated that jvsmice showed decreased tissue distribution and renal secretory clearance. In transport experiments using OCTN2-expressing cells, TEA and carnitine showed mutual trans-stimulation effects in their transport, implying a carnitine/TEA exchange mechanism. In addition, Na+ affected the affinity of carnitine for OCTN2, whereas Na+ is unlikely to be involved in TEA transport. This is the first molecular and physiological demonstration of the operation of an organic cation transporter in renal apical membrane. The results are consistent with the physiological coupling of carnitine reabsorption with the secretion of organic cations.
Organic cations such as tetraethylammonium (TEA) are distributed into various tissues and excreted into urine by specific transport systems in kidney. Previous studies suggested that organic cations are excreted by potential-dependent OCT transporters and by an organic cation/H+ antiporter across the renal epithelial basolateral and apical membranes, respectively (Zhang et al., 1998;Koepsell et al., 1999; Sweet and Pritchard, 1999), whereas no apically expressed transporters have been molecularly identified. We have isolated and characterized the new family of organic cation transporters, OCTN. The first member of the OCTN family, OCTN1, cloned from human fetal liver, transports organic cations such as TEA in a pH-dependent manner (Tamai et al., 1997; Yabuuchi et al., 1999). The second member, OCTN2, transports physiologically important carnitine in an Na+-dependent manner, as well as organic cations in an Na+-independent manner (Tamai et al., 1998; Wu et al., 1998, 1999; Ohashi et al., 1999). OCTN2 is present in various tissues, including kidney, skeletal muscle, heart, placenta, and others (Tamai et al., 1998). In 1988, the homozygous mutant mice, named juvenile visceral steatosis (jvs) mice, which exhibit cardiac hypertrophy, lipid accumulation in the liver, and hyperammonemia, (Koizumi et al., 1988; Horiuchi et al., 1993) were found with several histological changes (Koizumi et al., 1988; Narama et al., 1997) and alteration of carnitine disposition (Yokogawa et al., 1999a). The significance of OCTN2 as the carnitine transporter was clearly demonstrated using jvs mice, which exhibit the phenotype of systemic carnitine deficiency (SCD) caused by mutation of the OCTN2 gene (Leu352Arg) (Hashimoto et al., 1998; Lu et al., 1998;Nezu et al., 1999; Yokogawa et al., 1999c). However, no precise studies on organic cation transport by mouse OCTN2 have been done, and the physiological and pharmacological relevance of OCTN2 as an organic cation transporter remains to be clarified. Furthermore, the mechanism of the multifunctionality of OCTN2, transporting carnitine and organic cations in Na+-dependent and independent manners, respectively, is unclear. Because the jvs mice have genetically mutated OCTN2 and completely lack carnitine transport activity, they are expected to be useful to evaluate a contribution of OCTN2 as an organic cation transporter.
In the present study, we studied the multifunctionality of OCTN2 and the significance of OCTN2-mediated tissue distribution and renal excretion of the organic cation TEA by comparing the dispositions in wild type and jvs mice, as well as by examining transport in OCTN2-expressing cells.
Experimental Procedures
Materials.
[Methyl-3H]acetyl-l-carnitine hydrochloride (65 Ci/mmol) andl-[methyl-3H]carnitine hydrochloride (85 Ci/mmol) were purchased from Moravek Biochemicals Inc. (Brea, CA). [1-14C]Tetraethylammonium bromide (2.4 mCi/mmol), [N-methyl-3H]1-methyl-4-phenylpyridinium acetate (MPP) (82 Ci/mmol), [N-methyl-3H]verapamil hydrochloride (78.6 Ci/mmol) and [glycyl-2-3H]p-aminohippurate (2.45 Ci/mmol) were from New England Nuclear (Boston, MA). [Pyridinyl-5-3H]pyrilamine (28 Ci/mmol) was from Amersham Pharmacia Biotech (Buckinghamshire, UK). Other reagents, including cefazolin sodium, were obtained from Sigma Chemical Co. (St. Louis, MO), Wako Pure Chemical Industries (Osaka, Japan), and Nacalai Tesque, Inc. (Kyoto, Japan) and used without further purification. HEK 293 cells were obtained from Japanese Cancer Research Resources Bank (Tokyo, Japan).
Studies of Uptake and Efflux by Fibroblasts and HEK 293 Cells Expressing Mouse OCTN2.
Primary cultured fibroblasts from homozygous jvs and normal C57BL/6J embryos were prepared and cultured as described previously (Hashimoto et al., 1998). Briefly, the cells obtained were cultured in Dulbecco's modified Eagle's medium supplemented with 5% fetal calf serum (Life Technologies, Tokyo, Japan) on plastic disks in humidified 5% CO2 at 37°C and used for uptake experiments. In the transport measurement, the cells cultured on plastic disks were incubated with transport medium (Hanks' balanced salt solution) containing a radiolabeled test compound to initiate the uptake study. After an appropriate time, the disks were washed three times with ice-cold Hanks' balanced salt solution, solubilized with 1 N NaOH, and the radioactivity was measured in a liquid scintillation counter after neutralization with HCl and addition of liquid scintillation fluid, Clear-sol I (Nacalai Tesque).
For transport studies in HEK 293 cells expressing mouse OCTN2, the full-length wild-type or jvs mouse OCTN2 cDNA, subcloned into the BamHI sites of the expression vector pcDNA3 and the construct, pcDNA3/mouse OCTN2, was transfected into HEK 293 cells by means of the calcium phosphate precipitation method as described previously (Tamai et al., 1997). The cells were cultivated in Dulbecco's modified Eagle's medium containing 10% fetal calf serum, penicillin and streptomycin in a humidified incubator at 37°C under 5% CO2. After 24-h cultivation of the cells in the 15-cm dishes, pcDNA3/OCTN2 or pcDNA3 vector alone (Mock) was transfected by adding 10 μg of the plasmid DNA per dish. At 48 h after transfection, the cells were harvested with a rubber policeman, washed twice, and suspended in the transport medium containing 125 mM NaCl, 4.8 mM KCl, 5.6 mM d-glucose, 1.2 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 25 mM HEPES, pH 7.4. In an uptake measurement, the cell suspension was mixed with transport medium containing a radiolabeled test compound to initiate the uptake study. In the trans-stimulated uptake experiments, the cells were preloaded with TEA for 30 min and subsequently uptake of [3H]carnitine by the cells was measured. At appropriate times, 100- or 200-μl aliquots of the mixture were withdrawn and the cells were separated from the transport medium by centrifugation in a microtube containing a mixture of silicon oil and liquid paraffin with a density of 1.03 and 3 N KOH. The resultant cell pellets were solubilized in 3 N KOH, neutralized with HCl, and the associated radioactivity was measured in a liquid scintillation counter. In the trans-stimulated efflux experiments, the cells were preloaded with [14C]TEA for 30 min at 37°C. Then, the cells were spun down (0°C, 7,000g, 8 sec), suspended in ice-cold transport medium, and washed twice at 0°C. The cells were again suspended in the transport medium containing test compounds, and efflux was initiated at 37°C. At appropriate times, 100-μl aliquots of the mixture were withdrawn and the cell-associated radioactivity was measured as described above. Cellular protein content was determined according to the method of Bradford using a Bio-Rad protein assay kit (BioRad, Hercules, CA) and bovine serum albumin as the standard (Bradford, 1976). For Na+-free experiments, the obtained cells were suspended in Na+-free medium, in which Na+ was replaced isotonically withN-methyl-d-glucamine (NMG).
