


Abstract
Under reducing conditions of SDS-polyacrylamide gel electrophoresis, the CB1 receptor exists in its monomeric form as well as in an SDS-resistant high molecular weight form that appears to be devoid of G proteins. The CB1 cannabinoid receptor was immunoprecipitated from 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate-solubilized rat brain membranes using an antibody against the CB1receptor N terminus. The CB1 receptor was coimmunoprecipitated with its associated G proteins, specifically those of the Gαi/o family, but not Gαs, Gαq, or Gαz. The CB1receptor-Gαi/o complex existed in the absence of exogenous agonists, and the cannabinoid receptor agonist desacetyllevonantradol failed to alter the stoichiometry of the receptor-Gαi/o interaction. Guanosine-5′-O-(3-thio)triphosphate could disrupt the interaction. A peptide derived from the CB1 receptor juxtamembrane C-terminal domain, peptide CB1401-417, autonomously activates Gi/o proteins. Peptide CB1401-417 competitively disrupted the CB1receptor association with Gαo and Gαi3 but not Gαi1 or Gαi2. This G protein specificity was also observed in detergent extracts from membranes of the frontal cortex, striatum, and cerebellum. Alternative peptides, including peptides from the CB1 receptor third intracellular loop and the G protein activating peptide mastoparan-7, failed to promote uncoupling from Gαo. A CB2receptor juxtamembrane C-terminal peptide failed to disrupt the CB1 receptor-Gαo complex. These studies illustrate that the CB1 receptor can exist as an SDS-resistant multimer. In 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate detergent, the CB1 receptor exists in a complex with G proteins of the Gi/o family in the absence of exogenous agonists. Furthermore, this study provides the first description of domain specificity for interaction with a selective set of G proteins.
Δ9-Tetrahydrocannabinol, the major active compound of Cannabis species, and other cannabimimetic compounds (cannabinoids, aminoalkylindoles, and eicosanoid ethanolamides and esters) exert their biological actions on the nervous system by signaling through the G protein-coupled CB1 cannabinoid receptors (reviewed in Howlett, 1995). The CB1 receptor was found to be expressed predominantly in brain (Herkenham et al., 1991; Matsuda et al., 1993). The CB1 receptor signals through several biological mechanisms, including inhibition of adenylyl cyclase, modulation of N-type Ca2+ channels (see Howlett, 1995, for original references), activation of mitogen-activated protein kinase (MAPK), and expression of immediate early genes such askrox-24 (Bouaboula et al., 1997). A second subtype, the CB2 receptor, has been identified primarily in cells of immune origin (Galiegue et al., 1995). The CB2 receptor expressed in host cells has been shown to signal via inhibition of adenylyl cyclase (Felder et al., 1995) and activation of MAPK and expression of immediate early genes such as krox-24 (Bouaboula et al., 1996) but does not appear to modulate ion channels in host cells (Felder et al., 1995).
The importance of different domains of G protein-coupled receptors for interaction with G proteins has been established for many receptors (see Wess, 1998, for review). Recent studies from our laboratory have established that a CB1 receptor peptide fragment from the C-terminal juxtamembrane region, termed CB1401-417 (Table1), could autonomously activate Gi/o proteins in a dose-dependent fashion (Howlett et al., 1998; Mukhopadhyay et al., 1999). Gi/o activation could be detected as both activation of [35S]guanosine-5′-O-(3-thio)triphosphate ([35S]GTPγS) binding to G proteins in rat brain membranes and inhibition of adenylyl cyclase in neuronal cell membranes (Howlett et al., 1998). The activation of Gi by CB1401-417 was observed in cells that did not express CB1receptors and was blocked by pretreatment with pertussis toxin (Mukhopadhyay et al., 1999). These studies suggest that CB1401-417 represents a domain on the receptor that contributes to the interaction of the CB1receptor with Gi/o proteins. From these observations, we hypothesized that CB1401-417 directly interacts with Gi/o proteins, resulting in activation of the G proteins. This suggests that the C-terminal juxtamembrane region of the CB1 receptor serves as the Gi/o activation domain.
CB1 and CB2 cannabinoid receptor peptide fragments
Several approaches have been used to characterize the receptor-G protein coupling in various other G protein-coupled receptors. Immunoprecipitation procedures have been used successfully to characterize receptor-G protein association in the presence and absence of agonist treatment for G protein-coupled receptors (Matesic et al., 1991; Okuma and Reisine, 1992; Damaj et al., 1996; Gu and Schonbrunn, 1997). In the present communication, we used a similar approach to investigate the importance of the juxtamembrane C-terminal domain of the CB1 cannabinoid receptor for interaction with G proteins. We report here the conditions for immunoprecipitation of the CB1 receptor in association with Gα subtypes of the Gi/o family from 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate (CHAPS) detergent extracts. Using this methodology, we show that selective CB1 receptor-Gα interaction can be competitively disrupted by the C-terminal juxtamembrane fragment of the receptor. These studies illustrate that the CB1receptor can exist as a stable multimer in a complex with G proteins of the Gi/o family. The CB1receptor C-terminal juxtamembrane domain is important for interaction with selective G proteins within this family, suggesting the possibility for selectivity in G protein-mediated signaling pathways.
Experimental Procedures
Materials.
The standard chemicals used were of the highest grade and were purchased from Sigma Chemical Co. (St. Louis, MO) unless otherwise mentioned. Desacetyllevonantradol (DALN) was a gift from Pfizer, Inc. (Groton, CT). SR141716A was purchased from BIOMOL (Plymouth Meeting, PA). Mastoparan-7 was purchased from Research Biochemicals Inc. (Natick, MA). Urea was purchased from ICN (Costa Mesa, CA). SDS, acrylamide, bisacrylamide, ammonium persulfate, and polyvinylidine difluoride membranes were obtained from Bio-Rad (Hercules, CA). Anti-Gαo, a mouse monoclonal antibody against partially purified bovine brain Gαo protein, and anti-Gαi3, an affinity-purified rabbit antisera against a peptide specific for Gαi3, were purchased from BIOMOL. Anti-Gαi1/2, an affinity-purified rabbit antisera against the C-terminal peptide that is common to both Gαi1 and Gαi2, was purchased from Calbiochem (San Diego, CA). Rabbit antisera against peptides selective for either Gαi1 (3646) or Gαi2(1521) were generously provided by Dr. D. Manning, University of Pennsylvania (Williams et al., 1990). Affinity-purified rabbit polyclonal antibodies against Gβ1-4 were T-20, which was purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA), and BN1, which was a gift from Dr. N. Gautam, Washington University (Pronin and Gautam, 1994). Affinity-purified rabbit polyclonal antibodies against Gγ2 (BG) and Gγ (CG) were donated by Dr. N. Gautam (Pronin and Gautam, 1994). Anti-rabbit and anti-mouse IgG-horseradish peroxidase were from Jackson ImmunoResearch Laboratories (West Grove, PA). Rainbow molecular weight markers, enhanced chemiluminescence (ECL) reagents, and film were purchased from Amersham Life Sciences, Inc. (Arlington Heights, IL). Peptides CB1174-188, CB1301-317, and CB2302-320 were synthesized at Saint Louis University Peptide Facility, and other peptides were synthesized and purified by Princeton Biomolecules (Columbus, OH) (Howlett et al., 1998).
CB1 Receptor Antibody and Affinity Matrix Preparation.
