

Abstract
The present study examined the involvement of the 5-HT1A receptors in classical fear conditioning using the 5-HT1A agonist 8-hydroxy-2-(di-n-propyloamino)tetralin hydrobromide (8-OH-DPAT) and the selective “silent” 5-HT1A receptor antagonist (N-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-(2-pyridinyl)cyclo-hexane carboxamide trihydrochloride (WAY 100635). The drugs were administered both subcutaneously and bilaterally into the dorsal hippocampus of male C57BL/6J mice. The training was performed in a single trial in which a tone was followed by a footshock. The retention of context- and tone-dependent fear was examined in separate tests conducted either 1 or 24 hr after training. Subcutaneous 8-OH-DPAT (0.1–1.0 mg/kg), when injected before but not after training, caused a dose-dependent impairment of contextual fear in both 1 and 24 hr tests, whereas tone-dependent fear was less affected. Pretraining intrahippocampal injections of 5.0 μg but not 1.0 μg 8-OH-DPAT caused a severe deficit in contextual fear when tested 24 hr after training. When injected both subcutaneously and intrahippocampally, 8-OH-DPAT induced the 5-HT syndrome, indicative of postsynaptic 5-HT1A receptor activation at the dose ranges that impaired fear conditioning. However, the behavioral changes induced by 8-OH-DPAT at the time of training could not account for inhibitory effects of 8-OH-DPAT on fear conditioning. Neither subcutaneous (0.03 mg/kg) nor intrahippocampal (0.5 μg per mouse) WAY 100635 altered context- or tone-dependent fear. However, subcutaneous WAY 100635 blocked both the 5-HT syndrome and the impairment of fear conditioning induced by subcutaneous or intrahippocampal 8-OH-DPAT. In contrast, intrahippocampal WAY 100635 blocked the impairment caused by intrahippocampal but not subcutaneous 8-OH-DPAT, indicating the involvement of extrahippocampal 5-HT1A receptors in fear conditioning. It is concluded that the deficits in fear conditioning induced by 8-OH-DPAT are a result of postsynaptic 5-HT1Areceptor activation that interferes with learning processes operating at acquisition but not consolidation. Furthermore, the dorsohippocampal 5-HT1A receptors play an important but not exclusive role in the limbic circuitry subserving contextual fear conditioning.
- serotonin
- presynaptic and postsynaptic 5-HT1A receptors
- dorsal hippocampus
- fear conditioning
- 8-OH-DPAT
- WAY 100635
Accumulating evidence from animal and human studies indicates that forebrain 5-hydroxytryptamine (5-HT, serotonin) systems are involved in cognition (Ögren 1985; McEntee and Crook, 1991; Meltzer et al., 1998; Meneses, 1999). Among the 5-HT receptor subtypes shown to play a role in various learning and memory models (Meneses, 1999; Misane and Ögren, 2000), the 5-HT1A receptors are of particular interest. Receptor binding and in situ hybridization studies have shown that the postsynaptic 5-HT1A receptors are enriched in the limbic forebrain structures, such as hippocampal formation (the pyramidal cells of CA1 and CA3 areas, granular cells of dentate gyrus), lateral septum, and entorhinal cortex, which are known to play an important role in learning and memory. In addition, presynaptic 5-HT1A receptors (somatodendritic autoreceptors) are abundant in the midbrain raphe nuclei (Laporte et al., 1994; Gozlan et al., 1995; Khawaja, 1995). The representation of the 5-HT1A receptors in the limbic forebrain probably explains the marked effects of the 5-HT1A agonists such as 8-hydroxy-2-(di-n-propyloamino)tetralin hydrobromide (8-OH-DPAT) in various learning and memory tasks in rats and mice ranging from aversive conditioning (Carli et al., 1993; Misane et al., 1998; Misane and Ögren, 2000) to spatial learning (Carli and Samanin, 1992;Bertrand et al., 2000) (for review, see Meneses, 1999). Although the exact mechanisms underlying these effects are not well defined, they might be related to the inhibitory effects of the 5-HT1A agonists on limbic (Sprouse and Aghajanian, 1988; Grunschlag et al., 1997; Tada et al., 1999) and neocortical (Hajos et al., 1999) neuronal activity.
Classical fear conditioning is an associative learning paradigm for studying aversive learning and memory. In this model, a neutral conditioned stimulus (CS) such as sound or light flash is combined with an aversive unconditioned stimulus (US) such as footshock. After conditioning, the CS elicits a defensive reaction in the absence of the US (LeDoux, 1993, 1995; Fendt and Fanselow, 1999). An advantage of this model is that context- and tone-dependent fear are acquired in a single learning trial and that they can be assessed independently from each other. The single trial procedure allows also an exact timing of drug treatment in relation to training and retention.
The neuronal systems underlying context- and tone-dependent fear conditioning have been investigated in some detail (Rogan and LeDoux, 1996; for review, see Fendt and Fanselow, 1999), and both context- and tone-dependent fear conditioning depend on the integrity of the amygdala, which is considered to be the sensorimotor interface for conditioned fear (Rogan and LeDoux, 1996; Fendt and Fanselow, 1999). Although the hippocampus is a major afferent input to the amygdala, the role of the dorsal hippocampus in either context- and tone-dependent fear conditioning is still debated. Contextual fear conditioning, which involves multimodal sensory processing of continuously present (tonic) stimuli at the time of training, is assumed to depend on a “time-limited” function of the hippocampus (Selden et al., 1991;Kim and Fanselow, 1992; Kim et al., 1993; Holland and Bouton, 1999;Fanselow, 2000). In contrast, tone-dependent fear conditioning, which involves discrete (phasic) unisensory information processing, was suggested not to involve hippocampus (Kim and Fanselow, 1992; Phillips and LeDoux, 1992; Chen et al., 1996). However, more recent investigations suggest that the hippocampus may also be involved in tone-dependent fear conditioning (Maren et al., 1997; Radulovic et al., 1999).
Previous studies using nonselective 5-HTergic manipulations, such as 5-HT releasing compounds, 5-HT reuptake inhibitors, or neurotoxins, have provided evidence for a possible role of 5-HT in contextual fear conditioning in the rat (Archer, 1982; Archer et al., 1982, 1984; Inoue et al., 1996a,b; Hashimoto et al., 1997). Furthermore, combination studies of 5-HT1A antagonists and the 5-HT reuptake inhibitor citalopram have also implicated involvement of the 5-HT1A receptors in the conditioned freezing (Hashimoto et al., 1997). In mice, the role of 5-HT in fear conditioning has been studied using transgenics, that is, 5-HT1B and monoamine oxidase A knock-out mice (Kim et al., 1997; Malleret et al., 1999). To date, the role of 5-HT1A receptors in classical fear conditioning using tone-shock association has not been studied in any of the species.
This study examined the involvement of 5-HT1A receptors in both contextual (background) and tone-dependent fear conditioning in C57BL/6J mice by the use of the 5-HT1A receptor agonist 8-OH-DPAT and the selective 5-HT1A receptor antagonistN-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-N-(2-pyridinyl)cyclohexane carboxamide trihydrochloride (WAY 100635). In addition, symptoms of the 5-HT syndrome indicative of postsynaptic 5-HT1A receptor activation were assessed during training. To analyze the role of dorsohippocampal 5-HT1A receptors in fear conditioning, 8-OH-DPAT and WAY 100635 were injected subcutaneously and intrahippocampally.
MATERIALS AND METHODS
Subjects. A total of 325 male C57BL/6J mice aged 9–12 weeks (Centre D'Elevage, Le Genest St. Isle, France) were used. The mice were individually housed in standard Macrolon cages (Type 2: 22 × 16 × 13 cm) and maintained on a 12 hr light/dark cycle (lights on at 7 A.M.) with food and water freely available. All experimental procedures were in accordance with the European Council Directive (86/609/EEC) by permission of the Animal Section Law enforced by the District Government of Braunschweig, Lower Saxony, Germany.
