


Abstract
A family of G protein-coupled P2Y receptors that are activated by adenine and uridine nucleotides has been identified recently. Degenerate primers based on conserved sequences in these P2Y receptors were used to amplify turkey DNA, which was used to isolate the complete coding sequence of a cDNA that encodes a novel G protein-coupled receptor. Stable expression of this avian cDNA in 1321N1 human astrocytoma cells resulted in the conveyance of marked inositol phosphate responses to various nucleotides. Although this cloned avian receptor exhibited its highest homology to the previously cloned mammalian P2Y4 receptor, its pharmacological selectivity was not consistent with the avian receptor’s being a species homologue of the P2Y4 receptor. That is, whereas the P2Y4receptor is selectively activated by UTP and is not activated by ATP or AP4A, the novel avian receptor was potently activated by ATP and AP4A as well as by UTP. Taken together, these results describe the identification of an avian phospholipase C-coupled P2Y receptor that, like the mammalian P2Y2 receptor, is activated by both adenine and uridine nucleotides.
Extracellular adenine and uridine nucleotides modulate a variety of physiological responses (1, 2) through interaction with broadly distributed P2X and P2Y receptors (3, 4). P2X receptors are ligand-gated ion channels that are activated by adenine nucleotides, and P2Y receptors are G protein-coupled receptors that are activated by both adenine and uridine nucleotides (2).
Four G protein-coupled P2Y receptors (P2Y1, P2Y2, P2Y4, and P2Y6 receptors) have been cloned from mammalian species and shown to encode nucleotide-activated receptors in functional studies after transient or stable expression (5). These receptors share 35–54% sequence identity and are selectively activated by adenine nucleotides (P2Y1 receptor) (6, 7), by uridine nucleotides (P2Y4 and P2Y6 receptors) (8-11), and by both adenine and uridine nucleotides (P2Y2 receptor) (11, 12).
Three additional G protein-coupled receptors have been proposed to be members of the P2Y receptor family. The p2y31 receptor is a P2Y receptor originally cloned from a chick brain cDNA library (13). The amino acid sequence identity (65%) between the chick p2y3 and mammalian P2Y6 receptors is high, with long stretches of >80% identity. The pharmacological selectivity of the p2y3 receptor is also remarkably similar to that of the mammalian P2Y6 receptor, and both receptors are selectively activated by UDP.2 No mammalian homologue of the avian p2y3 receptor has been reported, and based on their similarities in sequence and activity, it is likely that the p2y3 receptor is the avian homologue of the mammalian P2Y6 receptor. The proposed p2y5 receptor (14) was originally cloned from activated chick T-lymphocytes as an orphan receptor designated the 6H1 receptor (15), and the proposed human p2y7 receptor was cloned recently from a human erythroleukemia cell line (16). Both of these G protein-coupled receptors were suggested to be members of the P2Y receptor family largely on the basis of binding studies with [35S]dATPαS (14, 16), which was shown recently not to be a general radioligand for P2Y receptors (17). Moreover, these receptors exhibit predicted amino acid sequences that are only 28–32% identical to that of known P2Y receptors, and nucleotide-promoted stimulation of a second-messenger signaling response is not observed after expression of either of these G protein-coupled receptors (18-20). The protein originally referred to as the p2y7 receptor has been shown recently to be a receptor for leukotrienes rather than for purine or pyrimidine nucleotides (20).
In this article, we report the cloning and expression of the cDNA of a novel avian G protein-coupled receptor that exhibits its highest homology to the previously reported UTP-selective P2Y4 receptor. This receptor is clearly a member of the P2Y receptor class of signaling proteins based on observation of nucleotide-promoted inositol lipid signaling responses in cells infected with the avian cDNA. However, the pharmacological selectivity of the expressed avian receptor differs markedly from that of the P2Y4 receptor and reveals the identification of an avian receptor that, like the mammalian P2Y2receptor, is activated by both adenine and uridine nucleotides.
Experimental Procedures
Materials.