Rabbit polyclonal antibodies were raised against the N-terminal 14 amino acids of the CB1 receptor as described previously (Howlett et al., 1998). Anti-CB11-14 was affinity purified using CB11-14 as the affinity ligand attached to agarose matrix according to the SulfoLink Immobilization procedure (Pierce, Rockford, IL). An affinity resin for the CB1 cannabinoid receptor was prepared by coupling affinity-purified anti-CB11-14 to Affi-Prep-Hz matrix (Bio-Rad) according to the manufacturer's instructions. This method binds periodate-oxidized carbohydrate moieties on the antibody heavy chain to hydrazide-activated methacrylate matrix (O'Shannessy and Hofman, 1987).
Membrane Solubilization and Immunoprecipitation.
Rat brain P2 membranes were prepared as described previously (Howlett et al., 1998). CHAPS solubilization of the membranes were carried out according to the method of Houston and Howlett (1993). For immunoprecipitation of the CB1 receptor and associated proteins, an aliquot of CHAPS-solubilized receptor from rat P2 membranes (100 μg protein) was incubated with 20 μl of anti-CB11-14-affinity matrix for 4 h at 4°C plus an additional 1.5 h at room temperature under constant rotation. The anti-CB11-14-affinity matrix was then sedimented at 14,000 rpm for 5 min and washed three times with TBS buffer (Tris · HCl, pH 7.4, 140 mM NaCl) containing 0.1% Tween 20. Immunoprecipitated protein was eluted from the matrix by mixing with Gly-HCl, pH 2.5 (100 mM), and the eluate was immediately neutralized with Tris · HCl, pH 8.0 (1.5 M). The protein from the neutralized eluate was precipitated by addition of 8 volumes of CHCl3/CH3OH/H2O (1:4:3), dissolved in Laemmli's sample buffer containing 5 mM EDTA, and heated at 65°C for 5 min. Samples were subjected to polyacrylamide gel electrophoresis (PAGE) on 10% polyacrylamide-0.1% SDS-6 M urea gels.
Western Immunoblot Analysis.
Electrophoretic transfer of proteins from the gel to polyvinylidene difluoride membranes was carried out in 10 mM 3-[cyclohexylamino]-1-propanesulfonic acid (CAPS) buffer with 0.01% SDS, pH 11, for 16 h (0–4°C) at 20 V using a Bio-Rad Trans-Blot Cell equipped with a cooling coil. Blots were rinsed with TBS buffer and incubated with blocking buffer (5% nonfat dry milk plus 5% normal goat serum in TBS) at room temperature for 1 h to eliminate nonspecific binding. Blots were then incubated with affinity-purified anti-CB11-14 combined with the indicated anti-Gα antibody (1:1000) in blocking buffer for 90 min at room temperature, followed by washing three times with TBS containing 0.1% Tween 20. Control experiments were performed using separate incubations with individual antibodies, and the results were the same as experiments stained with combined antibodies. In Fig.1, no bands are observed in the molecular weight range of the Gα proteins when staining was performed with affinity-purified anti-CB11-14. Thus, for studies shown in this series of experiments (except Figs. 1 and 7B), the costaining procedure was used. Blots were incubated with horseradish peroxidase-coupled anti-rabbit and anti-mouse IgG sequentially for 1 h at room temperature, followed by one rinse with TBS, seven rinses with TBS-Tween 20, and four rinses with water. Immunoreactive bands were detected by ECL. Densitometric scanning was analyzed using a modified version (version 1.59) of the National Institutes of Health Image Program (Scion Corp.).

Effects of agonists and antagonist on CB1cannabinoid receptor bands on Western blots. Rat brain membranes were incubated with the indicated ligands or vehicle for 1 h at 30°C in 20 mM Tris · HCl, pH 7.4; 3 mM MgCl2, and 1 mM EDTA. After centrifugation for 20 min at 20,000g, the pellets were solubilized with 8 M urea/5% SDS/1% β-mercaptoethanol/0.1 mM EDTA sample buffer, heated at 37°C for 10 min, and applied to SDS-10% polyacrylamide gels (Blumer et al., 1988). Western blots were stained with anti-CB11-14. Molecular weight markers indicated by dashes at the left of the blot are 10, 15, 30, 35, 50, 75, 105, 160, and 250 kDa. Lanes on blots show membranes treated with 1, vehicle; 2, 1 μM DALN; 3, 1 μM WIN 55122–2; 4, 1 μM anandamide; and 5, 1 μM SR141716A. The bands from four individual blots were scanned, and the ratios of the densities of the low molecular weight to high molecular weight bands were (mean ± S.E.) 1, 1.8 ± 0.1; 2, 2.3 ± 0.8; 3, 2.4 ± 0.2; 4, 2.0 ± 0.6; and 5, 2.0 ± 0.5. No significant differences were found using ANOVA.

Association of the CB1 receptor and Gαo or Gαi1/2 subtypes from different regions of rat brain and the susceptibility to disruption by peptide CB1401-417. A and B contain immunoprecipitates from CHAPS extracts of membranes (100 μg protein) from lanes 1 and 2, rat frontal cortex (FC); lanes 3 and 4, rat striatum (ST); and lanes 5 and 6, rat cerebellum (CB). Lanes 1, 3, and 5, CHAPS extract that had been incubated at 30°C for 30 min with peptide CB1401-417 (1 mM) before immunoprecipitation. Lanes 2, 4, and 6, vehicle controls. A, immunoblot was stained with anti-CB11-14 plus anti-Gαo. B, blot in A was treated with H2O2 (15% for 30 min at room temperature) and washed twice with TBS-Tween buffer and once with TBS buffer to inactivate the horseradish peroxidase conjugate (TechTip #120; Amersham Life Science). The blot was restained with anti-Gαi1/2. This experiment is representative of three studies with similar results.
Results
Monomer and Multimer Forms of CB1 Cannabinoid Receptor.
The CB1 cannabinoid receptor typically exhibits a pattern of two bands on Western blot analysis of rat brain P2 membrane preparations (Fig. 1). Affinity-purified anti-CB11-14 recognized a band at 64 kDa and a second band between 160 and 200 kDa. For rat brain membrane preparations, the higher apparent molecular weight band was frequently more pronounced (C. Song, H. McIntosh, and S. Mukhopadhyay, unpublished observations). This complex represents a protein-protein interaction that is stable to SDS, urea, and sulfhydril reagents used in the SDS/urea/PAGE procedures. Some conditions that appeared to enhance the CB1 cannabinoid receptor monomer form were the addition of 5 mM EDTA to the sample buffer, solubilization of the membranes in sample buffer rather than diluting the membrane suspensions with 2× sample buffer, heating the membranes in sample buffer for 5 min at 60°C rather than for 5 min at 100°C or 20 min at 60°C, use of 20% sucrose in sample buffer without glycerol, 6.5 M urea in SDS sample buffer, and use of a higher concentration of SDS in the sample buffer or running buffer (data not shown). Conditions that made no difference in augmenting the monomer form were presence of 5% β-mercaptoethanol, sonication of the samples in sample buffer rather than heating at 60°C, treatment for 30 min with alkaline phosphatase in 50 mM Tris · HCl, pH 8.0, and digestion of membranes for 18 h with N-glycosidase. Acid phosphatase treatment appeared to augment the monomer form relative to the high molecular weight aggregate; however, the procedure resulted in significant loss of protein from both bands, making quantification uninterpretable.
