


Abstract
Recombinant human A2B adenosine receptors (A2BARs) and receptors extended on the amino terminus with hexahistidine and the FLAG epitope, DYKDDDDK (H/F-A2B) were stably overexpressed (to >20,000 fmol/mg protein) in human embryonic kidney 293 cells (HEK-A2B). By Western blotting, the H/F-A2Breceptor runs as a 34.8-kDa glycoprotein. Pharmacological properties of A2BARs were characterized with125I-3-aminobenzyl-8-phenyl-(4-oxyacetic acid)-1-propylxanthine (KD, 36 nM). In competition binding assays, the affinity of agonists is reduced by substitution on either the N6- or the C-2 position of the adenine ring, whereas 5′-substitutions increase affinity, resulting in the potency order: 5′-N-ethylcarboxamidoadenosine (NECA) ≫N6-aminobenzyl-NECA ≈2-chloroadenosine > 2-[4-(2-carboxyethyl)phenethylamino]-NECA (CGS21680) >N6-aminobenzyladenosine. The A2BAR is potently blocked by the A2A-selective antagonist 4-(2-[7-amino-2-[2-furyl][1,2,4]triazolo-[2,3-a][1,3,5] triazin-5-yl-amino]ethyl)phenol (ZM241385; KI, 32 nM for A2B, 1.4 nM for A2A) and the A1 selective antagonist 8-cyclopentyl-1,3-dipropylxanthine (KI, 50.5 nM for A2B; 2.5 nM for A1). TheKI values for the antiasthmatic xanthines, theophylline (7.8 μM) and enprofylline (6.4 μM), are below their therapeutic plasma concentrations (20 to 50 μM), and agree withKI determinations for inhibition of NECA-stimulated cAMP accumulation in HEK-A2B cells. NECA orN6-(2-iodo)benzyl-5′-N-methylcarboxamidodoadenosine (IB-MECA) stimulate inositol trisphosphates and calcium accumulation in HEK-A2B or HEK-A3 cells, respectively, but only the A3 response is prevented by pertussis toxin. In human HMC-1 mast cells, A2BAR activation stimulates calcium mobilization and cAMP accumulation. We conclude that HEK-A2B cells and HMC-1 mast cells possess A2BAR glycoproteins that are coupled to both Gq/11 and Gs.
Of the four subtypes of adenosine receptors (ARs), A1, A2A, A2B, and A3 (Linden et al., 1994), radioligand binding assays have been extensively used for all except the A2BAR. Consequently, the pharmacological characterization of A2BARs has been based primarily on functional assays of cAMP accumulation in tissue slices or cultured cells. A2BARs have been functionally evaluated in NIH3T3 cells (Brackett and Daly, 1994) and demonstrated in aorta (Martin, 1992), chromaffin cells (Casado et al., 1992), astrocytes (Peakman and Hill, 1994), erythroleukemia cells (Strohmeier et al., 1995), astroglioma cells (Fredholm and Altiok, 1994), mast cells (Auchampach et al., 1997), intestinal smooth muscle (Burnstock, 1978), and intestinal epithelia (Strohmeier et al., 1995). On cloning of the rat A2BAR cDNA, highest levels of transcript were found in large intestine, cecum, and urinary bladder (Rivkees and Reppert, 1992). We have noted previously that a low-specific-activity radioligand, [1,3-3H]diethyl-8-phenylxanthine (DPX) (Fig.1), binds to a saturable site on HEK-A2B membranes (Robeva et al., 1996a). Here we describe a binding assay improved by the generation of new HEK-A2B cell line that expresses a high density of A2BARs. A second improvement in radioligand binding to A2BARs derives from the use of a high-specific-radioactivity radioligand,125I-3-(3-iodo-4-aminobenzyl)-8-(4-oxyacetate)phenyl-1-propyl-xanthine (125I-ABOPX).

The structure of some xanthine antagonists of A2B adenosine receptors that are referred to in the text.
Antipeptide antibodies raised against the A2Badenosine receptor recognize a putative nonglycosylated receptor with a molecular mass of 57 kDa (Puffinbarger et al., 1995). This differs from the 33-kDa glycoprotein predicted by the A2BAR receptor cDNA. Here we show that a purified, recombinant human A2B adenosine receptor extended on the amino terminus with hexahistidine and the FLAG epitope (DYKDDDDK) (H/F-A2B) is a glycoprotein with an apparent molecular mass of 34.8 kDa.
Adenosine stimulates bronchoconstriction and the degranulation of mast cells in people with asthma, but not in normal subjects (Cushley and Holgate, 1985). The A3AR has been implicated as the receptor that is activated by adenosine to trigger degranulation of rodent perivascular mast cells (Jin et al., 1997) and rat RBL-2H3 mast-like cells (Ramkumar et al., 1993). However, A2BARs appear to play a predominant role in the regulation of canine BR mast cell degranulation (Auchampach et al., 1997) and the slow release of interleukin-8 from human HMC-1 mast cells (Feoktistov and Biaggioni, 1995). It is curious, however, that A2B receptors are known to couple to cAMP accumulation, because the activation of protein kinase A is thought to inhibit mast cell degranulation (Lohse et al., 1987; Jin et al., 1997). Here we present evidence that in addition to coupling to Gs, A2B receptors couple to Gq/11 and calcium mobilization in HEK-A2B cells and HMC-1 mast cells. The antiasthma drugs theophylline and enprofylline effectively block human A2BARs and receptor-mediated mast cell degranulation at therapeutic concentrations.