Animal Experiments.
All the animal experiments were performed according to the Guidelines for the Care and Use of Laboratory Animals in Takara-mach Campus of Kanazawa University.Jvs mice, originally found among mice of the C3H.OH strain (Koizumi et al., 1988), and control wild mice C3H/HeJ (Japan SLC, Hamamatsu, Japan) were used for the following studies. By mating heterozygous male mice with heterozygous female mice, we obtained homozygous mutants (jvs/jvs). Eight-week-oldjvs male mice (jvs/jvs) and normal male C3H/Hej mice (+/+) were used to study the disposition of TEA after overnight starvation. Mice were anesthetized with pentobarbital and were bolus-injected with 8.8 mg/kg of [14C]TEA via the jugular vein. [3H]Inulin was simultaneously injected with [14C]TEA to evaluate glomerular filtration rate (GFR). Serial blood samples were collected from the intraorbital venous plexus using heparinized capillary tubes at designated time intervals in individual mice during the experiment. Urine samples were collected by washing the bladder with saline (0.5 ml) at designated times through a catheter. For determination of the apparent tissue-to-plasma concentration ratio (K p), the mice were decapitated at 4 h after a single intravenous injection of [14C]TEA. The tissues were quickly excised, rinsed well with ice-cold saline, blotted to dryness, and weighed. Plasma and urine samples were mixed with scintillation fluid for quantitation. Tissue samples (0.05–0.2 g) were dissolved in 1 ml of Soluene-350 (Packard Inc., Meriden, CT) by incubation at 50°C for 3 h. The dissolved samples were mixed with scintillation fluid, neutralized with 1 N HCl, and the associated radioactivity was measured. In the same manner with TEA, cefazolin at a dose of 20 mg/kg was intravenously injected to mice and the concentration of cefazolin in plasma and urine was measured by high-performance liquid chromatography. The used column and mobile phase for high-performance liquid chromatography analysis of cefazolin were TSKgel ODS-80Ts (4.6 × 150 mm; Tosoh, Tokyo, Japan) and acetonitrile/water containing 10 mM ammonium acetate (15:85), respectively.
Data Analysis.
Uptake of [3H]carnitine or [14C]TEA by in vitro experiments was usually expressed as the cell-to-medium concentration (C/M) ratio (in microliters per milligram of protein) obtained by dividing the uptake amount by the concentration of test compound in the medium. TheK m values of carnitine and TEA were 22.1 μM and 215 μM (Tamai et al., 2000), respectively, and concentrations of [3H]carnitine (10 nM) and [14C]TEA (50–100 μM) used in each experiment were low enough to be evaluated by the C/M ratio compared withK m values. Pharmacokinetic parameters such as the area under the plasma concentration-time curve (AUC), the elimination rate constant (k e), the steady-state distribution volume (Vdss) and the total body clearance (CLtot) were estimated according to model-independent moment analysis or compartment analysis using WinNonlin (SCI, Cary, NC). GFR was estimated in terms of inulin clearance. The renal clearance (CLr) and renal secretory clearance (CLs) were calculated by using the equations CLr = X / AUC and CLs = CLr − GFR, where X is the urinary excretion amount of [14C]TEA. All data are expressed as mean ± S.E.M. and statistical analysis was performed by using Student'st test, with p < 0.05 as the criterion of significance.
Results
Transport of [14C]TEA and [3H]Carnitine in Embryonic Fibroblasts from Wild-Type and jvsMice.
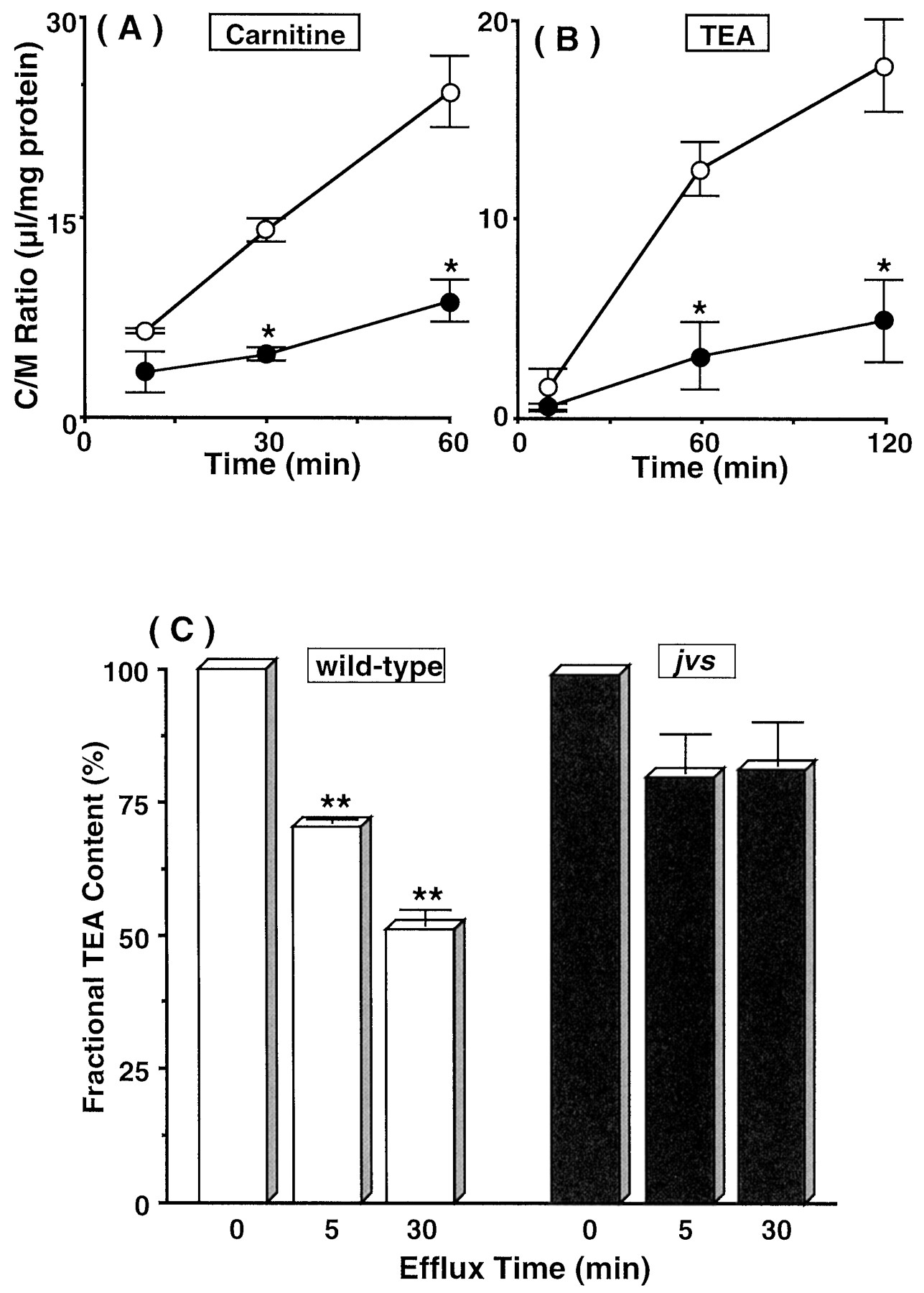
Fig. 1, A and B, show the uptakes of [3H]carnitine and [14C]TEA by embryonic fibroblasts from wild-type and jvs mice. Uptake of [14C]TEA by fibroblasts from jvsmice was significantly lower than that by the cells from normal mice, as was also the case for [3H]carnitine. Furthermore, after preincubation of fibroblasts with [14C]TEA, the efflux transport of [14C]TEA was measured (Fig. 1C). The efflux rate of [14C]TEA from wild-type fibroblasts was larger than that of jvs fibroblasts. The apparent rapid efflux of TEA from jvs mice-derived fibroblasts at the initial stage represents release of the cell-surface adsorbed [14C]TEA, because further decrease in cell-associated radioactivity was not observed. These findings demonstrated that TEA is bidirectionally transported in mouse embryonic fibroblasts and OCTN2 plays a role in the transport of TEA as well as carnitine.