For other G protein-coupled receptors, evidence exists that agonist binding may influence the receptor monomer/multimer equilibrium (Ciruela et al., 1995; Hebert et al., 1996; Cvejic and Devi, 1997). For A1 adenosine receptors, treatment of membranes with nonhydrolyzable GTP analogs, N-ethylmaleimide, or pertussis toxin failed to alter the fraction of the receptor in the dimeric state; however, treatment of membranes with agonists before solubilization in SDS resulted in loss of the dimeric form (Ciruela et al., 1995). An agonist influence on monomer/multimer ratio for the CB1 receptor was not observed in the present study (Fig. 1). Membranes that had been treated in the presence of the cannabinoid agonist DALN, the aminoalkylindole agonist WIN-55212-2, or the eicosanoid agonist anandamide failed to exhibit a different pattern of banding on Western analysis compared with control membranes. Furthermore, preincubation in the presence of the CB1 receptor antagonist SR141716A failed to alter the banding pattern. Neither pertussis toxin treatment nor incubation of CHAPS extracts with agonists, antagonist, or GTPγS altered the fraction of receptors in the high-molecular-weight band (S. Mukhopadhyay, unpublished observations). This observation is consistent with findings of Ciruela et al. (1995), who noted that the addition of agonists to the solubilized extract rather than membranes failed to deplete the amount of dimer.
The composition of the high molecular weight form of the CB1 receptor is not known. Western blot analysis indicates that antibodies against Gαo, Gαi1, Gαi2, Gβ1/2/3/4 (two different antibodies), Gγ2, or Gγ3 failed to detect immunoreactivity at this high molecular weight position (data not shown). These findings suggest that the high molecular weight band is not a receptor-G protein complex, nor is it a complex of receptor with any one of the G protein subunits.
Coimmunoprecipitation of Gα Proteins with CB1Receptor.
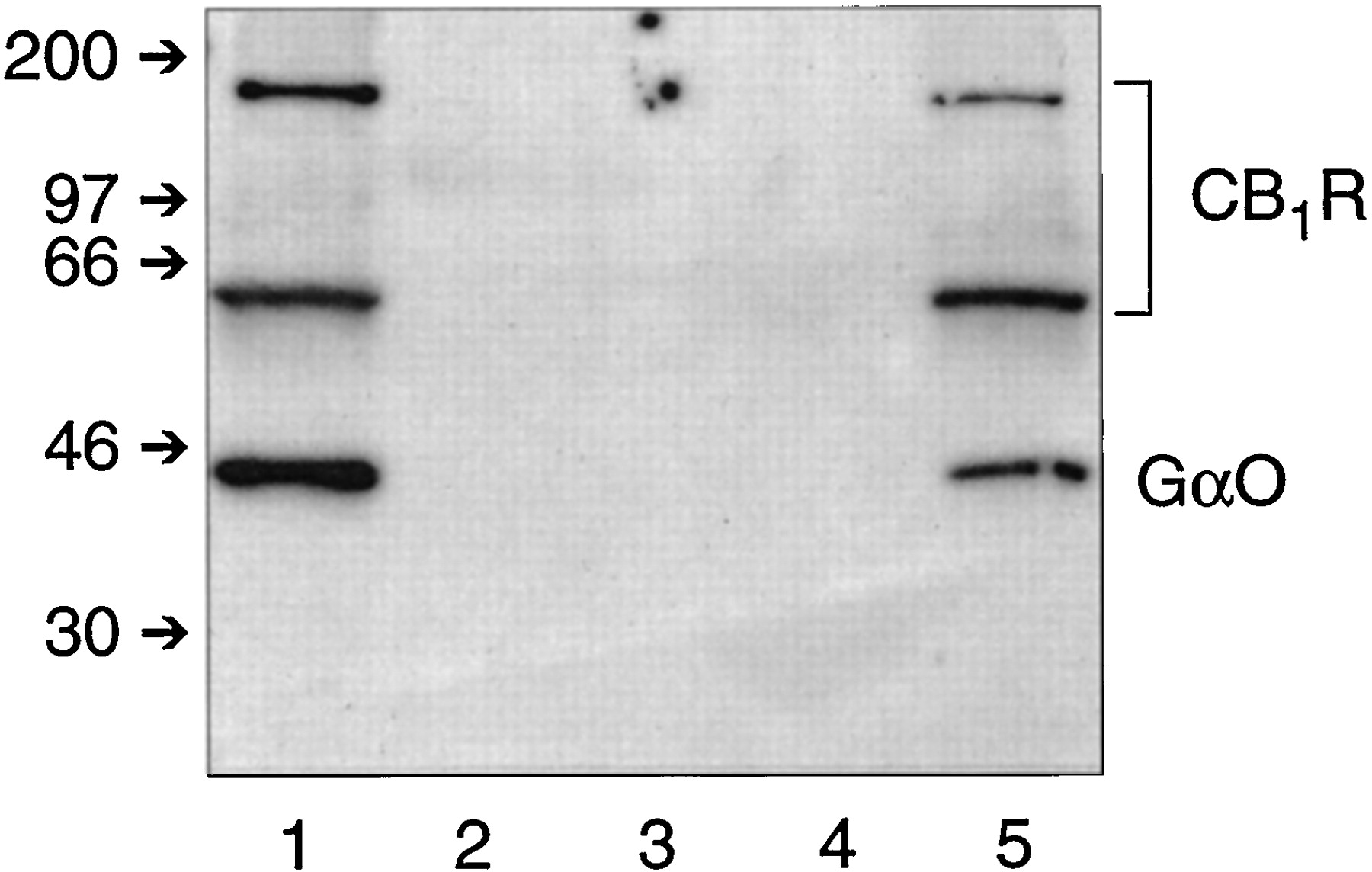
Matrix-bound anti-CB11-14 was able to immunoprecipitate the CB1 receptor in association with Gαo from CHAPS-solubilized rat brain membranes. Figure 2 demonstrates the feasibility of this coimmunoprecipitation method for resolving G proteins that are associated with the CB1receptor. Lane 1, containing CHAPS extract from rat brain membranes, shows the CB1 receptor as bands at 64 kDa and approximately 200 kDa, and αo detected at 40 kDa. The CHAPS extract was incubated with matrix-bound anti-CB11-14. After removing the unbound proteins followed by several washing steps, no leaching of CB1 receptor or αo was observed, as shown for the second and third washes of the affinity matrix in Fig.2, lanes 2 and 3, respectively. Matrix-bound protein that was eluted at low pH exhibited a nearly quantitative recovery of CB1 receptor (lane 5). Somewhat less of the high molecular weight form was recovered from the affinity matrix. A significant fraction of the αo was found to be associated with the CB1 receptor and appeared in the eluate as the CB1 receptor was dislodged from the affinity matrix. Coomassie staining of the gel showed that no extraneous bands were observed (data not shown). No detectable bands were observed from an eluate of CHAPS extract under mock immunoprecipitation conditions using matrix devoid of anti-CB11-14 (lane 4), indicating that nonspecific adherence of CB1receptor protein or Gαo to the matrix material did not occur during the procedure. Successful coimmunoprecipitation of Gαo with the CB1 receptor using a CB1 receptor antibody supports the idea that significant amounts of CB1 receptor remain precoupled with G proteins even in the absence of exogenous agonists. Because the CB1 receptor is detected as two bands, receptor content is quantified in subsequent studies by summing both band densities detected by ECL and densitometric scanning.