Experimental Procedures
Materials.
Wizard Megaprep columns and competent JM109 cells were obtained from Promega Corp. (Madison WI);R-N6-(2-phenylisopropyl)adenosine, 5′-N-ethylcarboxamidoadenosine (NECA), CPA, pepstatin A, leupeptin, aprotinin, phenylmethylsulfonyl fluoride, benzamidine, and forskolin were obtained from Sigma Chemical Corp. (St. Louis, MO). 2-[4-(2-Carboxyethyl)phenethylamino]-5N-N-ethylcarboxamidoadenosine (CGS21680), 2-chloroadenosine, DPX, and 8-(4-((2-aminoethyl)aminocarbonylmethyloxy)phenyl)-1,3-dipropylxanthine (XAC) were obtained from Research Biochemicals International (Natick, MA); adenosine deaminase was obtained from Boehringer Mannheim (Indianapolis, IN); lipofectin, G418, tissue culture media and serum, were obtained from Gibco BRL (Gaithersburg, MD).N6-Iodoaminobenzyladenosine (I-ABA), 8-(4-carboxyethenylphenyl)-1,3-dipropylxanthine (BW-A1433), 8-cyclopentyl-1,3-dipropylxanthine (CPX) and127I-ABOPX (BW-A522) were gifts from Dr. Susan Daluge of Glaxo Wellcome (Research Triangle Park, NC). Aminobenzyl-5′-N-ethylcarboxamidoadenosine (AB-NECA) was a gift from Dr. Ray Olsson of the University of South Florida (Tampa, FL); anti-FLAG m2 antibodies were obtained from Kodak IBI (New Haven, CT).
Stable Transfection of HEK 293 Cells.
cDNA for human A2BARs was prepared by polymerase chain reaction of human brain cDNA (Clonetech, Palo Alto, CA) and sequenced on both strands. cDNAs encoding human A1, A2A, and A3 ARs were gifts of Marlene Jacobson (Merck & Co, West Point, PA). DNA sequencing was carried out in the University of Virginia Biomolecular Research Facility with an ABI Prism 377 Automated DNA Sequencer. The four wild-type human adenosine receptor cDNAs were subcloned into the expression plasmid CLDN10B. To prepare H/F-A2B, the cDNA was subcloned into the pDoubleTrouble plasmid (Robeva et al., 1996b. The plasmids were amplified in competent JM109 cells and plasmid DNA isolated by using Wizard Megaprep columns (Promega Corporation, Madison, WI). Recombinant receptors were introduced into HEK 293 cells by lipofectin. Colonies were selected by growth of cells in 0.6 mg/ml G418. Stably transfected cells were maintained in Dulbecco’s modified Eagle’s medium/Ham’s F12 medium (DMEM/F12 medium) with 10% fetal calf serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 0.3 mg/ml G418. G418 was omitted from the last passage before harvest.
Radioligand Binding.
To prepare125I-ABOPX, 10 μl of 1 mM ABOPX in methanol/1 M NaOH (20:1) was added to 50 μl of 100 mM phosphate buffer, pH 7.3. One or 2 mCi of Na125I were added, followed by 10 μl of 1 mg/ml chloramine T freshly prepared in water. After incubating for 20 min at room temperature, 50 μl of 10 mg/ml Na-metabisulfite in water was added to quench the reaction. The reaction products were applied to a C18 HPLC column and eluted for 5 min with 4 mM phosphate, pH 6.0/methanol (65:35). The methanol concentration was then ramped to 100% over 15 min. ABOPX elutes in 11 to 12 min and 125I-ABOPX elutes at 18 to 19 min in a yield of 50 to 60% of the initial 125I. For equilibrium binding assays, the specific activity of125I-ABOPX was diluted 10- to 20-fold by the addition of 127I-ABOPX . Radioligand binding assays were conducted at 21°C for 2 to 3 h in 100 μl of buffer containing 10 mM HEPES, pH 7.4, 1 mM EDTA, 5 mM MgCl2, 20 μg membrane protein, and 1 U/ml adenosine deaminase. Nonspecific binding was measured in the presence of 10 μM 8(4-((2-aminoethyl)-aminocarbonylmethyloxy)phenyl)-1,3-dipropylxanthine (XAC). Competition binding experiments were carried out with 0.5 to 0.6 nM 125I-ABOPX. To detect bound radioligand, membranes were filtered over Whatman GF/C filters by using a Brandel cell harvester (Gaithersburg, MD) and washed 3 times over 15 to 20 s with ice-cold buffer (10 mM Tris, 1 mM MgCl2,pH 7.4).It is important to maintain the wash buffer in an ice-slurry and to prime the filtration apparatus with ice-cold wash buffer to prevent dissociation of specific binding during the wash of glass fiber filters. Bmaxand KD values were calculated by nonlinear least-squares interpolation to a single-site binding model.