Uptakes of [3H]carnitine (A) and [14C]TEA (B) and efflux of [14C]TEA (C) by embryonic fibroblasts from wild-type (○, open columns) andjvs (●, closed columns) mice. Uptakes of [3H]carnitine (10 nM) and [14C]TEA (50 μM) by fibroblasts were measured at 37°C and pH 7.4. In the [14C]TEA efflux study, cells were preloaded by incubating with 300 μM [14C]TEA for 60 min at 37°C. The effluxes of [14C]TEA were measured at 37°C and pH 7.4. *, Significantly different from the uptake by wild-type fibroblasts by Student's t test (p < 0.05). **, Significantly different from the content of [14C]TEA at 0 min by Student's t test (p < 0.05). The results are shown as means ± S.E.M. of four determinations.
Transport of [14C]TEA and [3H]Carnitine by Wild-Type and jvs OCTN2-Expressing HEK 293 Cells.
To compare the transport activities for carnitine and TEA via mouse OCTN2, we examined the uptakes of carnitine and TEA by HEK 293 cells transfected with cDNA of the wild-type OCTN2 and the mutated OCTN2 found in jvs mice. In the jvs-derived OCTN2-expressing cells, uptake of [3H]carnitine and [14C]TEA were both negligible, whereas wild-type OCTN2 exhibited significant increase of transport in usual transport medium containing Na+ (Fig.2A). When the uptakes by wild-type OCTN2-expressing cells were examined in the presence and absence of Na+ (Fig. 2B), [14C]TEA uptake was maintained in the absence of Na+, whereas [3H]carnitine uptake was disappeared. Accordingly, OCTN2 does not require Na+ for the transport of TEA, and the mutation in jvs OCTN2 (Leu352Arg) commonly leads to loss of transport activities of TEA and carnitine (Nezu et al., 1999).

Uptakes of TEA and carnitine by HEK 293 cells transfected with cDNA of wild-type and jvs-mutant OCTN2. Uptakes of [3H]carnitine (10 nM) and [14C]TEA (50 μM) by the cells were measured for 10 or 30 min, respectively, at 37°C and pH 7.4. (A) Uptakes of carnitine (left) and TEA (right) by cells transfected with the pcDNA vector alone (Mock, ▨) and the plasmid vector containing cDNA for wild-type OCTN2 (■) and jvs-mutant OCTN2 (▪) in transport buffer containing Na+. *, Significantly different from the uptake by jvs-mutant OCTN2-transfected cells by Student'st test (p < 0.05). (B) Uptakes of carnitine (left) and TEA (right) by cells transfected with the pcDNA vector alone (Mock ▨) and the plasmid vector containing cDNA for wild-type OCTN2 (■), were measured in the absence (−Na+) or presence (+Na+) of Na+ in the transport buffer. In the absence of Na+, Na+ was replaced with N-methyl-d-glucamine. *, Significantly different from the uptake by Mock cells by Student's ttest (p < 0.05). The results are shown as mean ± S.E.M. of three determinations.
Na+-Dependent Inhibition of OCTN2-Mediated [14C]TEA Uptake by Carnitine.
We next examined the inhibitory effect of various concentrations of carnitine on the OCTN2-mediated uptake of [14C]TEA in the presence and absence of extracellular Na+ (Fig.3). The reduction of [14C]TEA uptake by carnitine was concentration-dependent, being significantly greater in the presence of Na+. Accordingly, Na+ seems to influence the affinity of carnitine for OCTN2.