ATP and UTP were obtained from Pharmacia (Piscataway, NJ). 2MeSATP, 2MeSADP, and pyridoxal-phosphate-6-azophenyl-2′,4′-disulfonic acid were from Research Biochemicals International (Natick, MA). Hexokinase and all other nucleotides were from Boehringer Mannheim Biochemicals (Indianapolis, IN). Type I potato apyrase and AP4A were from Sigma Chemical (St. Louis, MO).myo-[2-3H]-inositol (20 Ci/mmol) was from American Radiolabeled Chemicals (St. Louis, MO). DNA sequencing kit AmpliCycle was from Perkin Elmer (Branchburg, NJ). All tissue culture reagents were from the Lineberger Comprehensive Cancer Center tissue culture facilities at the University of North Carolina.
The purity of ATP and UTP used in this study was higher than 99% as estimated by high performance liquid chromatography. Stock solutions of AP2A, AP3A, and AP4A were treated with apyrase at 10 units/ml, and the assay of these dinucleotides was carried out in the presence of 1 unit of apyrase/well. Similarly, all nucleotide diphosphate stock solutions were treated with 10 units/ml hexokinase and 25 mm glucose, and the assays were performed in the presence of 1 unit of hexokinase/well.
PCR amplification with degenerate primers and isolation of a novel cDNA.
With the goal of identifying sequences encoding novel P2 receptors, we carried out PCR amplifications using degenerate oligodeoxynucleotide primers corresponding to the third, sixth, and seventh transmembrane domains conserved between the P2Y1 and P2Y2 receptors. A novel sequence was amplified by PCR from turkey DNA using 5′-TT(CT)(CT)TIACITG(TC)ATI(AT) (GC)IG(CT)ICA-3′ as a sense primer and 5′-GC(GA)AAIACIG(TC)IA(AG)IAC-3′ as an antisense primer. Because this sequence exhibited high homology to members of the superfamily of G protein-coupled receptors, we decided to obtain the full-length clone and to express the receptor to study its pharmacological properties.
A turkey blood cDNA library was constructed using the λ-Zap cDNA synthesis kit (Stratagene, La Jolla, CA) as described in detail previously (21). A 325-bp DNA fragment of the novel turkey sequence (corresponding to the region between the predicted third and sixth transmembrane domains) was used as a probe for the isolation of the full-length clone. The probe was radiolabeled with [α32P]dCTP (Amersham, Arlington Heights, IL) by the random priming method and used to screen approximately 1.2 × 106 plaque-forming units of the turkey blood cDNA library. Nylon filters (Hybond N+; Amersham) containing plaque lifts were hybridized at 42° in 2× PIPES buffer (0.8 m NaCl, 20 mm PIPES, pH 6.5), 50% formamide, 0.5% sodium dodecyl sulfate, and 100 μg/ml salmon sperm DNA. The filters were washed with 0.1% standard saline citrate (15 mm sodium citrate, 0.15 m NaCl, pH 7.0), 0.5% sodium dodecyl sulfate at 55°, followed by autoradiography. Eight hybridizing clones were plaque-purified, and the pBluescript-containing insert of one of these positive clones was rescued from the purified λ phage by in vivo excision using the Exassist/SOLR system (Stratagene) and was sequenced in both strands by the dideoxynucleotide chain termination reaction using the AmpliCycle sequencing kit (Perkin-Elmer Cetus, Norwalk, CT).
Construction of retroviral vectors for the avian P2Y receptor.
Two oligonucleotide primers were designed to amplify the open reading frame of the novel avian cDNA. The upstream primer contained an EcoRI site in the 5′ end and included 31 bp of 5′-untranslated sequence. The downstream primer was designed to contain a XhoI site at the 5′ end, and to include 135 bp of 3′-untranslated sequence. PCR amplification was carried out for 30 amplification cycles using Pfu DNA polymerase (Stratagene). The amplified fragment was purified, digested with EcoRI andXhoI, and ligated with the similarly digested retroviral expression vector pLXSN.
Receptor expression in 1321N1 human astrocytoma cells.