Fear conditioning and behavioral observations. The fear conditioning experiments were performed as previously described (Stiedl and Spiess, 1997; Milanovic et al., 1998; Stiedl et al., 1999a) using a computer-controlled fear conditioning system (TSE, 303410, Bad Homburg, Germany). The scheme of tests is presented in Figure1. Fear conditioning was performed in a Plexiglas cage (36 × 21 × 20 cm) within a constantly illuminated (12 V, 10 W halogen lamp, 100–500 lux) fear conditioning box of dark gray color (context 1). In the conditioning box, a high-frequency loudspeaker (Conrad, KT-25-DT, Hirschau, Germany) provided constant background noise [white noise, 68 dB sound pressure level (SPL)].

Experimental paradigm for the fear conditioning tests with the training and memory test sequences. Mice were subjected to contextual background fear conditioning (signaled shock) during the training. US, Unconditioned stimulus (footshock, 0.7 mA, 2 sec, constant current); CS, conditioned stimulus (tone, 10 kHz, 75 dB SPL, pulsed 5 Hz); Δt = time interval. For details, see Materials and Methods.
The training (conditioning) consisted of a single trial. The mouse was exposed to the conditioning context (180 sec) followed by a tone (CS, 30 sec, 10 kHz, 75 dB SPL, pulsed 5 Hz). After termination of the tone, a footshock (US, 0.7 mA, 2 sec, constant current) was delivered through a stainless steel grid floor. The mouse was removed from the fear conditioning box 30 sec after shock termination to avoid an aversive association with the handling procedure. Only mice that received post-training injections were removed from the fear conditioning box directly after footshock exposure. Under these conditions, the context served as background stimulus. The fear conditioning chamber was thoroughly cleaned with 70% ethanol before each animal.
Before and during the exploratory phase in context 1, the behavior of the animals was assessed, and the presence or absence of the components of the serotonin (5-HT) syndrome (flat body posture, crouching, reciprocal forepaw treading, ballistic paw movements, lateral head weaving, hind-limb abduction, Straub tail, and tremor) (Tricklebank et al., 1984; Blanchard et al., 1997) were noted.
Memory tests were performed either 1 or 24 hr after fear conditioning (Stiedl et al., 1999a) (Fig. 1). Contextual memory was tested in the fear conditioning box (context 1) for 180 sec without CS or US presentation (with background noise). Subsequently, without delay in the case of the 1 hr test or with a 2 hr interval in the case of the 24 hr test, the tone-dependent memory test was performed in a novel context (context 2) (Stiedl and Spiess, 1997; Stiedl et al., 1999a). Context 2 represented an identically sized cage with a plain floor in a white-surrounding environment (350–500 lux) outside the fear conditioning box that was cleaned with 1% acetic acid before each animal. No background noise was provided. In the tone-dependent memory test, a 180 sec pause without stimulation (pre-CS phase) preceded a 180 sec period of auditory stimulation (Fig. 1, CS phase).
Freezing, defined as the lack of movement except for respiration, was assessed as the behavioral parameter of the defensive reaction of mice (Bolles and Riley, 1973; Fanselow and Bolles, 1979) by a time-sampling procedure throughout conditioning and memory tests. Every 10 sec (during 1 sec light-pulse), each mouse was instantly judged as either freezing or active by observers that were unaware of the training condition. Interobserver reliability was r = 0.97. In addition, activity-derived measures (inactivity, mean activity, and exploratory area) were recorded by a photo beam system (10 Hz detection rate) controlled by the fear conditioning system (Radulovic et al., 1998; Stiedl et al., 1999a,b). Activity (centimeters per second) consisted of locomotor activity and local body movements that were graphed as an activity plot (Fig. 2). The duration of inactivity was calculated by the fear conditioning system from the activity plots and expressed in percentage of time during which the mouse was inactive during a defined test phase (Stiedl et al., 1999a). Inactivity served as an additional measure of fear. The threshold for inactivity was defined as <1 cm/sec (Stiedl et al., 1999a). In addition, the mean activity (centimeters per second) and exploratory area (percentage of total area covered by a mouse) for each test-phase were calculated by the fear conditioning system. In the tone-dependent memory tests, changes in the test parameters from the pre-CS to the CS phase (calculated as Δ = CS − pre-CS value), served as indicators of the tone-specific response throughout this study.

Representative activity plots of two C57BL/6J mice obtained by the fear conditioning system. One mouse was injected with 8-OH-DPAT (0.5 mg/kg, s.c.) 15 min before training, and the control mouse received a subcutaneous saline injection. The threshold for inactivity (<1 cm/sec) is indicated in each plot. For details, see Materials and Methods.
Each experimental group consisted of 6–19 mice, and all experiments were performed during the light phase of a 12 hr light/dark cycle.
Stereotaxic operations. Double guide cannulae (C235, Plastics One, Roanoke, VA) were implanted in a stereotactic holder during 1.2% avertin anesthesia (0.02 ml/g, i.p.) under aseptic conditions as previously described (Radulovic et al., 1999; Stiedl et al., 2000). Each double guide cannula with inserted dummy cannula and dust cap was fixed to the skull by dental cement. The cannula was directed toward both dorsal hippocampi (Fig. 2). The coordinates were based on the stereotaxic plates of the atlas of Franklin and Paxinos (1997): anterior-posterior (AP) coordinates referred to bregma, lateral (L) coordinates to the midsagittal suture line, and ventral (V) coordinates to the surface of the skull: AP, −1.5 mm; L, ±1.0 mm, and V, −2.5 mm (injection site).
The animals were allowed to recover for 4–5 d before the experiments started. Mice that displayed signs of distress such as weight loss or apparent behavioral abnormalities were excluded from the experiment.
Drugs and drug administration. 8-OH-DPAT and WAY 100635 were obtained from RBI (Natick, MA). Both drugs were dissolved in saline (NaCl, 0.9%) and administered subcutaneously into the scruff of the neck in a volume of 8 ml/kg under brief (∼60 sec) isoflurane inhalation anesthesia (Forene, Abbott, Wiesbaden, Germany).
For intrahippocampal (i.h.) injections, 8-OH-DPAT and WAY 100635 were dissolved in sterile artificial CSF (aCSF) on the day of injection. The aCSF solution contained (in mm): NaCl 130, NaHCO3 24, MgSO4·7H2O 1.5, CaCl2·2H2O 2.0, KCl 3.5, NaH2PO4·H2O 1.25, and glucose 10, adjusted to pH 7.4 and 300 mOsm. A microinfusion pump (CMA/100, CMA/Microdialysis, Solna, Sweden) was used to infuse WAY 100635 (0.5 μg per mouse) and 8-OH-DPAT (1.0 and 5.0 μg per mouse) bilaterally into the dorsal hippocampus (0.25 μl per side) at a flow rate of 0.33 μl/min. Two 25 μl syringes were mounted onto the pump and a double injection cannula (Plastics One) was connected by polyethylene tubing to the syringes. The microinjector delivered the injection solution 1 mm below the tip of the guide cannula at the depth of 2.5 mm. Before injection, the cap and the dummy cannula were removed. Intracerebral injections were applied during ∼90 sec isoflurane inhalation anesthesia. After infusion, the dummy cannula and the cap were placed back on the guide cannula.
In most experiments, WAY 100635 and 8-OH-DPAT were administered 30 and 15 min, respectively, before training. Some experiments were performed with immediate post-training subcutaneous administration of 8-OH-DPAT.
Histology. After the behavioral experiments, the injection sites were confirmed in each mouse that received an intrahippocampal injection by bilateral injection of 0.25 μl methylene blue solution and subsequent histological evaluation. Coronal brain sections were used to verify the exact site of injection. A representative guide cannula placement and injection sites are shown in Figure3.

The injection sites in dorsal hippocampus are indicated on the right hemisphere according to the coronal section from the Mouse Brain Atlas (Franklin and Paxinos, 1997) (A). Representative photomicrographs of coronal brain section of bilateral dorsal hippocampal injection of methylene blue before and after counterstaining with nuclear fast red are shown (B). DG, Dentate gyrus;CA1, CA1 area of the hippocampus; CA3, CA3 area of the hippocampus.
Only animals that received symmetrical and bilateral injections into the dorsal hippocampi were included in the final analysis of fear conditioning data, which are presented in Results.
Among the total of 152 animals with intracerebral aCSF, 8-OH-DPAT, or WAY 100635 injections, 55 mice were excluded because of misplacement of cannula as well the failures of equipment during the fear conditioning experiments. The data from mice with misplaced cannula were also evaluated and used as nonspecific controls for the localized site injections.