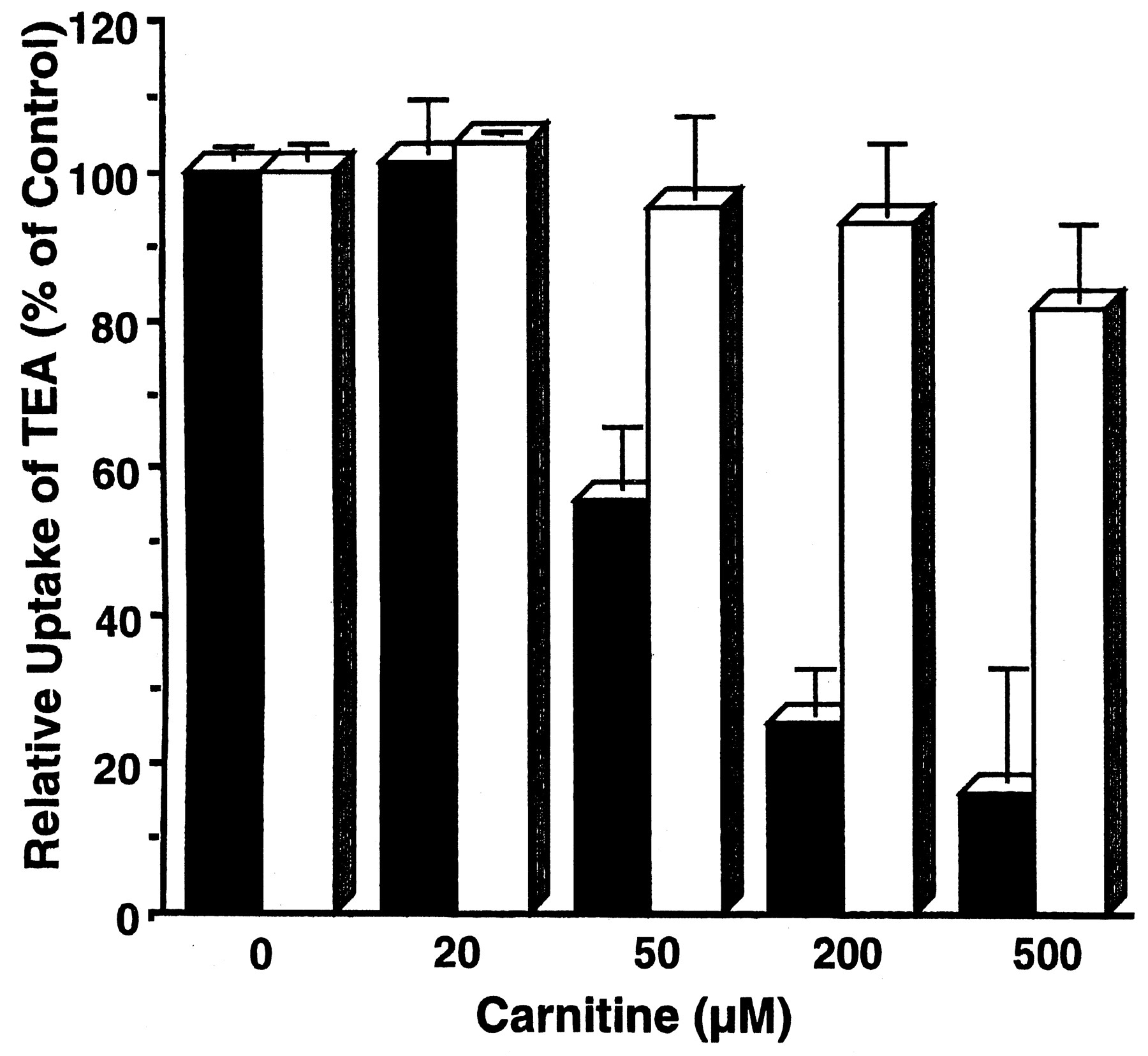
Na+-dependent inhibition by carnitine of TEA uptake by OCTN2-expressing HEK 293 cells. The uptakes of [14C]TEA (100 μM) were measured for 30 min at 37°C in transport buffer, pH 7.4, in the presence of various concentrations of carnitine. The results show the expressed-OCTN2 specific uptake obtained by subtracting the uptake by Mock cells from that by the OCTN2-expressing cells in the presence (▪) or absence (■) of Na+ in the transport buffer. Uptake in the absence of Na+ was determined by replacing Na+ withN-methyl-d-glucamine. The results are shown as means ± S.E.M. of three determinations.
Mutual trans-Stimulation Effects on TEA and Carnitine Transport by OCTN2.
To examine the functional coupling of the transport of TEA and carnitine, trans-stimulation effects via OCTN2 were measured. Based on the Na+dependence of carnitine transport and the physiological concentration of Na+, trans-stimulation effects were evaluated by measuring influx and efflux transports of carnitine and TEA, respectively. After preloading of OCTN2-expressing HEK 293 or Mock cells with TEA at 100 μM, 500 μM, or 1 mM, the uptake of a tracer concentration of [3H]carnitine was evaluated in the presence or absence of Na+ (Fig.4, A and B). In Mock cells, a higher concentration of TEA (1 mM) was used to keep the intracellular TEA concentration at a level similar to that in OCTN2-expressing cells and a similar level of TEA uptake at 500 μM, which showed significant effect on carnitine uptake, by OCTN2-expressing cells. When OCTN2-expressing cells were preloaded with TEA, uptake of [3H]carnitine was significantly increased compared with that by the cells without preloading. The extent of the increment was dependent on the concentration of TEA preloaded, with a higher uptake at higher concentrations of preloaded TEA (Fig. 4A). In the absence of Na+, however, no stimulation of the uptake of [3H]carnitine by TEA was observed (Fig. 4B). Thus, efflux of TEA seems to facilitate the Na+-dependent carnitine influx.

Trans-stimulation of [3H]carnitine uptake (A, B) and [14C]TEA efflux (C) by OCTN2-expressing HEK 293 cells. In trans-stimulation of [3H]carnitine uptake, OCTN2-expressing (shown as OCTN2) or Mock (shown as Mock) cells were preloaded with 0, 100, 500 μM or 1 mM TEA for 30 min. Subsequently, uptakes of 10 nM [3H]carnitine were measured in the presence (A) or absence (B) of Na+ in the transport buffer. Intrans-stimulation of [14C]TEA efflux, cells were preloaded by incubating with 100 (OCTN2-expressing cells, solid lines) or 200 μM (Mock, broken line) [14C]TEA for 30 min at 37°C (C). The effluxes of [14C]TEA were measured in the absence (■ as control) or presence of 250 μM carnitine (○), 10 mM TEA (▵) or 1 mM quinidine (░). The results are shown as means ± S.E.M. of three determinations. *, Significantly different from control of OCTN2-mediated uptake by Student's t-test (p < 0.05). **, Significantly different from control of OCTN2-mediated efflux by Student's t test (p < 0.05).
After preincubation of OCTN2-expressing and Mock cells with [14C]TEA, the efflux transport of [14C]TEA was measured in the presence and absence of carnitine in the extracellular medium (Fig. 4C). The efflux rate of [14C]TEA was higher in OCTN2-expressing cells than in Mock cells. In the presence of a strong inhibitor of OCTN2, quinidine (Ohashi et al., 1999; Table 2), the efflux rate of [14C]TEA in OCTN2-expressing cells was decreased level comparable with that in Mock cells, confirming that [14C]TEA is exported from the cells via OCTN2. In the presence of 250 μM carnitine, which is a normal urinary level (Leschke et al., 1984), in the extracellular medium, significantly enhanced efflux of [14C]TEA was observed in OCTN2-expressing cells, whereas such an effect was not observed in the Mock cells (data not shown). Addition of 10 mM unlabeled TEA to the extracellular efflux medium enhanced the OCTN2-mediated efflux of [14C]TEA at the initial 1 min, whereas 2 min after initiation of efflux, the efflux rate of [14C]TEA was retarded compared with the control. This result can be explained by assuming that the unlabeled TEA added to the extracellular efflux medium was gradually taken up by the cells and exerted an inhibitory effect on [14C]TEA efflux in the same manner as observed in the case of quinidine. Accordingly, these results suggest that OCTN2-mediated TEA efflux is linked with influx of carnitine, as well as TEA itself, by an exchange transport mechanism.