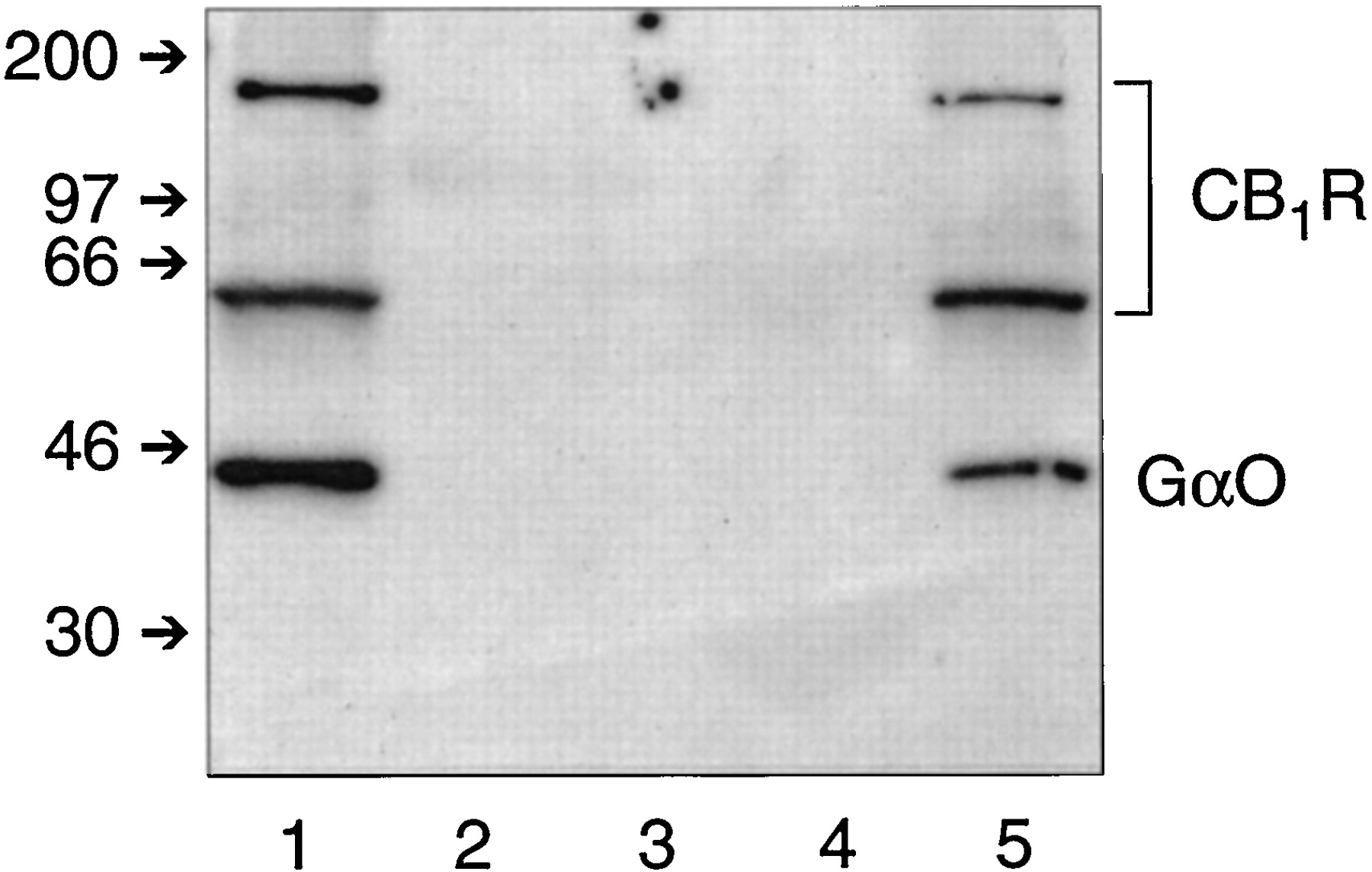
Coimmunoprecipitation of the CB1receptor-Gαo complex from CHAPS extracts of rat brain membranes. Western analysis was carried out using anti-CB11-14 and anti-Gαo as described in the text. Lane 1, CHAPS extract from rat brain membranes (100 μg protein) that was applied to the CB1 receptor affinity matrix; lanes 2 and 3, the second and third washes of the affinity matrix, respectively; lane 4, eluate after a mock immunoprecipitation with AffiPrep matrix that was not coupled to anti-CB11-14; and lane 5, protein eluted from the affinity matrix. Small arrows on the left indicate molecular weight markers (kDa). This experiment is representative of seven studies with similar results.
Immunoprecipitates of the CB1 receptor from the CHAPS extracts of rat brain exhibited a strong Gαo signal on Western analysis (Fig.3, lane 1). In some experiments, we could resolve two isoforms of Gαo in the SDS-urea PAGE system (see Fig. 7). CB1 receptor also coimmunoprecipitated with Gαi1 and Gαi2 (Figs. 3, lane 2, and 6, A and B) and Gαi3 (Fig. 3, lane 3). The commercially available antibody to Gαi1/2 detected a doublet in the range of Gα proteins as well as a nonspecific minor band at a lower apparent molecular weight. Studies using Dr. Manning's antibodies that were specific for Gαi1 or Gαi2 identified the higher and lower bands of the doublet as Gαi1 and Gαi2, respectively, and the minor band was not observed with these antibodies. In comparable experimental conditions, the CB1 receptor was not able to coimmunoprecipate with Gαs, Gαq or Gαz(data not shown). Thus, all three Gαi subtypes and both isoforms of Gαo remain precoupled to the CB1 receptor in detergent extracts. However, Gs, which stimulates adenylyl cyclase, and Gq, which mediates regulation of phospholipase Cβ, failed to associate with the CB1 receptor.