Statistical analysis. The overall treatment effects were examined using one-way ANOVA. For each significant F ratio, Fisher's protected least significant difference (PLSD) test was used to analyze the statistical significance of appropriate multiple comparisons (Kirk, 1968). A probability level of p < 0.05 was accepted as statistically significant in all of the studies, and all of the post hoc tests were two-tailed.
RESULTS
Effects of subcutaneous drug administration on fear conditioning
Effects of subcutaneous pretraining 8-OH-DPAT on fear conditioning (24 hr retention)
Injection of 8-OH-DPAT (0.05–1.0 mg/kg, s.c.) 15 min before training resulted in a dose-dependent decrease in mean activity (F(4,45) = 17.17, p < 0.001) and exploratory area (F(4,45) = 26.76, p < 0.001) and increase in inactivity (F(4,45) = 17.30, p < 0.001) during the 180 sec contextual exploration phase (Table1). A significant effect for all of these measurements was seen already from the 0.1 mg/kg dose (p < 0.001, Fisher's PLSD test). Freezing, regardless of treatment, was only occasional (data not shown). All animals showed clear attentive responses to the 30 sec tone by transient suppression of activity (Fig. 2) and the reduction of mean activity, which was not different among treatment groups (F(4,45) = 0.87, p > 0.48). No startle-like responses to tone were observed. There was a dose-dependent attenuation of shock response (F(4,45) = 2.89,p < 0.05) in the 8-OH-DPAT-treated animals, with significant effect at the 0.5 and 1.0 mg/kg doses (p < 0.05 and p < 0.01 vs saline control, respectively) (Table 1, Fig. 2).
Effects of 8-OH-DPAT on training performance in 24 and 1 hr tests in C57BL/6J mice
When tested 24 hr after training, 8-OH-DPAT caused a dose-dependent impairment of contextual fear conditioning, as indicated by significant reduction of freezing (F(4,45) = 37.09, p < 0.001) and inactivity (F(4,45) = 21.38, p < 0.001) as well as increase in mean activity (F(4,45) = 18.34, p < 0.001) and exploratory area (F(4,45) = 15.52, p < 0.001) when compared with saline controls (Fig. 4). In this test, 8-OH-DPAT produced a significant effect for all parameters from the 0.5 mg/kg dose (p < 0.001 vs saline control), whereas the exploratory area was increased already at the 0.1 mg/kg dose (p < 0.05 vs saline control).

The effects of pretraining subcutaneous 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (24 hr retention). 8-OH-DPAT (0.05–1.0 mg/kg, s.c.) was administered 15 min before training. The saline control group was tested concurrently with 8-OH-DPAT-treated groups. Contextual memory test was performed 24 hr after training. Two hours later, mice were subjected to the tone-dependent memory (pre-CS and CS phases) test. Vertical bars represent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. The statistical analysis was performed by one-way ANOVA followed by Fisher's PLSD test (*p < 0.05, **p < 0.01, and ***p < 0.001 vs saline control group;n = 9–11). For details, see Materials and Methods and Figure 1.
In the subsequent tone-dependent memory test, 8-OH-DPAT affected both pre-CS and CS phases (Fig. 4). In the pre-CS phase, saline controls had low freezing scores and low level of inactivity as well as mean activity that were comparable with those found during training (Table1). In the pre-CS phase, a significant effect was found for freezing (F(4,45) = 4.62, p < 0.01) and inactivity (F(4,45) = 2.93, p < 0.05). Mice that received the lowest 0.05 mg/kg dose of 8-OH-DPAT showed higher freezing scores than the control group (p < 0.05 vs saline control).
In the CS phase, a significant treatment effect for freezing (F(4,45) = 7.20,p < 0.001), inactivity (F(4,45) = 4.44, p < 0.01), and mean activity (F(4,45) = 3.34, p < 0.05) was found. Lower freezing scores and inactivity were found in the groups treated with 8-OH-DPAT (0.5 and 1.0 mg/kg) (Fig. 4).
ANOVA also revealed a tone-specific change (Δ = CS-pre-CS) among treatment groups for freezing (F(4,45)=4.19, p < 0.01) but not for inactivity (F(4,45) =1.20, p > 0.32). Post hoc comparisons indicated that these tone-induced changes were decreased at the 0.5 mg/kg but not at the 1.0 mg/kg dose of 8-OH-DPAT (data not shown).
On the basis of these results, the 0.5 mg/kg dose of 8-OH-DPAT was chosen for combination studies with the selective 5-HT1A receptor antagonist WAY 100635.
Effects of subcutaneous pretraining 8-OH-DPAT on fear conditioning (1 hr retention)
In this experiment, effects of pretraining 8-OH-DPAT (0.5 and 1.0 mg/kg, s.c.) on training performance were similar to those found in the experiment with 24 hr retention (Table 1).
When examined 1 hr after training, 8-OH-DPAT (0.5 and 1.0 mg/kg) caused an impairment of contextual fear conditioning (Fig.5). A significant treatment effect was found for freezing (F(2,20) = 16.68,p < 0.001), inactivity (F(2,20) = 7.87, p < 0.01), mean activity (F(2,20) = 8.60,p < 0.01), and exploratory area (F(2,20) = 10.23, p < 0.001).

The effects of pretraining subcutaneous 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (1 hr retention). 8-OH-DPAT (0.5–1.0 mg/kg, s.c.) was administered 15 min before training. The saline control group was tested concurrently with 8-OH-DPAT-treated groups. Contextual memory test was performed 1 hr after training. Mice were subjected to the tone-dependent memory test (pre-CS and CS phases) directly after contextual memory test.Vertical bars represent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. *p< 0.05, **p < 0.01, and ***p< 0.001 versus saline control group, n = 7–8. For details of statistical analysis and general information, see Figures 1and 4.
In the tone-dependent memory test, overall differences in fear responses were not found in either the pre-CS or CS phase (Fig. 5). However, ANOVA revealed differences in tone-specific changes (Δ = CS–pre-CS) for inactivity (F(2,20)= 7.26, p < 0.01), mean activity (F(2,20) = 4.92, p < 0.05), and exploratory area (F(2,20) = 8.87, p < 0.01), and a trend for changes in freezing (F(2,20) = 3.27, p = 0.058). No clear dose dependency was found because the increases in fear responses in CS phase in the animals treated with the 0.5 mg/kg dose of 8-OH-DPAT were lower than those in the control group, but no significant differences were found at the 1.0 mg/kg dose (data not shown).
Effects of subcutaneous pretraining 8-OH-DPAT and WAY 100635 on fear conditioning (24 hr retention)
This experiment was designed to show the crucial role of the 5-HT1A receptors in the impairment of context- and tone-dependent fear by 8-OH-DPAT. For this reason, the selective and “silent” 5-HT1A receptor antagonist WAY 100635 was injected in combination with 8-OH-DPAT (0.5 mg/kg, s.c.). The doses of WAY 100635 (0.03–0.3 mg/kg, s.c.) were chosen on the basis of previous studies in the rat (Misane and Ögren, 2000), and they were in the dose range known to block 5-HT1A receptors in vivo (Fletcher et al., 1996).
In this experiment, ANOVA revealed a significant overall treatment effect on the contextual exploratory phase during training: mean activity (F(4,38) = 35.17,p < 0.001), exploratory area (F(4,38) = 61.49,p < 0.001), and inactivity (F(4,38) = 38.99, p < 0.001). Similar to the dose–response studies, 8-OH-DPAT (0.5 mg/kg) when administered 15 min before training caused a marked decrease in activity in the 180 sec contextual exploratory phase (all test parameters, p < 0.001 vs saline + saline control). WAY 100635 (0.03–0.3 mg/kg, s.c.) by itself did not alter any of these parameters. However, the 0.03 mg/kg dose of WAY 100635 completely normalized activity in the 8-OH-DPAT-treated mice (data not shown).
Freezing, regardless of treatment, was observed only occasionally. All animals showed clear attentive responses to the 30 sec tone by a transient suppression of activity and the reduction of mean activity, which did not differ among treatment groups (F(4,38) = 0.87, p > 0.48) (data not shown). No startle-like responses to tone were observed.