Inhibitory effects of several endogenous compounds and xenobiotics on carnitine uptake by OCTN2-expressing HEK 293 cells
Pharmacokinetic Analysis of [14C]TEA in Wild-Type andjvs Mice.
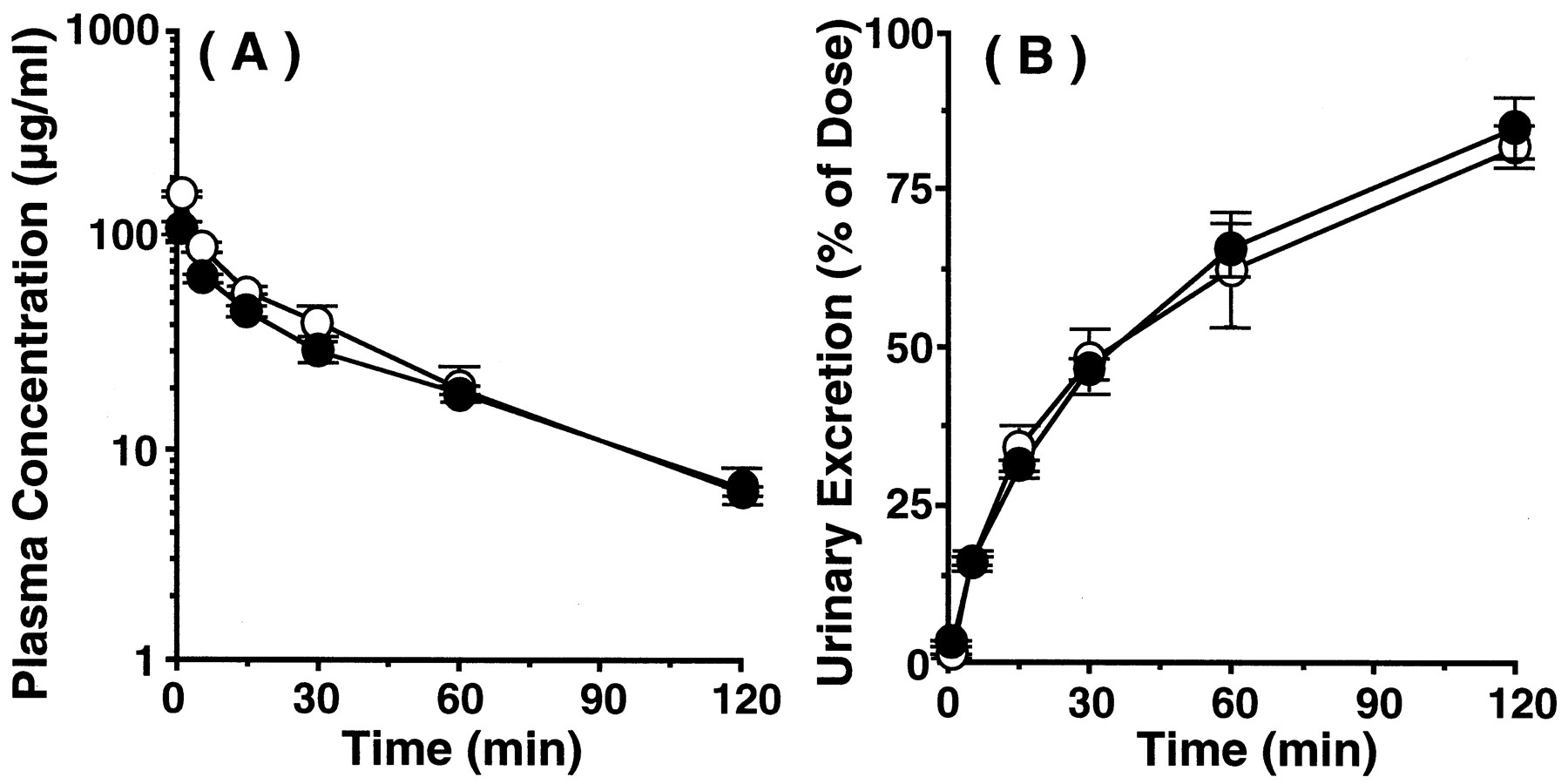
Pharmacokinetic characteristics of TEA after intravenous administration to wild-type and jvs mice were compared. The time profiles of plasma concentration and cumulative urinary excretion of radioactivity of [14C]TEA are shown in Fig. 5, A and B. The radioactivities in plasma and urine can largely be ascribed to intact [14C]TEA, because Mintun et al. (1980) reported that 96% of [14C]TEA was excreted in urine as an intact form after i.v. administration in rats. Marked difference in plasma concentration-time curves at the elimination phase (i.e., 30 min and after) were observed between the two types of mice (Fig. 5A). By the compartment analysis, the rate constant of elimination phase injvs mice was significantly decreased compared with wild-type mice (wild-type, 0.116 ± 0.01 min−1;jvs, 0.059 ± 0.006 min−1;p < 0.05), whereas the rate constants of distribution phase were not changed between wild-type and jvs mice (wild-type, 0.195 ± 0.105 min−1;jvs, 0.229 ± 0.040 min−1,p > 0.05). The difference of the slopes at the linear terminal phase suggested that jvs mice have a lower elimination rate than wild-type mice. The cumulative urinary excretion of radioactivity in jvs mice was significantly lower than that of wild-type mice from 60 min after administration (Fig. 5B). Pharmacokinetic parameters of [14C]TEA were calculated from these results. The AUC from time 0 to infinity forjvs mice (919 ± 86 μg/min/ml) was significantly increased in comparison with that of wild-type mice (553 ± 43 μg/min/ml, p < 0.05). GFR, which was estimated from inulin clearance, and the renal clearance (CLr) of TEA were 9.78 ± 0.86 ml/min/kg and 16.0 ± 0.8 ml/min/kg in wild-type mice and 6.34 ± 0.68 ml/min/kg and 9.77 ± 1.19 ml/min/kg forjvs mice, respectively. Based on these values, the renal secretory clearances (CLs), which represent the renal capability for active secretion of TEA, were evaluated to be 6.22 ± 0.42 ml/min/kg and 3.43 ± 0.77 ml/min/kg for wild-type andjvs mice, respectively, showing a 45% decrease injvs mice compared with wild-type mice. GFR in jvsmice was lower than that in wild mice, so the renal function ofjvs mice was partially impaired. To evaluate the alteration of other epithelial function than OCTN2 in jvs mice, the pharmacokinetic characteristics of cefazolin were evaluated after intravenous administration to wild-type and jvs mice. Cefazolin is an anionic cephalosporin antibiotic that is not metabolized in the body and mainly excreted into the urine (Brogard et al., 1978; Yamazaki et al., 1981; Tsuji et al., 1983). Furthermore, the mechanisms responsible for the excretion of cefazolin are glomerular filtration and active tubular secretion, which is mediated by organic anion transport system in humans and several animals, including mice (Nightingale et al., 1975; Tsuchiya et al., 1978; Kasher et al., 1983;Tsuji et al., 1985; Takano et al., 1989; Hori et al., 1993). The time profiles of plasma concentration and cumulative urinary excretion of intact cefazolin are shown in Figs. 6A and 6B and they were very similar between the two types of mice. Furthermore, pharmacokinetic parameters of cefazolin calculated were comparable without significant differences between wild-type andjvs mice. For example, the AUC from time 0 to infinity for wild-type and jvs mice were 4151 ± 443 μg/min/ml and 3586 ± 216 μg/min/ml, respectively. The CLr and CLs values of cefazolin were 4.39 ± 0.71 ml/min/kg and 2.83 ± 0.49 ml/min/kg, respectively, in wild-type mice and 5.31 ± 0.34 ml/min/kg and 4.41 ± 0.37 ml/min/kg, respectively, forjvs mice. Based on these observations, renal secretory transport activity of organic anions seemed to be maintained injvs mice. Accordingly, the observed decrease in CLs of TEA in jvs mice should be ascribed mainly to the loss of specific secretory activity by OCTN2 for TEA.