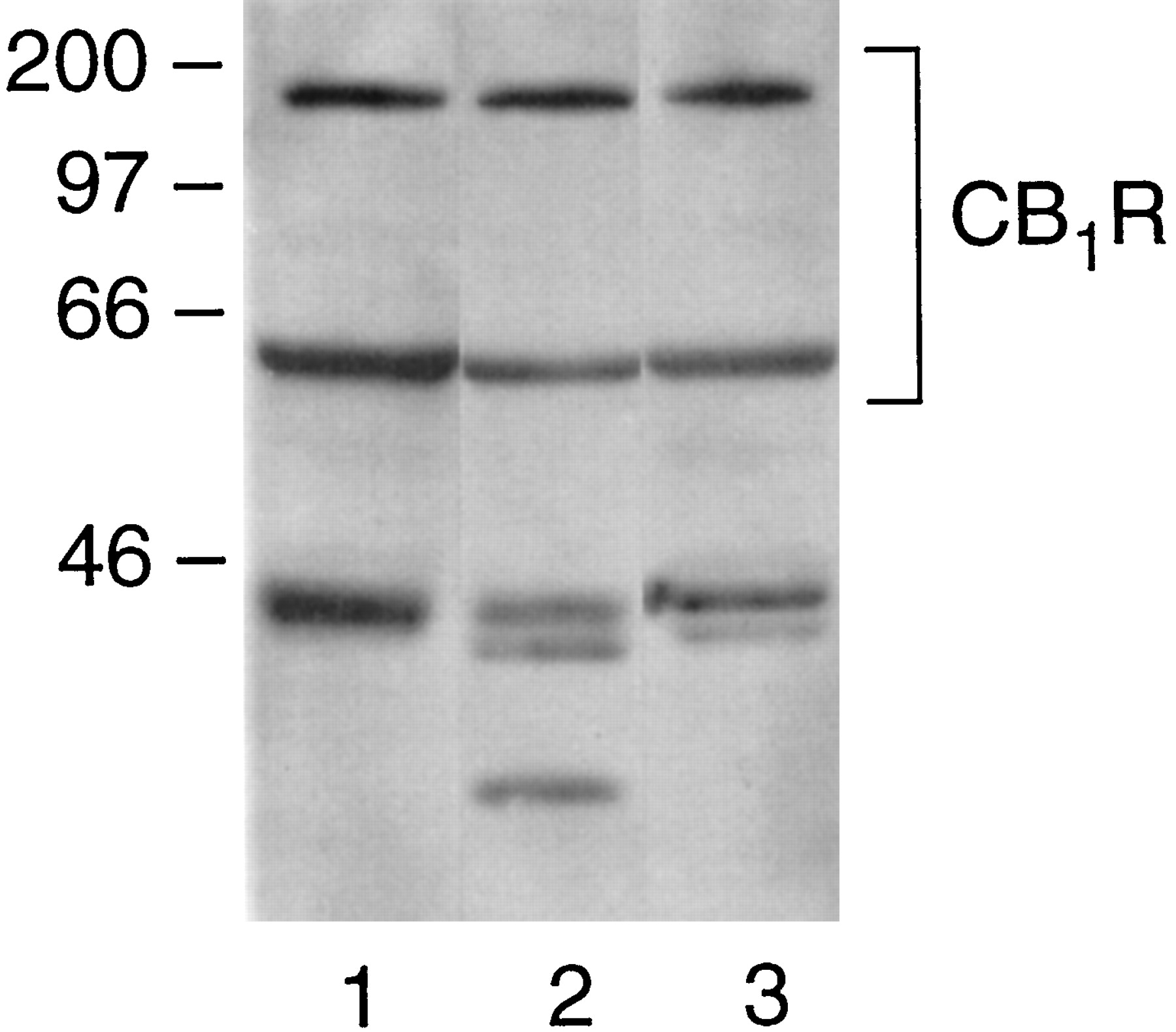
Gα subtypes coimmunoprecipitated with the CB1 receptor from CHAPS-solubilized rat brain membranes. Each lane was loaded with immunoprecipitated protein from a CHAPS extract of rat brain membranes (100 μg protein), and Western blot analysis was performed. Lanes were stained with anti-CB11-14 plus the following antibodies: lane 1, anti-Gαo; lane 2, anti-Gαi1/2; and lane 3, anti-Gαi3. This experiment is representative of 10 studies with similar results.
GTP Analogs and Agonists Can Disrupt CB1 Receptor-G Protein Complex.
To determine the effect of agonist and antagonist binding on the coupling of the CB1 receptor to G proteins, CHAPS-solubilized rat brain membranes were treated with the CB1 receptor agonist DALN in the presence or absence of the nonhydrolyzable guanine nucleotide analog GTPγS. After equilibration, immunoprecipitation and Western analyses were performed. As shown in Fig. 4, agonist alone failed to cause any significant alteration in the CB1receptor-Gαo complex with respect to control (lane 2 compared with lane 1). In the presence of GTPγS, DALN caused a slight decrease in the Gαo band density (lane 3). GTPγS alone caused a significant dissociation of the Gαo subunit from the CB1receptor as evident from the pronounced decrease in Gαo signal in the Western blot (lane 4). The CB1 receptor antagonist SR141716A failed to cause any significant alteration in the band density of Gαo associated with the CB1 receptor. No changes in the densities of the CB1 receptor bands were observed after any of these treatments. These findings demonstrate that in CHAPS solution, the CB1 receptor-Gαocomplex exists in a form that can dissociate when GTP or its analogs bind to the Gαo protein. The nonhydrolyzable GTP analog stabilizes free Gαo by slowing the reassociation with the receptor and shifting the equilibrium to a greater population of free Gαo. An agonist might be expected to facilitate this shift in equilibrium; however, this was not observed. Perhaps under these experimental conditions, the role of the agonist is to stimulate the receptor to function as a “guanine nucleotide exchange factor” and thereby facilitate the release of the GTP analog and allow reassembly of the CB1 receptor-Gαo complex.

Effect of guanine nucleotide triphosphate, agonists and antagonist on CB1 receptor-Gαoassociation. CHAPS extracts from rat brain membranes (100 μg protein) were incubated for 20 min at 30°C in the presence of the indicated agents. Treatments were followed by immunoprecipitation and Western blot analysis. All lanes were immunostained with both anti-CB11-14 and anti-Gαo. Lanes on the blots show membranes treated with 1, no additions; 2, DALN (1 μM); 3, DALN (1 μM) plus GTPγS (100 μM); 4, GTPγS (100 μM); and 5, SR141716A (1 μM). The bands from three individual experiments were scanned, and density values were (mean ± S.E.) 1, 100%; 2, 99 ± 1.8%; 3, 60 ± 4.3%; 4, 28 ± 5.1%; and 5, 87 ± 6.4%.
We determined that incubation of rat brain membranes or CHAPS-solubilized extracts with pertussis toxin (A subunit) facilitated a shift in the population from CB1receptor-Gαo complexes to dissociated receptors and Gαo subunits (Howlett et al., 1999). Inasmuch as pertussis toxin ADP-ribosylates a Cys residue on Gαo that is critical for the interaction with G protein-coupled receptors, this finding corroborates the notion that the CB1 receptor-Gαocomplex in detergent solution can be disrupted on experimental manipulation.
Peptide CB1401-417 Competes with CB1Receptor for Interaction with Gαo and Gαi3but Not Gαi1 or Gαi2 in CHAPS Extracts of Rat Brain Membranes.
It was hypothesized that the juxtamembrane C-terminal domain of the CB1 receptor is of primary importance in the CB1 receptor-Gα interaction. If this were the case, then peptide CB1401-417 should compete for binding to the receptor recognition site on Gi/o proteins. To determine whether CB1401-417 can disrupt the equilibrium between CB1receptor-Gαo complex and free proteins, CHAPS extract from rat brain membranes was incubated with CB1401-417, immunoprecipitated with the anti-CB11-14 affinity matrix, and assessed by Western blotting. Shown in Fig. 5, preincubation with peptide CB1401-417 significantly reduced the Gαo band density (49 ± 4.3%) without any change in the band densities for the CB1 receptor (101 ± 7.5%). A concentration-response relationship study indicated that under these conditions, no disruption was observed below 0.2 mM peptide, and half-maximal disruption was observed at 0.75 mM (data not shown). Lower concentrations (0.5 mM) of CB1401-417 could maximally compete for the CB1receptor-Gαo association if a longer preincubation period (1 h) were allowed. At room temperature, peptide CB1401-417 induced very little disruption of the CB1 receptor-Gαoassociation, even at higher concentrations (1 mM) and prolonged (1-h) preincubation (data not shown). This finding demonstrates that peptide CB1401-417 competes for the CB1 receptor-Gαoassociation.

Effect of equilibrating peptide CB1401-417 with the CB1 receptor-Gα complex. CHAPS extract from rat brain membranes (100 μg protein) was incubated in the absence or presence of peptide CB1401-417 (1 mM) at 30°C for 30 min (lanes 1 and 2, respectively). The CB1receptor was immunoprecipitated, and Western blot analysis was performed as described in the text. A, representative immunoblot was stained with anti-CB11-14 plus anti-Gαo. The ratio of the relative densities of the Gαo to CB1 receptor band (both bands were quantified and added to obtain total CB1 receptor density) is shown, where control is 100% (mean ± S.E., three separate experiments). B, representative immunoblot was stained with anti-CB11-14 plus anti-Gαi1/2. The ratio of the relative densities of Gαi1/2 to CB1 receptors (bands added) is shown, where control is 100% (mean ± S.E., three separate experiments).