ANOVA revealed a significant treatment effect for mean activity during shock exposure (F(4,38) = 4.84,p < 0.01). WAY 100635 (0.03–0.3 mg/kg) by itself did not alter shock response. However, WAY 100635 (0.03 mg/kg) completely normalized the attenuated shock response in the 8-OH-DPAT-treated animals (p < 0.05 vs saline + 8-OH-DPAT-treated group and p > 0.81 vs saline + saline control group) (data not shown).
In the contextual memory test (Fig. 6), ANOVA indicated a significant main treatment effect for freezing (F(4,38) = 32.71, p < 0.001), inactivity (F4,38 = 14.67,p < 0.001), mean activity (F(4,38) = 13.31, p < 0.01), and exploratory area (F(4,38) = 11.68, p < 0.001). WAY 100635 (0.03–0.3 mg/kg) by itself did not influence contextual memory. However, WAY 100635 (0.03 mg/kg) completely blocked the impairment caused by 8-OH-DPAT as indicated by all test parameters (freezing, inactivity, mean activity, and exploratory area).

The combined effects of subcutaneous WAY 100635 and 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (24 hr retention). Mice were injected with WAY 100635 (0.03–0.3 mg/kg, s.c.) and/or 8-OH-DPAT (0.5 mg/kg, s.c.) 30 and 15 min before training, respectively. The saline + saline (8 ml/kg, s.c.) control group was tested concurrently with WAY 100635- and 8-OH-DPAT-treated groups. Contextual memory test was performed 24 hr after training. Mice were subjected to the tone-dependent memory test (pre-CS and CS phases) 2 hr after contextual memory test. Vertical bars represent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. *p < 0.05, **p < 0.01, and ***p < 0.001 versus saline + saline control group;†p < 0.05,†† p < 0.01, and†††p < 0.001 versus saline + 8-OH-DPAT-treated group, n = 8–10. For details of statistical analysis and general information, see Figures 1 and4.
In the pre-CS phase of the tone-dependent memory test (Fig. 6), a significant treatment effect was found for freezing (F(4,38) = 9.29, p < 0.001) and exploratory area (F(4,38) = 5.12, p < 0.01) but not for inactivity (F(4,38) = 1.78, p > 0.15) and mean activity (F(4,38) = 1.71, p > 0.16). 8-OH-DPAT caused a decrease in freezing (p < 0.001) and an increase in mean activity and exploratory area (p < 0.05) when compared with the saline + saline control group, and these effects were partly (mean activity and exploratory area) blocked by the 0.03 mg/kg dose of WAY 100635. At the 0.3 mg/kg dose, WAY 100635 caused a decrease in freezing and an increase in exploratory area.
In the CS phase, all parameters, that is, freezing (F(4,38) = 22.99, p < 0.001), inactivity (F(4,38) = 8.24,p < 0.001), mean activity (F(4,38) = 12.36, p < 0.001), and exploratory area (F(4,38)= 6.57, p < 0.001), differed among treatment groups. WAY 100635 (0.03 mg/kg, s.c.), which by itself did not alter any of the test parameters, completely or partly blocked the impairment of the conditioned fear responses caused by the 0.5 mg/kg dose of 8-OH-DPAT. At the 0.3 mg/kg dose, WAY 100635 caused a slight decrease in inactivity and an increase in mean activity and exploratory area (Fig.6).
ANOVA revealed differences in tone-specific changes (Δ = CS–pre-CS) for freezing (F(4,38) = 8.50, p < 0.001). A similar trend was found for inactivity (F(4,38) = 2.54,p = 0.055) but not for mean activity (F(4,38) = 1.37, p > 0.26) and exploratory area (F(4,38) = 1.72, p > 0.16). WAY 100635 (0.03–0.3 mg/kg) itself did not influence these tone-induced changes, but at the 0.03 mg/kg dose it completely or partly antagonized the decrease in Δ values of freezing (p > 0.99 vs saline + saline control and p < 0.001 vs saline + 8-OH-DPAT group), inactivity (p > 0.55 vs saline + saline control and p < 0.05 vs saline + 8-OH-DPAT group), mean activity (p > 0.77 vs saline + saline control and p > 0.14 vs saline + 8-OH-DPAT group), and exploratory area (p > 0.39 vs saline + saline control and p > 0.20 vs saline + 8-OH-DPAT group) caused by 8-OH-DPAT (data not shown).
Effects of subcutaneous post-training 8-OH-DPAT on fear conditioning (24 hr retention)
In this experiment, all animals were drug naive at the time of training. ANOVA revealed no differences between subsequent saline or 8-OH-DPAT-treatment (0.5–1.0 mg/kg, s.c.) groups for inactivity (F(2,21) = 1.16, p > 0.33), mean activity (F(2,21) = 0.45,p > 0.64), or exploratory area (F(2,21) = 0.41, p > 0.67) (data not shown). No differences were found for either tone or shock responses (data not shown).
Injection of 8-OH-DPAT immediately after training did not affect either context- or tone-dependent memory: context freezing (F(2,21) = 0.14, p > 0.87); context inactivity (F(2,21) = 0.90, p > 0.42); context mean activity (F(2,21) = 0.98, p > 0.39); context exploratory area (F(2,21) = 0.36, p > 0.70); pre-CS freezing (F(2,21)= 2.30, p > 0.12); pre-CS inactivity (F(2,21) = 1.08, p > 0.35); pre-CS mean activity (F2,21 = 1.74,p > 0.19); pre-CS exploratory area (F(2,21) = 0.96, p > 0.39); CS freezing (F(2,21) = 0.24,p > 0.78); CS inactivity (F(2,21) = 0.80, p > 0.46); CS mean activity (F(2,21) = 1.07, p > 0.35); CS exploratory area (F(2,21) = 0.18, p > 0.83) (Tables 2,3).
Effects of post-training 8-OH-DPAT in 1 and 24 hr contextual memory tests in C57BL/6J mice
Effects of post-training 8-OH-DPAT in 1 and 24 hr tone-dependent memory tests in C57BL/6J mice
Effects of subcutaneous post-training 8-OH-DPAT on fear conditioning (1 hr retention)
Similar to the 24 hr test, no differences among treatment groups were found in the training phase of the 1 hr test when 8-OH-DPAT (0.5–1.0 mg/kg, s.c.) was injected immediately after training (data not shown).
In this experiment, 8-OH-DPAT did not affect the contextual memory, as indicated by freezing (F(2,16) = 0.73,p > 0.49), inactivity (F(2,16) = 0.72, p > 0.50), mean activity (F(2,16) = 0.57,p > 0.57), and exploratory area (F(2,16) = 0.25, p > 0.77) (Table 2).
In contrast, differences were revealed in the tone-dependent memory test (Table 3). In the pre-CS phase, a significant treatment effect was found for inactivity (F(2,16) = 4.22,p < 0.05), mean activity (F(2,16) = 7.78, p < 0.01), and exploratory area (F(2,16) = 6.21, p = 0.01) but not for freezing (F(2,16) = 2.26, p > 0.13). Post hoc comparisons revealed higher inactivity (p < 0.05 vs saline control), lower mean activity (p < 0.01 vs saline control), and decreased exploratory area (p < 0.01 vs saline control) at the 1.0 mg/kg dose of 8-OH-DPAT. Similar results were obtained in the CS phase: inactivity (F(2,16) = 6.71, p < 0.01), mean activity (F(2,16)= 8.21, p < 0.01), exploratory area (F(2,16) = 5.51, p < 0.05). In addition, it was a significant treatment effect also for freezing (F(2,16) = 4.04,p < 0.05). Higher freezing scores (p < 0.05 vs saline control), higher inactivity (p < 0.01 vs saline control), lower mean activity (p < 0.001 vs saline control), and decreased exploratory area (p < 0.01 vs saline control) were found at the 1.0 mg/kg dose of 8-OH-DPAT.
There was no overall treatment effect, however, for the values of the change (from pre-CS to CS phase) in freezing (F(2,16) = 3.03, p > 0.07), inactivity (F(2,16) = 2.70,p > 0.09), mean activity (F(2,16) = 0.76,p > 0.48), or exploratory area (F(2,16) = 0.13, p > 0.87) (Table 3). Despite this, freezing and inactivity increased more at the 1.0 mg/kg dose of 8-OH-DPAT (p < 0.05 vs saline control).