A recombinant retrovirus harboring the coding sequence for the novel avian receptor was produced by transfection of the amphotrophic packaging cells PA317. Transfection was achieved by the calcium phosphate precipitation method as described previously (22). After incubation of transfected PA317 cells for 48 hr at 32° in the presence of 5 mm butyrate, the cell supernatant containing packaged retroviruses was collected, filtered, and used to infect 1321N1 human astrocytoma cells. Infection was carried out for 2 hr in the presence of 8 μg/ml polybrene. After 48 hr, infected 1321N1 cells were selected for geneticin resistance with 600 μg/ml G-418 until mock-infected cells treated with the same concentration of G-418 died. Clonal cell lines that stably expressed the avian P2Y receptor were obtained by plating serial dilutions of geneticin-resistant cells. Cell populations originating from single cells were expanded and screened for receptor expression.
Cell culture.
The murine packaging cell line PA317 was grown in DMEM containing 4 g/liter glucose, 10% fetal bovine serum, and no antibiotics. 1321N1 human astrocytoma cells that stably expressed the avian receptor were grown in DMEM supplemented with 5% fetal bovine serum and 600 μg/ml G-418. All cells were maintained at 37° in a humidified atmosphere of 5% CO2/95% air except where indicated.
Phosphoinositide hydrolysis assays.
1321N1 human astrocytoma cells were seeded in 48-well plates and assayed 3–4 days after subculture. Twenty four hours before the assay, the inositol lipid pool was radiolabeled by incubation in 200 μl of serum-free, inositol-free DMEM containing 2 μCi/mlmyo-[3H]inositol. Before the assay, cells were supplemented with 40 mm (final concentration in the assay) HEPES, pH 7.4, placed in a 37° water bath, and supplemented with 10 mm LiCl (final concentration). Ten minutes after LiCl addition, cells were challenged with agonists for an additional 10 min. Incubations were terminated by aspiration of the drug-containing medium and addition of 500 μl of boiling 10 mm EDTA, pH 8.0. [3H]Inositol phosphates were isolated by anion exchange chromatography on Dowex AG1-X8 columns as previously described (23).
Results
A novel cDNA was cloned from a turkey blood cDNA library as detailed in Experimental Procedures. This 2000-bp insert was sequenced in both strands, which revealed an open reading frame of 1122 bp encoding a 374-amino acid protein. Fig. 1shows the nucleotide sequence and deduced amino acid sequence. Two potential initiation codons (nucleotides 374–376 and 392–394 in Fig.1) are preceded in the nucleotide sequence by an in-frame stop codon. Based on the consensus translation initiation sequence proposed by Kozak (24), the first initiation codon most likely represents the amino terminus of the receptor protein. The second ATG codon matches with the initiation codon in the best alignment with the most homologous protein, the human P2Y4 receptor (see below). The predicted molecular mass of the protein (with the first ATG codon) is 42.59 kDa. Hydropathy analysis revealed seven stretches of hydrophobic amino acids corresponding to the putative transmembrane domains characteristic of G protein-coupled receptors. The protein encoded by this cDNA includes potential N-glycosylation sites (Asn24) in the amino terminus and in the second extracellular loop (Asn187). Potential protein kinase C phosphorylation sites occur in the second intracellular loop (residue 149) and in the third intracellular loop (residues 242 and 252). A potential protein kinase A phosphorylation site also is found at residue 247 in the third intracellular loop. The predicted amino-acid sequence includes a number of residues conserved in most of the G protein-coupled receptors, including two conserved cysteine residues in the putative first and second extracellular loops that might be involved in a disulfide bond, as well as proline residues in transmembrane domains 4, 5, 6, and 7 that occur as conserved residues in all G protein-coupled receptors. The four positively charged amino acids in transmembrane segments VI and VII that have been suggested to play a role in nucleotide binding of P2Y receptors (25) are also conserved in the avian receptor.

Nucleotide and deduced amino acid sequence of the turkey P2Y receptor DNA. Bold, two potential initiation codons; underlined, seven putative transmembrane domains; ▪, two potential N-linked glycosylation sites; ∗, potential phosphorylation sites. This nucleotide sequence has been submitted to the GenBank database with accession number AF031897.