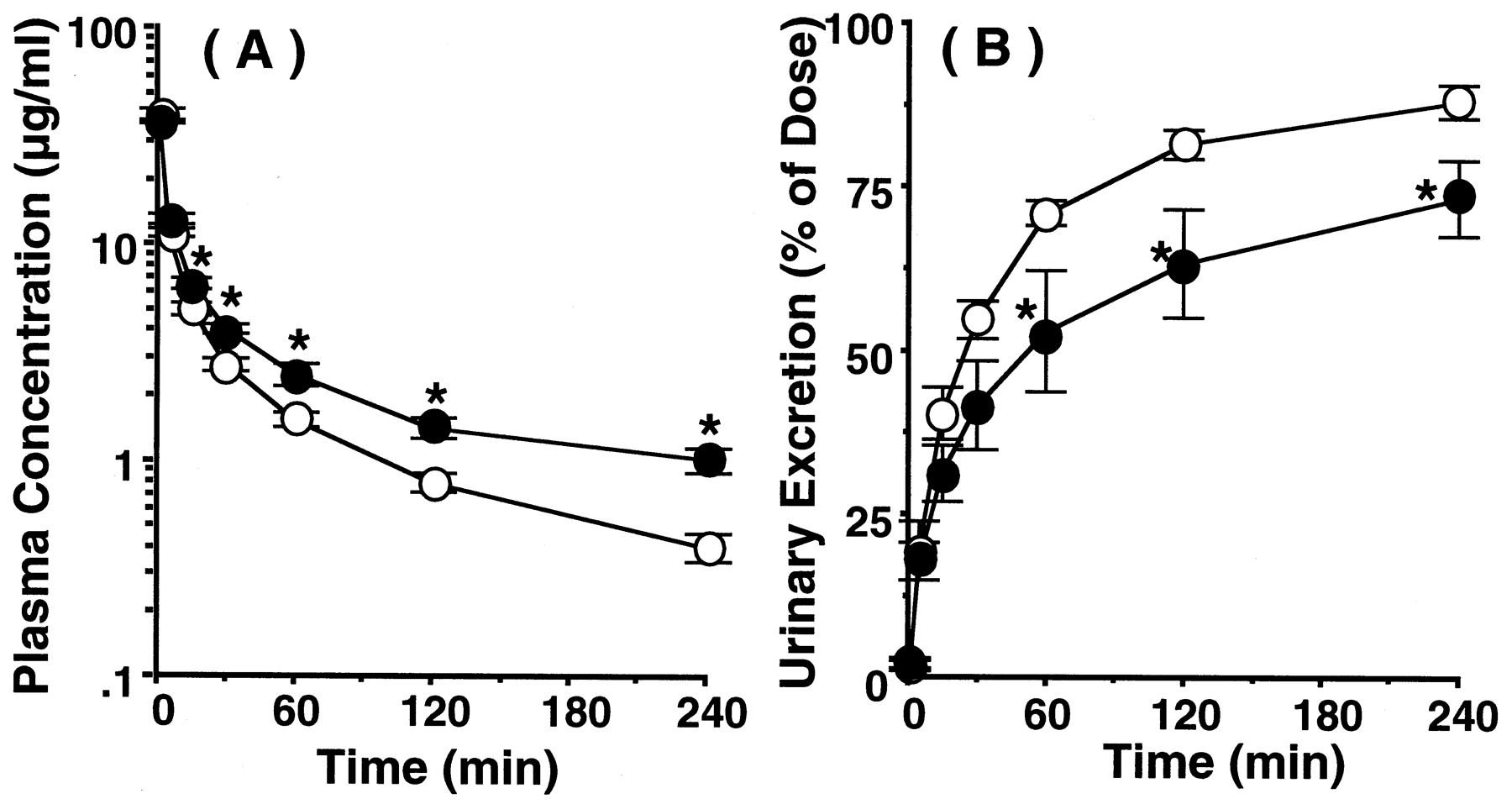
Plasma concentration (A) and cumulative urinary excretion (B) of [14C]TEA after intravenous administration to wild and jvs mice. [14C]TEA was intravenously administered at a dose of 8.8 mg/kg to wild-type (○) and jvs mice (●). *, Significantly different from wild-type mice by Student'st test (p < 0.05). The results are shown as means ± S.E.M. of four to five mice.

Plasma concentration (A) and cumulative urinary excretion (B) of cefazolin after intravenous administration to wild andjvs mice. Cefazolin was intravenously administered at a dose of 20 mg/kg to wild-type (○) and jvs mice (●). The results are shown as means ± S.E.M. of three to four mice.
Tissue Distribution of [14C]TEA in Wild-Type andjvs Mice.
We examined the tissue distribution of TEA in vivo. The values of tissue-to-plasma concentration ratio (K p) of [14C]TEA at 4 h after intravenous administration in wild-type andjvs mice are summarized in Table1 and compared with those of carnitine that have been reported previously (Yokogawa et al., 1999a). TheK p values of TEA in brain, lung, liver, and spleen in jvs mice were significantly lower than those in wild-type mice. In kidney, however, the K pvalue of jvs mice (25.0 ± 4.6) was significantly higher than that of wild-type mice (9.89 ± 1.16). TheK p values of other tissues were not significantly changed between wild-type and jvs mice. The specific increase of K p values in kidney may be explained by the decrease of renal apical secretory transport activity and the decrease of K p values in other tissues may be explained by the decreased tissue uptake activities.
Tissue-to-plasma concentration ratio (K p) of [14C] TEA in wild-type and jvs mice
Inhibition and Transport of Various Compounds in OCTN2-Expressing HEK 293 Cells.