To characterize additional recombinant human adenosine receptor subtypes, radioligands and compounds used to detect nonspecific binding, respectively, were: A1, [3H]CPX, 10 μM CPX; A2A, 125I-2-[2-(4-amino-phenyl)ethylamino]adenosine (125I-APE) or125I-ZM241385, 10 μM NECA, or 1 μM ZM241385; and A3, 125I-ABA, 100 μM NECA. In competition binding assays, IC50 values for competing compounds were derived by fitting the data to a four-parameter logistic equation: B = specific/(1+(IC50)n) + nonspecific, where B = specific binding and n = the Hill slope. KI was derived from IC50 as described previously (Linden, 1982).
Western Blots.
Membranes expressing H/F-A2BARs were solubilized in digitonin and purified by anti-FLAG affinity chromatography (Robeva et al., 1996b). Purified receptors were subjected to SDS polyacrylamide gel electrophoresis. In some instances, receptors were incubated with 0.5 U of N-glycosidase F for 18 h at 37°C. Electrophoresed receptors were transferred to Westran polyvinylidene difluoride membranes. The membranes were incubated overnight at 4°C in 10% milk Blotto (50 mM Tris, 80 mM NaCl, 20% milk, 0.2% Tween 20, pH 8). Blots were rinsed with Tris-buffered saline/Tween 20 (TBS/T) buffer (20 mM Tris, 137 mM NaCl, 0.3% Tween 20, pH 7.6), incubated with anti-FLAG antibody (5 μg/ml) in TBS/T containing 1 mg/ml BSA for 1 h at room temperature, washed in TBS/T, and incubated for 1 h with horseradish peroxidase-conjugated sheep antimouse IgG F(ab′)2 fragments diluted 1:10,000. After washing in TBS/T, blots were visualized by enhanced chemiluminescence. If untransfected HEK cells replaced HEK-A2B cells, no detectable protein or immunoreactivity was eluted from anti-FLAG affinity columns.
cAMP Measurements.
HEK-A2B or Chinese hamster ovary (CHO)-K1 cells were grown to near confluence on 150-mm diameter plates. The cells were removed by replacing the medium with PBS containing 5 mM EDTA for 5 min. Cells were pelleted by centrifugation at 250g for 5 min, washed once in DMEM, and resuspended in DMEM supplemented with 1 U/ml adenosine deaminase and 10 mM HEPES, pH 7.2, resulting in a cell density of 250,000 cells/ml. The cells were equilibrated at ambient temperature for 1 h. To initiate cAMP accumulation, 50 μl of test compound was added to 200 μl of suspended cells and incubated for 10 min at 37°C in a shaking water bath. The cells were lysed by the addition of 500 μl of 0.15 N HCl. After centrifugation at 2,000g for 10 min, 500 μl of supernatant was removed, acetylated, and acetyl-cAMP measured by automated radioimmunoassay. The pA2 of antagonists was determined by the method of Schild (Schild, 1957).
Inositol Trisphosphate (IP3) Measurements.
HEK-A2B or HEK-A3 cells were grown on 100-mm tissue culture plates. The medium was removed and the cells were washed once with inositol-free DMEM/F12 and incubated for 24 to 48 h with 2.5 μCi/ml [myo-3H]inositol in inositol-free DMEM/F12 plus 2% fetal calf serum. For treatment of cells with pertussis toxin, 200 ng/ml toxin was added for 18 h. After tritium labeling, the cells were washed and resuspended in HEPES-buffered DMEM (20 mM HEPES, pH 7.2) plus 1 U/ml adenosine deaminase and 100 mM LiCl and pipetted into test tubes (106 cells/0.2 ml). Cells were maintained at 37°C for 30 min in a shaking water bath. Assays were terminated by the addition of 400 μl of stop solution (0.5 M HClO4, 5 mM EDTA, and 1 mM diethylenetriaminepentaacetic acid) and 1 mg/ml phytic acid. The tubes were placed on ice for 30 min and 5 M K2CO3 was added to raise the pH to 8 to 9. Samples were centrifuged to remove the KClO4 precipitate and the supernatants were collected and filtered through 0.45-μm Gelman Acrodisc filters and then applied to columns containing 1 ml of anion exchange resin (AG 1-X8, chloride form, 200 to 400 mesh; Bio-Rad Laboratories, Richmond, CA). Columns were washed with 5 ml of water, then 5 ml of 40 mM HCl. IP3 fractions were eluted with 5 ml of 170 mM HCl and radioactivity counted by liquid scintillation counting.
Measurement of Intracellular Ca2+.
Cells on tissue culture plates were loaded with 1 μM 1-[2-(5-carboxyoxazol-2-yl)-6-aminobenzofuran-5-oxyl]-2′-amino-5′-methylphenoxy)-ethane-N,N,N′,N′-tetraacetic acid/acetoxymethyl ester in a buffer containing 100 mM NaCl, 5 mM KCl, 1 mM MgSO4, 1 mM KH2PO4, 25 mM NaHCO3, 0.5 mM CaCl2, 2.7 g/liter d-glucose, 20 mM Na-HEPES, pH 7.4, and 0.25% BSA for 45 min. Cells were washed, detached, and resuspended in the same buffer without BSA, plus 1 U/ml adenosine deaminase to a density of 106 cells/ml. Fluorescence was monitored in an SLM 1100 spectrofluorometer in a thermostated stirred cuvette at 37°C at an emission wavelength of 510 nm and excitation wavelengths of 340 and 380 nm.