The protein encoded by the turkey cDNA is most homologous with members of the P2Y receptor family of signaling proteins (Fig.2). The sequence SILFLTCISVHR found in the predicted third transmembrane domain of the novel avian receptor is highly conserved in all four of the mammalian P2Y receptors. Among the P2Y receptors, the highest amino acid identity (56%) was with the human P2Y4 receptor followed by the human P2Y2 receptor (47%), chick p2y3 receptor (38%), turkey P2Y1 receptor (38%), human P2Y1 receptor (38%), and human P2Y6 receptor (35%).

Alignment of deduced amino acid sequences of the turkey P2Y receptor, and the human P2Y4, P2Y2, P2Y1, and rat P2Y6 receptors. Shaded areas indicate amino acid identity to the turkey receptor. Gaps (−) were introduced to improve the alignment.
The retroviral vector pLXSN alone or vector engineered to contain the novel avian cDNA was transfected in the viral packaging cell line PA317, and retroviruses from these constructs were used to infect 1321N1 human astrocytoma cells. These cells do not express endogenous P2Y receptors, and therefore, have been broadly used by us and others for the heterologous expression of P2Y receptors (7, 9, 11, 26, 27). Clonal cell lines derived from populations of infected cells were isolated and tested for receptor expression.
Expression of the avian receptor in 1321N1 cells resulted in high basal levels of [3H]inositol phosphates. These elevated basal levels were reduced by 30 ± 5 and 70 ± 5% by treatment of the cells with 1 unit/ml hexokinase or 1 unit/ml apyrase, respectively. These results suggest that basal activation of the expressed receptor occurred as a consequence of release of cellular ATP, as we have shown previously to be the case in studies of the cloned P2Y1 (26) and P2Y2(28) receptors. Addition of 0.3 μm or 100 μm UTP or ATP to the medium resulted in marked increases in [3H]inositol phosphate accumulation in 1321N1 cells expressing the avian receptor (Fig.3). In contrast, nucleotides had no effect on the accumulation of [3H]inositol phosphates in 1321N1 cells infected with retrovirus construct containing empty pLXSN vector. These results demonstrate that the protein encoded by the turkey cDNA exhibits the sensitivity to activation by nucleotide molecules that is characteristic of the P2Y receptor family of signaling proteins.