To confirm the specificity of OCTN2, the inhibitory effects of various endogenous compounds and xenobiotics onl-[3H]carnitine uptake by OCTN2-expressing cells were examined (Table2). Endogenous cations such as choline, acetylcholine, serotonin, dopamine, norepinephrine, and thiamine showed significant inhibitory effects (p < 0.05), whereas guanidine, N 1-methylnicotinamide, epinephrine and histamine, and such organic anions as α-ketoglutarate and p-aminohippuric acid were not inhibitory. Furthermore, cationic xenobiotics such as MPP, acetyl-β-methylcholine, pyrilamine, diphenhydramine, procainamide, lidocaine, quinidine, and verapamil demonstrated significant inhibitory effects.
To explore the possible substrates of OCTN2, we measured uptakes of several compounds by OCTN2-expressing cells. Uptakes were measured at 5 or 30 min, depending on the tested compounds. As shown in Table3, [3H]carnitine and its acyl-derivative [3H]acetyl-l-carnitine were good substrates for OCTN2. Although the background uptake observed in Mock cells was rather high, [3H]verapamil, which is a potent inhibitor of carnitine uptake by OCTN2 (Table 2), exhibited significantly increased uptake in OCTN2-expressing cells. [3H]Pyrilamine, a moderate inhibitor of carnitine transport, was also transported by OCTN2. Furthermore, uptake of [3H]MPP, which is a weak inhibitor of OCTN2-mediated carnitine uptake, was slightly but significantly increased, whereas the organic anion [3H]p-aminohippuric acid, which had no inhibitory effect on carnitine transport, was not transported.
Transport of carnitine and various organic cations by OCTN2-expressing HEK 293 cells
Discussion
Cationic xenobiotics and endogenous metabolic products are eliminated through kidney, liver, and intestine by transporters that exhibit broad substrate specificity (Pritchard and Miller, 1993; Oude Elferink et al., 1995; Hunter and Hirst, 1997). In renal epithelial cells, membrane potential-dependent organic cation transporters (OCTs) are functional at basolateral membrane (Zhang et al., 1998; Koepsell et al., 1999; Sweet and Pritchard, 1999), but the apical membrane transporters, which were suggested to be proton or organic cation/organic cation exchange transporter(s), have not been clearly identified. We found that OCTN1, which shows low structural similarity with OCTs, is strongly expressed in kidney and transports cationic compounds (Tamai et al., 1997; Yabuuchi et al., 1999). Subsequently, we (Tamai et al., 1998; Ohashi et al., 1999) and Wu et al. (1998, 1999) have isolated and characterized OCTN2 as an organic cation and high-affinity Na+-dependent carnitine transporter in human and rat. Because OCTNs transport organic cations as well as carnitine, they may contribute in part to the tissue distribution and renal excretion of organic cations (Wu et al., 1998). However, the relevance of OCTNs to organic cation transport remains uncertain, because of the absence of an in vivo evaluation of them. Injvs mice, which show the SCD phenotype (Koizumi et al., 1988), there is a functional defect of OCTN2 owing to a single mutated amino acid residue (L352R) of OCTN2, and the physiological significance of OCTN2 as the carnitine transporter was clearly demonstrated in vivo (Lu et al., 1998; Nezu et al., 1999). In the present study, we studied the role of OCTN2 in the transport of organic cations and the relationship of this with carnitine transport, by means of molecular and in vivo pharmacokinetical analyses using jvs mice.
The uptake and efflux of [14C]TEA by embryonic fibroblasts from jvs mice were decreased compared with those by wild mice (Fig. 1). Furthermore, the mutated OCTN2 found injvs mice lacked transport activity for TEA as well as carnitine (Fig. 2A). These results suggest that OCTN2 transports TEA bidirectionally and that jvs mice are suitable to evaluate the OCTN2-mediated organic cation transport. Because it was reported that P478L and Y211F mutations of human and rat OCTN2 affected carnitine transport without change in TEA transport activity (Seth et al., 1999), the functional site for carnitine on OCTN2 is presumed to be partly shared but not identical with that of TEA. Based on these observations, the region of mutation on OCTN2 of jvs mice (L352R) may be essential in transporting both compounds. Accordingly, we evaluated the significance of OCTN2 as an organic cation transporter by using jvs mice. The disposition of TEA is significantly altered in jvs mice (Fig. 5, A and B; Table 1). The plasma concentration of [14C]TEA in jvsmice was higher and urinary excretion was delayed compared with those of wild-type mice. In jvs mice, the GFR estimated in terms of the clearance of [3H]inulin, which was administered simultaneously with [14C]TEA, was also lower than that of wild-type mice. These results indicated that renal function (glomerular filtration and/or renal blood flow) injvs mice may be impaired nonspecifically. One reason for the decrease of GFR in jvs mice might be severe cardiomyopathy (Horiuchi et al., 1993). Recently, it was demonstrated that the rat multispecific organic anion transporter OAT1 mediates the renal secretion of β-lactam antibiotics, including cefazolin, at the renal basolateral membrane (Jariyawat et al., 1999). Furthermore, mouse and human type-I inorganic phosphate/organic anion transporters are responsible for the urinary secretion of many organic anions such as cefazolin at the renal apical membrane (Uchino et al., 2000a,b). So, we examined the effect of cefazolin on the in vitro transport of carnitine by wild-type and jvs mice fibroblasts and observed no effect of cefazolin on carnitine uptake (data not shown). In addition, in the present study, the secretory clearance of cefazolin examined in wild-type and jvs mice was maintained (Fig. 6, A and B). However the renal secretory clearance of [14C]TEA was significantly low injvs mice (55% of wild-type mice). So, the decrease in CLs of [14C]TEA in jvs mice is thought to be specific. Furthermore, the increase ofK p value of TEA was observed only in kidney (Table 1), which is in contrast to the previous observation that theK p value of [3H]carnitine in kidney of jvs mice was lower than that of wild-type mice (Yokogawa et al., 1999a). In contrast, K p values of [14C]TEA in other tissues were decreased injvs mice compared with those in wild-type mice (Table 1), which is similar to the observation in [3H]carnitine distribution (Yokogawa et al., 1999). Accordingly, the high K p value of [14C]TEA in jvs-mice kidney is specific and can be explained by an increase of accumulation owing to the decrease in secretory transport across the apical membrane. From these results, the apparent decrease of excretion rate of [14C]TEA into urine can be ascribed to the specific decrease of renal secretory transport of [14C]TEA in addition to that of GFR and/or renal blood flow. Furthermore, efflux of [14C]TEA from the OCTN2-expressing cells was accelerated in the presence of extracellular TEA, suggesting an exchange transport mechanism. Accordingly, OCTN2 might be one of the organic cation transporters expressed in the renal apical membrane, possibly an organic cation/organic cation exchanger.