Results
Western Blotting of H/F-A2B Adenosine Receptors.
To estimate the molecular mass of H/F-A2BARs, receptors were solubilized in digitonin, purified over anti-FLAG affinity columns and detected by anti-FLAG western blotting. To determine whether the A2BAR was a glycoprotein, some of the purified receptors were incubated withN-glycosidase F before electrophoresis. The results are illustrated in Fig. 2. The H/F-A2BAR had an apparent molecular mass of 34.8 kDa reduced to 31.3 kDa after enzymatic deglycosylation. Some purified deglycosylated receptors seemed to form dimers with an apparent molecular mass of 65 kDa.The amount of dimerized receptors detected in purified receptor preparations increased with incubation time, receptor concentration, and temperature and may be an artifact of receptor purification, without physiological significance. The properties of the H/F-A2BAR monomeric glycoprotein are consistent with its expected size and the existence of consensus sites for N-liked glycosylation, based on the receptor cDNA sequence.

Western blot analysis of purified H/F-A2Badenosine receptors. Purified receptors (50 ng) were subjected to western blotting with anti-FLAG antibodies without (−) or with (+) prior incubation overnight at 37°C with N-glycosidase F (N-Gly-F). The apparent molecular masses of the glycosylated and deglycosylated receptors, respectively, are 34.8 and 31.3 kDa. The upper band in the deglycosylated lane has a molecular mass of 65 kDa.
Radioligand Binding Studies.
Clonal lines of HEK-A2B cells grown in G418 were screened for maximal 125I-ABOPX binding. Figure3 shows equilibrium binding of125I-ABOPX to membranes derived from the clone that was found to have the highest level of expression of the wild-type A2BAR, with a Bmax in excess of 20,000 fmol/mg protein. A single saturable binding site was detected in transfected, but not in untransfected cells. There is an endogenous A2BAR that stimulates cAMP production in untransfected HEK 293 cells, but the density of this receptor is too low to be detected by radioligand binding with125I-ABOPX. The pharmacological properties of the 125I-ABOPX binding site were characterized in competition binding studies with a series of 11 agonists and eight antagonists, as summarized in Table 1. The binding affinity of agonists was reduced by substitutions on either the N6- or the C2 position of adenine. NECA, which is unsubstituted on the N6 or the C2 position, binds to A2BARs with more than 60-fold higher affinity than any of the other agonists tested.

Equilibrium binding of125I/127I-ABOPX to membranes derived from HEK-A2B cells. A, Specific (▪) and nonspecific equilibrium binding (■) were determined with isotope dilution as described in the methods. B, Scatchard transformation of specific binding. The Bmax value is 21,400 fmol/mg protein; the KD value is 37 nM. Each point is the mean ± S.E.M. of triplicate determinations. The results are typical of quadruplicate experiments.
Competition by various compounds for 125I-ABOPX binding to recombinant human A2BARs
Among antagonists, the xanthines used therapeutically to treat asthma, enprofylline and theophylline, were both found to compete for125I-ABOPX binding to A2BARs with KIvalues <10 μM (Fig. 4A; Table2). It is notable also that ZM241385 and CPX (Fig. 4A), compounds reported to be selective antagonists of A2AARs (Poucher et al., 1995) and A1ARs (Lohse et al., 1993), respectively, bind with high affinity to A2BARs. The adenosine receptor subtype selectively of I-ABOPX, ZM241385, CPX, theophylline, and enprofylline were determined in radioligand binding assays for the four AR subtypes (Table 2). If A2BARs are excluded, ZM241385 and CPX are highly selective for A2AARs and A1ARs, respectively, as reported previously. However, ZM241385 has only 22.1-fold selectivity for human A2A over A2B receptors, and CPX has only 22.4-fold selectivity for A1 over A2BARs. The high potency of CPX for A2B receptors was confirmed in CHO-A2B cells (Fig. 4B). The stable expression of recombinant human A2B receptors in CHO cells was lower than in HEK cells and could not be accurately measured by radioligand binding. Nevertheless, NECA stimulated cAMP accumulation in CHO-A2B cells, whereas it had only a slight effect in untransfected cells. By Schild analysis, CPX blocked the action of NECA in CHO-A2B cells with aKI of 22 nM (Fig. 4B). The high affinity of ZM241385 also has been confirmed. In a recent study, [3H]ZM241385 was found to bind to recombinant human A2B receptors with aKD of 34 nM (Ji and Jacobson, 1999).

A, competition by the enprofylline and theophylline for 125I-ABOPX binding to HEK-A2B membranes. Each tube contained 208,000 cpm (0.56 nM) 125I-ABOPX and 20 μg of membrane protein. Each point is the mean ± S.E.M. of triplicate determinations; where omitted, the S.E.M. is smaller than the symbols. KI values derived from triplicate experiments are summarized in Table 1. B, competition by CPX to inhibit NECA-stimulated cyclic AMP accumulation in CHO-A2B cells. The concentration of CPX (nm) is O (●), 30 (•), 150 (▴), and 750 (♦). The inset is a Schild plot, the pA2 is 7.66.
Adenosine receptor subtype selectivity of antagonists
Schild Analyses of Theophylline and Enprofylline Effects.