Effects of ATP and UTP on 1321N1 cells expressing the turkey P2Y receptor. [3H]Inositol phosphate accumulation was measured in response to 0.3 and 100 μmATP, and 0.3 and 100 μm UTP or 1 mm carbachol in 1321N1 cells infected with pLXSN (A) or pLXSN harboring the turkey P2Y receptor cDNA (B). The results are presented as mean ± standard error from a representative experiment carried out in triplicate and repeated at least three times with similar results.
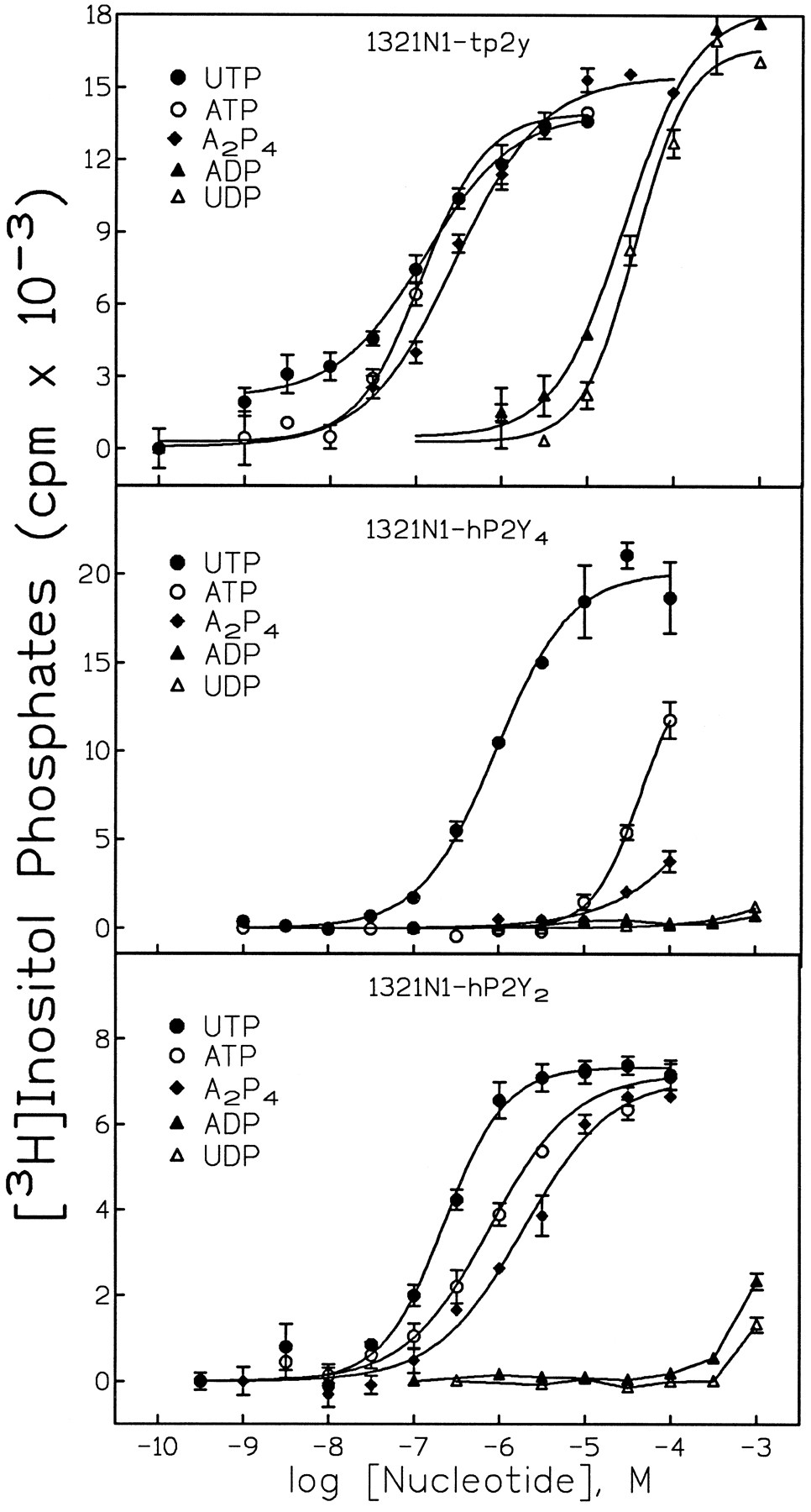
To define the pharmacological selectivity of the avian receptor, concentration-effect curves for adenine and uridine nucleotides were generated in 1321N1 cells expressing this receptor. UTP and ATP were the most potent agonists, with EC50 values of 103 ± 20 and 128 ± 20 nm, respectively (Fig.4; Table1). UDP and ADP were also full agonists. However, the EC50 values of the diphosphate molecules were 500- to 1000-fold higher than those of the triphosphates. The diadenosine polyphosphate AP4A was also a potent full agonist with an EC50 value of 1.31 ± 0.56 μm. In contrast, AP3A and AP2A were inactive (data not shown). The rank order of agonist potency for the avian receptor was UTP = ATP > AP4A > ATPγS > App(NH)p > 2MeSATP > UDP = ADP (Table1). 2MeSADP, ADPβS, AMP, adenosine, α,β-methylene-ATP, and β,γ-methylene-ATP were inactive (Table 1; data not shown).

Effect of adenine and uridine nucleotides on inositol lipid hydrolysis in 1321N1 human astrocytoma cells stably expressing the turkey P2Y receptor, the human P2Y4receptor, or the human P2Y2 receptor. The capacity of the indicated concentrations of ATP (○), UTP (•), AP4A (♦), ADP (▴), or UDP (▵) to stimulate the hydrolysis of inositol lipids by 1321N1 human astrocytoma cells stably expressing the turkey P2Y receptor (A), the human P2Y4 receptor (B), or the human P2Y2 receptor (C) was determined as described in Experimental Procedures. The data shown are the mean ± standard error of triplicate assays from a representative experiment repeated at least three times.