Effects of intrahippocampal drug administration on fear conditioning
Effects of intrahippocampal 8-OH-DPAT on fear conditioning (24 hr retention)
This experiment was designed to show the involvement of hippocampal 5-HT1A receptors in context- and tone-dependent fear conditioning. The doses for intrahippocampal 8-OH-DPAT were chosen on the basis of data obtained in dose–response studies with subcutaneous 8-OH-DPAT.
Bilateral injections of 8-OH-DPAT (1.0–5.0 μg per mouse) into dorsal hippocampus resulted in a dose-dependent decrease in mean activity (F(2,29) = 10.53, p < 0.001) and exploratory area (F(2,29) = 14.30, p < 0.001) and an increase in inactivity (F(2,29) = 7.76, p < 0.01) during the 180 sec exploration period (Table4). A significant effect for all these measurements was found at the 5.0 μg (p < 0.001, Fisher's PLSD test) but not at the 1.0 μg dose. Freezing, independently of treatment, was only occasional (data not shown). A transient reduction of activity caused by the tone presentation did not differ among treatment groups (F(2,29)= 1.26, p > 0.29). No startle-like responses to tone were observed. A dose-dependent attenuation of shock response (F(4,45) = 3.51, p < 0.05) was observed in the 8-OH-DPAT-treated animals with significant effect at the 5.0 μg dose (p < 0.05 vs aCSF control) (Table 4).
Effects of intrahippocampal 8-OH-DPAT on training performance in C57BL/6J mice
When tested 24 hr after training, intrahippocampal 8-OH-DPAT caused a dose-dependent impairment of context-dependent memory as indicated by a significant reduction of freezing (F(2,29) = 34.20,p < 0.001) and inactivity (F(2,29) = 13.40, p < 0.001) as well as an increase in mean activity (F(2,29) = 20.00, p < 0.001) and exploratory area (F(2,29) = 14.05, p < 0.001) when compared with aCSF controls (Fig. 7). 8-OH-DPAT affected all of these parameters at the 5.0 μg (p < 0.001 vs aCSF control) but not at the 1.0 μg dose.

The effects of pretraining intrahippocampal 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (24 hr retention). 8-OH-DPAT (1.0 or 5.0 μg per mouse) was administered via chronic cannulae bilaterally into the dorsal hippocampus (i.h.) 15 min before training. The aCSF (intrahippocampal, bilaterally, 0.25 μl per side) control group was tested concurrently with 8-OH-DPAT-treated groups. Contextual memory test was performed 24 hr after training. Tone-dependent memory test (pre-CS and CS phases) was performed 2 hr later. Vertical bars represent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. *p < 0.05, **p < 0.01, and ***p < 0.001 versus aCSF control group,n = 8–13. For details of statistical analysis and general information, see Figures 1 and 4.
In the subsequent tone-dependent memory test, 8-OH-DPAT affected both pre-CS and CS phases (Fig. 7). In the pre-CS phase, a significant treatment effect was found for inactivity (F(2,29) = 2.93, p < 0.05), mean activity (F(2,29) = 2.93,p < 0.05), and exploratory area (F(2,29) = 2.93, p < 0.05). An attenuation of generalized fear was found at the 5.0 μg dose of 8-OH-DPAT.
In the CS phase, ANOVA revealed a significant treatment effect for freezing (F(2,29) = 7.20,p < 0.001), inactivity (F(2,29) = 4.44, p < 0.01), and mean activity (F(2,29) = 3.34, p < 0.05). Lower freezing scores and inactivity were found in the group treated with the 5.0 μg dose of 8-OH-DPAT (Fig. 7). However, ANOVA revealed a tone-specific change (Δ = CS–pre-CS) among treatment groups only for freezing (F(2,29) = 4.19, p < 0.01), and this parameter was decreased by the 5.0 μg dose of 8-OH-DPAT (data not shown).
On the basis of these results, the 5.0 μg dose of 8-OH-DPAT was chosen for combination studies with WAY 100635.
Effects of subcutaneous and intrahippocampal WAY 100635 and intrahippocampal 8-OH-DPAT on fear conditioning (24 hr retention)
This experiment was designed to further analyze the involvement of hippocampal 5-HT1A receptors in the effects of 8-OH-DPAT on context- and tone-dependent fear. The 0.5 μg dose for intrahippocampal injection of WAY 100635 was estimated on the basis of data obtained with subcutaneous drug administration.
In this experiment, ANOVA revealed a significant overall treatment effect on the contextual exploratory phase, that is, mean activity (F(4,47) = 9.22, p < 0.001), exploratory area (F(4,47) = 13.77, p < 0.001), and inactivity (F(4,47) = 7.74, p < 0.001). Intrahippocampal WAY 100635 (0.5 μg/mouse), which by itself did not alter any of these exploratory parameters, completely normalized a decrease in mean activity and an increase in inactivity caused by the 5.0 μg dose of 8-OH-DPAT (data not shown). When combined with intrahippocampal 8-OH-DPAT, systemic WAY 100635 (0.03 mg/kg, s.c.) not only restored all of the exploratory parameters to the control level but rather produced an increase in activity (inactivity, mean activity, and exploratory area; p < 0.001,p < 0.001, and p < 0.01 vs aCSF + aCSF control, respectively) (data not shown). Freezing, regardless of treatment, occurred only occasionally. The tone-induced reduction of mean activity did not differ among treatment groups (F(4,47) = 1.34, p > 0.26) (data not shown). No startle-like responses to tone were observed. There was no significant overall treatment effect for mean activity during shock exposure (F(4,47) = 2.24, p > 0.07). However, shock response was attenuated in the aCSF + 8-OH-DPAT-treated animals (p < 0.01 vs aCSF + aCSF control). This effect was normalized by intrahippocampal (0.5 μg/mouse) but not systemic (0.03 mg/kg, s.c.) WAY 100635, which by itself did not alter shock activity (data not shown).
In the contextual memory test (Fig.8), ANOVA indicated a significant main treatment effect for freezing (F(4,47)= 23.98, p < 0.001), inactivity (F(4,47) = 16.77, p < 0.001), mean activity (F(4,47)= 13.40, p < 0.001), and exploratory area (F(4,47) = 9.51, p < 0.001). Intrahippocampal WAY 100635 (0.5 μg per mouse) by itself did not alter conditioned contextual fear. However, it completely blocked the impairment of conditioned fear responses caused by intrahippocampal 8-OH-DPAT. Similar results were obtained with the subcutaneous 0.03 mg/kg dose of WAY 100635.