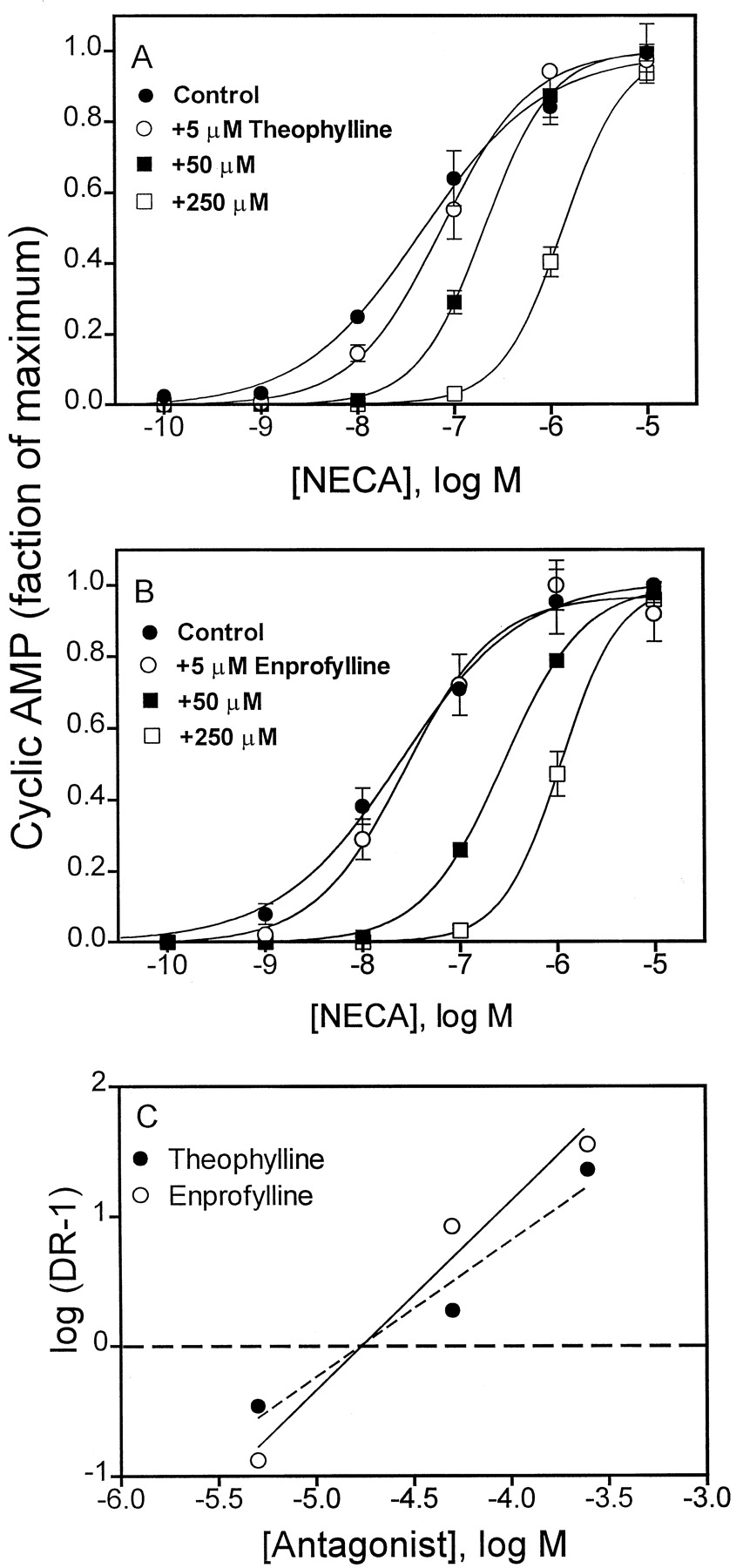
We next set out to estimate KI values from pA2 values for enprofylline and theophylline as antagonists of NECA-stimulated cAMP accumulation in HEK-A2Bcells to determine whether this estimate of binding affinity agrees with KI values derived by radioligand binding assays. As illustrated in Fig. 5,KI values for theophylline and enprofylline are both 10 to 17 μM, in reasonably good agreement with the binding constants reported in Table 1. Enprofylline and theophylline caused an increase in the slope of NECA does response curves. It is notable that basal cAMP in HEK-A2B cells was dose-dependently decreased by either theophylline or enprofylline from a level of about 20 pmol/ml to about 6 pmol/ml. This may be because of a small degree of constitutive activity by the overexpressed receptor, or the effects of endogenous adenosine released by the cells and not degraded by added adenosine deaminase. Inhibition of phosphodiesterase by these xanthines may also contribute to this effect on the slope of the NECA dose–response curve.

Schild analysis of theophylline and enprofylline to inhibit NECA-stimulated cAMP accumulation in HEK-A2B cells. A, competition by theophylline. B, competition by enprofylline. Each point is the mean ± S.E. of triplicate determinations. Basal levels for cAMP ranged from 3 to 20 pmol/ml; the maximum response to NECA was 240–270 pmol/ml. C, Schild analyses of the data derived from A and B; pA2 values for theophylline and enprofylline ranged from 10 to 17 μM in triplicate experiments.
Potency Order of Agonists in cAMP Assays.