Pharmacological selectivity of the avian P2Y receptor stably expressed in 1321N1 human astrocytoma cells.
We recently reported that adenosine 3′,5′-bisphosphate is a selective P2Y1 receptor antagonist (23). Suramin and pyridoxal-phosphate-6-azophenyl-2′,4′-disulfonic acid also have been shown previously to be competitive antagonists of the P2Y1 and other P2 receptors. None of these three molecules at 100-μm concentration antagonized the stimulatory effects of UTP in 1321N1 cells expressing the avian P2Y receptor (data not shown).
The pharmacological selectivity of the avian P2Y receptor was compared with that of previously cloned P2Y receptors by generating a series of agonist concentration-effect curves in 1321N1 cells stably expressing the avian receptor, the human P2Y4 receptor, or the human P2Y2 receptor. Although the highest sequence homology of the avian receptor was observed with the human P2Y4 receptor, the pharmacological selectivities of these two receptors were remarkably different (compare Fig. 4A and Fig. 4B). That is, whereas the human P2Y4 receptor is a UTP-selective receptor, the avian receptor was potently activated by both adenine and uridine nucleotide triphosphates and by the diadenosine polyphosphate molecule AP4A. Surprisingly, in spite of a relatively lower homology, the pharmacological selectivity of the avian receptor was much more similar to that of the human P2Y2receptor than to that of the human P2Y4 receptor. The avian and human P2Y1 receptors, which are activated by adenine but not by uridine nucleotides or AP4A [(28); data not shown], and the human P2Y6 receptor, which is selectively activated by UDP (11), exhibit pharmacological selectivities that differ markedly from that of this novel avian P2Y receptor.
Discussion
A cDNA has been isolated from a turkey blood cDNA library that encodes a 374-amino-acid protein exhibiting sequence similarity with confirmed members of the P2Y receptor family. The predicted amino acid sequence of this G protein-coupled receptor has the conserved 10–12 amino acid sequence found in the third transmembrane region of all of the cloned P2Y receptors, and exhibits other features common to all G-protein coupled receptors. Stable expression of this avian receptor in 1321N1 human astrocytoma cells conferred inositol phosphate responses to nucleotides.
Although the sequence similarity of this avian P2Y receptor compared with the mammalian P2Y receptors is highest with the UTP-selective human P2Y4 receptor, it is apparently not a species homologue of the P2Y4 receptor. That is, the novel avian receptor is activated by adenine nucleotides as well as by UTP. Indeed, the pharmacological selectivity exhibited by the avian receptor indicates that we have identified a receptor that is activated by both uridine and adenine nucleotides, as is the mammalian P2Y2 receptor. Although this avian P2Y receptor could be, in fact, a species homologue of the P2Y2 receptor, this seems unlikely, because it is more homologous to the P2Y4 receptor, and its sequence identity with the human P2Y2 receptor is less than 50%. The other possibilities, that this is a novel receptor for which a yet-to-be cloned mammalian homologue exists or that this is a novel avian P2Y receptor for which no clear mammalian counterpart exists, remain to be directly addressed.
It will be important to determine rigorously whether a sequence analogous to the avian receptor can be cloned from a mammalian species. Along these lines, the only clearly described avian homologue of a mammalian P2Y receptor is for the P2Y1 receptor. The avian and human P2Y1 receptors are 85% identical, and these receptors exhibit essentially identical pharmacological selectivities (7). The recently reported avian p2y3 receptor is likely a species homologue of the previously cloned mammalian P2Y6 receptor; these proteins are 65% identical and exhibit essentially the same pharmacological specificities.2 The interspecies differences between avian and mammalian β-adrenergic receptors range from a >80% identity of β2-adrenergic receptors3 to the approximately 60% identity of β1-adrenergic receptors (29). A β-adrenergic receptor for which no mammalian homologue had been found previously was cloned from turkey (30).