As shown in Table 1, the K p values of [14C]TEA in several tissues such as brain, lung, liver and spleen in jvs mice were lower than those in wild-type mice, as observed previously in our carnitine study (Yokogawa et al., 1999a). However, no significant difference inK p values of [14C]inulin, which cannot cross the membrane easily and is a marker of vascular and interstitial fluids, was observed between wild-type and jvs mice in most tissues (data not shown). The alteration of K pvalues in heart, brain and lung is apparently correlated with the tissue expression profile of OCTN2 (Tamai et al., 2000). This implies that the OCTN2 contributes at least in part to distribution of TEA in these tissues. The K p values of [3H]carnitine in the kidney and liver injvs mice were significantly lower than that of wild-type mice (Yokogawa et al., 1999a) and jvs mice lack in ability of reabsorption of carnitine in kidney (Horiuchi et al., 1994). So, the finding suggested that OCTN2 is located on the brush-border membrane in kidney and on the basolateral (sinusoidal) membrane in liver. Therefore, OCTN2 contributes greatly to the hepatic uptake of [14C]TEA from blood, which is in accordance with our previous observation in hepatic carnitine transport (Yokogawa et al., 1999b,c). Here, however, the K pvalues of [14C]TEA in lung, heart, liver, kidney, spleen, and gut in jvs mice are unity or above, so TEA is likely to be concentrated by an active transport system(s) other than OCTN2 (Koepsell et al., 1999). Furthermore, alteration ofK p values of [14C]TEA was not necessarily correlated with that of [3H]carnitine in a few tissues (Table1). So, other transporters, such as OCT family and OCTN1, which accept TEA as substrate, may greatly contribute to the distribution of organic cations in these tissues.
Another point of interest is the possible coupling of carnitine influx and TEA efflux. Efflux of TEA from the cells was enhanced by expression of OCTN2, demonstrating that TEA is bidirectionally transported by OCTN2, which is in accordance with the observation in wild-type fibroblasts. The reduced efflux in the presence of quinidine confirmed that TEA efflux is mediated by OCTN2, because quinidine is a strong inhibitor of OCTN2 (Table 2) and is taken up efficiently even by Mock cells (Yabuuchi et al., 1999). In the present study, OCTN2-mediated efflux of TEA was enhanced in the presence of extracellular carnitine. Furthermore, Na+-dependent carnitine uptake by the OCTN2-expressing cells was stimulated by preloaded TEA in a concentration-dependent manner (Figs. 4, A-C). These observations indicate that the influx of carnitine and the efflux of TEA are linked and the coupling enhances the two transport processes. As shown in Fig.2B, carnitine transport via OCTN2 requires Na+, whereas TEA transport does not, and Na+ affects the affinity for OCTN2 (Fig. 3). From these observations, the molecular mechanism of carnitine/TEA transport via OCTN2 at the renal apical membrane was postulated as shown in Fig.7. Because the luminal side of the membrane is rich in Na+, carnitine is taken up efficiently by the epithelial cells via OCTN2. When the transporting site is oriented from the outer to the inner cellular domain by transporting carnitine, the site will be inactivated for carnitine transport because of the inside-directed Na+gradient (intracellular low concentration of Na+). So, TEA, which is accumulated from the blood by basolateral transporter OCTs, can be a better substrate and is transported out of the cells apparently via an exchange mechanism between carnitine and TEA. So, the difference of Na+ dependence between carnitine and TEA transports determines the direction under physiological circumstances. A similar finding was obtained for the y+L-type amino acid transport system, which exchanges intracellular cationic amino acid with extracellular neutral amino acid in an Na+-dependent manner at the renal epithelial cells (Chillaron et al., 1996; Pfeiffer et al., 1999).

Proposed model of the transport mechanisms of carnitine and organic cations by OCTN2. After binding of Na+ to the outer surface of OCTN2, the carnitine binding site is activated as shown by the change of the shaded triangle (1) to a square (2) and carnitine is transported into the inner side of the membrane (3 and 4). Because of the presence of the inward-directed Na+ gradient, the affinity of carnitine for OCTN2 is reduced on the inner membrane side, and TEA binds to OCTN2 (5 and 6). Subsequently, TEA is transported out of the cells. TEA is transported into cells from the outside in the absence of Na+. This model explains the Na+ dependence and independence of carnitine and TEA transport, respectively. Furthermore, the two substrates are transported via an exchange mechanism and exhibit a mutual trans-stimulation effect. When the transporter is loaded with carnitine or TEA, it functions more efficiently than the unloaded form, resulting in a trans-stimulation effect. Because carnitine is a zwitterion and TEA is a cation, the anionic moiety of carnitine may require a specific site, which is not involved in TEA binding, on OCTN2. The Na+ binding site must be specific to carnitine and was therefore assumed to be located near the binding site of carboxyl group of carnitine, which should not overlap with the TEA binding site.
As summarized in Table 2, various cationic compounds inhibit carnitine transport by OCTN2. In addition, some of them are transported by OCTN2 (Table 3). Interestingly, among them, carnitine and its structural analog, acetylcarnitine, are transported in a Na+-dependent manner, whereas cationic compounds such as TEA show negligible Na+ dependence (Tamai et al., 1998; Wu et al., 1998, 1999; Ohashi et al., 1999). So, those compounds that do not require Na+ may be exchanged with carnitine, as observed in the case of TEA efflux, suggesting that reabsorption of carnitine is accompanied with accelerated elimination of organic cations and that the secretory transport of organic cations via OCTN2 energizes reabsorption of carnitine. The circulating organic cations are concentrated in the epithelial cells by the basolaterally localized OCT transporter and are subsequently secreted into urine across the apical membrane by OCTN2 as one of the apical-type organic cation transporters in the renal tubular epithelial cells.
In conclusion, an involvement of OCTN2 in the tissue distribution and renal secretion of organic cations was demonstrated by using OCTN2-expressing cells and jvs mice. Furthermore, molecular mechanism of multifunctionality of OCTN2 was proposed by hypothesizing an exchange transport of Na+-dependent carnitine reabsorption and Na+-independent secretion of organic cations.
Acknowledgments
We thank Dr. H. Uchino and Mr. Y. Yamaguchi for helpful discussions and technical assistance.
Footnotes
- Received July 7, 2000.
- Accepted November 11, 2000.
-
Send reprint requests to: Prof. Akira Tsuji, Ph.D., Department of Pharmacobio-Dynamics, Faculty of Pharmaceutical Sciences, Kanazawa University, 13–1 Takara-machi, Kanazawa 920-0934, Japan. E-mail: tsuji{at}kenroku.kanazawa-u.ac.jp
-
This study was partly supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports and Culture, Japan.
Abbreviations
- TEA
- tetraethylammonium
- jvs
- juvenile visceral steatosis
- SCD
- systemic carnitine deficiency
- MPP
- 1-methyl-4-phenylpyridinium
- HEK
- human embryonic kidney
- NMG
- N-methylglucamine
- GFR
- glomerular filtration rate
- AUC
- area under plasma concentration-time curve
- CLr
- renal clearance
- CLs
- renal secretory clearance
- Kp
- tissue-to-plasma concentration ratio
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}