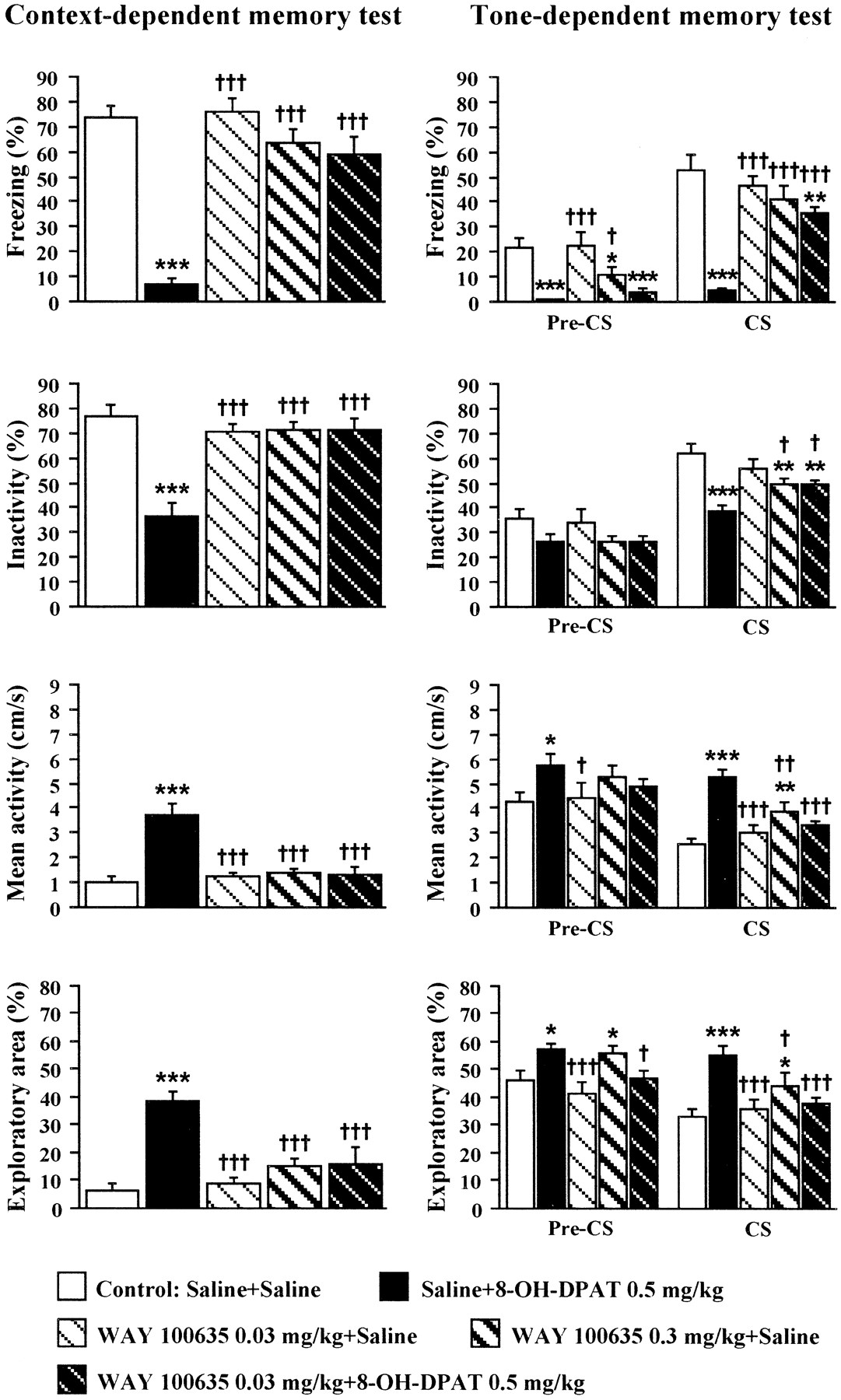
The combined effects of subcutaneous or intrahippocampal WAY 100635 and intrahippocampal 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (24 hr retention). Mice were injected with WAY 100635 (0.03 mg/kg, s.c., or 0.5 μg per mouse, i.h., bilaterally) and/or 8-OH-DPAT (5.0 μg per mouse, i.h., bilaterally) 30 and 15 min before training, respectively. The aCSF + aCSF (i.h., bilaterally, 0.25 μl per side) control group was tested concurrently with WAY 100635- and/or 8-OH-DPAT-treated groups. Contextual memory test was performed 24 hr after training. Mice were subjected to the tone-dependent memory test (pre-CS and CS phases) 2 hr after contextual memory test. Vertical bars represent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. *p < 0.05, **p < 0.01, and ***p < 0.001 versus aCSF + aCSF control group;†p < 0.05,††p < 0.01, and†††p < 0.001 versus aCSF + 8-OH-DPAT-treated group, n = 10–11. For details of statistical analysis and general information, see Figures 1 and4.
In the pre-CS phase of the tone-dependent memory test (Fig. 8), controls with double aCSF intrahippocampal injections showed a comparatively high level of generalized fear. There was a significant treatment effect for freezing (F(4,47)= 2.87, p < 0.05), inactivity (F(4,47) = 2.95, p < 0.05), and mean activity (F(4,47) = 2.90, p < 0.05), but not for exploratory area (F(4,47) = 1.99, p > 0.11). Intrahippocampal WAY 100635 (0.5 μg per mouse) reduced pre-CS freezing. However, both subcutaneous and intrahippocampal WAY 100635 blocked or tended to block the attenuation of fear responses caused by the 5.0 μg dose of 8-OH-DPAT (inactivity, mean activity, and exploratory area).
In the CS phase (Fig. 8), only freezing (F(4,47) = 4.14, p < 0.01) but not inactivity (F(4,47) = 2.08, p > 0.09), mean activity (F(4,47) = 2.44, p > 0.06), or exploratory area (F(4,47) = 1.88, p > 0.12) differed among treatment groups. However, a decrease in freezing (p < 0.001 vs aCSF + aCSF control) and inactivity (p < 0.05 vs aCSF + aCSF control) and an increase in mean activity (p < 0.05 vs aCSF + aCSF control) and exploratory area (p < 0.05 vs aCSF + aCSF control) were seen in the 8-OH-DPAT-treated mice. The 8-OH-DPAT effects were fully reversed by both systemic (0.03 mg/kg, s.c.) and intrahippocampal (0.5 μg per mouse) WAY 100635.
Neither ANOVA nor Fisher's PLSD test revealed any differences in Δ values between pre-CS and CS phases for any of the test parameters (data not shown).
Effects of intrahippocampal WAY 100635 and subcutaneous 8-OH-DPAT on fear conditioning (24 hr retention)
This experiment was performed to analyze the possible involvement of extrahippocampal 5-HT1A receptors in the context- and tone-dependent fear conditioning.
ANOVA revealed a significant overall treatment effect on the 180 sec contextual exploratory phase, that is, mean activity (F(2,34) = 45.56,p < 0.001), exploratory area (F(2,34) = 53.03, p < 0.001), and inactivity (F(2,34)= 25.84, p < 0.001). Intrahippocampal WAY 100635 (0.5 μg per mouse) failed to block the decrease in exploratory activity caused by the subcutaneous 8-OH-DPAT (0.5 mg/kg) (data not shown).
Freezing, independently of treatment, was only occasional. The reduction of mean activity during tone exposure did not differ among treatment groups (F(2,34) = 1.20,p > 0.23) (data not shown). No startle-like responses to tone were observed. There was a significant overall treatment effect for mean activity during shock exposure (F(2,34) = 5.69, p < 0.01). An attenuated shock response was found in the aCSF + 8-OH-DPAT-treated animals (p < 0.01 vs aCSF + saline control), and this was normalized by intrahippocampal WAY 100635 (data not shown).
In the contextual memory test (Fig. 9), ANOVA indicated a significant main treatment effect for freezing (F(2,34) = 117.65, p< 0.001), inactivity (F(2,34) = 29.75, p < 0.001), mean activity (F(2,34) = 16.29,p < 0.001), and exploratory area (F(2,34) = 30.97, p < 0.001). Intrahippocampal WAY 100635 (0.5 μg per mouse) failed to block the impairment of contextual fear conditioning caused by 8-OH-DPAT.

The combined effects of intrahippocampal WAY 100635 and subcutaneous 8-OH-DPAT on context- and tone-dependent memory in C57BL/6J mice (24 hr retention). Mice were injected with WAY 100635 (0.5 μg per mouse, i.h., bilaterally) and/or 8-OH-DPAT (0.5 mg/kg, s.c.) 30 and 15 min before training, respectively. The aCSF (i.h., bilaterally, 0.25 μl per side) + saline (8 ml/kg, s.c.) control group was tested concurrently with WAY 100635- and/or 8-OH-DPAT-treated groups. Contextual memory test was performed 24 hr after training. Mice were subjected to the tone-dependent memory test (pre-CS and CS phases) 2 hr after contextual memory test. Vertical barsrepresent means (±SEM) of freezing, inactivity, mean activity, and exploratory area. *p < 0.05 and ***p < 0.001 versus aCSF + saline control,n = 8–19. For details of statistical analysis and general information, see Figures 1 and 4.
In the pre-CS phase of the tone-dependent memory test (Fig. 9), there was a significant treatment effect for freezing (F(2,34) = 27.17, p < 0.001) and for exploratory area (F(2,34) = 3.74, p < 0.05).
In the CS phase (Fig. 9), freezing (F(2,34) = 60.74, p < 0.001), inactivity (F(2,34) = 19.19,p < 0.001), mean activity (F(2,34) = 11.33, p < 0.001), and exploratory area (F(2,34)= 10.95, p < 0.001) differed among treatment groups. Intrahippocampal WAY 100635 did not modulate the inhibitory effects of subcutaneous 8-OH-DPAT on fear responses in either pre-CS or CS phases.