As a further validation of the new radioligand binding assay, we determined the potencies of three agonists in cAMP assays. The absolute potencies of all three agonists is greater in cAMP than in radioligand binding assays, but the ratio of affinities derived from functional and binding assays is similar. Thus, IB-MECA is 169- and 162-fold less potent than NECA in functional and binding assays, respectively. CGS21680 is 530- and 1090-fold less potent than NECA.
Coupling of A2BARs to Phospholipase C.
The activation of A2BARs in HEK-A2B cells was found to increase the cellular content of IP3. This response was not affected by pretreating the cells with pertussis toxin (Fig. 6A). In contrast, pertussis intoxication to inactivate Gi and Go family G proteins abolished the ability of IB-MECA to stimulate phospholipase C in HEK-A3cells (Fig. 6B). The data are consistent with the idea that A3 receptors are coupled to phospholipase C via the βγ subunits of Gi/o G proteins, whereas A2B receptors are coupled to phospholipase C via Gq/11. A dose-response curve for NECA to stimulate IP3 in HEK-A2Bcells is shown in Fig. 6C. The ED50 value for NECA, 19 nM, is close to its ED50 to stimulate cAMP accumulation (29 nM; Fig. 7). We considered the possibility that there might be cross-talk between the cAMP and IP3 pathways in HEK cells. However, as illustrated in Fig. 8, two activators of adenylyl cyclase, isoproterenol and forskolin, increase cAMP levels in HEK-A2B cell and minimally influence IP3 levels, whereas UTP increases IP3 but not cAMP. The effects of UTP to stimulate phospholipase C or NECA to stimulate cAMP accumulation are not affected by pretreatment of cells with pertussis toxin (data not shown). The data are consistent with the interpretation that A2BARs are dually coupled to Gs and to Gq/11.

Stimulation of phospholipase C by recombinant adenosine receptors. HEK-A2B (A) or HEK-A3 (B) cells were pretreated for 16 h without or with 100 ng/ml pertussis toxin (PTX) and then stimulated with NECA or IB-MECA. Control concentrations of [3H]IP3 were 500–600 cpm/tube. C, dose-response curve for NECA to stimulate [3H]IP3 accumulation in HEK-A2Bcells. The ED50 value of NECA is 19 nM. Each bar or point is the mean ± S.E. of triplicate determinations. The results are typical of three to five experiments.

Dose-response curves of agonists to stimulate cAMP accumulation in HEK-A2B cells. Each point is the mean ± S.E. of triplicate determinations. ED50 values are: NECA, 28.6 nM; IB-MECA, 4.8 μM; CGS21680, 15.2 μM. The results are typical of triplicate experiments.

Stimulation of IP3 (A) and cAMP (B) accumulation by various compounds in HEK-A2B cells. Cells were treated with 1 μM isoproterenol (ISO), 10 μM forskolin (Forsk), 10 μM UTP, or 10 μM NECA, and cAMP and IP3accumulation were determined as described in Experimental Procedures. Each bar is the mean ± S.E. of duplicate (IP3) or triplicate (cAMP) determinations. The results are typical of three to four replicate experiments.
The coupling of A2BARs to PLC was further characterized by measuring calcium mobilization in response to A2BAR activation in HEK-A2Bcells. Figure 9 shows that the response to NECA is not affected by intoxication of cells with pertussis toxin but is largely attenuated by 100 μM theophylline or enprofylline. The data are consistent with the interpretation that NECA stimulates calcium mobilization as a consequence of activating A2BARs that are coupled to Gq/11.

NECA stimulation of calcium mobilization in HEK-A2B cells. Cells in suspension (106/ml) were stimulated with 1 μM NECA (arrows). A, control cells or cells pretreated with 200 ng/ml pertussis toxin (PTX) for 18 h. B, control cells or cells pretreated with enprofylline or theophylline for 5 min. The results are typical of triplicate experiments.
Coupling of A2BARs to Calcium Mobilization in Human Mast Cells.
Adenosine stimulates the degranulation of rodent and canine mast cells by activating A3 and A2B receptors, respectively (Ramkumar et al., 1993; Auchampach et al., 1997; Jin et al., 1997). A2BARs are thought to be responsible for triggering interleukin-8 release in the human mast cell tumor line, HMC-1 (Feoktistov and Biaggioni, 1995). Figure10 shows that calcium mobilization and cAMP accumulation in HMC-1 cells are stimulated with a potency order of NECA > IB-MECA. The response to NECA was not affected by pretreating HMC-1 cells with pertussis toxin. These data are consistent with the involvement of an A2BAR in human mast cell calcium mobilization. However, we could not detect specific125I-ABOPX binding to HMC-1 cell membranes, which suggests that A2B receptor density is too low to be detected with this radioligand.

Stimulation of calcium and cAMP accumulation in HMC-1 mast cells. A, calcium mobilization in response to NECA or IB-MECA added at the arrow. B, concentration response curves for cAMP accumulation. Each point is the mean ± S.E. of triplicate determinations. The results are typical of triplicate experiments.
Discussion
Physical Properties of A2B Adenosine Receptors.