Parallel experiments were performed to determine whether peptide CB1401-417 could uncouple CB1 receptor-Gαiinteractions in CHAPS extracts from rat brain membranes (Figs. 5B and6). No significant reduction in the Gαi1/2 band density compared with control was observed after incubation with peptide CB1401-417. This would suggest that the CB1 receptor-Gαi1/2association was refractory to competition with peptide CB1401-417. Alternatively, the peptide may not be accessible to the receptor recognition site and therefore may not compete under the incubation conditions used. To clarify whether both of the subtypes detected by this antibody were refractory to competition by peptide CB1401-417, antibodies that were selective for each of the three Gαisubtypes were used for detection in the competition protocol (Fig. 6). Gαi1 and Gαi2 were both refractory to competition by peptide CB1401-417 (Fig. 6, A and B), whereas the CB1receptor-Gαi3 complex was disrupted by peptide CB1401-417 (Fig. 6C). The Gαi3 band density relative to the CB1 receptor density was reduced by 90% after equilibration with peptide CB1401-417.

Effect of peptide CB1401-417 on the association of the CB1 receptor with Gαisubtypes. CHAPS extract from rat brain membranes (100 μg protein) was incubated in the absence (lanes 1) or presence (lanes 2) of peptide CB1401-417 (1 mM) at 30°C for 30 min, the CB1receptor was immunoprecipitated, and Western blot analysis was performed. Blots were stained with anti-CB11-14 plus anti-Gαi1 (A), anti-Gαi2 (B), and anti-Gαi3 (C). This experiment is representative of three studies with similar results.
The CB1 cannabinoid receptor is differentially expressed in regions of the brain as indicated by [3H]CP55940 binding in brain slices (Herkenham et al., 1991). Researchers from Childers' laboratory have noted that the CB1 receptor exhibits variations in coupling efficiency to G proteins in different regions of the rat brain (Breivogel et al., 1997). To determine whether the CB1 receptor association with G proteins is regionally specific, we quantified the precoupled CB1 receptor-Gαo complex in three regions of rat brain (Fig. 7). The relative band density ratios of Gαo to CB1 receptor were 0.88, 0.54, and 2.51 for cortex, striatum, and cerebellum, respectively (A). The relative association of the CB1 receptor with Gαo was greater in the cerebellum than in the frontal cortex, and relatively little CB1receptor-Gαo complex was found in the striatum. The relative band density ratios of Gαi1/2 to CB1 receptor were 0.86, 1.26, and 1.46 for cortex, striatum, and cerebellum, respectively (Fig. 7B). The regional differences in coupling to Gαi1/2 were not as pronounced as for Gαo, and coupling to Gαi1/2 was relatively high in the striatum. These findings suggest that in the absence of exogenous agonists, regional differences exist in the preference of CB1 receptor-G protein coupling in the brain. Of note, the CB1 receptor in the striatum associates to a greater extent with Gαi1/2, and the CB1 receptor in the cerebellum associates to a greater extent with Gαo.
We determined whether regional specificity exists in the requirement for the juxtamembrane C-terminal domain in the association of CB1 receptors with G proteins. Peptide CB1401-417 was able to compete equally well for the CB1 receptor-Gαointeraction in all three brain regions (Fig. 7A). As was shown for the membranes from the whole brain, peptide CB1401-417 failed to disrupt the CB1 receptor-Gαi1/2interaction in any of the three brain regions (Fig. 7B). These region-specific data are internally consistent with data obtained from the whole brain.
Specificity of CB1 Receptor Juxtamembrane C-Terminal Peptide for Gα Interactions.
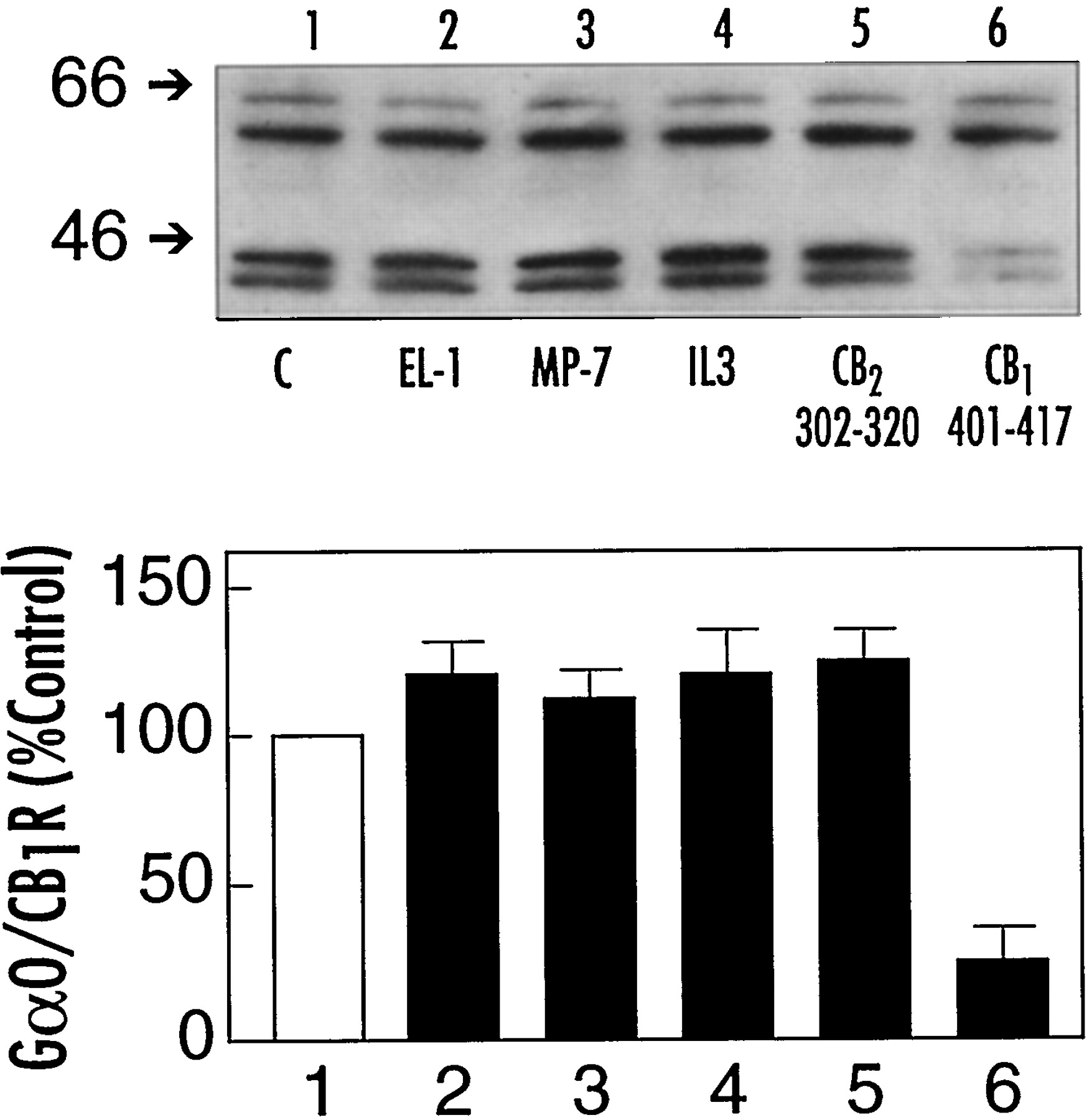
To determine the specificity of peptide CB1401-417 to disrupt the CB1 receptor-Gαointeraction, a series of peptides derived from other domains of the CB1 receptor were tested (Fig.8). CB1401-417 disrupted the interaction of the CB1 receptor with Gαo as previously demonstrated (lane 6 compared with lane 1). The CB1 receptor first extracellular loop (peptide EL1, Table 1) is not believed to contribute to ligand binding or signal transduction of the CB1 receptor (Howlett et al., 1998). As expected, this peptide did not disrupt the CB1receptor-Gαo interaction (lane 2 compared with lane 1).