ANOVA revealed differences in Δ values of tone-induced specific changes (CS–pre-CS) for all test parameters, that is, freezing (F(2,34) = 30.23, p < 0.001), inactivity (F(2,34) = 6.22,p < 0.01), mean activity (F(2,34) = 5.15, p < 0.05), and exploratory area (F(2,34) = 6.57, p < 0.01). The decrease in these changes found in the 8-OH-DPAT-treated mice was not antagonized by intrahippocampal WAY 100635 (data not shown).
Observations of the 5-HT syndrome
The behavioral observations in the home cages showed that 8-OH-DPAT (0.05–1.0 mg/kg, s.c.) induced a dose-dependent development of the 5-HT syndrome with a very rapid onset (2–3 min after injection). The severity of the 5-HT syndrome closely correlated with a progressive decrease in locomotor activity (locomotion and rearing). In support of this, the 5-HT syndrome started with unstable, ataxic gate and dyscoordination. Hind limb abduction, flattening of back, and eye narrowing were noted already at the 0.1 mg/kg dose. At the 0.5–1.0 mg/kg doses, the mice mostly laid in the corners of their cage; Straub tail was an additional sign of the 5-HT syndrome. Locomotion and rearing were almost or completely abolished at the 0.1–1.0 mg/kg doses of 8-OH-DPAT.
When 8-OH-DPAT was administered intrahippocampally, a clear 5-HT syndrome was seen at the 5 μg dose but not the 1 μg dose, and its symptomology and strength were comparable with that seen at the doses of 0.5–1.0 mg/kg administered subcutaneously.
The 5-HT syndrome was observed continuously for 15 min after the 8-OH-DPAT injections when mice were transferred to the fear conditioning box. As noted in the home cages, the dramatic decrease in locomotor activity accompanied the 5-HT syndrome, and only small local movements were seen in the animals treated with 0.5–1.0 mg/kg subcutaneously and the 5 μg intrahippocampal doses of 8-OH-DPAT (Table 1, Fig. 3). However, in this novel situation the pattern of the 5-HT syndrome differed: crouching prevailed but back flattening occurred; lateral head weaving and occasional forepaw treading, ballistic moves, and grooming were observed. The rest of symptoms were noted already in the home cages, that is, eye narrowing, Straub tail, and hindlimb abduction remained. This symptomology was seen at the 0.5–1.0 mg/kg (s.c.) and 5.0 μg (i.h.) doses, and some of the signs were noted at the 0.1 mg/kg (s.c.) dose of 8-OH-DPAT.
Signs of the 5-HT syndrome (head weaving) were also noted in the 1 hr memory test after the immediate post-training administration of 8-OH-DPAT (0.5 and 1.0 mg/kg). In the 1 hr test, when 8-OH-DPAT was injected 15 min before training, occasional symptoms of the 5-HT syndrome were visible in animals treated with 1.0 mg/kg but not 0.5 mg/kg dose of 8-OH-DPAT.
WAY 100635 when given subcutaneously (0.03 mg/kg) but not intrahippocampally (0.5 μg per mouse) completely abolished all symptoms of the 5-HT syndrome induced by 8-OH-DPAT (0.5 mg/kg, s.c.). However, the 5-HT syndrome induced by the intrahippocampal 8-OH-DPAT (5.0 μg per mouse) was blocked by both subcutaneous and intrahippocampal WAY 100635.
DISCUSSION
The effects of 8-OH-DPAT on context- and tone-dependent fear conditioning
The major finding of this study is that the selective 5-HT1A receptor agonist 8-OH-DPAT, when injected subcutaneously or intrahippocampally before training, caused a marked impairment of contextual fear conditioning tested 1 and 24 hr after training, indicative of deficits in information processing from short- to long-term memory. In addition, pretraining subcutaneous 8-OH-DPAT impaired a tone-dependent fear conditioning tested both 1 and 24 hr after training. However, the lack of dose-dependent effect by subcutaneous 8-OH-DPAT in the tone-dependent memory test requires further examination. There was also a clear trend for an attenuation of conditioned tone-dependent fear after intrahippocampal 8-OH-DPAT. However, because no significant differences in the tone-induced specific changes were found because of the high variability in the performance of mice, the specific involvement of dorsohippocampal 5-HT1A receptors in tone-dependent fear awaits further evaluation.
Unlike pretraining 8-OH-DPAT, the post-training activation of 5-HT1A receptors by the subcutaneous 8-OH-DPAT did not affect either short- or long-term retention of contextual fear. The facilitatory effects of the high 1.0 mg/kg dose of 8-OH-DPAT on consolidation of tone-dependent fear needs further evaluation, but it could be caused by nonspecific drug effects present at the time of testing because it was observed only in the 1 hr and not in the 24 hr test. The lack of generalized fear after pretraining 8-OH-DPAT administration suggests that an association between context 1 and shock was not formed. Therefore, it seems likely that subcutaneous injection of 8-OH-DPAT before training inhibits processing of CSs and their association with the US.
Taken together, the present results show that activation of the brain 5-HT1A receptors at the time of training disrupts acquisition of contextual and, to some extent, tone-dependent fear. In contrast, post-training 5-HT1A receptor activation may cause time-limited alterations in the consolidation of tone-dependent fear.
The role of nonspecific factors in fear conditioning
There are several factors that have to be taken into account in the interpretation of the present data. First, the drug states at both training and retention are known to influence subsequent performance in aversive learning tasks (Overton, 1978). However, data with pre- and post-training systemic 8-OH-DPAT administration indicated that there was no carry-over drug effect in the 24 hr tests, and even in the 1 hr tests nonspecific drug effects could be dissociated from specific memory-related effects (see below).
Second, the 5-HT1A agonists can alter sensorimotor reactivity at the time of conditioning (Rigdon and Weatherspoon, 1992; Sipes and Geyer, 1995; Dulawa et al., 1997), which could influence memory tests by alteration of multi- or unisensory input at the initial stage of information processing. In addition, 5-HT1A agonists induce behavioral effects such as changes in general locomotor activity (Curzon, 1990; Evenden and Angeby-Möller, 1990; Chojnacka-Wojcik, 1992; Blanchard et al., 1997) and nociceptive thresholds (Berge et al., 1985; Hamon et al., 1990) as well as induction of the 5-HT syndrome (Tricklebank et al., 1984; Blanchard et al., 1997).
These “nonspecific” effects depend on the mouse strain and the dose levels. In this study, the subcutaneous 8-OH-DPAT induced the well defined 5-HT syndrome in individually housed C57BL/6J mice already at the 0.1 mg/kg dose. This dose is approximately 5–10 times lower than those described previously (Yamada et al., 1988; Blanchard et al., 1997). This might be because of several factors such as differences in mice strains, housing, observation conditions, and finally, the route of 8-OH-DPAT administration.
Although 8-OH-DPAT caused a dose-dependent attenuation of the shock activity, all mice, regardless of treatment, reacted to the shock by flinching, running, jumping, and sometimes vocalizing. The shock response could always be confirmed by a severalfold increase in activity that was also clearly seen on the activity plots provided by the fear conditioning system (Fig. 2). The shock level used in the present study was well above the threshold for a flinch response (withdrawal of forepaws from the grid floor), which is in the range of 0.3–0.4 mA in the naive C57BL/6J mice. Furthermore, the 0.1 mg/kg dose of 8-OH-DPAT, which did not reduce shock activity, produced a deficit of contextual memory. In addition, the weaker shock activity at the higher doses of 8-OH-DPAT could be a result of the decrease in motor activity caused by the 5-HT syndrome (Evenden and Angeby-Möller, 1990). Importantly, WAY 100635, which by itself did not alter the shock response, completely normalized the shock activity in the 8-OH-DPAT-treated animals and also antagonized the 5-HT syndrome. Moreover, a clear attentive response to tone (reduction of activity) was seen in all animals regardless of treatment. Therefore, alterations in US (shock) or CS (tone) perception cannot simply account for the deficits in subsequent memory tests caused by pretraining subcutaneous or intrahippocampal 8-OH-DPAT. It is therefore hypothesized that pretraining 8-OH-DPAT produces CS–US (context–tone–shock) dissociations at the early stages of US and CS processing.