In this study, we have characterized the physical, pharmacological, and coupling characteristics of recombinant human A2BARs. Based on their cDNA sequences, the wild-type and H/F-A2BARs are predicted to have molecular masses of 36 and 38.5 kDa, respectively. There are consensus sites for N-linked glycosylation on asparagines 153 and 163 in the second extracellular loop of the human receptor. The first site is conserved in the A2B sequences of all species that have been cloned to date (Linden and Jacobson, 1998). In a previous study, Western blots conducted with antipeptide antibodies derived from the human A2BAR sequence detected immunoreactivity in 50- to 55-kDa nonglycoproteins found in various human tissues (Puffinbarger et al., 1995). Hence, we considered the possibility that the antipeptide immunoreactivity might detect a cross-reacting epitope on a nonreceptor protein. To explore this possibility, the physical properties of the purified H/F-A2BAR were investigated. Figure 1 indicates that the purified H/F-A2BAR has characteristics close to those expected from the cDNA sequence (i.e., by western blotting with anti-FLAG antibodies it appears as a 35-kDa glycoprotein). These data imply that the immunoreactivity previously detected in tissues may not represent the A2BAR.
125I-ABOPX Binding to A2BARs.
125I-ABOPX is described here as a new and improved radioiodinated ligand for the detection recombinant A2BARs. It is nonselective; hence, it cannot be used to detect A2BAR on cells or tissues that possess multiple subtypes of adenosine receptors. It has an affinity (KD = 37 nM) that is low enough to require isotope dilution to saturate binding sites. The radioligand cannot be used to detect low levels of endogenous receptors on untransfected HEK cells or HMC-1 mast cells. Nevertheless, the results indicate that125I-ABOPX binds with a high signal-to-noise ratio to recombinant A2BARs on HEK-A2B cells. When used in conjunction with a new HEK-A2B cell line that has a high density of A2BARs (in excess of 20,000 fmol/mg protein), specific binding of 125I-ABOPX is >80% of total binding (Fig. 4). Hence this compound can used efficiently to screen competing ligands in competition radioligand binding assays.
Agonist Binding to A2BARs.
The results of competition binding assays generally confirm the assessment, based on functional data, that A2BARs have a relatively low affinity for agonists (Daly et al., 1983). The affinity of agonists for the A2BAR is reduced by the presence of bulky substituents on either the N6- or the C2-position of the adenine ring. Thus,N6-aminobenzyl-NECA binds with 65-fold lower affinity than NECA. The introduction of a relatively small chlorine atom at the C2-position has a minor effect on affinity, thus 2-Cl-CPA binds with only 1.17-fold lower affinity than CPA. However, CGS21680, which consists of NECA with a bulky 2-aralkyl substituent, binds with an affinity more than 1000 times lower than NECA. The data also are consistent with the previous observation that 2-phenylethoxy-9-methyladenine is a 2-substituted adenine-analog antagonist that discriminates well between A2Areceptors on coronary arteries and A2B receptors on the aorta (Martin et al., 1993). The introduction ofN-ethylcarboxamide on the 5′-position of the ribose ring of adenosine moderately enhances agonist affinity for A2BARs inasmuch as the affinity ofN6-aminobenzyl-5′-NECA (AB-NECA) is 5.8-fold higher than the affinity of the same compound lacing the 5′ substitution, ABA.
Antagonist Binding to A2BARs.
KI values for antagonists noted in this study generally agree with literature KIestimates for human receptors based on functional assays (Brackett and Daly, 1994; Alexander et al., 1996; Cooper et al., 1997). We found in this study that A2BARs have a high affinity for CPX and ZM241385. It is significant in this regard that CPX or ZM241385 are often used in the concentration range of 0.1 to 1 μM to selectively block A1 or A2Areceptors, respectively. At 1 μM, either of these antagonists would occupy more than 90% of human A2BARs. TheKI value of enprofylline for binding to A2B receptors, 6 μM, agrees closely with functional estimates of KI value based in inhibition of agonist-stimulated cAMP accumulation in NIH-3T3 cells (Brackett and Daly, 1994) and HEL cells (Feoktistov and Biaggioni, 1995). It was noted in 1982 that the KIvalue of enprofylline needed to inhibit cAMP accumulation in rat hippocampal slices, approximate 7 μM, is 20 times more potent than the KI for binding to rat A1 receptors. Hence, it seems likely that the receptor responsible for cAMP accumulation in hippocampal slices is A2B.
Coupling Characteristics of A2BARs.
We demonstrate in this study that the over-expressed recombinant human A2BARs in HEK 293 cells couple not only to cAMP accumulation, but also to phospholipase C activation and calcium mobilization. It is possible that the coupling of recombinant receptors to one or both of these signaling pathways could be an artifact of receptor over-expression, resulting in indiscriminate coupling of recombinant receptors to various HEK cell G proteins. However, there is an endogenous A2BAR on HEK 293 cells (Townsend-Nicholson, 1997) that also couples to the accumulation of cAMP and calcium as well as to the activation of the mitogen-activated protein kinase isoform extracellular-signal regulated kinase (Gao et al., 1999). We provide evidence here that calcium mobilization in response to NECA in HEK-A2B cells is not a result of cAMP accumulation (Fig. 8); rather, it is probably caused by the direct coupling of A2BARs to Gq/11.