Specificity of peptide CB1401-417 to dissociate the CB1 receptor-Gαo complex. CHAPS extracts from rat brain membranes (100 μg protein) were incubated at 30°C for 30 min with the indicated peptides (1 mM): lane 1, control without peptide (C); lane 2, peptide CB1174-188 (EL-1); lane 3, mastoparan-7 (MP-7); lane 4, peptides CB1310-317 plus CB1316-327 plus CB1329-344, each at 1 mM (IL3); lane 5, peptide CB2302-320; and lane 6, peptide CB1401-417 (see Table 1). After equilibration, the mixture was subjected to immunoprecipitation and Western blot analysis. Proteins in the immunoprecipitates were detected by staining the blots with anti-CB11-14 plus anti-Gαo. Bottom, relative densities from three or four individual experiments (mean ± S.E.).
Three third intracellular loop peptides were synthesized to mimic the domains comprising the N-terminal side (CB1301-317), the middle (CB1316-327), and the C-terminal side (CB1329-344) of the third intracellular loop. The N-terminal peptide fragment is of particular note because it exhibits some activity to inhibit adenylyl cyclase in N18TG2 membranes, although the response is not as robust as that of peptide CB1401-417 (Howlett et al., 1998). This third intracellular loop peptide has little ability to stimulate [35S]GTPγS binding to G proteins in rat brain membranes unless accompanied by the other two peptides composing the remainder of the third intracellular loop (Howlett et al., 1998). Under conditions of incubation that allowed peptide CB1401-417 to effectively compete for the CB1 receptor interaction with Gαo, the combination of the three third intracellular loop peptides was totally ineffective (lane 4 compared with lane 1). In parallel experiments using CHAPS-solubilized membranes from N18TG2 cells, it was shown that the combination of third intracellular loop peptides could not disrupt the interaction between the CB1 receptor and Gαi3(data not shown). These data suggest that in contrast with the juxtamembrane C-terminal region, the third intracellular loop domain may have little importance in the CB1receptor-Gαo or Gαi3interactions.
Mastoparan-7 is an active analog of mastoparan, a bee venom peptide that has the ability to autonomously activate G proteins (Higashijima et al., 1990). Mastoparan-7 had no ability to disrupt the CB1 receptor-Gαo complex under conditions in which peptide CB1401-417 evoked a robust response (lane 3). One property of mastoparan and mastoparan-7 that correlates with the ability to activate G proteins is the ability of these peptides to form an amphipathic α-helical structure in a hydrophobic environment (Higashijima et al., 1990). We determined that in contrast to mastoparan, CB1401-417 could not form an α-helical structure in a hydrophobic environment (methanol, trifluoroethanol) but could in the presence of a negatively charged environment (SDS, phosphatidic acid) (Mukhopadhyay et al., 1999). Furthermore, the α-helical structure was not a requirement for peptide CB1401-417 to stimulate Gifunctions in membrane preparations (Mukhopadhyay et al., 1999). Thus, although both peptides are cationic and can form α-helical structure in certain environments, differences exist in the mechanism of signaling to G proteins between mastoparan analogs and the juxtamembrane C-terminal domain of the CB1receptor.
The CB2 receptor is able to couple to Gi proteins to inhibit adenylyl cyclase in recombinant host cells (Felder et al., 1995). To test whether the C-terminal juxtamembrane region of the CB2receptor can serve the same coupling function as does the homologous region of the CB1 receptor, a peptide fragment from this region was incubated with the CHAPS extract of rat brain membranes. As shown in Fig. 8, lane 5, peptide CB2302-320 from the CB2receptor did not compete for the CB1receptor-Gαo complex. CB2302-320 possesses two of the three cationic amino acids at the N-terminal side of peptide CB1401-417 and has three residues that are identical and two that are similar within the N-terminal six amino acids. This homology is insufficient to mimic the ability of peptide CB1401-417 to inhibit adenylyl cyclase (S. Mukhopadhyay and A. C. Howlett, unpublished observations) or to disrupt the CB1receptor-Gαo association.
Discussion
A high molecular weight form of the CB1receptor was routinely observed in rat brain membrane preparations. The high molecular weight CB1 receptor band on SDS-PAGE is devoid of G proteins or their subunits and thus cannot represent a receptor-G protein complex. The absence of G proteins in the high molecular weight forms of other G protein-coupled receptors has also been demonstrated (Ciruela et al., 1995; Wreggett and Wells, 1995; Ng et al., 1996). For the CB1 receptor, the high molecular weight form continues to exist after enzymatic deglycosylation (C. Song and A. C. Howlett, unpublished observations). Similarly, glycosylation has not been found to be involved in multimer formation for other G protein-coupled receptors that have been subjected to enzymatic deglycosylation (Ciruela et al., 1995; Ng et al., 1996; Cvejic and Devi, 1997) or G protein-coupled receptors expressed in Sf9 insect cells that fail to efficiently glycosylate recombinant mammalian proteins (Ng et al., 1994; Schreurs et al., 1995).
Evidence from protein cross-linking studies suggests that G protein-coupled receptors may exist as multimers in their native membrane environment (Herberg et al., 1984; Hebert et al., 1996; Ng et al., 1996; Cvejic and Devi, 1997). Multimers persist through detergent solubilization and immunoaffinity purification of the CB1 receptor as well as other G protein-coupled receptors. The CB1 receptor SDS-resistant complex is also resistant to disruption of disulfide linkages. For the rhodopsin-like subfamily, the transmembrane helices are important for dimerization of β2-adrenergic and D2 dopamine receptors, which could be selectively dissociated by incubation with peptides corresponding to helices VI and VII (Hebert et al., 1996; Ng et al., 1996). Some G protein-coupled receptor dimers could be disrupted by extended (30 min) exposure of solubilized, immunoprecipitated receptors to high temperature (65°C or 90°C) or acid (pH 3; Ng et al., 1996). However, consistent with our findings for the CB1 receptor, extreme conditions are necessary to promote protein-protein dissociation of the high molecular weight complex.
The functional significance of CB1 receptor multimers has yet to be understood. Ligand binding selectivity may differ as a function of the oligomeric state of the receptor, as shown for the D2 dopamine receptor, in which the dimeric form recognizes multiple classes of ligand whereas the monomer is more selective (Ng et al., 1996). Wreggett and Wells (1995) provided radioligand binding data supporting a model of a tetrameric receptor complex that exhibits cooperativity in ligand binding in the absence or presence of G proteins. In that model, the G proteins act by modulating the degree of cooperativity. Dimerization may be an important step in hormone receptor signaling, as has been suggested for the β2-adrenergic receptor (Hebert et al., 1996) and mGlu1/mGlu2 receptor heterodimers (Kuner et al., 1999). Alternatively, receptor dimer to monomer conversion may play an early role in the agonist-evoked internalization process as proposed for the δ-opioid receptor (Cvejic and Devi, 1997).