Although residual signs of the 5-HT syndrome were seen in 8-OH-DPAT-treated animals in the 1 hr tests with both pre- and post-training administration, the lack of drug effects in the contextual memory test, when 8-OH-DPAT was injected after training, cannot be explained simply by a decrease in activity. Thus, the 8-OH-DPAT-treated mice, when transferred to context 2 for the tone-dependent memory test, showed a clear increase in activity and exploratory area. In the tone-dependent tests, the nonspecific drug effects could be dissociated by the tone-induced changes in the test parameters.
In conclusion, the behavioral changes at the time of training could not account for the inhibitory effects of 8-OH-DPAT on fear conditioning.
8-OH-DPAT and 5-HT1A receptors in fear conditioning
In addition to 5-HT1A receptors, 8-OH-DPAT also has a relatively high affinity to the cloned rodent and human 5-HT7 receptors (Ruat et al., 1993; Boess and Martin, 1994; Eglen et al., 1997). However, this receptor subtype seems to be of little importance for the present results because WAY 100635, which completely antagonized the impairment of both context- and tone-dependent fear induced by the subcutaneous 8-OH-DPAT, does not bind to the 5-HT7 receptor (Hoyer et al., 1994;Routledge, 1996). On the other hand, an inverted U-shape activity by subcutaneous 8-OH-DPAT in 1 and 24 hr tone-dependent tests suggests that nonserotonergic mechanisms might contribute to the effects of 8-OH-DPAT at higher doses. For instance, (±)-8-OH-DPAT has been shown to increase basal dopamine turnover and release in the rat medial prefrontal cortex (Rasmusson et al., 1994; Tanda et al., 1994) at the doses used in the present study.
Because 5-HT1A receptors are localized both pre- and postsynaptically, the understanding of their relative role in the actions of 8-OH-DPAT is of particular importance for the analysis of mechanisms underlying aversive conditioning. The present data indicate that the deficits in fear conditioning after both intrahippocampal and subcutaneous 8-OH-DPAT administration are caused by postsynaptic 5-HT1A receptor activation, because in the dorsal hippocampus only postsynaptic 5-HT1A receptors are localized (Hall et al., 1985; Miquel et al., 1992; Kia et al., 1996). This finding is consistent with studies in another aversive learning paradigm, that is, passive avoidance (Carli et al., 1993;Mendelson et al., 1993; Misane et al., 1998). Both subcutaneous and intrahippocampal 8-OH-DPAT induced the 5-HT syndrome indicative of the postsynaptic receptor activation at the dose range that impaired fear conditioning. In addition, subcutaneous WAY 100635 (0.03 mg/kg) fully blocked the 5-HT syndrome and the deficits in fear conditioning induced by 8-OH-DPAT. Although at the 0.03 mg/kg dose, which was used in the combination studies with 8-OH-DPAT, WAY 100635 influenced neither context- nor tone-dependent fear; a slight attenuation of tone-dependent fear was found at a higher dose (0.3 mg/kg) of WAY 100635. The lack of effect of this dose on tone-induced specific changes awaits further evaluation.
Specific role of dorsohippocampal 5-HT1A receptors in fear conditioning
Although dorsohippocampal 5-HT1A receptors are involved in the regulation of motor functions and the 5-HT syndrome, their role in fear conditioning can be dissociated from their other behavioral effects. Behavioral observations in those mice with misplaced cannulae (data not shown) as well as the failure of intrahippocampal WAY 100635 to block the 5-HT syndrome caused by subcutaneous 8-OH-DPAT showed that the 5-HT syndrome is not exclusively mediated by the dorsohippocampal 5-HT1Areceptors, but it also involves postsynaptic 5-HT1A receptors in other hippocampal subregions as well as nonhippocampal forebrain structures. Furthermore, animals with unilateral dorsohippocampal 8-OH-DPAT (5.0 μg per mouse) injections or bilateral injections into hippocampal subareas more caudal from the dorsal hippocampus displayed the 5-HT syndrome and low activity during training (data not shown). However, only those mice with precise bilateral dorsohippocampal injections showed marked deficits in the subsequent memory tests, supporting a specific involvement of dorsohippocampal 5-HT1A receptors in the contextual fear conditioning and possibly also in tone-dependent fear conditioning.
The present data clearly show that dorsal hippocampus is a part of the limbic circuitry involved in fear conditioning. Although the hippocampal formation is believed to mainly play a significant role in declarative memory, e.g., spatial learning tasks (Aggleton et al., 1986; Squire, 1992), there is increasing evidence for its involvement also in the acquisition of tasks that depend on multiple context (CS) and/or tone (CS) and shock (US) associations (Fendt and Fanselow, 1999), such as classical fear conditioning and passive avoidance. Thus, in addition to its “spatial attributes,” the hippocampus processes converging multisensory inputs representing nonspatial contextual cues and their multiple associations (Wallenstein et al., 1998; Shapiro and Eichenbaum, 1999; Sharp, 1999; Wood et al., 1999). However, even “simple” Pavlovian learning tasks such as contextual fear conditioning contain elements of spatial (place) learning, although they can also be acquired with nonspatial strategies as described in mice that were hippocampally lesioned before training (Frankland et al., 1998).
Interestingly, stimulation of postsynaptic 5-HT1Areceptors results in an impairment of fear conditioning that, in part, resembles deficits seen in animals with hippocampal lesions. Both manipulations affect more profoundly contextual than tone-dependent fear (Maren et al., 1997; this study), but the deficits by the subcutaneous 8-OH-DPAT are clearly more selective. Thus, impairments observed after post-training hippocampal lesions are usually greater than those produced before training (Maren et al., 1997, 1998;Frankland et al., 1998), whereas post-training 8-OH-DPAT does not disrupt fear conditioning even at high doses. Although post-training 8-OH-DPAT might produce deficits under low levels of fear, a severe retrograde amnesia of contextual fear results from hippocampal lesions even after intensive training (Kim and Fanselow, 1992; Anagnostaras et al., 1999). In view of the neuroanatomy of the hippocampus, these differences between selective pharmacological manipulations and lesions are not surprising because the hippocampal lesions will nonselectively disrupt most afferent cortical (entorhinal cortex via fimbria fornix) input to the hippocampus as well as its output via the subiculum to the cortex. This means that cortical input required for retrieval function will not enter the hippocampus after lesioning.
The failure of intrahippocampal WAY 100635 to block either the 5-HT syndrome or the impairment of conditioned fear caused by subcutaneous 8-OH-DPAT indicates that in addition to dorsohippocampal 5-HT1A receptors, those in extrahippocampal limbic structures such as entorhinal cortex, lateral septum, and amygdala are likely to be involved in different aspects of fear conditioning. Several studies have shown that the lateral septum (Thomas et al., 1991; Vouimba et al., 1998; Radulovic et al., 1999) and the entorhinal cortex (Maren and Fanselow, 1997) are involved in fear conditioning, and both structures are enriched with the 5-HT1A receptors (Laporte et al., 1994; Gozlan et al., 1995; Khawaja, 1995).
The inhibitory effects of 8-OH-DPAT on fear conditioning could be related to its powerful inhibitory action on hippocampal (Tada et al., 1999) and possibly, entorhinal cortex (Schmitz et al., 1998) pyramidal neuronal activity. On one hand, inhibition of entorhinal cortical cells will inhibit major extrinsic synaptic input to the hippocampal formation. On the other hand, inhibition of the CA1 glutamatergic pyramidal cells (Tada et al., 1999) will markedly reduce some of the hippocampal efferent outputs to other limbic structures, including the amygdala and the entorhinal cortex.
Footnotes
This work was supported by the Max Planck Society (J.S.) and a grant from the Swedish Medical Research Council (MFR; project K98-14X-11588-03A) (S.O.Ö.) and The Research Funds from Karolinska Institutet (S.O.Ö.). We thank Sonja Kriks and Tanja Lorenz for excellent technical assistance.
O.S. and I.M. contributed equally to this work.
Correspondence should be addressed to Dr. Oliver Stiedl, Department of Molecular Neuroendocrinology, Max Planck Institute for Experimental Medicine, Hermann-Rein-Strasse 3, D-37075 Goettingen, Germany, E-mail:stiedl{at}em.mpg.de, or Dr. Ilga Misane, Karolinska Institutet, Division of Behavioral Neuroscience, SE-171 77 Stockholm, Sweden, E-mail:ilga.misane{at}neuro.ki.se.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}