The ED50 values of NECA required to stimulate cAMP accumulation in untransfected HEK and HEK-A2B cells are 2.7 μM (Gao et al., 1999) and 28.6 nM (Fig. 7), respectively, a difference of approximately 100-fold. Spare receptor theory dictates that under some circumstances the potency of an agonist in functional assays is proportional to receptor density (Nickerson, 1956; Nickerson, 1956). If this relationship holds for the endogenous A2BAR of HEK cells versus HEK-A2B cells, than the endogenous level of receptor expression in HEK cells can be approximated as about 200 fmol/mg protein. This should be considered a high estimate because lower levels of receptors are likely to have coupling efficiency greater than (or similar to) overexpressed receptors.
A2BARs and Mast Cell Function.
The A2BAR has been implicated as the adenosine receptor that is responsible for stimulating the degranulation of canine BR mastocytoma cells (Auchampach et al., 1997) and for triggering the slow release of interleukin-8 from human HMC-1 mast cells (Feoktistov and Biaggioni, 1995). In this study, we show that A2BARs also trigger a rapid mobilization of calcium in HMC-1 cells. The agonist potency order (NECA > IB-MECA) is consistent with calcium mobilization in HMC-1 cells being mediated by an A2BAR. It is notable that a different AR subtype, the A3AR, mediates the degranulation of hamster perivascular mast cells (Shepherd et al., 1996; Jin et al., 1997) and rat RBL-2H3 mast-like cells (Ramkumar et al., 1993). It is calcium mobilization rather than cAMP accumulation that is likely to trigger mast cell degranulation in response to A2BAR activation, because cAMP seems to inhibit mast cell degranulation (Church and Hughes, 1985; Hughes and Church, 1986; Hughes et al., 1987; Jin et al., 1997).
Xanthines in the Treatment of Asthma.
We found that enprofylline and theophylline block human A2BARs with KI values in the range of 6 to 8 μM. Optimal plasma levels of theses compounds for the treatment of asthma have been reported to be 4 μg/ml (19 μM) for enprofylline, and 10 μg/ml (55 μM) for theophylline (Vilsvik et al., 1990). Both enprofylline and theophylline have lower affinities for the A3AR than for the other human adenosine receptor subtypes. Theophylline is nonselective among A1, A2A, and A2B subtypes, whereas enprofylline is somewhat selective for the A2BAR subtype (Table 2). Blockade of A2AARs by theophylline may be counterproductive in the treatment of asthma because activation of A2AARs has inhibitory effects on mast cells and several other types of inflammatory cells (Sullivan and Linden, 1998). Adverse side effects of theophylline that are not shared by enprofylline may be mediated principally by A1AR blockade. These include diuresis, free fatty acid release, gastric secretion, and central nervous system stimulation (Persson, 1986).
The observation that enprofylline is effective in the treatment of human asthma (Chapman et al., 1990, 1994) suggests that activation of the A2BAR may contribute to bronchoconstriction in human disease. The radioligand 125I-ABOPX cannot be used to detect low levels of endogenous receptors on untransfected HEK cells or HMC-1 mast cells. Nevertheless, the availability of improved radioligand binding assays described here should facilitate the search for new A2B-selective antagonists that may be useful for the treatment of asthma.
Acknowledgments
We thank Dr. J. H. Butterfield (The Mayo Clinic, Rochester, MN) for his kind gift of HMC-1 mast cells and Melissa Marshall for technical assistance.
Footnotes
-
Send reprint requests to: Dr. Joel Linden, Departments of Internal Medicine and Molecular Physiology, Box MR4 6012, Room MR4 6071, Health Sciences Center, University of Virginia, Charlottesville, Virginia. E-mail:jlinden{at}virginia.edu
-
This work was supported by National Institutes of Health Grant RO1-HK37942.
- Abbreviations:
- AR
- adenosine receptor
- DPX
- 1,3-diethyl-8-phenylxanthine
- CPX
- 8-cyclopentyl-1,3-dipropylxanthine
- XAL
- 8-(4-((2-aminoethyl)aminocarbonyl-methyloxy)phenyl)-1,3-dipropylxanthine
- ABOPX
- 3-(3,4-aminobenzyl)-8-(4-oxyacetate)phenyl-1-propyl-xanthine
- H/F-A2B
- recombinant human A2B adenosine receptors extended on the amino terminus with hexahistidine and the FLAG epitope (DYKDDDDK)
- NECA
- 5′-N-ethylcarboxamidoadenosine
- CPA
- N6-cyclopentyladenosine
- CGS21680
- 2-[4-(2-carboxyethyl)phenethylamino]-5N-N-ethylcarboxamidoadenosine
- XAC
- 8-(4-((2-aminoethyl)aminocarbonylmethyloxy)phenyl)-1,3-dipropylxanthine
- ABA
- N6-aminobenzyladenosine
- BW-A1433
- 8-(4-carboxyethenylphenyl)-1,3-dipropylxanthine
- AB-NECA
- aminobenzyl-5′-N-ethylcarboxamidoadenosine
- DMEM
- Dulbecco’s modified Eagle’s medium
- APE
- 2-[2-(4-amino-phenyl)ethylamino]adenosine
- TBS/T
- Tris-buffered saline/Tween 20
- CHO
- Chinese hamster ovary
- IP3
- inositol trisphosphate
- IB-MECA
- N6-(2-iodo)benzyl-5′-N-methylcarboxamidodoadenosine
- Received March 5, 1999.
- Accepted June 21, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}