The present results demonstrate that the CB1receptor is associated with Gαo, Gαi1, Gαi2, or Gαi3 after disruption of the membrane by CHAPS detergent, and this complex can be coimmunoprecipitated under these mild detergent conditions. Other laboratories have used antibodies against Gα subunits to immunoprecipitate associated receptor proteins (Okuma and Reisine, 1992; Damaj et al., 1996); however, the effectiveness of antibodies directed against the Gα C terminus to interact with receptors is compromised by the presence of agonists (Okuma and Reisine, 1992). Immunoprecipitation with antibodies against the receptor have provided evidence for G protein association with M2 muscarinic (Matesic et al., 1991) and sst1 and sst2A somatostatin (Gu and Schonbrunn, 1997) receptors.
CB1 receptor-Gαi/ocomplexes formed in the absence of pretreatment with exogenous agonist ligands, in a manner similar to what has been observed for κ-opioid (Frances et al., 1990) and D2-dopamine (Senogles et al., 1987) receptors. However, this finding contrasts with reports of other G protein-coupled receptors that require the presence of agonists to stabilize a complex with G proteins during detergent solubilization. For example, G proteins were complexed with only the agonist-occupied muscarinic (Matesic et al., 1989; 1991), somatostatin (Murray-Whelan and Schlegel, 1992; Gu and Schonbrunn, 1997), and interleukin-8 (Damaj et al., 1996) receptors. The finding that a CB1 receptor-G protein complex exists spontaneously in detergent extracts supports the idea that the CB1 receptor is “precoupled” to G proteins in the cell membrane. Consistent with this idea is the finding that the CB1 receptor is constitutively active in recombinant overexpression systems (Bouaboula et al., 1997) and in brain membranes and N18TG2 cells that endogenously express the CB1 receptor (J. P. Meschler and A. C. Howlett, unpublished observations). We estimate that the immunoprecipitation procedure yields about 85% recovery of the Gαo that was present in the CHAPS extract from rat brain membranes, suggesting that the CB1receptor entraps a majority of the available Gαo.
Specific competition by peptide CB1401-417 for the CB1 receptor-Gαo or Gαi3 interactions suggests that this peptide is able to occupy a locus on these G proteins that represents a site of protein-protein association. The ability of peptide CB1 401-417 to autonomously activate G proteins suggests that this locus is the activation site (Howlett et al., 1998;Mukhopadhyay et al., 1999). Studies using synthetic peptides have implicated this domain of the receptor in G protein coupling for rhodopsin/transducin (Konig et al., 1989), β-adrenergic receptor/Gαs (Munch et al., 1991),N-formyl peptide receptor/Gαi2(Schreiber et al., 1994), angiotensin II receptor/Gαi1, Gαi2, and Gαo (Shirai et al., 1995; Sano et al., 1997), and δ-opioid receptor/Gαi/o (Merkouris et al., 1996). Peptides from rhodopsin and the N-formyl peptide receptor competed for protein-protein interactions between receptors and G proteins, but no activation of signal transduction was demonstrated (Konig et al., 1989; Schreiber et al., 1994). The juxtamembrane C-terminal peptide from β-adrenergic and δ-opioid receptors antagonized agonist-stimulated signal transduction, suggesting a competition for binding to Gα but failure to evoke activation (Munch et al., 1991; Merkouris et al., 1996). The angiotensin II receptor juxtamembrane C-terminal peptide, like peptide CB1401-417, was found to activate G proteins in GTPγS binding assays (Shirai et al., 1995).
It appears from the selectivity that peptide CB1401-417 shows for CB1receptor-Gαo and -Gαi3complexes that Gαi1 and Gαi2 may not possess a high-affinity site for peptide CB1401-417. This would mean that Gαi1 and Gαi2 are not targets for activation by the juxtamembrane C-terminal domain of the CB1 receptor. The observation that a peptide fragment of the homologous region of the CB2receptor fails to compete for the CB1receptor-Gαo complex suggests that this region of the CB2 receptor fails to perform the analogous function of activation of Gαo. These findings argue for selectivity of domains on G protein-coupled receptors that can bind to and activate specific G proteins. Of note, it is the C-terminal domain of the prostaglandin EP3 receptor family of splice variants that defines the G protein coupling selectivity (Namba et al., 1993).
The intrinsic efficacy of agonist-receptor stimulated G protein signaling can be assessed by measuring net agonist-stimulated [35S]GTPγS binding. The ratio of apparentB max of agonist-stimulated [35S]GTPγS binding andB max of receptor radioligand binding is an indicator of the receptor/transducer amplification factor (Breivogel et al., 1997). Recent studies from researchers at Childers' laboratory suggest that the CB1 receptor/G protein amplification factor varies widely among different regions of the brain (Breivogel et al., 1997). Our data suggest that precoupling of the CB1 receptor with different subtypes of G proteins in neurons from various brain regions may influence receptor/transducer amplification. One can envision that a brain region that exhibits predominantly CB1receptor-Gαo or -Gαi3complexes in their precoupled state may respond preferentially to agonists that induce a conformational change in the juxtamembrane C-terminal domain of the receptor. In contrast, neurons in brain regions that exhibit preferential CB1receptor-Gαi1 or -Gαi2coupling may respond more efficiently to agonists that induce conformational changes in an alternative intracellular surface of the CB1 receptor. Previous studies have suggested that G proteins and the cationic allosteric site influence the affinity states of aminoalkylindole agonists differently from cannabinoid agonists (Houston and Howlett, 1998). It may be that this difference is a result of alternative conformations of the receptor induced by these two structurally diverse classes of CB1 receptor agonists.
In summary, we have shown that the CB1 receptor can exist as an SDS-resistant multimer. In CHAPS detergent, the CB1 receptor exists in a complex with G proteins of the Gi/o family that, unlike other G protein-coupled receptors, does not require the presence of agonists. We have used a novel approach of using peptides representing specific domains of the receptor to competitively disrupt interactions with coimmunoprecipitated proteins. With this methodology, we demonstrated that a peptide derived from the juxtamembrane C-terminal domain of the CB1 receptor can uncouple specific CB1 receptor-Gαo and Gαi3 interactions, suggesting that this domain represents the Gα activation site for these G proteins. The finding that receptor interactions with Gαi1 and Gαi2 are not affected suggests the possibility for selectivity in G protein-mediated signaling pathways. Further studies are in progress to elucidate the mechanisms of interaction of the CB1 receptor with other Gi/o proteins.
Acknowledgments
We thank Kelly Jester for whole rat brain preparations and Rusoun Belue for performing the rat brain dissections and membrane preparations from these regions.
Footnotes
-
Send reprint requests to: Dr. Allyn C. Howlett, Department of Pharmacological and Physiological Science, Saint Louis University School of Medicine, 1402 South Grand Blvd., St. Louis, MO 63104. E-mail: howletta{at}slu.edu
-
↵1 Present address: National Enzyme Company, Forsyth, MO 65653.
-
This work was supported in part by NIDA Grants R01-DA03690 and K05-DA00182.
- Abbreviations:
- MAPK
- mitogen-activated protein kinase
- CAPS
- 3-[cyclohexylamino]-1-propanesulfonic acid
- CHAPS
- 3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate
- DALN
- desacetyllevonantradol
- ECL
- enhanced chemiluminescence
- GTPγS
- guanosine-5′-O-(3-thio)- triphosphate
- PAGE
- polyacrylamide gel electrophoresis
- Received June 4, 1999.
- Accepted October 8, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}