Recently Bogdanov et al. reported the cloning and expression of a Xenopus laevis P2Y receptor (Xlp2y) (31). This receptor has unique characteristics that include a 216-amino-acid carboxyl-terminal tail and similar responsiveness to all naturally occurring nucleotide triphosphates. Activation of this receptor expressed in X. laevis oocytes results in persistent activation of a Ca2+-dependent Cl− current. The Xlp2y receptor also is closely related to the P2Y4 and P2Y2 receptors (62% and 56% identical, respectively) between transmembrane domains 1 and 7. The homology between the amphibian P2Y receptor and the avian P2Y receptor reported here is also high (57% identical over the overlapping regions). However, structural differences between both receptor proteins suggest that the avian P2Y receptor is not a species homologue of the amphibian receptor reported by Bogdanov et al. (31).
The receptor nomenclature used to date, together with an urgency to assign a definitive name to cloned G protein-coupled receptors, has created some ambiguities regarding the nature of several of the proposed members of the P2Y receptor family. For example, two cloned G protein-coupled receptors called the p2y5 and p2y7 receptors (14, 16) were proposed to be members of the P2Y receptor family, but this probably will prove not to be the case (18-20). To avoid more confusion in the nomenclature of P2Y receptors, and in conformity with the IUPHAR recommendations on receptor nomenclature (32) as adopted by the subcommittee on P2 receptors (33), we do not propose a number designation for the receptor described here.
The cloning of the novel P2Y receptor from a turkey blood cDNA library suggests that this receptor is expressed in blood cells. Over the past decade, we have studied extensively a P2Y1receptor on turkey erythrocytes that has been cloned from turkey (26), chick (6), and several mammalian species (34, 35) including human (7,36, 37). We have conducted a broad biochemical and pharmacological analysis of the P2Y1 receptor in the turkey erythrocyte model system (38, 39), and these previous studies distinguish the P2Y1 receptor from the avian P2Y receptor reported here. For example, the P2Y1receptor expressed in turkey erythrocytes is most potently activated by 2MeSATP and 2MeSADP, whereas the avian receptor reported here is only weakly activated by 2MeSATP, and 2MeSADP is ineffective. In contrast, the EC50 for UTP is 30 nm for the novel avian receptor, but UTP is inactive at the turkey erythrocyte P2Y1 receptor. Finally, AP4A is inactive at the turkey erythrocyte P2Y1 receptor but is one of the most potent agonists at the novel avian receptor. These data are consistent with the idea that the newly identified avian receptor is not expressed in erythrocytes. We are currently investigating the expression of this receptor in other avian blood cell populations and other tissues and are examining human genomic libraries to determine if a clear mammalian homologue of this receptor exists.
Acknowledgments
We are indebted to Arvind Mohanram for excellent technical assistance, to Joel Schachter, Qing Li, Eduardo Lazarowski, and Rob Nicholas for helpful discussions and suggestions, and to Brenda Asam for typing the manuscript.
Footnotes
- Received July 9, 1997.
- Accepted August 25, 1997.
-
Send reprint requests to: José L. Boyer, Department of Pharmacology, CB# 7365, Mary Ellen Jones Bldg., University of North Carolina, Chapel Hill, NC 27599. E-mail:boyerl{at}med.unc.edu
-
↵3 F. del Toro, J. L. Boyer, T. K. Harden, and R. A. Nicholas, manuscript in preparation.
-
This work was supported by United States Public Health Service Grants HL54889 and GM38213.
-
↵1 The nomenclature used here is that suggested by the International Union of Pharmacology Committee on Receptor Nomenclature and Drug Classification (32) and adapted by the Purinoceptor Subcommittee of the same organization (33).
-
↵2 Q. Li, M. Olesky, T. K. Harden, and R. A. Nicholas, manuscript in preperation.
Abbreviations
- 2MeSATP
- 2-methylthio-ATP
- 2MeSADP
- 2-methylthio-ADP
- EGTA
- ethylene glycol bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid
- DMEM
- Dulbecco’s modified Eagle’s medium
- HEPES
- 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
- PIPES
- piperazine-N,N′-bis-2-ethanesulfonic acid
- APxA
- diadenosine x-phosphate, wherex is the number of phosphates
- PCR
- polymerase chain reaction
- bp
- base pair(s)
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}